新規細胞死阻害剤
IM-93 は急性尿細管壊死と好中球細胞外
トラップを阻害し、急性腎傷害をレスキューする
3
第1章 序論
4
密接に関連する。この炎症では、特に好中球が形成する細胞外トラップ (Neutrophil extracellular trap, NET) が組織傷害を増悪させることが分かってきた6。活性化し
5
第2章 実験材料と実験方法
2 – 1. 実験動物 7〜12週齢のマウスを用いた。野生型マウス (C57BL/6J) は日本クレアから購入 した。CD169-DTRマウスは以前に当研究室で樹立したものを使用した14。実験に用 いたマウスは全て東京薬科大学実験動物管理施設のSPF環境下で飼育した。動物を 使用した実験は全て東京薬科大学動物実験委員会の承認のもと (承認番号:L14-21, L15-03, L16-14, L17-23, L17-24, L17-25) 、東京薬科大学動物実験規定およびガイ ドラインに従って行なった。 2 – 2. 細胞株NIH3T3細胞は Dulbecco’s modified Eagle’s medium (D-MEM; Wako) に10% 牛胎児血清 (FBS; Thermo Fisher Scientific) 、1% ペニシリン・ストレプトマイ シン (Wako) を添加した培養液で37℃、5% CO2存在下にて培養した。
2 – 3. 試薬
6
2 – 4. 細胞死アッセイ
NIH3T3細胞を7.5×103 cells/wellで96穴プレート (CORNING) に播種して、
37℃、5% CO2存在下にて24時間培養した。培地を新鮮な培養液に交換した後、tBHP
(最終濃度; 50 µM、培養時間; 12時間) 、または、エラスチン (最終濃度; 0.5 µM、 培養時間; 24時間) を添加して酸化ストレスに伴う細胞死を誘導した。細胞生存率 はCell Counting Kit-8 (DOJINDO LABORATORIES) を用いて酵素活性によって 評価した。細胞死阻害剤 (IM-93、DFO、Fer-1、Z-VAD) を、細胞死誘導剤を加え る直前に添加して細胞死阻害活性を評価した。
2 – 4. 腎臓IRIモデルマウス
1) 腎臓虚血再灌流傷害 (IRI) の誘導
IRは唐澤らの方法を参考にした15。麻酔下で、マウスの後腹膜左側に切り込みを
7 3) 細胞死阻害剤の投与実験 IRの3時間後に細胞死阻害剤を静脈内投与した。IM-93; 10 mg/kg 体重、 DFO; 100 mg/kg 体重、Fer-1; 5 mg/kg 体重で投与を行った。 4) 生存率評価 IRI誘導後に7日間、1日毎に生死を判定して生存率を算出した。 5) 生化学検査 IRI誘導前、および誘導後1日目、2日目に血清を採取して、血清クレアチニン濃 度を測定して腎機能を評価した。生化学検査はオリエンタル酵母工業に委託した。 2 – 5. フローサイトメトリー解析 1) 腎臓細胞の調製 腎臓細胞の分散は唐澤らの方法を参考にした15。マウスの腎臓を摘出して皮膜を 剥離した。細かく切り刻み、消化酵素溶液 (500 μg/ml Collagenase IIs、10 μg/ml DNase I / 1×HBSS (Ca2+, Mg2+ (+); Gibco)) に懸濁し、200 rpm/minで激しく攪拌
しながら、37℃で20分間、インキュベートした。細胞を1×HBSS(Ca2+, Mg2+ (-);
Gibco) で洗浄後、EDTA 溶液 (2 mM EDTA / 1×HBSS(Ca2+, Mg2+(-)) に懸濁して、
8
BD Biosciences) を用いて解析した。データ解析はFlowJoソフトウェア(Tree Star) で行った。
3) 抗体
Fc Blocker (抗マウスCD16/32抗体, clone; 2.4G2) はTONBO biosciencesから購 入した。抗マウスCD45.2抗体 (clone; 104) 、抗マウスCD11b抗体 (clone; M1/70) 、 抗マウスF4/80抗体 (clone; CI:A3-1) 、抗マウスLy6C抗体 (clone; HK1.4) 、抗マ ウスLy6G抗体 (clone; 1A8) 、死細胞除去のために用いた7-amino-actinomycin D (7-AAD) はBioLegendから購入した。 2 – 6. 免疫組織化学的染色 1) 腎臓組織における好中球の検出 腎臓の免疫染色は唐澤らの方法を参考にした15。腎臓を摘出してすぐに、OCTコ ンパウンド (サクラファインテックジャパン) に包埋して凍結ブロックを作製した。 クライオスタット (3050S, Leica) で5 µm厚の薄切切片を作成した。内因性のペル オキシダーゼ活性を除去するため、3% 過酸化水素/メタノール溶液で処理した。洗 浄後、内因性ビオチンをBiotin Blocking System (DAKO) で阻害した。TNブロッ キングバッファー (PerkinElmer) で非特異的結合をブロック後、ビオチン化抗マ ウスLy6G抗体 (clone; 1A8) を反応させ、、TSA BIOTIN SYSTEM Kit
