平 成 2 5 年 度
新 潟 市 衛 生 環 境 研 究 所 年 報
( 調査研究報告 )
第 38 号
Annual Report
of
Niigata City Institute of Public Health and Environment
新 潟 市 衛 生 環 境 研 究 所
も く じ
(1)16SrRNA遺伝子の塩基配列解析を用いたレジオネラ属菌の同定--- 1
(2)Campylobacter jejuni におけるギラン・バレー症候群関連遺伝子の保有状
況について
-
---
5
(3)環境水などから検出した大腸菌の病原因子保有率について(第2報)---
9
(4)新潟市における 2013/14 シーズンのインフルエンザウイルス分離・検出状況
について--- 12
(5)新潟市における 2012/13 および 2013/14 シーズンのインフルエンザウイルス
薬剤耐性遺伝子について状況--- 15
(6)下水における下痢症ウイルス遺伝子の検出状況について(第2報)--- 20
(7)市販二枚貝からの下痢症ウイルス遺伝子の検出状況について(第2報)
-
-
25
(8)新潟市河川における水生底生生物実態調査(第12報)---
-
-
32
(9)生物指標による水質評価法の検討について
-
---
41
(10)新潟市における酸性雨の状況調査---
-
-
45
(11)新潟市沿岸海域における水質の現状と汚濁調査について---
-
-
54
(12)新潟市における海塩影響調査(第4報)---
-
-
65
- 1 - 調査研究報告(1)
16S rRNA 遺伝子の塩基配列解析を用いたレジオネラ属菌の同定
山本 一成
概 要
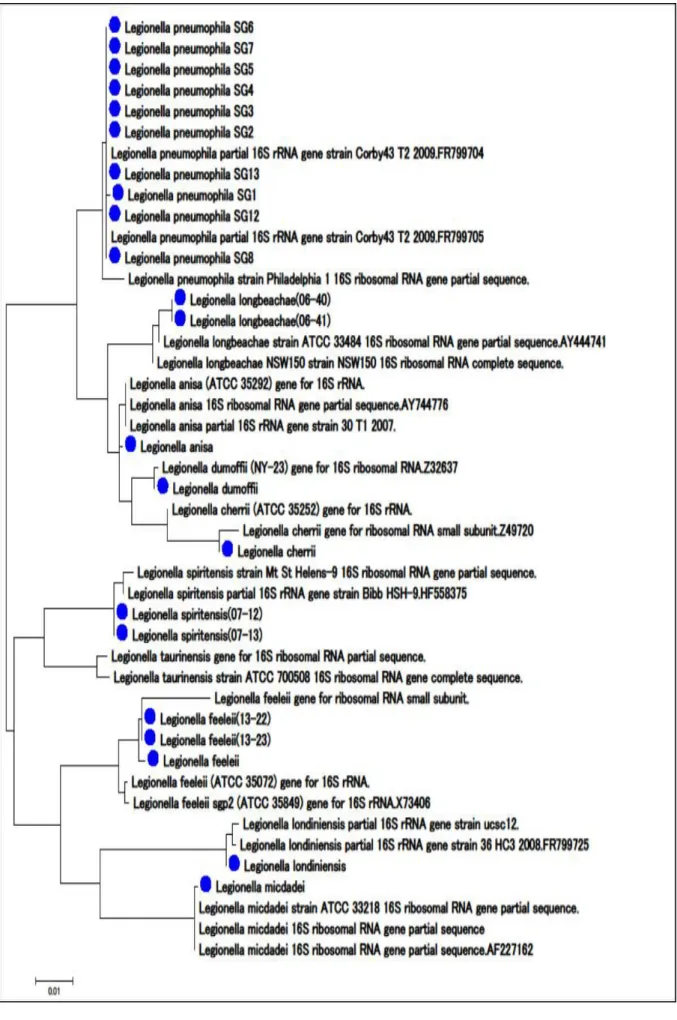
近年,分離菌同定の迅速法として 16S リボゾーム RNA(16S rRNA)遺伝子解析が利用されて いる。一般的にレジオネラ属菌の種鑑別法として特異抗血清,イムノクロマトおよび核酸を用 いた同定法がある。今回,当所の保存菌株で同定されているもの 16 株および過去の手法で同定 できず保存されていた 6 株について 16SrRNA 遺伝子解析によるレジオネラ属菌の同定について 検討した。同定されていた 16 株は,過去の同定と一致した。また,未同定たった 6 株はLegionellalongbeachae(L. longbeachae )2 株,Legionella spiritensis (L. spiritensis) 2 株,およびLegionella feeleii(L. feeleii) 2 株と 16S rRNA 遺伝子解析で同定された。 また,各菌株は系統樹解析において各々のクラスターに分類された。
1 はじめに
レジオネラ属菌は,河川や湿った土壌などの自然界広く分布しており,細菌や藻類などの代 謝産物を栄養源としたり,または,細菌捕食性のアメーバなどの原虫の細胞内で増殖する細胞 内寄生性細菌である。レジオネラ属菌は 2011 年 8 月時点で 52 種の菌種が報告されている1)。 レジオネラ属菌は,生化学的性状試験での菌種鑑別・同定は非常に難しい。同定方法としては、 市販の特異抗血清による方法,イムノクロマト法による方法および核酸を用いた方法がある。 核酸を用いた方法には,ポリメラーゼ連鎖反応によるLegionella pneumophilaとレジオネラ属 菌の同定(PCR 法),DNA-DNA ハイブリダイゼーション( DDH レジオネラ’極東’)による同定 および 16S rRNA 遺伝子解析によるシークエンス法がある。 16S rRNA は,生物の本質に関わる機能を持った部分であり,遺伝子の長さが約 1,500bp で塩 基配列の保存性が高い部分と変異しやすい部位も含まれており,近縁種でも比較が可能である と言われている2)。米国薬局方,ヨーロッパ薬局方および世界中の監督官庁は,細菌および親 近の遺伝子解析による同定を推奨している。また,日本においても 2006 年に,日本薬局方は参 考情報として,遺伝子解析による微生物の迅速同定法に関する章を追加し,16S rRNA 遺伝子解 析による同定が多く利用されている3)。今回,保存菌株を用いて 16S rRNA 遺伝子解析を行い若 干の知見を得たので報告する。2 材料と方法
1)材料
当所で過去に分離し保存されている菌株,Legionella pneumophila(L. pneumophila)血清 群 1~8,12,13,, Legionella Cherrii(L. Cherrii),Legionella feeleii(L. feeleii),
