九州大学学術情報リポジトリ
Kyushu University Institutional Repository
年輪生態学的手法を用いた屋久島におけるスギ老齢 林の長期森林動態の解明
伊髙, 静
Department of Forest and Forest Products Sciences, Graduate School of Bioresource and Bioenvironmental Sciences, Kyushu University
https://doi.org/10.15017/26651
出版情報:Kyushu University, 2012, 博士(農学), 課程博士 バージョン:
権利関係:
Dendroecological Analysis of Long-term Forest Dynamics of
Old-growth Cryptomeria japonica Forest on Yakushima Island
Shizu Itaka
2013
1
Contents
Chapters
1 . Ge ne ra l Introd uc tion - - - 4
1.1. Obj ecti ves - - - 4
1.2. M ethods of Thi s St ud y - - - 5
1.3. Yakus hi m a Is l and - - - 7
1.4. Cr ypt omeri a j aponi ca Forest on Yakushi m a Is l and - - - 8
1.5. Stud y Area - - - 9
2. Es tima tion of Grow th Ra tes Ba sed on Tree -ring Anal ys is of Cry ptome ria ja ponica on Yak ushima Is land, Ja pa n - - - 13
2.1. Int roducti on - - - 13
2. 2. M at eri al and M et hods - - - 1 4 2 .2.1. S am pli ng and C ross -dati ng Trees - - - 14
2 .2.2. Age Es ti m at ion - - - 1 5 2 .2.3 . Growt h -rat e C al cul at ion - - - 18
2. 3 . R es ul ts - - - 19
2. 4 . Di s cus s i on - - - 2 4 3. Ide ntif ying De ndroec ologic al G row th Re leas es in Old-grow th Cry ptome ria japonica Fores t on Yak ushima Is land, Ja pan - - - 29
3.1. Int roduct i on - - - 29
3.2. Mat eri al and Methods - - - 30
2
3 .2.1. S am pli ng and C ross -dati ng Trees - - - 30 3 .2.2 . St anding Tree M oni toring and Mapping - - - 30 3 .2. 3. Age Es ti mati on - - - 34 3 .2.4 . Growth -rate C al cul at ion - - - 3 4 3 .2.5. R el eas e Anal ys i s - - - 3 4 3.3 . R es ul ts - - - 3 7 3 .3.1. Boundar y Li ne - - - 37 3 .3.2. Dist urbance Hist or y - - - 37 3. 4 . Di s cus si on - - - 40
4. Comparing 3 0 - ye ars Diame te r Cens use s a nd Tree -ring Chronologies on Old-grow th Cry ptome ria ja ponica from Yak us hima Is la nd , Ja pan - - - 45 4.1. Int roduct i on - - - 45 4.2. Mat eri al and Methods - - - 4 6 4 .2 .1 . D i am et e r C en s u s - - - 4 6 4 . 2. 2 . S a m pl i n g an d C r os s -d a t i n g Tr e e s - - - 4 7 4 . 2 . 3 . Anal ys i s - - - 4 7 4. 3. R esult s - - - 5 1 4 .3.1. Di am et er Cens us vs. Tree -ri ng Meas urem ent - - - 51 4 .3.2. Nail M eas urem ent vs . Tree -ring M eas urem ent - - - 51 4. 4 . Di s cus si o n - - - 57
5. Gene ral Disc us sion - - - 63
Re fe re nc es - - - 67
3
Summa r y - - - 7 4 S u m m a r y ( i n J a pa n e s e ) - - - 7 7
Ac kno wl ed gem ent s - - - 7 9
4
1. Ge ne ral introduction 1.1. Ob jecti ves
J apanes e buil t up t hei r culture i n ri ch forest res ource s over s everal t en thous ands of years (Yas uda, 1980 ). Cr yptomeri a j aponi ca (L. f.) D. Don occurs nat urall y from t he northern li mit of Aom ori P refect ure to the southern l imi t of Yakus him a Is l and i n Kagoshim a P refect ure, but becaus e o f anci ent loggi ng acti vit y, ex tens ive nat ural forest s of Cr. j aponi ca current l y onl y exi st i n Aki t a and Kochi prefectures , and on Yakushim a Isl and (M aeda, 1983). Loggi ng records do not exist for m os t of thes e forest s and l itt l e i s known about how the current forest st ruct ure devel oped . C r. japoni ca on Yakus hi ma Is l and l ive m ore than one thousand years (S uzu ki and Ts ukahara, 1987) , and the ol d-growt h Cr. japoni ca forests on Yakus hi ma Is l and had been affect ed b y l oggi ng act iviti es. Currentl y t he f orest on t he is land consis ts of 200 –300 year-ol d Cr. Japoni ca that regenerat ed aft er l oggi ng act ivi ti es and 400 t o over 1000 year-ol d t rees t hat have survived loggi ng act ivi ti es. (K yus hu R egi onal Fores t Offi ce/ Yaku -s hi ma Envi ronm ent Cons ervat ion C ent re, 1996; Yoshi da and Im anaga , 1990 ). The Cr. j aponi ca forest on Yakus him a Isl and is a val uabl e si t e enabl es estim ati on o f long-t erm growth dynam i cs and effects of hum an dis turbance. However, growt h d ynam i cs and t he effects of hum an dis turbances on t he growt h of Cr.
japoni ca on Yakus him a Is l and have not been st udi ed . Cons ervati on of a forest s uch as this requi re s an underst a nding of t he growt h d ynami cs and the effect s of hum an dist urbances on the growth of Cr. japoni ca, becaus e i t m a y help res earchers eluci dat e past forest s t ruct ure and provi de us eful informati on for t he l ong-t erm forest m anagem ent s trat egy for old -growt h C r.
japoni ca forest on Yakushi m a Isl and .
Tree-ring anal ysi s provi des uni que dat a sou rce as t he y cover a wi de range
5
of tim e. Tree -ri ng inform ati on is general l y us ed for recons t ructi on of pas t envi ronm ents (Fri tts , 1976), t ree growth (Cherubi ni et al ., 1998; Ot a et al., 2007) and dist urbance ( Lori m er and Frel i ch, 1989) , or dat ing hist ori cal mat eri al s (M it sutani , 1987). To estim ate l ong -t erm pat t erns in forest growth, cont inuous cens us i s general l y carri ed out . However, t he t im e span of monit oring dat a i s li mit e d to onl y s everal decades . There a re som e res earches about dynam i cs and di st urbance his tor y of 300 -400 year old -growt h forest (Abram s et al ., 199 9; Ot a et al., 2007; Zi egl er, 2004 ).
Us ing vari at ions i n t ree -ring width, it i s possi bl e t o extrapol ate i nforma ti on regardi ng growth d ynami cs and evaluat ing t he di st urbance his tor y.
This thesis att em pt ed t o cl ari f y t he l ong -t erm growth d ynam ics and the effect s of hum an di sturbances on the growth of C r. j aponi ca bas ed on dendroecologi cal anal ys i s . And al so gr owth f rom di am et er cens us es and tree-ri ng dat a were compare d to know error range, becaus e cens us will be a ver y i m port ant t ool t o know further growth d ynam i cs .
1.2. Method s of Thi s Stu dy
To unders t and long-term growth p att ern of C r. japoni ca on Yakushim a Is l and over the l as t several hundred yea rs , dendroecol ogi cal anal ys i s was em pl o yed (C hapt er 2 ). P revious studi es regardi ng growth of Cr. j aponi ca on Yakus hi ma Is land have m onit ored growt h si nce 1973 ( Yos hi da and Im anaga , 1990; Takas him a , 2009), whi l e others have exami ned growt h or d ynami cs of Cr. j aponi ca over a s pan of 10 -20 years ( Kim ura, 1994; Tak yu et al ., 2005);
however, 10 -30 year s mi ght not be l ong enough to underst and l ong -t erm growt h patt ern of thi s s peci es, whi ch can l ive for over 1000 years . Furthermor e, previous research using stem anal ysi s of C r. j aponi ca on Yakushi ma Is land em pl o yed onl y 3 sampl e t rees aged 45 -149 years
6
(Ham aoka, 1933), and no previous s tudies have exami ned a l arge enough sampl e of indi vi dual s coveri ng a range of ages that s pans hund reds of years . The l ong-t erm growth p at t ern of Cr. j aponi ca over t he cours e of s everal hundred years rem ai ns poorl y underst ood. T herefore, C r. j aponi ca s am pl es that regenerat ed s ubsequent t o t he i naugural year of loggi ng whi ch ranged in age from about 200 -300 years were used to devel op an unders t andi ng of growt h p att erns . Annual bas al area i ncrem ent (BA I) was calcul at ed from tree-ri ng s eri es. B ecaus e growt h rates for trees of di fferent ages and s iz es shoul d be bas ed on ring -area seri es, whi ch are l es s depen dent on st em siz e or age than ring -widt h s eries and provide an accurat e quanti fi cat ion of wood product ion (P hi pps, 1979; LeBl anc, 1990).