(PerkinElmer) で検出した。核染色には 4',6-diamidino-2-phenylindole (DAPI; DOJINDO) を用い、FluorSave (ミリポア) で封入した。
2) 腎臓組織における好中球NET形成の検出
9
/ 1×HBSS (Ca2+, Mg2+ (+)) で処理を行った。ウサギ由来抗シトルリン化ヒストン
H3抗体 (anti-CitH3; Abcam)、続いてヤギ由来抗ウサギIgG抗体 (Invitrogen) を 反応させシトルリン化ヒストン(NET)を検出した。核染色にはDAPIを用い、 FluorSave (ミリポア) で封入した。
3) NET形成量の算出
染色した凍結切片を蛍光顕微鏡 (BZ-X710; Keyence) で観察した。Ly6G陽性、 あるいはCitH3陽性の領域面積をイメージ解析ソフトウェア (ImageJ) でそれぞれ 算出した。Ly6G陽性面積 (好中球) に占めるCitH3+ (NET形成部位) の面積を算出
した。
2 – 7. In vitro ヒト好中球NET形成誘導実験
1) ヒト末梢血好中球の単離精製
10
30分間前処理をした後、PMA (最終濃度 0.8 nM) を添加した。37℃、5% CO2存在
下で30分間培養後、SYTOX Green (最終濃度 0.5 µM) とHochest33342(Hoechst, 最終濃度 1 µg/ml)を添加し、さらに30分間培養した。
3) NET陽性好中球の検出
NET形成好中球数は四元らの方法を利用して定量した16。ハイコンテンツイメ
ージングシステムOperetta CLS (PerkinElmer) を用いて Sytox Green および Hoechst陽性細胞数を計測した。NET陽性好中球の割合は、Sytox Green 陽性細胞 数 (NET陽性細胞数) /Hoechst陽性細胞数 (総好中球数) x 100 (%) により算出した。
2 – 8. ROS産生量の定量
1) マウス骨髄好中球の単離精製
マウスの腓骨および大腿骨にMACS buffer (0.5 % BSA / 2 mM EDTA) を勢い よく流し入れ、骨髄細胞を採取した。70 µm セルストレーナー (BD Falcon) で濾 過し、500g、4℃で5分間遠心した。回収した細胞を10 μg/mL Fc Blocker溶液に懸 濁し、4℃、10分間インキュベートした。洗浄後、抗Ly6Gビオチン化抗体 (clone; 1A8, BioLegend) 溶液に懸濁し、4℃、25分間インキュベートした。洗浄後、抗ビオチン マイクロビーズ (Miltenyi) 溶液に懸濁し、4℃、15分間インキュベートした。洗浄 後、Ly6G+細胞を自動磁気細胞分離装置 (AutoMACS Pro; Miltenyi) で精製した。
11
2 – 9. 脂質過酸化量の定量
ヒト末梢血好中球を5×104 cells/wellで96穴丸底プレート (Thermo Fisher
Scientific) に播種し、IM-93、およびフェロトーシス阻害剤を添加して、37℃、5% CO2存在下にて30分間前処理をした後、PMA (最終濃度 1.6 nM) を添加した。37℃、
5% CO2存在下で30分間培養後、C11-Bodipy (最終濃度 2 µM) を添加し、さらに30
12
第
3 章 実験結果