Legionella anisa(L. anisa), Legionelladumoffii(L. dumoffii),Legionella micdadei
(L. micdadei), Legionellalondiniensis(L. londiniensis)の 16 株および未同定 6 株を 材料とした。
2)
方法
保存菌株を BCYEα培地に起し,国立感染症研究所の病原体検出マニュアル,レジオネラ症1))
に準じて実施した。塩基配列の決定には,Applied Biosysytems 3500 ジェネティックアナライ ザーを用い,ダイレクトシーケンス法により行った。
- 2 -
得られた塩基配列を DNA Data Bank of Japan(DDBJ) の BLAST(http://blast.ddbj.nig.ac.jp/
top.-j.html)で相同性解析を行い,遺伝子解析ソフト MEGA5 を用いて系統樹解析等を実施した。
3 結果
DDBJ BLAST における相同性解析の結果,100 から 98%と高い相同性が得られた。また,アク セッション番号の菌株との距離は,0.000 から 0.009 であった(表)。また,Legionella属菌 6 株は,L. longbeachae 2 株, L. spiritensis 2 株,およびL. feeleii 2 株と 16S rRNA 遺伝 子解析で同定された。系統樹解析においても各々のクラスターに分類された(図)。
4 考察
今回の結果より,16S rRNA 遺伝子解析および系統樹解析を用いることは,通常の方法で同定 できないときに有用な手法と思われた。しかし,L.pneumophilaの血清群の鑑別は不可能であ り,ヒトへの感染において報告数の多いL.pneumophila SG1 の同定には,特異抗血清との組み 合わせが必要である。 今後,特異抗血清と 16S rRNA 遺伝子解析を組み合わせることにより,正確な同定が可能と思わ れる。 今回は,レジオネラ菌について 16S rRNA 遺伝子解析の検討を行ったが,16S rRNA は全ての 細菌に普 遍的に存在し機能的に保存されているため,近年,微生物の迅速法として使用されるている。 このことより,他の菌においても生化学的性状等で同定不可能の場合,利用可能と思われるた め,今後も検討していく必要がある。文献
1) 国立感染症研究所:病原体検出マニュアル,レジオネラ症(平成 23 年 10 月 7 日改訂). 2) Wikipedia:
16S rRNA 系統解析, https://www.google.co.jp/?gfe_rd=cr&ei=S8hwU-boC8bJ8geLmYGwCw#q=16S+rRNA.- 3 -
表 アクセッション番号の菌株と相同性および距離
同 定 株
ACCESSION Ident Distance
Legionella_pneumophila_SG1
FR799704
100
0.000
Legionella_pneumophila_SG2
FR799705
100
0.000
Legionella_pneumophila_SG3
FR799705
100
0.000
Legionella_pneumophila_SG4
FR799705
100
0.000
Legionella_pneumophila_SG5
FR799705
100
0.000
Legionella_pneumophila_SG6
FR799705
100
0.000
Legionella_pneumophila_SG7
FR799705
100
0.000
Legionella_pneumophila_SG8
FR799705
100
0.000
Legionella_pneumophila_SG12
FR799705
100
0.000
Legionella_pneumophila_SG13
FR799705
100
0.000
Legionella_cherrii
Z49720
99
0.006
Legionella_feeleii
X73406
98
0.009
Legionella_anisa
AY744776
99
0.002
Legionella_dumoffii
Z32637
99
0.001
Legionella_micdadei
AF227162
99
0.001
Legionella_londiniensis
FR799725
99
0.002
Legionella_longbeachae(06-40) AY444741
99
0.005
Legionella_longbeachae(06-41) AY444741
99
0.005
Legionella_spiritensis(07-12)