Hi stori cal des cripti ons rel at ed t o l oggi ng have been in exis tence si nce 1563 (Kanet ani and Yoshim aru, 2007) . However s cal e of l oggi ng act ivit y, its effect on growt h of the s urvi vi ng t rees and t he germi nati on years of Cr.
japoni ca whi ch t hrived because of gap form at ion have not been st udi ed. To reveal past l oggi ng act ivit y, i ts effects of hum an di st urbances on t he growt h of Cr. j aponi ca and t he germ ination year of regenerat ed Cr. japoni ca through gap form ations , dendroecologi cal approaches were us ed (C hapt er 3 ).
Dendroecol ogi cal approaches have been proven t o be extrem el y us eful i n evaluati ng the dist urbance hi stor y of a st and wi th compl ex age s t ruct ure over ti m e and one of t he fundam ent al dendroecologi cal approaches t o evaluati ng t he dis turbance histor y i s identi fi cati on of rel eases ( Lori m er and Frel i ch, 1989). C al cul at ion of rel eases i s a powerful and unique t ool that refl ect s disturbances at a hi gh t emporal resoluti on (Bl ack and Abram s, 2003).
7
To est im at e l ong -t erm patt erns in forest growth, general l y obt ain from cont inuous di am et er cens us or t ree -ri ngs m easurem ent s . However, rel at ivel y lit tl e res earch has focused on comp aring i nvent ori es and t ree -ring dat a (Biondi, 1999; Cl ark et al ., 2007). B iondi (1999) report ed t hat t ree -ri ng dat a was clos el y m atched wit h repeat ed forest invent ori es . We focus ed on 30 years di am et er cens us and t ree -ri ng chronologi es of C r. j aponi ca t o compare growth from di am eter census es and t ree -ring dat a t o know error range i n order t o consider about the rol e of ol d -growth forest cens us (C hapter 4 ).
1. 3. Yak ush i ma I sland
Yakus hi ma Is l and is l ocat ed at 30 20′N, 130 31 ′E, about 60 km from the southern end of K yu shu, sout hern J apan and has an area of 504 .9 km2. Thi s nearl y ci rcul ar is l and has about 130 km of s horeline. M t. Mi yanoura, locat ed at t he cent er of t he i sl and, reaches an alt it u de of 1,93 6 m and forms the isl and's hi ghest poi nt . Precipit at ion l evels on Yakus hi ma Is l and are som e of t he hi ghest i n t he worl d and range from 2,400 –5,000 mm year– 1 on the coast to 5,000– 7,400 m m year– 1 wi thi n m ount ai nous areas (Takahar a and Mats um ot o, 2 002). This heav y rai nfall i s caus ed b y as cendi ng air currents under the infl uence of t he warm P aci fi c current as well as frequent t yphoons (Takahara and M at s umoto, 2002). Wit hi n t he roughl y 2,000 m el evat ional di fference bet ween the fl atl ands and m ount ai n peak forest s range from sub-t ropi cal and t emperat e rainforest s, mix ed coni fer -broadl eaved fores t cont aining Cr. japonica, to evergreen dwarf bam boo grass land surrounding mount ain peaks (K yushu R egi onal Forest Offi ce/Yaku -shim a Envi ronm ent Conservat ion C ent re, 1996). The presence of hi gh preci pi t ati on and vert i cal dist ribut ion has l ed t o about 90% of t he i sl and developi ng ri ch forest s wit h
8
hi gh divers e flora, whi ch cont ain ol d Cr. j aponi ca, m an y endem ic and endangered s peci es (Kanetani and Yoshim aru , 2007).
1. 4. C rypt om eri a j aponi ca Forest on Yak ushi ma Isl and
At alt it udes bet ween 700 and 1, 80 0 m , t he veget ati on on the i sl and consis ts prim ari l y o f a m ixed coni fer -broadl eaved fores t dom inat ed b y old -growt h C r. j aponi ca (Mi yawaki , 1980) . Cr. japoni ca on Yakus hi m a Is l and l ive more t han one t hous and year s (Suz uki and Ts ukahara, 1987).
Thes e old -growt h Cr. japoni ca forest s on Yakus him a Is l and had been affect ed b y l oggi ng acti vi ti es ( Yoshida and Im anaga , 19 9 0). The most anci ent record rel at ed t o l oggi ng of Cr. j apon i ca from Yakus him a Is l and i s from 1563 when l oggi ng was done t o rebuil d the Kagos him a shri ne (Kanet ani and Yos him aru , 2007). S ys tem ati c l arge s cal e of loggi ng act ivi ti es of C r. japoni ca occurred over a 300 year period begi nni ng i n 1642 (Ham aoka, 1933; Kaki noki, 1954; Yos hi da and Im anaga , 1990 ). C anop y ga p form ati on b y l oggi ng act ivi ti es l ed to rege nerati ons of C r. j aponi ca (S uzuki , 1997). Currentl y t hi s forest consi st s of 200–300 year ol d, regenerat ed Cr.
japoni ca as well as 400 t o over 1 ,000 year ol d C r. j aponi ca that survived loggi ng acti vit i es (Kyus hu R egional Forest Offi ce/ Yaku -s hi ma Envi ronm ent Cons ervat ion C entre, 1996; Yoshida and Im anaga , 1990 ). The l oggi ng i s prohibit ed s ince 1982 for over 800 year old Cr. j aponi ca and s ince 2002 for the all natural Cr. japoni ca in Yakus him a Isl and (Kanet ani and Yos him aru, 2007; Kum am ot o R egional Forest Offi ce , 1982; S at o and Teraoka, 2012).
Thes e old -growt h fores ts on Yakushim a Is l and have been cons erved as Forest Ecos yst em Prot ect ed Area (FEPA), whi ch i ncl udes the worl d herit age -list ed area (Tagawa, 1994). FEPA is divi ded i nt o the core and buffer areas under t he concept of biosphere res erves , whi ch has been
9
evol ved b y UNES CO's M an and Bi osphere P rogram (Tagawa, 1994) . In the core area, n o hum an acti vit y s uch as logg ing is all owed . In t he buffer area surroundi ng t he core area, hum an activiti es are res tri ct ed, and s el ect ive loggi ng is all owed onl y in Cr. j aponi ca plant at ions (Kagoshi ma prefecture, 2012). S uch pl ant at i ons i n t he buffer z one are expect ed to be t rans form e d to naturall y regenerat ed s t ands ( K yus hu Regi onal Forest Offi ce , 2013).
Outsi de FEPA, t here i s producti on area of Cr. japoni ca, and cl earcutti ng s ys t em or s el ecti on s ys t em has been carri ed out m ainl y i n plant ati ons (K yus hu R egi onal Forest Offi ce/ Yaku -shi m a Envi ronm ent C ons ervat ion Cent re, 1996) .
1. 5. Stud y Area
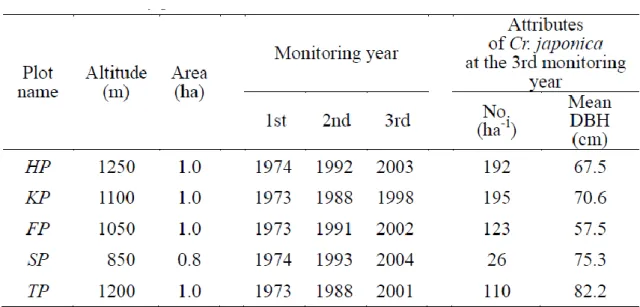
During 1973 -1974, fi ve perm anent plot s were es t ablis hed b y t he Kum am oto R egi onal Fores t Offi ce , nam ed t he Hana yam a plot (HP), Kohana yam a pl ot (KP), Fut aridake-no-kom i chi plot (FP), Shi rat ani pl ot (S P) and Tenm on-n o -m ori pl ot (TP) (Fi g. 1 -1). St ud y pl ot s were covered i n natural, uneven -age d, mix ed coni fer-broadl eaved fores t dominated b y C r.
japoni ca (Takashim a, 2009) . Stud y pl ots were locat ed bet ween 850 and 1,250 m a.s .l ., wit h SP having t he l owest el evat ion of the four pl ots , and S P had an area of 0.8 ha (100 m × 80 m ), whil e t he ot her plots had areas of 1.0 ha (100 m × 100 m) (Tabl e 1 -1). Al l st ud y pl ot s have previous l y been affect ed b y l oggi ng act ivi ti es (Yoshida and Im anaga, 1990).
Growi ng stoc k wit hi n SP was l ess t han in the other plots (Tabl e 1-1), whi ch was poss ibl y t he result of more recent hum an acti vi t y w ithin t he area and was suggest ed b y a record of t he mot her t ree m ethod having been impl em ent ed during 1897 wit hi n a nei ghbori ng secti on o f forest , aft er whi ch regenerati ons were rare (Kum am ot o R egi onal Forest Offi ce, 1982).