3 – 1. 新規細胞死阻害剤 (IM-93) は酸化ストレスに伴う細胞死とフェロトーシス を抑制する IM-93は過酸化水素による細胞死阻害活性を指標に開発された薬剤である。本研 究では、このIM-93が抑制する細胞死様式を検討した。最初に、IM-93の、過酸化 水素以外の酸化ストレス誘導物質による細胞死阻害活性の有無を検討した。 NIH3T3細胞を細胞死阻害剤存在下あるいは非存在下で、50 M tBHPで処理し、 12時間後の細胞生存率をCell Counting Kit-8で定量した。図1に示す通り、IM-93 は過酸化水素だけでなく、tBHPによるNIH3T3の細胞死も効率よく抑制することが わかった (図1A) 。このように、IM-93は酸化ストレスによる細胞死を広く阻害す るが、この酸化ストレスによる細胞死は、形態学的にネクローシス様の細胞死であ ることは分かっているものの、細胞死実行の分子機構は分かっていない。最近、こ の酸化ストレスによる細胞死と関連する新たな細胞死としてフェロトーシスが定義 された。当初フェロトーシスは、Ras遺伝子に変異を有するがん細胞に誘導される 死として報告された17。この細胞死は、既知のregulated cell deathとは異なる細胞13 IM-93はこのエラスチンで誘導されるNIH3T3の細胞死を抑制できることが明らか になった (図1B) 。これらの結果から、我々は、酸化ストレスに伴う細胞死とフェ ロトーシスの間には、共通のメカニズムが存在すること、およびIM-93は他のフェ ロトーシス阻害剤と同様に、この2つの細胞死を抑制できる、と結論づけた。 3 – 2. IM-93はマウスの虚血再灌流傷害における急性腎障害を抑制する
IRIにおいて、活性酸素種 (Reactive Oxygen Species; ROS) による実質細胞の壊 死は、病態を規定する重要な因子である。最近、IRIにおける実質細胞の壊死には フェロトーシスが関与しており、フェロトーシス阻害剤の予防的投与が、IRIの程 度を軽減することが報告された5。IM-93が、フェロトーシス、および酸化ストレス
14 改善した (図2B) 。次に、炎症誘導後2日間の血清クレアチニン (Cr) 濃度を測定し た。糸球体でろ過されたCrは、尿細管で再吸収されず全量が尿中に排泄されるため、 腎機能を反映する指標として用いられる。DFO、およびFer-1投与群では、術後2日 目には血清Cr濃度が2.0 mg/dlを超え、腎機能が不可逆的に低下することが分かった。 一方、IM-93投与群では、血清Crが術後2日をピークに改善した (図2C) 。以上の結 果から、IM-93は虚血再灌流の誘導後に投与しても急性腎傷害を緩和できることが わかった。 3 – 3. IM-93は炎症部位への好中球浸潤数には影響を与えない
DFO, Fer-1がin vitroにおけるNIH3T3のフェロトーシス、および酸化ストレスに 伴う細胞死はIM-93と同程度かそれ以上に抑制するのに対して、IRに伴うAKIでは IM-93だけが強い治療効果を発揮できる理由を考察した。CD169発現マクロファー ジを消失させると、炎症部位への好中球浸潤が亢進する。マクロファージ消失マウ スでは血管内皮の白血球接着因子ICAM-1の発現が亢進する。抗Gr-1抗体による好 中球の消去や、好中球と血管内皮との接着を阻害する抗ICAM-1抗体は、 CD169-DTRマウスの死亡率を改善する15ことから、IM-93は、腎臓への好中球浸潤 を抑制することによって、AKIにおける炎症を緩和しているのではないかと考えた。 そこで、CD169発現マクロファージを消失させたCD169-DTRマウスにIRIを誘導し、 再灌流の3時間後にIM-93を尾静脈投与した。IR8時間後の腎臓における好中球の割 合をフローサイトメトリーで調べた。腎臓の好中球 (図3A; 7AAD−、CD45.2+、
CD11b+、Ly6G+) の割合は、Vehicle (生理食塩水) 投与群では、sham control群の
19.1%から48.2%に増加していた (図3B) が、Ly6C+単球 (図3A; 7AAD−、CD45.2+、
CD11b+、Ly6G−、Ly6C+) の割合には変化がなかった (図3C) 。一方で、AKIの改
15 解析の結果と同様に、生理食塩水投与群とIM-93投与群の好中球数に有意な差はな かった (図3D) 。これらの結果から、IM-93の腎IRIに伴うAKIの炎症抑制効果は、 炎症部位への好中球浸潤を阻害することよる効果ではなく、集積した好中球の活性 化状態および炎症応答を抑制することによるものである可能性が示唆された。 3 – 4. IM-93は好中球の細胞外トラップ (NET) 形成を抑制する 炎症部位に集積した活性化好中球は、クロマチンを脱凝集し、顆粒内タンパク質 とともに細胞外に放出する。これは好中球細胞外トラップ (neutrophil
extracellular trap; NET) と呼ばれ、NET形成を伴う好中球の死をネトーシス (NETosis) と呼ぶ。NET形成は病原性微生物の捕獲と殺菌に有効な21一方で、過剰
な顆粒タンパクが2次的な組織傷害の原因となると考えられており22、IRIでは、こ
の好中球のNET形成がさらなる組織傷害を引き起こすと考えられている6 。