HF558375
99
0.004
Legionella_spiritensis(07-13)
HF558375
99
0.004
Legionella_feeleii(13-22)
X73406
98
0.009
Legionella_feeleii(13-23)
X73406
98
0.009
- 4 -
- 5 - 調査研究報告(2)
Campylobacter jejuni
におけるギラン・バレー症候群関連遺伝子の保
有状況について
山本一成 齊藤哲也概 要
Campylobacter jejuni(以下 C. jejuni)はギラン・バレー症候群を高頻度に誘発する原因 菌として知られている。下痢症患者便,牛の便や胆汁,および市販の鶏肉から検出された C. jejuniのギラン・バレー症候群発症に係る遺伝子の保有状況について調査した。その結果,最 もギラン・バレー症候群を発症させるリスクの高いC. jejuniは牛よりも鶏に多く分布してい ると思われる。
1 はじめに
カンピロバクターは,発熱,腹痛,下痢,血便などの消化器症状を引き起こす食中毒の起因 菌である。現在日本では,細菌性食中毒事件の原因菌として上位を占めており,本市において もカンピロバクターを原因菌とする食中毒事件が年間を通して 1~2 件発生している。 カンピロバクターによる腸炎症状の予後は一般的に良好であるが,まれに多発性神経障害の 一種であるギラン・バレー症候群(Guillain-Barré syndrome:以下 GBS)を発症することがある。 特にC. jejuniは,GBS を高頻度に発症させる先行感染の原因とされている。 C. jejuniによる GBS の発症機序には,菌体表層にあるリポオリゴ糖(以下 LOS)が関与してい る。この LOS の一部に神経細胞を構成するガングリオシド(GM1,GD1a)との分子相同性を有する ものがある。C. jejuni に感染した場合に生体内で作り出される抗体が,ガングリオシド様と なり,交叉抗体として神経細胞に作用することで GBS が発症するとされている。C. jejuni 菌 外膜に,このガングリオシド様 LOS が形成されるには,シアル酸転移酵素,N-アセチルガラク トサミン転移酵素,およびガラクトース転移酵素の 3 種類の酵素が必要である。この 3 種転移 酵素遺伝子をコードする遺伝子領域 cstⅡ,cgtA および cgtB の保有状況により C. jejuni の GBS 発症に関するリスクを評価することが可能である1)。 今回,牛由来,市販鶏肉由来および下痢症患者便由来のC. jejuniについて転移酵素遺伝子 の保有状況を調査したので報告する。2 材料および方法
〈材料〉 C. jejuni菌株収集のために牛の便 30 件,牛の胆汁 30 件,市販の鶏肉 8 件を検査した。 その内,牛の便由来の 12 株,牛の胆汁由来の 7 株,市販鶏肉由来の 5 株,および 2007 年から 2010 年までに分離された下痢症患者由来の 22 株,計 46 株を検体とした。 〈PCR 法〉M. Koga2),Irving Nachamkin,3)らが報告した cstⅡ,cgtA および cgtB を検出するプライ
マーを用い,PCR 法にて検査を行った。試薬は TaKaRa Taq を用いた。 〈血清型別試験〉
カンピロバクター免疫血清(デンカ生研)を用いて Penner 血清群別試験を行った。既存の血 清で凝集が見られないものを Unknown Type (以下 UT)とした。
- 6 -
3 結果
結果を図 1 および表 1 に示す。牛の便および胆汁から検出されたC. jejuniは cstⅡを 19 株中 8 株(44%)が保有しているものの,他の cgtA,cgtB は保有していなかった。一方で, 鶏肉から検出されたC. jejuniでは 8 株中 5 株(63%)が cstⅡ,cgtA および cgtB をすべて 保有していた。下痢症患者便から検出されたC. jejuniは 22 株中 3 株(14%)が 3 種の遺伝 子をすべて保有していた。また cstⅡを単独で保有するものが 22 株中 2 株(9%)であった。 血清群別結果と GBS 関連遺伝子との内訳を表 2,表 3 および表 4 に示す。B 群 3 株(牛の胆 汁由来 1 株,患者便由来 2 株),D 群 4 株(牛の便由来 1 株,牛の胆汁由来 1 株,患者便由来 2 株),L 群 1 株(鶏肉由来 1 株),C 群,F 群,G 群,K 群,Y 群,各 1 株(すべて患者由来), UT が 33 株であった。 