10
Fi g. 1 -1 Locati on of st ud y pl ot s on Yakus him a Isl and.
1
2 3
4
11
Tabl e 1-1 Stud y pl ot att ri but es
12
All pl ot s, however, contained al most t he s am e number of Cr. japoni ca stumps, and t he forest st ruct ure of thes e plots m a y have been s imi l ar prior to l arge -s cal e loggi ng act iviti es (Takashi ma, 2009) .
All l ivi ng t rees wit h di amet ers at breast hei ght (DBH, approxim at el y 1.2 m) ≥ 4 cm were measured three times at intervals of 10-19 years within each stud y plot (Tabl e 1 -1 ).
13
2. Es tima tion of Grow th Ra tes Ba sed on Tree -ring Anal ys is of Cry ptome ria ja ponica on Ya kus hima Island, Ja pa n
2.1. In trodu cti on
Previous st udi es regarding growth of Cr. japoni ca on Yakus him a Is l and have m onit ored growth s ince 1973 (Yoshi da and Im anaga, 1990; Takas him a, 2009), whil e others have exam ined growt h or d ynam i cs of Cr. japoni ca over a s pan of 10 -20 year s (Kim ura, 1994; Tak yu et al ., 2005); however, 10 -30 years mi ght not be l ong enough to underst and long -t erm growt h patt ern of thi s s peci es , whi ch can li ve for over 1000 years . Furtherm ore, previ ous res earch usi ng s t em anal ys i s of C r. j aponica on Yakushi m a Is l and emplo ye d onl y 3 s ampl e t rees aged 45 -149 years (Ham aoka, 1933), and no previ ous studi es have ex am ined a l arge enough sam pl e of indi vi dual s covering a range of ages t hat spans hundreds of year s. The long-t erm growt h patt ern of Cr. japoni ca over t he course of s everal hundred years remains poorl y underst ood.
Using vari ati ons i n t ree -ri ng width, it is pos si bl e to ext rapolat e informati on regardi ng growt h rat e. In fact, recent st udi es have us ed tree-ri ng widths to i dent i f y and quant if y growth t rends (C herubi ni et al., 1998 ; Ot a et al., 2007). The tim e span of monit oring dat a i s li mit ed t o onl y several decades ; however, tree -ri ng anal ysis usi ng s ampl e cores enabl es much more long -t erm growth patt erns to be understood. The obj ecti ve of t he pres ent st ud y was t o underst and long-t erm growt h p att ern of Cr. j aponi ca on Yakus hi ma Is l and over t he la st s everal hundred years . R egenerat ed C r.
japoni ca, whi ch ranged in age from about 200 -300 years, were exami ned in order to underst and t he long-t erm growt h patt ern wi thin t hese i ndi vi duals so that i nform ation pertinent t o t he sust ainabl e m anagem ent of Cr. japoni ca
14
forest on Yakus hi ma Is l and coul d be obt ai ned. In the pres ent s tud y, s am pl e cores were col l ect ed from Cr. j aponi ca i ndividual s locat ed wi thi n four perm anent s tud y pl ot s on Yakushim a Is l and i n order t o provi de dat a for us e in the i nvesti gat ion of l on g-t erm growt h patt erns .
2.2. Materi al and Meth ods
2.2.1. S ampli ng an d Cros s -d ating Trees
During 2005 –2008, sampl e t rees more t han 30 cm of DBH cl ass es were randoml y s el ect ed in each plot . One or t wo s ampl es from each of the DBH cl asses were cored usi ng a n increm ent borer wi th 80 cm l ength and di am et ers were m eas ured where the s am pling cores were obt ai ned. S am pl ed cores were glued ont o wooden mounts and s anded until i ndivi dual t ree -rings were cl earl y vi si bl e. Each tree -ri ng wi dt h was m easured on a TA Unisl ide Velm ex m achi ne (0.001 m il lim et er precisi on; Vel m ex Inc., Bl oom fi el d, NY, US A). Dat ing of raw t ree -ri ng widths and ass oci at ed m easurem ent errors were evaluat ed usi ng t he COFECHA program (Holm es, 1983). When there were t wo cores from the s am e t ree, t he m ean t ree ring width of the t wo cores was used t o creat e a singl e ri ng -wi dt h series for each t ree.
Four plots , HP, K P, FP and SP were us ed as s tud y s it es i n t h is st ud y (Fi g.
1-1 and Tabl e 1 -1 ). Within st ud y plots, DBH dis tribut ions obt ai ned from t he second measurem ent int erval were norm al for C r. japoni ca individual s ≤ 100 -110 cm , whi ch repres ent ed t rees that regenerat ed at around t he sam e tim e as the gaps were m ade b y l arge -s cal e loggi ng act ivi ti es . DBH dis t ribut ions were uni form for indi vi duals wit h DBH great er t han 120 cm, whi ch represent ed old -aged t rees t hat had not been t arget ed b y l oggi ng operat ions (Yos hi da and Im anaga, 1990). Aft er the t hi rd m easurem ent was conduct ed, cert ai n t rees whose DBH cl ass had been 110 cm during t he
15
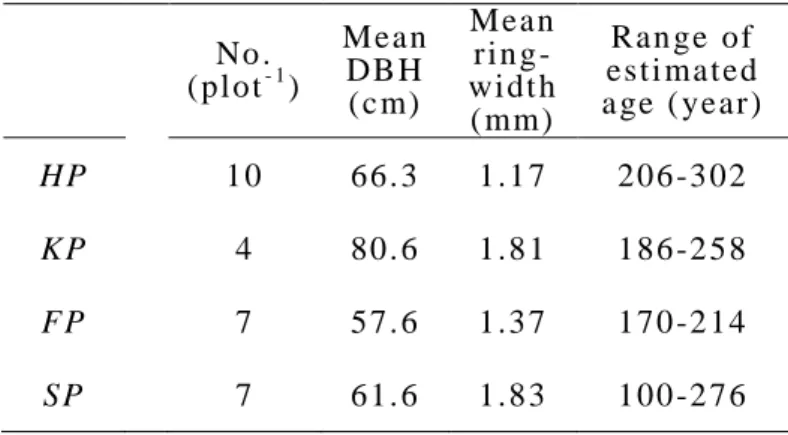
second m eas urem ent int erval had grown t o a DBH of 120 cm . In t he pres ent stud y, s ampl e cores from t rees with DBH ≤ 120 cm were us ed (Fi g. 2 -1 ).
Cores were t aken from 68 t rees , but onl y 28 t rees were anal yz ed as cert ai n cores were broken or too s hort for age estim ati o n (Tabl e 2 -1 ). The reas on for t he hi gh proport ion of broken cores is uncl ear, but m a y be due t o the frequent t yphoons i n t he area that shake bi g t rees and m a y caus e them to break insi de.
2.2.2. Age E s ti mation
Sam pl e cores oft en l acked pit h, the chronol ogi cal cent er of a t ree.
Missi ng part s of t ree -ri ng radi us were estim at ed usi ng the t wo m et hods out li ned below.
1. Measuring arc of inner t ree -ri ng
When sam pl e cores pass ed clos e enough to t he chronologi cal cent er t hat arcs of the i nner rings were vi si bl e, m is sing radi us l engt hs were es tim at ed usi ng t he equati on (Duncan, 1989):
r = L2/ 8 h + h / 2 (1)
where r: l engt h of t he mi ss ing radi us , L: length of arc, h: hei ght of an arc . Est im at ed l engths of mis si ng radi us was di vided b y the average t ree -ring wi dt h of t he i nnermost 20 ri ngs i n order t o obt ai n an est im at e of age. W hen the l ength of t he mi s sing radius appears t o be wi thi n 50 mm, t hen t he m ean absolut e error i s ±21 years of age (Duncan, 1989). In t hi s s tud y, t he m ean length of t he mis si ng radi us was 30.7 mm .
2. Age-di am et er m odel
When sam pl e cores had no vi si bl e inner ring arcs, t he mi ssi ng l engths, whi ch are cal cul at ed b y subt ract ing the l engt h of sam pl e core from t he
0
16
Fi g. 2 -1 Di am eter d ist ributi on of li vi ng C r. j aponi ca i ndivi duals (3 rd measurem ent i nt erval).
SP
K P H P
0
17
Tabl e 2-1 S ampl e t ree at t r i but es No .
(pl ot- 1)
Mean DB H (c m)
Mean ri n g- wi dt h ( mm)
Ran ge of est i mat ed a ge ( year)
HP 10 66. 3 1.1 7 206 -3 02
KP 4 80. 6 1.8 1 186 -2 58
FP 7 57. 6 1.3 7 170 -2 14
SP 7 61. 6 1.8 3 100 -2 76
18
radius at the di am et er of t he coring hei ght , and then dividing by t he average tree-ri ng widt h of t he 20 i nnermo st ri ngs to obt ai n an est im at e of age (Nort on et al ., 1987). When there were t wo cores from t he s am e t ree, t his age-di am et er m odel was us ed to cal cul at e tree age us ing a m ean ring -wi dth seri es . The m ean errors are est im at ed to be l ess t han ± 15% where t he c ore length repres ent s 80% of t he radi us (Norton et al.,1987). For thi s st ud y, t he mean core length was 80.3%.