そこで、IM-93のNET形成阻害効果をまずin vitroで検討した。ヒト末梢血好中球 をPMAで刺激し、Hoechstと、SYTOX Greenで染色した16。Hoechstと異なり、
16
3Dで示したように、IM-93投与マウスの腎臓における浸潤好中球量は、非投与群と 同程度であった。しかし、CitH3陽性好中球の割合は、IM-93投与群で有意に減少 した (図4D) 。
18
19 図2 IM-93は虚血再灌流に伴う急性腎傷害の病態を改善する (A) 薬剤投与のタイムコース。CD169-DTRマウスにDTを腹腔内投与し、CD169発 現マクロファージを一過性に消失させた後、腎虚血再灌流傷害を誘導した。再灌流 の3時間後に細胞死阻害剤を静脈内投与した。 (B) 術後7日間の生存率。フェロトーシス阻害剤 (DFO, Fer-1) 投与群では、不可逆 的に腎機能障害が進行し、術後5日目までに全例が死亡した。IM-93投与群では、 control (コントロール、薬剤非投与群) と比較し、生存率が有意に改善した。(C) 虚 血再灌流後の血清クレアチニン値。Controlやフェロトーシス阻害剤投与マウスの血 清クレアチニン値は術後2日目までに約2.0 g/mlまで上昇したのに対し、IM-93投 与マウスでは血清クレアチニン値の上昇が有意に抑制された。*< 0.05。
20
3
A
S S C F S C FSC FSC 7AAD C D 45 .2 C D 45 .2 CD11b Ly6G L y6 C 29.9Sham operation kidney
21
図3 IM-93は炎症局所への好中球浸潤を抑制しない
(A-C) 虚血再灌流傷害の誘導8時間後の腎臓における、好中球 (7AAD-, CD45.2+,
CD11b+, Ly6G+) 、および単球 (7AAD-, CD45.2+, CD11b+, Ly6C+) の割合をフロ
22
4
A
Hoechst
SYTOX
PMA + IM-93 PMA + DFO PMA + Fer-1
23 図4 IM-93は好中球のNET形成を抑制する
24
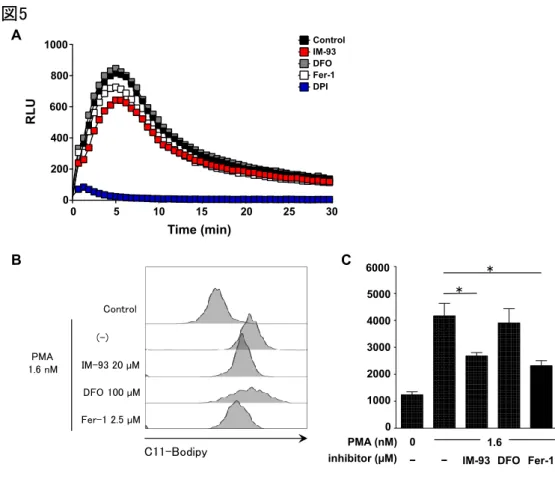
図5 IM-93は脂質過酸化の抑制を介して、好中球のNET形成を抑制する
(A) IM-93はROS産生を阻害しない。PMA (1 µM) で刺激後の、マウス骨髄好中球 におけるROS産生量をルミノール反応を指標にリアルタイムで測定した。IM-93; 10 µM、DFO; 100 µM、Fer-1; 2.5 µM、DPI; 10 µM。NADPHオキシダーゼ阻害剤 のDPI (青) は、PMA刺激によるROSの産生を完全に抑えた。一方、IM-93 (赤) 、 およびフェロトーシス阻害剤 (DFO; 灰色、Fer-1; 白)はいずれも、ROS産生量を 抑制しなかった。(B, C) IM-93とFer-1は脂質過酸化を阻害する。ヒト末梢血好中 球をIM-93、あるいはフェロトーシス阻害剤で処理した後、PMA (1.6 nM, 1時間) で 刺激した。(B) C-11 Bodipyは酸化されると緑色蛍光を発する色素。C-11 Bodipyの 蛍光量を指標に、脂質過酸化の程度を定量した。IM-93;20 µM、DFO; 100 µM、 Fer-1;2.5 µM。(C) (B) のヒストグラムの平均蛍光強度をグラフ化した。DFOでは 脂質過酸化物量に変化は見られなかったが、Fer-1とIM-93は同程度に減少した。 R L U 0 200 400 600 800 1000 IM-93 Fer-1 DFO DMSO DPI IM-93 Fer-1 DFO DMSO DPI Control DPI IM-93 DFO Fer-1 Time (min) 0 5 10 15 20 25 30 A B C 1000 2000 3000 4000 5000 6000 0 0 − 1.6 PMA (nM)
28
第
5 章 参考文献
1. Linkermann, A. Nonapoptotic cell death in acute kidney injury and transplantation. Kidney Int 89, 46-57 (2016).
2. Linkermann, A. et al. Two independent pathways of regulated necrosis mediate ischemia-reperfusion injury. Proc Natl Acad Sci U S A 110, 12024-12029 (2013).