1~5:鶏肉由来,6~17:牛の便由来,18~24:牛の胆汁由来 N:Negative control 図1 PCR による cstⅡ,cgtA および cgtB の泳動結果 1 2 3 4 5 6 7 8 9 10 11 12 13 14 15 16 17 18 19 20 21 22 23 24 N 1 2 3 4 5 6 7 8 9 10 11 12 13 14 15 1 2 3 4 5 6 7 8 9 10 11 12 13 14 15 16 17 18 19 20 21 22 23 24 N 16 17 18 19 20 21 22 23 24 N cstⅡ 204bp cgtA 527bp cgtB 502bp- 7 - 表 1 GBS 関連遺伝子の保有状況 調査数 C. jejuni陽性数 cstⅡ cgtA cgtB 牛の便 30 12 6 0 0 牛の胆汁 30 7 2 0 0 鶏肉 8 5 5 5 5 患者便 22 5 3 3 表 2 牛 の 便 お よ び 胆 汁 由 来 の C . j e j u n i の 血 清 群 と G B S 関 連 遺 伝 子 の 検 査 結 果 表 3 市 販 鶏 肉 由 来 の C . j e j u n i の 血 清 群 と G B S 関 連 遺 伝 子 の 検 査 結 果 表 4 下 痢 症 患 者 便 由 来 の C . j e j u n i の 血 清 群 と G B S 関 連 遺 伝 子 の 検 査 結 果
4 考察
今回の調査では,最も GBS 発症のリスクが高い cstⅡ,cgtA および cgtB を同時に保有する C. jejuni は鶏肉由来もしくは下痢症患者由来であった。牛由来,および下痢症患者由来と比 較すると,鶏肉由来において GBS を高頻度に発生させる可能性のあるC. jejuni が最も優位で あった。 下痢症患者便由来のC. jejuni は鶏肉由来には劣るものの,比較的高い割合(22 株中 3 株 の 14%)で cstⅡ,cgtA および cgtB を同時に保有していた。この患者の下痢発症時の喫食状 遺伝子保有パターン 血清群 Penner cstⅡ cgtA cgtB B D L UT 合計 + + + + - - 2 6 8 - - - 1 11 11 遺伝子保有パターン 血清群 Penner cstⅡ cgtA cgtB B D L UT 合計 + + + 1 4 5 + - - - - - 遺伝子保有パターン 血清群 Penner cstⅡ cgtA cgtB B C D F G K L Y UT 合計 + + + 1 2 3 + - - 1 1 2 - - - 1 1 2 1 1 11 17- 8 - 況を確認することはできなかったが,鶏肉を喫食していた可能性が示唆される。 一方で,牛の便および胆汁から検出された C. jejuniの一部は cstⅡのみ保有しており,他 の cgtA および cgtB は保有していなかった。 血清群については日本では GBS 患者から Penner O 群が多く検出されるとの報告があるが,一 方で国外での研究では関連がないとの報告もある4)。今回の調査では cstⅡ,cgtA および cgtB を同時に保有するC. jejuni 1 株(患者便由来)が B 群を示し,2 株(患者便由来)が D 群を示 した。更に cstⅡを単独で保有するC. jejuni 1 株(患者便由来)も B 群を示し,2 株(牛由来) が D 群を示した。GBS 関連遺伝子を保有するC. jejuniの血清群が B 群もしくは D 群である確 率が高いことが示唆されるが,この点については更なる調査を進める必要がある。 また菌株収集に用いた検体のうち,市販鶏肉からのC. jejuniの検出率は 8 株中 5 株で 63% であった。市販されている未加熱の鶏肉は高い確率でC. jejuniに汚染されていることがわか った。 今後,保健所と連携してC. jejuniの感染予防に資するために,市販鶏肉の検査数を増やし, 新たに食中毒等で分離されたヒト由来の菌株の検査を加え,GBS 発症に関与する転移酵素遺伝 子の保有状況を引き続き調査していく必要がある。
参考文献
1) Michiaki Koga,:Comprehensive analysis of bacterial risk factors for the development of Guillain-Barré syndrome after Campylobacter jejuni enteritis,J Infect Dis. (2006) 2) Michiaki Koga,: Campylobactor gene polymorphism as a determinant of clinical features of Guillain-Barré syndrome,AAN Enterprises, Inc.(2005)