Cores were not t aken at ground l evel ; rat her, m ost were t aken at 1.2 m above ground l evel. The ex act age of s ampl ed t rees was es ti mat ed bas ed on st em anal ys i s of C r. japoni ca on Yakus him a Is l and. The age to reach t he coring hei ght was es tim at ed usi ng the rel at ions hi p bet ween hei ght and tree-ri ng num ber of dis c from st em anal ysis (Togo, 1981).
2.2. 3. Grow th -rate Cal cul ation
Bas al area increm ent (BA I ) was used to est im at e growt h rat e, s ince growt h rat es for t rees of di fferent ages and siz es s houl d be bas ed on ri ng-area s eri es, whi ch are l ess dependent on st em siz e or age t han ri ng-wi dt h s eri es and provi de an accurat e quanti fi cati on of wood producti on (P hipps 1979; LeBl anc , 1990). BA I i s calcul at ed from raw ri ng -wi dt hs as foll ows:
BAI = π (Dt / 2 )2 – π (Dt- 1 / 2)2 ( 2) where Dt is t he di amet er of t he cori ng hei ght for year t . Di am et er of t he coring hei ght for ye ar t was cal cul ated usi ng the di am et er val ue at coring hei ght (wit hout bark) coll ect ed i n t he fi el d or from m onit or ing res ult s.
However, i f t he m eas ured di am et er was short er t han t wi ce sam pl e core length, we cal cul at ed t he diam et er as an additi onal value of length of t he
19
core and est im at ed mis si ng radi us .
BA I result s were gr ouped i nt o each age cl as s and di amet er cl as s , and Tuke y's param et ri c mul tipl e com paris on procedure s were appl i ed t o t est whether there are s i gni fi cant di fferences i n pai rs of di fferent class es.
We com pared recent growth rat e obt ai ned from 30 - year m onitori ng dat a wit h past growth rat e cal cul at ed from t ree-ring dat a t hat was dat ed from 1850 to 1900, because s am pl e siz e was l ess t han 20 i ndivi dual s t hat have tree ri ngs before 1850 . For t his com pari son, BA I dat a were grouped int o for each of di am et er cl ass es , and nonparam etric Wil coxon –M ann –Whi tne y t est was us ed t o t est whet her there are di fferences in BA I bet ween recent and past growth rat es of each di am et er cl ass. We did not anal yz e dat a from t he di am et er cl as s es m ore than 70cm becaus e sam p l e siz e was very s m al l (l ess than 2) i n thes e cl ass es for t ree -ring dat a.
2. 3. Resul ts
Di am et er of t he cori ng hei ght of s ampled t rees was cal cul at ed for each year bas ed on t ree -ring s eri es (Fi g. 2 -2 ), and age -di am et er rel ati onships were i nferred from estim at ed age and t ree -ri ng widths fo r trees of di fferent age (Fi g. 2-3). For trees s am pl ed i n t he pres ent st ud y, di amet er cl as s es ranged from 30 -120 cm, age cl asses ranged from 100 -300 years , and individual di am et er growt h curves showed hi gh diversit y (Fi g. 2-3).
St andard devi ati on of t ree -ri ng widt hs and BA I values al so s howed a wide range (Fi gs . 2 -4 and 2-5), which s ugge st ed a hi gh degree of vari abil it y bet ween i ndi vi dual growth pat t erns.
Tree-ring wi dths of 28 i ndivi duals from four s tud y pl ots were al s o grouped i nt o 10 - yea r age cl asses as wel l as 10 cm di am et er cl asses and averaged for present at ion of the result s (Fi g. 2 -4 ). Tree -ri ng growt h rat es
20
Fi g. 2 -2 Tree di am et er growth.
21
Fi g. 2 -3 Age-di am et er rel ati ons hip.
22
Fi g. 2-4 Growt h rate of t ree -ri ng width for each of age (A) and di am et er cl asses (B). The e rror bar indi cat es the st andard devi ati on .
23
Fi g. 2 -5 Growth rat e of basal area i ncrem ent for each of age (A) and di am et er cl ass es (B). The error bar i ndi cates the standard devi at ion.
24
increas ed and peaked wi thin t he 20 -50 year age cl as s and t he 10 cm di am et er cl ass , aft er whi ch growt h rat e decreas ed gradual l y (Fi g. 2-4 ).
Mean of t ree-ri ng wi dth was great er t han 1 m m i n i ndividual s below the 160 year age cl ass (Fi g. 2-4 (A)).
BA I values of 28 i ndi vi duals from four s tud y pl ots were grouped into 10 - year age cl ass es as well as 10 cm di am et er cl as s es and averaged (Fi g.
2-5). R esult s of m ult ipl e comparisons showed si gni fi cant di fferences in BA I growt h rat e wi thin i ndi vi duals in age cl ass es bel ow 50 year s and di am eter cl asses bel ow 30 cm. In fact, BA I growt h rat e i ncreas ed rapi dl y up unt il the 50 year age cl as s and t he 30 cm di am et er cl ass before it reached a ceili ng;
however, a sl ow i ncreas e i n BA I growt h rat e was obs erved from t he 50 year to the 110 year age cl as s before i t gradual l y decreas ed (Fi g. 2 -5 (A)).
Furthermore, a s low i ncreas e in BA I growt h rat e from the 30 cm di am et er cl ass to t he 50 cm diam et er cl ass was observed before it reached a ceil ing and subsequent l y s howed an i ncreas e between t he 70 and 80 cm di am et er cl asses; however, t he obs ervati on of t hi s s econdar y i ncreas e could have been t he res ul t of an insuffi ci ent num ber of s amp l e siz e more than 70 di am et er cl ass es (Fi g. 2 -5(B)).
Com paring recent 30 - year m onit oring dat a and t ree -ri ng anal ys i s from 1850 -1900 showed s i gni fi cant di fferences in m ean BA I growth rat e i n all di am et er cl as s es from 10 to 6 0 cm; t he past growt h was consi st entl y l arger that t he recent one (Fi g. 2 -6).
2. 4. Dis cu ss ion
The present stud y at t em pt ed to cl ari f y t he long-t erm growth p att erns of C r.
japoni ca on Yakushim a Is l and over l ast several hundred years usi ng tree-ri ng anal ys is .
25
Fi g. 2-6 Growt h rate of basal area increm ent of t ree-ring anal ys i s from 1850 -1900 and m oni tori ng result duri ng t he last 30 years . The error bar indi cat es t he st andard deviati on.
26
Accordi ng to the resul t s of t ree -ri ng anal ys i s, t ree -ring wi dth i ncreas ed unt il the 20 -50 year age cl ass and t he 10 cm di amet er cl ass , aft er whi ch poi nt growth rat e gradual l y decreas ed (Fi g. 2 -4). The decline obs erved wit hi n thi s age cl as s coul d be t ypi cal of tree -ri ng width gro wth pat t erns , whi ch are wide ri ngs near t he pit h and narrower rings toward out si de (P hipps, 1979). Cr. j aponi ca on Yakus hi ma Is l and are known for t hei r sl ow growt h, and it has been report ed that t ree -ri ng wi dt h was l es s t han 1 m m (Numat a, 1986); however, m ean of t ree-ri ng widths of sampl e trees l ess t han 160 years in age obs erved in t he pres ent stud y were greater t han 1 m m (Fi g.
2-4(A)).
General l y, growt h rate (m3 year- 1) culmi nati on of art i fi ci al Cr. j aponi ca forests occurs aft er approxim at el y 1 5 -40 years (Ot om o, 1983); however, thi s growt h pat t ern has not been wel l st udi ed in ot her natural C r. j aponi ca forests i n J apan. The pres ent stud y de monst rat ed that BAI growt h rat e increas ed rapi dl y i n trees under t he 50 year age cl as s and di spl a yed a subs equent gra dual ris e unti l it peaked within t he 110 year age cl as s , aft er whi ch poi nt i t gradual l y decreas ed (Fi g. 2 -5 (A)). Therefore, t he obs erved growt h rat e peak i n age of t he nat ural Cr. japoni ca forest on Yakushim a Is l and was si gnifi cantl y great er t han t hat of a rt i fi ci al C r. j aponi ca fores ts . However, i ndivi dual vari ati on in t ree growth patt erns i s a comm on characterist i c of nat ural forest s, becaus e the growi ng condi tions of each individual t ree can di ffer (K i mura , 1994). Wide st andard devi ati ons of tree-ri ng wid ths and BA I val ues mi ght also be a charact erist ic of nat ural forests (Fi g s . 2 -4 and 2 -5 ).