3. Dixon, S.J. et al. Ferroptosis: an iron-dependent form of nonapoptotic cell death. Cell 149, 1060-1072 (2012).
4. Friedmann Angeli, J.P. et al. Inactivation of the ferroptosis regulator Gpx4 triggers acute renal failure in mice. Nat Cell Biol 16, 1180-1191 (2014).
5. Linkermann, A. et al. Synchronized renal tubular cell death involves ferroptosis. Proc Natl Acad Sci U S A 111, 16836-16841 (2014).
6. Savchenko, A.S. et al. VWF-mediated leukocyte recruitment with chromatin decondensation by PAD4 increases myocardial ischemia/reperfusion injury in mice. Blood 123, 141-148 (2014).
7. Brinkmann, V. et al. Neutrophil extracellular traps kill bacteria. Science 303, 1532-1535 (2004).
8. Jorch, S.K. & Kubes, P. An emerging role for neutrophil extracellular traps in noninfectious disease. Nat Med 23, 279-287 (2017).
9. Branzk, N. & Papayannopoulos, V. Molecular mechanisms regulating NETosis in infection and disease. Semin Immunopathol 35, 513-530 (2013).
10. Wong, S.L. et al. Diabetes primes neutrophils to undergo NETosis, which impairs wound healing. Nat Med 21, 815-819 (2015).
29
12. Dodo, K., Katoh, M., Shimizu, T., Takahashi, M. & Sodeoka, M. Inhibition of hydrogen peroxide-induced necrotic cell death with 3-amino-2-indolylmaleimide derivatives. Bioorg Med Chem Lett 15, 3114-3118 (2005).
13. Sodeoka, M. & Dodo, K. Development of selective inhibitors of necrosis. Chem Rec 10, 308-314 (2010).
14. Miyake, Y. et al. Critical role of macrophages in the marginal zone in the suppression of immune responses to apoptotic cell-associated antigens. J Clin Invest 117, 2268-2278 (2007).
15. Karasawa, K. et al. Vascular-Resident CD169-Positive Monocytes and Macrophages Control Neutrophil Accumulation in the Kidney with Ischemia-Reperfusion Injury. J Am Soc Nephrol 26, 896-906 (2015).
16. Yotsumoto, S. et al. Hyperoxidation of ether-linked phospholipids accelerates neutrophil extracellular trap formation. Sci Rep 7, 16026 (2017).
17. Dolma, S., Lessnick, S.L., Hahn, W.C. & Stockwell, B.R. Identification of genotype-selective antitumor agents using synthetic lethal chemical screening in engineered human tumor cells. Cancer Cell 3, 285-296 (2003).
18. Yang, W.S. et al. Regulation of ferroptotic cancer cell death by GPX4. Cell 156, 317-331 (2014).
19. Barbosa, P.R. et al. Inhibition of mitochondrial respiratory chain in the brain of rats after renal ischemia is prevented by N-acetylcysteine and deferoxamine. Metab Brain Dis 25, 219-225 (2010).
20. Bernardi, R.M. et al. N-acetylcysteine and deferrioxamine protects against acute renal failure induced by ischemia/reperfusion in rats. Rev Bras Ter Intensiva 24, 219-223 (2012).
30
22. Yipp, B.G. & Kubes, P. NETosis: how vital is it? Blood 122, 2784-2794 (2013).
23. Leshner, M. et al. PAD4 mediated histone hypercitrullination induces heterochromatin decondensation and chromatin unfolding to form neutrophil extracellular trap-like structures. Front Immunol 3, 307 (2012).
24. Masuda, S. et al. NETosis markers: Quest for specific, objective, and quantitative markers. Clin Chim Acta 459, 89-93 (2016).
25. Sorensen, O.E. & Borregaard, N. Neutrophil extracellular traps - the dark side of neutrophils. J Clin Invest 126, 1612-1620 (2016).
26. Kessenbrock, K. et al. Netting neutrophils in autoimmune small-vessel vasculitis. Nat Med 15, 623-625 (2009).