3) Irving Nachamkin,: Campylobactor jejuni from Patients with Guillain-Barré Sundrome Preferentially Expresses aGD1a-Like Epitope,Infection and Immunity.(2002)
4) Parker, C.T.et al: Comparison of Campylobacter jejuni lipooligosaccharide biosynthesis loci from a variety of sources, J. Clin.Microbiol.,Jun.,p.2771~2781(2005)
- 9 - 調査研究報告(3)
環境水などから検出した大腸菌の病原因子保有率について(第2報)
足立玲子 高橋裕子概 要
平成 24 年度に引き続き,河川水及び下水から分離した大腸菌 157 株について大腸菌病原因子 のうち 9 種のマルチプレックス PCR を実施した。その結果 eae,astA,afaD,aggR 遺伝子を 検出し,病原因子の検出率は 8.3 %であった。はじめに
大腸菌(Escherichia coli)はヒトなどの腸管内正常細菌叢の一つであり,ほとんどは病原 性を示さないが,一部に下痢などを引き起こすものがあり,「下痢原性大腸菌」と総称されてい る。 下痢原性大腸菌は乳幼児の胃腸炎の病原菌として 1927 年 T.Adam(アダム)が最初に報告し たとされ,その後も多くの報告がなされてきた1)。 2012 年 1 月から下痢原性大腸菌の分類が改定され,腸管出血性大腸菌(以下 EHEC),腸管毒 素性大腸菌(以下 ETEC),腸管侵入性大腸菌(以下 EIEC),腸管病原性大腸菌(以下 EPEC), 腸管付着性大腸菌(以下 EAggEC),その他の下痢原性大腸菌の 6 種類に区別するようになった。 これらの下痢原性大腸菌と正常細菌叢の大腸菌とを区別するためには病原因子を検索する必 要がある2)。 これらの人を取り巻く環境中での分布状況を把握し,食品などへの汚染要因を検討する基礎 資料とするため,河川水と,下水処理場の浄化前の流入水(以下 下水)について昨年度に引 き続き病原因子保有状況を調査したので報告する。材料
昨年度と同様に環境基準監視のための河川水および下水を検体とした。河川水は平成 25 年 5, 8,11 月,平成 26 年 2 月に,下水については平成 25 年 5,6,7,9,10,11 月,平成 26 年 1, 2 月に検査を実施した。方法
材料からの大腸菌株の採取および病原因子の検索は,昨年度と同様に行った。 すなわち,河川水については「要測定指標(大腸菌)の測定について」3)に従い測定し,測 定後の大腸菌のコロニーを無作為に供試した。大腸菌数測定に用いた酵素基質培地は ES コリマ ーク寒天培地(栄研化学)を使用した。 大腸菌と判定した菌株について,平成 23 年度細菌研修テキストの方法より大腸菌病原因子の PCR を実施した。EHEC については VT 遺伝子,ETEC については LT,ST 遺伝子,EIEC については invE 遺伝子,EPEC については eae 遺伝子,bfpA 遺伝子,EAggEC については aggR 遺伝子を,そ の他の下痢原性大腸菌については afaD 遺伝子,astA 遺伝子の 9 遺伝子についてマルチプレッ クス PCR を実施した。 病原因子を検出した株について,病原大腸菌免疫血清(デンカ生研)1号セットを用いたス ライド凝集反応による O 群血清型別を実施した。O 群が決定した株については,さらに同免疫 血清2号セットを用いて試験管内での凝集での H 型別を行った。 下水については,ES コリマーク寒天培地と下水を混釈し,37℃20 時間培養後のコロニーを無- 10 -
作為に釣菌し,性状確認した菌株を河川水と同様に実施した。
結果
河川水 98 株,下水 59 株の計 157 株について大腸菌病原因子を検索した。
PCR による検出状況およびその血清型別を表1に示した。eae 遺伝子を河川水から 5 株,下水 から 2 株検出した。aggR 遺伝子を下水から 1 株,astA 遺伝子を河川水から 4 株そして afaD 遺 伝子を下水から1株検出した。
VT 遺伝子, LT 遺伝子,ST 遺伝子,bfpA 遺伝子および invE 遺伝子は,河川水,下水からの いずれからも検出しなかった。