BA I growth rate showed an i nit i al i ncrease under t he 30 cm di am et er cl ass , a slow i ncreas e wit hi n the 30 t o 50 cm di amet er cl ass, and peaked wit hi n t he 70-80 cm di am et er cl ass before i ncreasing again; however, results from
27
more t han 70 cm diam et er cl ass m i ght have s kewed res ul ts due t o an inadequat e s am pl e s ize (Fi g. 2 -5(B)). Typi call y, in order t o m ake a m ean tree-ri ng chronol ogy, 20 -30 t rees are required ( C ook and Kai ri uksti s , 1990);
however, in t he pres ent st ud y, the s ampl e siz e for t he 70 cm di am et er cl ass was below 15 whil e t he s ampl e siz e for t he 80 cm di am et er cl ass was below 10. At l eas t growth rat e increased t o the 50 cm di am et er cl as s , aft er which poi nt BA I growt h rat e was sti ll hi gh (approxi m at el y 25 cm2 year- 1).
Monit ori ng res ult s obt ai ned over t he l ast 30 years have s hown t hat t he mean BA I growt h rate of C r. j aponi ca t rees i n all di am et er class es from 1 0 to 60cm consi st entl y had si gni fi cant di fferences com p ari ng t o m ean BA I growt h rat e of tree -ring anal ysi s from 1850 -1900 (Fi g. 2 -6 ). Thes e result s indi cat ed t hat growt h over t he l ast 30 years was much sl ower than growt h that occurred s everal hundred years ago, sugges ti ng t hat growth condi ti ons of Cr. j aponi ca were bett er i n t he past . One poss ibl e expl anat ion coul d be that l arge -s cal e l ogging acti vi ti es t hat occurred about 350 years ago and cont inued about 300 years encouraged gr owth b y providi ng bet t er l i ght and sp at i al conditi ons . There m a y be other fact ors c ausi ng growt h di fferences over a few hundred years; di fferences i n mi croclim at e condit ions and tree-age m i ght be s uch fact ors.
In concl us ion, t he res ul ts of the present st ud y emphas iz ed t hat BA I growt h of Cr. japoni ca on Yakushim a Isl and showed an initi al i ncreas e and peaked i n t he 110 ye ar age cl as s and the 50 cm di am et er cl as s, whil e l arge di am et er t rees m ai nt ai ned hi gh growt h rat es. M ean of t ree-ri ng width was great er t han 1 mm wit hi n indi vi dual s bel ow t he 160 year age cl as s and growt h rat e was hi gher 10 0-150 years ago t han i t was wit hi n the l ast 30 years. These result s cl ari fi ed BA I gr owth p att erns of regenerat ed Cr.
japoni ca aft er l arge -scal e l oggi ng activit ies; pas t growth was much bet t er
28
than recent one and i t m i ght have been affect ed mainl y b y l oggi ng act ivi ti es . This sugges t s t hat hum an or nat ural dist urbances m a y be very im port ant t o encourage growt h of Cr. j aponi ca over l ong-t erm forest m anagem ent st rat egy for old -growt h Cr. japoni ca fores t on Yakushi m a Isl and or other regions . Furt her res earch s h oul d focus on usi ng t ree -ri ng dat a of s tumps and fall en l ogs i n order t o underst and growt h patt erns prior to ext ensi ve l oggi ng.
29
3. Ide ntif ying De ndroecological G row th Re lease s in Old-grow th Cry ptome ria ja ponica Fores t on Ya kus hima Is la nd , J apa n
3.1. In trodu cti on
Cons ervat ion of old -growt h Cr. j aponi ca forest s requi re s an underst anding of t he effects of hum an dis turbances on the growt h of Cr.
japoni ca and d yna mi cs of Cr. j aponi ca fores t, becaus e it m a y help res earchers elucidat e past forest st ructure and p rovi de us eful inform at ion for t he l ong-t erm fores t m anagem ent s t rat egy i n t he buffer and producti on areas . His tori cal descriptions rel at ed to loggi ng have been in exis t ence since 1563 (Kanet ani and Yoshi m aru, 2007) . However s cale of logging act ivi t y, it s effect on growt h of t he s urviving trees and t he germi nat ion years of C r. japoni ca whi ch t hri ved because of gap form ati on have not been studi ed.
Dendroecol ogi cal approaches have been proven t o be ext rem el y us eful in evaluati ng the dist urbance hi stor y of a st a nd wi th compl ex age s t ruct ure over ti m e and one of t he fundam ent al dendroecologi cal approaches t o evaluati ng t he dis turbance histor y i s identi fi cati on of rel eases ( Lori m er and Frel i ch, 1989). C al cul at ion of rel eases i s a powerful and unique t ool that refl ect s dis turbances at a hi gh t emporal resoluti on (Bl ack and Abram s, 2003). R egi onal st udi es of di st urbance regi mes have been us eful in underst anding speci es d ynami cs and have s erved as gui des for restoring natural veget ati on compl ex es (Bonni cks en and St one, 198 0 ). The objecti ve s of the pres ent s tudy w ere; 1) t o pi npoint t he ti m e and the s cal e of disturbances to veri fy anci ent records of l oggi ng act ivi t y, 2) t o det erm ine the effects of l oggi ng on growt h of ol d -aged Cr. japoni ca and 3) to reveal the germi nat ion yea r of regenerat ed Cr. japoni ca through gap form ati ons
30
usi ng dendroecol ogi cal approaches and posit ional i nformat ion. For t his stud y we focus ed on det ecting rel eases i n t he FP st ud y pl ot . The FP area has been desi gnat ed as recreati on forest s ince 1971.
3. 2. Materi al and Meth ods
3.2.1. S ampli ng an d Cros s -d ating Trees
Four plots , HP, K P, FP and S P were used as st ud y s it e i n t hi s st ud y (Fi g.
1-1 and Tabl e 1 -1 ). The methods of taki ng sampl e cores, m eas uri ng tree-ri ng wi dt h and evaluating error of m easurem ent s were t he s am e as
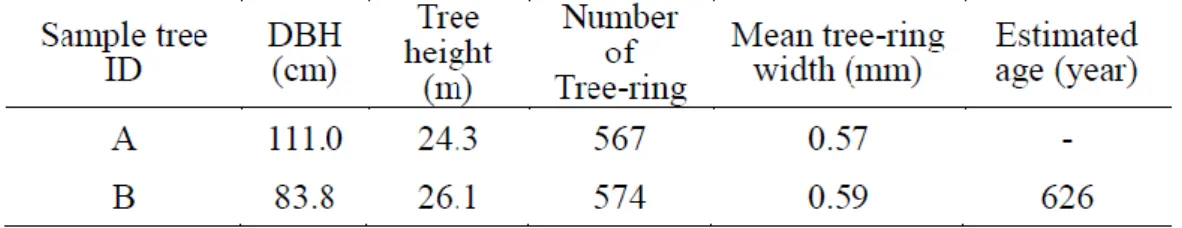
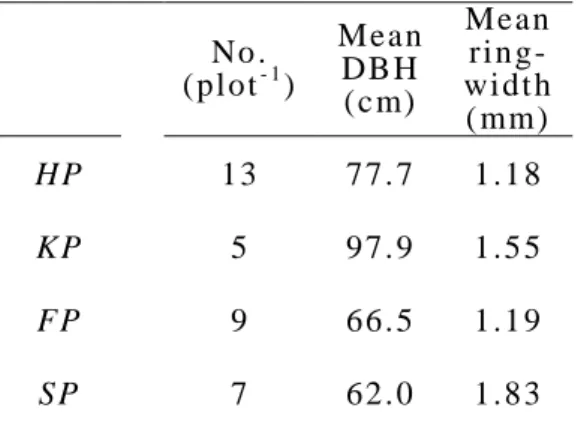
“2.2.1. Sampling and Cross-dating Trees”. To detect releases for the last 550 years we us ed tree -ri ng dat a from FP st ud y pl ot and succeeded in obt aining t wo long sampl e cores, whi l e t aki ng cores from l arge di am et er trees was s o di ffi cult t hat m ostl y i nnermos t of cores were broken. In FP stud y plot , two old -aged trees t hat mi ght be regenerat ed before t he st arti ng year of l arge s cal e l oggi ng acti vit y i n 1642 and six regenerat ed t rees t hat were expect ed t o have regenerat ed aft er 1642, were us ed t o det ect rel eas es (Tabl e s 3 -1 and 3 -2 ). A l arge data s et of t ree ring m eas urem ents was needed to cal cul at e s peci es -speci fi c rel eas e crit eri a. Therefore, we suppl em ent ed our dat a wit h 34 t ree -ri ng dat a sets from t he four st ud y s i t es (Tabl e 3-3).