病原因子別にみると,eae 遺伝子が 4.4%,astA 遺伝子 2.5%,afaD 遺伝子 0.6%,aggR 遺 伝子 0.6%の検出率であり,検体別では,河川水 9.2%,下水 6.8%であった。
全体では,8.3%であり,複数の病原因子を保有する株はなかった。
血清型別では,下水中の eae 遺伝子保有株 1 株が O63:H6,同じく afaD 遺伝子を持った株が O168:HNM(NM は非運動性)であった。他の病原遺伝子が検出された 11 株は全て OUT であった。
表1.病原因子検出状況と血清型別(平成 25 年度)
河川水 血清型別 下水 血清型別 病原因子別検出数
(率%)
eae 遺伝子 5 全て OUT 2 O63:H6,OUT 7 (4.4)
astA 遺伝子 4 全て OUT 4 (2.5) afaD 遺伝子 1 O168:HNM 1 (0.6) aggR 遺伝子 1 OUT 1 (0.6) 小計 9 4 13 検出率(%) 9.2 6.8 8.3
考察
昨年度に引き続き,環境での循環水を検体として,ヒトの下痢症にかかわる大腸菌の病原因 子の保有状況を調査した。 平成 25 年度の調査では病原因子を検出した大腸菌は 13 株(6 パターン)8.3%であった,VT 遺伝子,LT,ST 遺伝子,invE 遺伝子,bfpA 遺伝子は検出されなかった。昨年度検出されなか った aggR 遺伝子が下水から検出された。 表 2.由来別の病原因子検出率の年度別状況(検出率 %) 河川水・H24 河川水・H25 下水・H24 下水・H25 VT 遺伝子 0.4 0 0 0 eae 遺伝子 3.4 5.1 0 3.4 astA 遺伝子 9.6 4.1 3.8 0 afaD 遺伝子 0.7 0 0 1.7 aggR 遺伝子 0 0 0 1.7 いずれかの保有 14.4 9.2 3.8 6.8 昨年度の結果と比較すると,VT および aggR 遺伝子で検出,不検出の差はあったが,いずれ も検出率は著しく低く,他の病原因子についても検出率を考慮すると大きな差異とは認められ- 11 - ないと思われる。 また,昨年度からを通じて,下水より河川水の方が全体の病原性因子の検出率が高い傾向が うかがわれる。 今後さらにデータを重ねて,上記のような傾向を確認するとともに,食品など人との接触の より高い材料や胃腸炎の起因となったものとの比較検討が必要である。
参考文献
1)坂崎利一 編集:新訂 食水系感染症と細菌性食中毒 中央法規 2000 年 2)特集 下痢原性大腸菌 2011 年現在:病原微生物検出情報 Vol.33 No.1 2012 年 1 月 3)平成 23 年 3 月 24 日付け環水大水発第 110324001 号 4)環境水などから検出した大腸菌の病原因子保有率について 新潟市衛生環境研究所平成 24 年度年報調査研究報告(2)- 12 - 調査研究報告(4)
新潟市における 2013/14 シーズンのインフルエンザウイルス分離・検出
状況について
小林 元概 要
新潟市における 2013/14 シーズンのインフルエンザ患者報告数は 2014 年第 2 週(2014 年 1 月6 日から 12 日)に定点あたり報告数が 1.00 を上回り,2014 年第 9 週(2014 年 2 月 24 日から 3 月 2 日)に 60.85 となり,ピークを迎えた。今シーズンのインフルエンザウイルスの分離・検 出状況は 2013 年第 52 週までは AH3 亜型インフルエンザウイルス(以下 AH3 亜型)が主流 であったが,2014 年第 1 週以降は AH1pdm09 インフルエンザウイルス(以下 AH1pdm09) が主流となった。B 型インフルエンザウイルス(以下 B 型)は山形系統,ビクトリア系統とも に検出されたが,大半が山形系統であった。分離したインフルエンザウイルスの HI 試験を行 ったところ,AH1pdm09 および B 型はすべてワクチン類似株であった。AH3 亜型では流行後 期に抗原性の異なる株が検出された。1.はじめに
インフルエンザは冬期に発生,患者が次第に増加し 4~5 月にかけて減少する季節性流行を 毎年繰り返し,年によって差はあるものの推定国内で平均1,000 万人以上の罹患者が発生する 身近な疾患である。また,「感染症の予防及び感染症の患者に対する医療に関する法律」で 5 類感染症に分類され,感染症法に基づく感染症発生動向調査事業においては定点把握対象疾患 として,全国約5,000 のインフルエンザ定点医療機関(小児科 3,000,内科 2,000)から週ごとに, 患者の発生状況などの情報が収集・解析されている。さらに,地方衛生研究所では主に病原体 定点(インフルエンザ定点の約10%)で採取された検査材料からウイルス分離を実施し,抗原 解析などの性状検査を実施している。この検査結果は病原体検出情報システムに登録し,国立 感染症研究所感染症情報センターで集約され,インフルエンザの流行予測およびワクチン株選 定に活用されている。当所においても感染症発生動向調査の一環として,インフルエンザウイ ルスの分離・検出を行っており,2013/14 シーズンのインフルエンザウイルスの分離・検出状 況と分離株の性状について若干の知見を得たので報告する。2.材料と方法
2013 年第 36 週から 2014 年第 23 週までに病原体定点医療機関などでインフルエンザあるい はインフルエンザ様疾患と診断された患者より採取された咽頭ぬぐい液などの検体を材料とし てウイルス分離培養を行い分離された 88 株のインフルエンザウイルスを対象とした。ウイル ス分離にはMDCK,LLC-MK2,Hep-2,HeLa,RD-18s,Caco2 及び Vero 細胞を用いて常法 に従って細胞培養を行った。分離したウイルス株の型別同定は国立感染症研究所インフルエン ザウイルス研究センターから配布された「2013/2014 シーズン用インフルエンザウイルス同定 キット」を用いてインフルエンザ診断マニュアル 1)に準拠し,モルモット血球を使用して赤血 球凝集阻止試験(HI 試験)により実施した。また,HI 試験が実施できなかったものについては リアルタイムPCR 法(インフルエンザ診断マニュアル準拠)により同定を行った。3.結果
2013/2014 シーズンに当所で分離されたインフルエンザウイルス 88 株のうち AH1pdm09- 13 - は51 株(58.0%),AH3 亜型は 19 株(21.6%),B 型山形系統は 17 株(19.3%),B 型ビクトリ ア系統は1 株(1.1%)であった。また,亜型別の推移は 2013 年第 43 週から第 51 週までは AH3 亜型のみが分離されていたが第52 週には AH3 亜型に加え,AH1pdm09,B 型山形系統も分離 された。それ以降はAH1pdm09 の分離数が増加し主流となったが,AH3 亜型,B 型も数は少 ないものの,流行終期まで継続して分離された。 分離された 88 株のうち,85 株については HI 試験により同定が可能であったが,3 株 (AH1pdm09 1 株,AH3 亜型 2 株)については HI 試験に必要な HA 価が得られず,リアルタイ ムPCR により同定を行った。HI 試験を実施した 85 株のうち,AH1pdm09 50 株および B 型 18 株については,すべてワクチン株類似株(HI 試験で 2 管差以内)であったが,AH3 亜型 17 株のうち3 株はワクチン株に比べ HI 価が大きく低下したもの(HI 試験で 4 管差)であった。 図1:インフルエンザウイルス分離・検出報告数の割合(2013/14 シーズン:新潟市) 図2:検体採取週別インフルエンザウイルス分離・検出報告数(2013/14 シーズン:新潟市)
4.考察
新潟市でのインフルエンザ定点あたり報告数の推移は2014 年第 2 週(2014 年 1 月 6 日から 12 日)に定点あたり報告数が 1.00 を上回り,2014 年第 9 週(2014 年 2 月 24 日から 3 月 2 日) に 60.85 となり,今シーズンのピークを迎えた。全国での推移 2)は2013 年第 51 週(2013 年 12 月 16 日から 22 日)に流行開始の指標である 1.00 を上回り,2014 年第 5 週(2014 年 1 月 27 日から2 月 2 日)にピークを迎え,その後緩やかに減少しており,流行のピークの大きさや時期 に若干の違いはあるものの,流行の開始時期および終息時期については当市との大きな違いは なかった。 58.0% 21.6% 19.3% 1.1% AH1pdm09 AH3 B/Yamagata B/Victoria 0 2 4 6 8 10 12 分 離 ・ 検 出 報 告 数 ( 株 数 ) 検体採取週 B/Victoria B/Yamagata AH3 AH1pdm09- 14 - 図3:インフルエンザウイルス定点患者報告数(2013/14 シーズン) 今シーズンのインフルエンザウイルスの分離状況は2013 年第 52 週までは AH3 亜型が主流 であったが,2014 年第 1 週以降は AH1pdm09 が主流となった。B 型は山形系統,ビクトリア 系統ともに分離されたが,大半が山形系統であった。このことは全国的な傾向 3)と大きな違い はなかった。 