3.2.2. Standi ng Tree Mon itorin g and Mappin g
Di am et er and s peci es nam e of al l li ving t rees wit h DBH ≥ 4 cm have been recorded three di fferent t im es s ince 1973 or 1974 wi thi n each s tud y pl ot (Tabl e 1 -1 ). El evati ons were m eas ured on a 20 m gri d at corners of the sub-bl ocks and posi t ion s of al l softwood and domi nant broad-l eafed t rees were mapped (Takashim a, 2009 ). For C r. japoni ca t rees in t he FP s tud y pl ot
31
Tabl e 3 -1 S am pl e t ree att ribut es: old -aged s ampl e t rees of the FP st ud y plot
32
Tabl e 3 -2 S ampl e t ree att ri but es: regenerat ed s am pl e t rees of t he FP s tud y plot
33
Tabl e 3-3 S ampl e t ree at t r i but es No .
(pl ot- 1)
Mean DB H (c m)
Mean ri n g- wi dt h ( mm)
HP 13 77. 7 1.1 8
KP 5 97. 9 1.5 5
FP 9 66. 5 1.1 9
SP 7 62. 0 1.8 3
34
regenerati on t ypes were al so recorded ; trees regenerat ed from t he ground, logs or s tump s. Wi t hin the FP s tud y pl ot Cr. japonica s nags and st um ps (DBH ≥ 10 cm) were mapped and their DBH were recorded in 2005 (Takas hi m a, 2009 ). Fi g. 3 -1 shows positi ons of l iving t rees, s nags and stumps in t he FP s tud y plot .
3.2.3. Age E s ti mation
The m et hod of age es tim ati on was s am e as “2.2.2. Age Es tim at ion , 1.
Measuring arc of inner tree -ri ng”. In t his st ud y, we es tim at ed age of 7 sampl e cores from FP st ud y pl ot . The m ean l ength of t he mi ss ing radii was 28.27 mm . One ol d -aged s ampl e t ree had a core whi ch was too short , thus we di d not calcul at e the age to avoi d a l arge m argi n of error.
3.2.4. Grow th -rate Cal cul ation
BA I was cal cul at ed using the s am e form at ion as “2.2.3. Growt h -rat e Calculation”.
3.2.5. Rel eas e Analysis
Releas e anal ys is us i ng t ree -ri ng wi dt h is a us eful approach for evaluat ing the dis turbance his tor y of a s tand wit h compl ex age st ructure (Lori m er and Frel i ch, 1989). For t he anal ysi s, the percent age growth change (%GC ), whi ch was t he percent age di f ference bet ween preceding and subs equent 10 - yr m eans of t ree -ri ng width, was calcul at ed using the formul a bel ow (Nowacki and Abram s, 1997):
%GC = (M2 - M1) / M1 × 100 (3)
where M1: precedi ng 10 - yr m ean, M2: subsequent 10 - yr m ean. A 10 - yr s pan
35
Fi g. 3 -1 Locati on of li vi ng t rees, s tumps and s nags in t he FP stud y pl ot . A and B: o ld -aged t rees . a-f: regenerat ed t rees .
36
for radi al -growth averaging was used t o det ect sust ained growt h i ncreas es in percent age to di s count the influence of cl im at e and ot her short -t erm growt h pert urbat ions (Leak, 1987). %GC of ei ght Cr. j aponi ca s ampl e trees in FP s tud y plot were cal cul at ed t o det ect growth i ncreas es caus ed b y gap form ati ons from hum an or natural disturbance.
To obt ain rel eas e cri teri a, the boundar y l ine m et hod was us ed (Bl ack and Abram s , 2003) becaus e t hi s m et hod s ol ved the dendroecol ogical probl ems of ring wi dt h decreasing caused b y agi ng and narrow ring wi dt h s howi ng ext rem el y l arge %GC. Thi s m et hod uses two st eps : (1) em pi ri cal esti mati on of t he m axi mum g rowt h change bas ed on prior growth, and (2) s cali ng of t he rel eas es rel ati ve to t he boundar y li ne (Splecht na et al., 2005).
In t he fi rst st ep t he boundar y l i ne m et hod i s det erm ined based on the rel at ions hi p between %GC and prior growth values, whi ch was m e an growth over t he pri or 10 yea rs . For cal cul at ing t he speci es -specifi c boundar y l ine, a large dat a s et of t ree ri ng m easurem ents was needed. Therefore, we suppl em ent ed t ree -ri ng dat a from the FP st ud y s it e wi th dat a from another three perm anent st ud y plot s on Yakus him a Is l and; tot al number of individual s used was 34 (Tabl e 3-3). We divided t he dat a s et i nto ni ne pri or growt h cl ass es (cl ass wi dt h 0.5 m m), averaged the t en hi ghest growt h change values for ever y growth cl as s, and fit l inear, power, l ogari thm i c and exponenti al curves and s el ect ed t he functi on t hat yi elded the hi ghest R2 value. In t he s econd st ep, all t he rel eas es were eval uat ed rel ative t o t he boundar y li ne. We i dent i fi ed pot ent i al rel eas es according to a procedure devel oped b y Bl ack and Abrams (2003) as fol lows. Onl y %GC val ues great er than 10% were ret ained. A t i me seri es graph of %GC shows increases at poi nts of pot enti al rel ease, and onl y t he m axi mum %GC for each ascent was used so that each peak woul d be considered onl y once as a
37
pot ent i al rel eas e. Onl y t hes e pot ent i al rel eas es were then eval uat ed rel ati ve to the boundar y l ine. We i denti fi ed an y %GC peak more t han 20% of t he boundar y li ne at the gi ven pri or growth rat e as m oderate rel eas e and an y peak ex ceeding 50% of t he val ue of the bounda r y l ine as a m ajor rel eas e.
3. 3. Resul ts
3.3.1. Bou ndary Lin e
The best fitt ed equat ion as t he boundar y l ine was:
%GC = –91.88 l n (P G) + 137.56 (4)
where P G: prior growt h. The R2 val ue of above equati on was 0.9 6 (Fi g.
3-2(A)). All cal cul at ed %GC rang ed from – 77.7 to 277.9% for pri or gro wth from 0.12 to 4.71 mm (Fi g. 3 -2(B) ). Fi g. 3 -2 (B) als o i ncl udes the li nes indi cati ng 50% and 20% of the boundar y l ine, whi ch are t hresholds us ed to define m aj or and m oderat e rel eases , respect ivel y.
3.3.2. Dis turban ce His tory
Fi g. 3 -3 s hows t he dis t ribut ion of rel ease for s am pl e t rees (A) and (B).
Sam pl e t ree (A) s howed m aj or rel eas es i n 1751, 1774, 1778 and 1996, and sampl e t ree (B) s howed them i n 1629, 1687, 1689, 1691, 1821, 1845, 1892, 1905 and 1939. The BA I val ue of sam pl e tree (A) increas ed from the mi ddl e of 1700s t o t he begi nni ng of 1900s. The BA I of s ampl e t ree (B) i ncreas ed from t he begi nning of 1800s to the end of 1900s . Thes e i ncreas es of BAI value occurred aft er t he frequent m ajor rel eas es from the m i ddl e o f 1700s for sampl e t ree (A) and from t he beginni ng of 1800s for sampl e t ree (B).
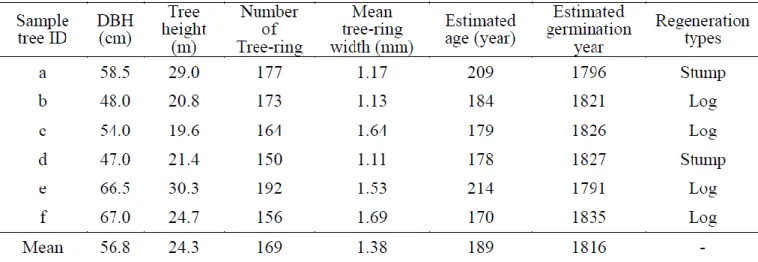
Tabl e 3-2 shows t he es tim at ed age from regenerat ed l iving t rees and
38
Fi g. 3 -2 Li near approxim at ion of averaged value of t en hi ghest growt h change for ever y growt h cl ass (A). Boundar y l ine and pl ot of percent growt h change (%GC ) val ues wit h respect to pri or growt h for 34 sam pl e t rees (B).
39
Fi g. 3 -3 P ercent growth change (%GC ) and bas al area i ncrement (BA I) for sampl e t rees (A) and (B).
40
regenerati on t ypes . Even t hough t he y were l ocat ed in t wo different areas (Fi g. 3 -1 ), regeneration years were wit hin t he rel ati vel y n arrow range bet ween t he years 1791 and 183 5 . This t imi ng was consi st ent wit h a m aj or rel eas e foll owed b y hi gh BAI values for bot h s am pl e t rees (A) and (B).
Fi g. 3 -4 s hows the number of s ampl e t rees showi ng moderat e and m ajor rel eas es wi thin each of 10 - year cl as s for ol d -aged and rege nerat ed t rees . Ol d -aged t rees showed m aj or and m oderat e rel eas es from the 1450’s to 1990’s. Regenerated trees showed major and mode rate releases from the 1820’s to 1990’s.
3. 4. Dis cu ss ion
The present s tud y a tt em pt ed t o pi npoi nt the t im e of di st urbance of C r.
japoni ca on Yakus him a Is l and over l as t s everal hundred years usi ng tree-ri ng anal ys i s . Ol d -aged sampl e t rees (A) and (B) s howed i ncreasi ng growt h alt hough they were approxim at el y 5 00 to 600 years old, whil e t he growt h rat e of t rees norm all y decli nes as a tree ages (Gower et al., 1996).
The s am pl e t ree (A) showed m ajor rel eas e from the mi ddl e of 1700s and the sampl e t ree (B) s howed m aj or rel ease fro m the beginni ng of 1800s (Fi g.
3-3). Both t rees s howed a rel ati vel y hi gh BAI val ue for about 150 years aft er t hes e r el eas es (Fi g. 3 -3). In old growt h natural Cr. j aponi ca fores t i n Akit a, the growt h of 160 –200 years old Cr. japoni ca i ncre as ed aft er thi nning (Nis hiz ono et al ., 2006). This stud y cl arifi ed that m uch ol der Cr.
japoni ca trees o n Yakushi m a Isl and al so increas ed their growth rat e s aft er dis turbances.
Est im at ed germi nat i on years of regenerated t rees were bet ween 1791 and 1835, whi ch were aft er the m ajor rel ease of ol d -aged s ampl e t rees foll owed b y l ong-l as ti ng hi gh BA I val ue s , and al l of t hem regenerat ed on st um ps or
41
Fi g. 3-4 Decadal dist ribut ion of m aj or and m oderat e releases of t wo old -aged and six regenerat ed t rees for FP st ud y plot . Tot al number of rel ease (A), and rel ease for each sam pl e t ree s (B).
42
logs (Tabl e 3 -2 ). Hence, t hese regenerat ed t rees m i ght have grown up in improved l i ght and bett er condi tion s of com peti tion on nei ghbori ng t rees becaus e of loggi ng act ivit y. Eve n though the regenerat ed s am pl e t rees were locat ed in t wo s eparate pl aces, germi nat ion year of s am pl e t rees cent ered on a short peri od of t im e (Tabl e 3-2 and Fi g. 3-1). This res ult s how s t here was loggi ng acti vi t y in t he s am e t im e point i n bot h thes e areas o f the st ud y si t e.
A m aj or rel eas e of t he old -aged sampl e t ree (A) was det ect ed duri ng t he 1990 ’s , but s ampl e t ree (B) did not show rel eas e for t he 1990 ’s (Fi gs. 3-3 and 3 -4 ). M ajor rel eases of regenerated t rees for t he 1970 ’s and 1980 ’s were al so det ect ed (Fi g. 3-4). These releases mi ght have been caused b y nat ural dis turbance, because the FP st ud y pl ot has been s t ri ct l y prot ect ed since 1971. The m ajor natural dist urbance in Yakus him a Is land mi ght be l and sli de and t yphoon. S him okawa and Jitousono (1984) report ed that l and s li de ma y happen ever y 1000 years i n st eep or d rai nage basin in Yakushim a Is l and. In FP st ud y pl ot on gentl e sl ope, however, land sli de m i ght not happen at l east l as t 600 years, j udgi ng from the number of t ree -ri ng for t he sampl e trees (Tabl e 3-1 ) and the exist ence of m an y l arge t rees and s tumps (Fi g. 3 -1 ). Yakushi ma Is l and i s s uscept ibl e t o t yphoons , which m a y caus e the canop y gaps i n t he st ud y plot . The weather s t ati on of Yakushim a recorded wi nd veloci ti es exceedi ng 55 m s- 1 ei ght tim e s from 1938 t o 2012 (J apan M et eorologi cal Agenc y, 2013), m eaning powerful t yphoons hi t about ever y 10 years in Yakushi m a Is l and. However, Takas him a (2009) reported that onl y a few Cr. j aponi ca have been recruit ed in perm anent s tud y pl ot s incl uding FP st ud y plot bas ed on m onit oring res ult s si nce 1973. In thes e plots, som e l os s es of api cal parts of the crowns were observed ( Ishii et al ., 2010), whil e whol e crown dam aged or uproot ed t rees are rarel y observed, especi al l y i n l arger t rees . Onl y one bi g C r. japoni ca wi th a DBH of 250 cm
43
in K P st ud y plot was fel l ed b y t he t yphoon (No. 19) in 1997 , but no recrui tm ent of Cr. j aponi ca was observed. This s uggest s di st urbances s ince 1970 ’s mi ght have been sm al l er s cal e t han previ ous loggi ng acti vit y and happened at t he i ndi vidual tree l evel . In addi ti on, s uch sm al l s cal e nat ural dis turbance m a y als o occur all the ti me even before 1970 ’s and during the large s cal e logging a ct ivit y.
There are hi st ori cal des cri pti on s s howing t he earli es t l oggi ng occurred i n 1563 and t he s t arti ng year of s ys t em ati c loggi ng act ivit y was 1642. In the FP st ud y s it e som e moderate rel eases were det ect ed si nce 1450 ’s , but no major rel eas es occurred unti l 1629. Ba sed on t he t ree -ri ng anal ys i s of stumps, there m a y have been som e loggi ng acti vi ti es bef ore 1642 (Ushij im a et al ., 2006), and s o t hes e moderat e rel eas es before 1642 m a y have been caus ed b y logging as well as natural dis turbances . However, thes e loggi ng act ivi ti es mi ght have been s m all er scal e than l at er s ys t emati c l oggi ng act ivi ti es, becaus e onl y m oderate rel eases occurred .
In concl us ion, t hi s s tud y emphasiz ed t hat s ys t em at i c large s cal e l oggi ng act ivi ti es of Cr. japoni ca occurred as part of t he hist ori cal record. In our stud y s it e, logging act ivi t y st art ed about 1630 and l arge scal e loggi ng act ivi t y occurred from the mi ddle of 1700s . Large s cal e loggi ng acti vi t y encouraged growth rat e s i n older t rees about 500 t o 600 years ol d ; gap form ati on m a y be i m port ant for regenerat ion of Cr. japoni ca and sm all s cal e dis turbance as soci at ed wi th indi vidua l t ree l evel mi ght be inadequat e to stim ul at e regenerati on of Cr. j aponi ca. Low l evels of dis t urbances als o occurred before 1630 and these rel eases were li kel y t o be caus ed b y l oggi ng but mi ght have been sm al l scal e. These res ul ts suggest past l oggi ng act ivi ti es are im port ant t o encourage growt h and regen erati on of C r.
japoni ca.
44
Current l y, loggi ng of C r. japoni ca i s basi call y not all owed in t he core area i n FEPA. However, s el ect ive loggings are carried out for Cr. j aponi ca pl ant ati ons i n t he buffer area i n FEPA and i n t he producti on area outsi de FEPA. Our findi ngs sugges t that group selection s ys t em is m ore appropri at e rat her than s ingl e -t ree s el ecti on in order t o encourage natural regenerat ion and growth of rem aining t rees i n such areas. Int eres ti ngl y, Im a da (1986) had al read y propos ed the group s el ect ion sys t em wit h 240 yea r rot ati on for product ion area of C r. j aponi ca forest on Yakus hi ma Is l and, and t hi s s ys t e m has been experim ent al l y i mpl em ent ed. Thus, i t coul d be very val uabl e t o evaluat e such an ex peri m ent al pract i ce to furt her confi rm t he effect s of loggi ngs.
45
4. Comparing 3 0 - ye ars Diame te r Cens use s a nd Tree -ring Chronologies on O ld-grow th Cry ptome ria ja ponica from Yak us hima Is la nd , Ja pan
4.1. In trodu cti on
Cons ervat ion of t he old -growt h forest requi r es an underst andi ng of long-t erm growth i n C r. japoni ca, becaus e i denti fi cati on of l ong -t erm patt erns in fores t growth is needed t o underst and fores t dynam i cs , wi th di rect i mpli cati ons i n fores t m anagem ent and sil vi cul ture (Biondi , 1999).
However, growth o f nat ural forest is not well known, becaus e forest st ructure of nat ural forest i s compl ex and its growth process i s various . Conti nuous di am eter cens us or t ree -ri ngs m eas urem ents are required to estim at e long -t erm pat t erns i n forest growt h .
The most comm on m ethod of growt h es tim ati on is m easuri ng t ree di am et ers repeat edl y and cal cul ati ng di amet er change (C l ark et al., 2007).
This approach has t he advant ages t hat diam et ers can be m easured rapidl y (Cl ark et al., 2007), it is possibl e to coll ect dat a i n prot ec t ion area wi thout dam aging t rees, and it i s eas y t o get inform at ion about surroundi ng actual sit uati on , but not possi bl e to get past i nform ati on before st art ing census.
However t here are several di s advant ages t o di am et er m eas urem ents . The di am et er m easurem ents have substant i al error (Barker et al ., 2002; Kit ahara et al., 2009), and negati ve growt h coul d be obs erved (Cl ark et al ., 2007) al though posi tive gr owth should occur each year. Annual gr owth rat es are oft en unknown, becaus e i nt ervals bet ween m eas urem e nts can be long (C l ark et al., 2007). Especiall y i t is di ffi cult t o m easure bi g C r. j aponi ca t rees because of i rregul ar s hape, covering t hi ck moos of t ree surface, and poor foot ing caus ed b y man y bi g stumps and logs on forest fl oor. Thus, it i s
46
concerned t hat as t he di am et ers are bi gger, m eas urem ent s m a y be l es s accurate.
In comparis on, advant age of t ree -ri ng m eas urem ent is t hat t ree -ring dat a provide an accurat e repres ent ati on of yea r -t o- year growth pat t erns (Bi ondi , 1999), and negati ve growth can ’t be observed. Tree-ring inform ati on al so represent s past growth back to t he year of germ inati on of t he t ree. However, there are s om e di s advant ages . Tree -ri ng dat a mi ght i ncl ude fal s e or mis si ng ri ngs. As one or two s ample cores are t aken general l y from one t ree, t he act ual t ree circum ference i s not known (Cl ark et al ., 2007). S am pl e cores shri nk aft er t aki ng from t rees and are not be abl e t o be t aken from prot ect ed areas or pl aces where t rees do not produce ident ifi abl e annual rings .
Relati vel y l itt l e res earch has focused on com pari ng inventori es and dendrochronol ogi cal records (Biondi, 1999; C l ark et al ., 2007). Biondi (1999) reported that tree -ri ng data was cl osel y m at ched with repeat ed fores t invent ori es . We focus ed on 30 years di am et er cens us and t ree -ri ng chronol ogi es on ol d -growt h Cr. j aponi ca fores t from Yakushi m a Isl and, whi ch cons is ts of few hundred year old trees. The obj ective of t hi s st ud y was t o compare growt h from di am et er census es and t ree -ri ng dat a t o know error range i n order to consi der about the r ol e of cens us for nat ural old -growt h Cr. japonica forest on Yakus him a Is l and .
4.2. Materi al and M eth ods 4.2.1. Di ameter Cen sus
DBH of al l li ving t rees wit h DBH ≥ 4 cm were recorded three di fferent tim es since 1973 in the fi ve plots (Tabl e 1 -1). DBH m easu rem ent was conduct ed usi ng cal iper in 0.5 cm round bas ed up to 90 cm or 100 cm cal iper s iz e. Two m eas urem ents from di fferent di rections at ri ght angl es t o
47
each ot her were ob t ai ned for each t ree, and the m easurem ent val ues were averaged. Di am eter tape was u s ed for each t ree that was not abl e t o be measured usi ng caliper, in 0.5 cm round based. P ol e was us ed for each t ree that was st andi ng on poor footi ng caus ed b y m an y bi g s tum ps and logs on forest fl oor, in 5 cm round based. At t he breast hei ght we placed n um ber pl at es and m easured DBH ever y census at s am e pl ace. Furthermore, the rest lengths of t he nai ls , whi ch att ached t he num ber pl at es t o the s tems (Fi g.
4-1 ), were randoml y sel ect ed and m eas ured i n 0.1 cm round based during at thi rd m eas urem ent for 224 t r ees i n st ud y plots of HP, FP and S P (Fi g. 1 -1 and Tabl e 1 -1 ).
4.2. 2. S ampli ng an d Cros s -d ating Trees
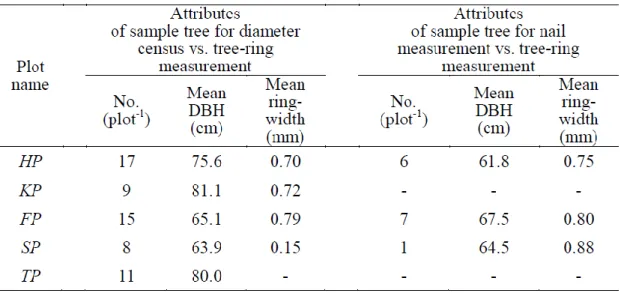
Tree-ring width of s ampl e c ores from 49 i ndi vi duals from st udy pl ot s HP, KP, FP and SP, were m eas ured (Fi g. 1 -1 and Tabl e 1 -1 ). The m ethods of taki ng s am pl e cores , m easuri ng t ree -ri ng wi dth and eval uating error of measurem ent s were t he s am e as “2 .2.1. S am pl ing and C ros s -dat ing Trees ”.
In T P s tud y pl ot (Fi g. 1 -1 and Tabl e 1 -1 ), 12 sampl e cores from 11 individual s were t aken in 2012. To know percent age of sh ri nk, cores were ins erted i nto st raw and t he siz e of core (wit hout bark) was m arked on the st raw i mm edi at el y a ft er t he t aki ng cores and t he l engt h were m eas ured . Aft er ai r dri ed, l ength of cores w ere m easured and percent age of s hri nk is cal cul at ed. The res ult s from t ree -ring m eas urem ent were i ncreas ed b y average shrink percent age 1.5%, whi ch were added t o the res ult of t ree -ring measurem ent .
4.2. 3. An al ys is
The period bet ween t he fi rst and second measurem ent s of di amet er census
48
Fi g. 4 -1 Dri vi ng a nail i n to t he st em (Yoshida, 2007 ).
49
is t erm ed as ‘fi rst peri od’, between t he second and thi rd m eas urem ents as
‘second period’, and between the first and third measurements as ‘all period’. The first period and second period were termed 10 -19 years and all period was t erm ed 28 -30 years . For fi rs t , second and al l peri od s, di am et er growt h per 10 years (wit hout bark) was cal cul at ed from t he cens us records . The percent age of bark t hi ckness was esti mat ed us ing t he rel at ions hi p bet ween di am et er and bark thi ckness of di s c from s t em anal ys i s (Togo, 1981). We cal cul at ed the di am et er growth of 4 9 indi vi duals , whi ch were identi cal t o t rees whos e s am pl e cores were t aken from 4 s tud y plots (HP, KP, FP and SP) (Tabl e 4 -1 ). For t he s am e peri od, di am et er growt h per 10 years was cal cul at ed from t he sum of t ree -ri ng wi dt hs . Diam et er growt h per 10 years from 14 i ndivi dual s, for which res t lengths of the nail s were m eas ured and als o s am pl e cores were t aken, was calcul at ed.
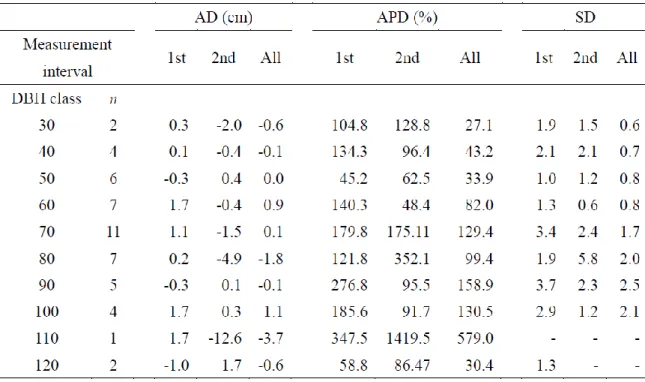
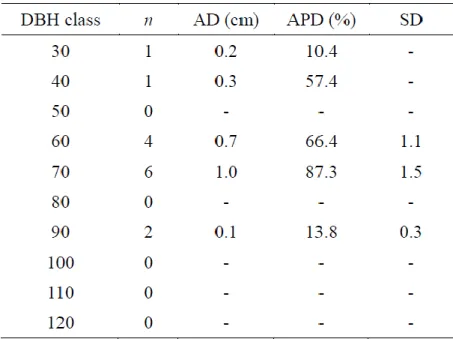
Pai red t t es t was appl i ed to t es t t he null h ypot hesi s of no di fference bet ween di am et er cens us/ nai l m easurem ent and t ree -ring m easurem ent . For the cont inuous vari abl es , we cal cul at ed average di fference (AD), average percent age di fference (APD), s t andard devi ati on (SD) of the di fferences , whi ch are commonl y us ed m ea sures of random m easurem ent error (Kit ahara et al ., 2009). For t he cal cul at ion t ree -ri ng measurem ent s were defined as true value. AD defi ned as the average of t he di am et er cens us /nai l measurem ent mi nus t he t ree -ri ng meas urem ent, and AP D defined as the average of t he abs ol ut e di fference divided b y the t ree -ring measurem ent . Di am et er growth per 10 years from diam et er cens us , t ree -ring m eas urem ent and rest l engths of t h e nai ls were grouped into each di am et er class , and AD, AP D and S D for each di am et er cl ass we re also cal cul at ed.
From t he all dat a of res t l engt h of nai l, percent age t hat nails have fall en awa y from t he st em and nai ls have been pushed out of the st em, were
50
Tabl e 4-1 S ampl e t ree att ri but es at the 3rd moni tori ng year