分離したインフルエンザウイルスのHI 試験の結果では,AH1pdm および B 型はすべてワク チン株類似株であったが,AH3 亜型では流行後期(2014 年第 11 週,第 12 週,第 14 週)にワク チン株に比べHI 価が大きく低下した株が検出された。AH3 亜型ウイルスについてはヒトでの 流行ウイルスを孵化鶏卵で分離増殖させるとアミノ酸の置換が生じ(卵馴化),抗原性が MDCK 細胞で分離したものに比べ著しく異なる傾向を示すといわれている 4)。今シーズンのワクチン 株に対して作製したフェレット感染抗血清は今シーズンにMDCK 細胞で分離された 74%の国 内流行株に対して大きく反応性が低下しているとの報告5)もあり,当所で分離したAH3 亜型も このことによる影響を受けている可能性もあるが,今回の検査結果だけでは判断できない。ま た,2014/15 シーズンのワクチンについては AH3 亜型のウイルス株の変更が公表されており, 来シーズンの動向にも注意が必要である。 インフルエンザウイルスの分離培養・検出はインフルエンザの流行状況の把握やワクチン株 の選定等,公衆衛生上重要な役割を担っており,今後も継続して実施する必要があると考える。
謝辞
検査材料の採取にご協力いただきました病原体定点医療機関の方々に深謝いたします。文献
1)国立感染症研究所:病原体検出マニュアル,インフルエンザ診断マニュアル(第2 版) 2)厚生労働省健康局結核感染症課・国立感染症研究所:今冬のインフルエンザの発生動向 (2013/14 シーズン) 3 ) 国 立 感 染 症 研 究 所 :IASR イ ン フ ル エ ン ザ ウ イ ル ス 分 離 ・ 検 出 速 報 , http://www.nih.go.jp/niid/ja/iasr-inf.html 4)国立感染症研究所:平成25 年度インフルエンザ HA ワクチン製造株の検討について(回答) , 感染研第232 号 5)国立感染症研究所:平成26 年度インフルエンザ HA ワクチン製造株の検討について(回答) , 感染研第626 号 0 10 20 30 40 50 60 70 患 者 報 告 数 ( 人 ) 診断週 新潟市 新潟県 全国- 15 - 調査研究報告(5)
新潟市における 2012/13 および 2013/14 シーズンのインフルエンザウイ
ルス薬剤耐性遺伝子について
山本 一成
概 要
新潟市における 2012/13(2013 年 4 月から 8 月)および 2013/14(2013 年 9 月から 2014 年 3 月)シーズンの流行株は,AH3N2 亜型インフルエンザウイルス(A/H3N2 亜型),A(H1N1)pdm09 亜型インフルエンザウイルス(A(H1N1)pdm09 亜型),B 型インフルエンザウイルス(B 型)の Yamagata 系統および Victoria 系統であった。2012/13 シーズンは,A/H3N2 亜型が主流であっ た。2013/14 シーズンは,A/H3N2 亜型から A(H1N1)pdm09 亜型および B 型の Yamagata 系統の混 合へと遷移した。A/H3N2 亜型(36 検体),A(H1N1)pdm09 亜型(46 検体),Yamagata 系統(27 検体)および Victoria 系統(2 検体)について薬剤耐性遺伝子の変異を調査したところ,2013 年第 7 週の A(H1N1)pdm09 亜型 1 件でアミノ酸配列解析において H275H/Y が見つかったため,国立感染症研究所で薬剤感 受性試験を実施したところ,オセルタミビル・べラミビルの両薬剤に対し感受性の低下が確認 された。
はじめに
インフルエンザは冬季に発生し,4 月~5 月に減少する冬期流行パターンを示す疾患である。 インフルエンザの原因となるインフルエンザウイルスは A 型,B 型,C 型があり,ヒトのイン フルエンザの原因となるウイルスは A 型と B 型が主である。A 型,B 型インフルエンザウイル スは人畜共通感染で突然変異が起こりやすく,ヒトではウイルスに対する免疫の持続も短く, 毎年流行を繰り返している。過去に不連続変異により,スペインかぜ(A/H1N1 亜型),アジア かぜ(A/H2N2 亜型),香港かぜ(A/H3N2 亜型),ソ連かぜ(A/H1N1 亜型),新型インフルエ ンザ(A(H1N1)pdm09 亜型)などが出現しパンデミックを起こしてきた。 インフルエンザは,5 類感染症(感染症の予防及び感染症の患者に対する医療に関する法律) に分類されており,感染症法に基づく感染症発生動向調査では,全国約 5,000 のインフルエン ザ定点医療機関(小児科 3,000,内科 2,000)から,インフルエンザと診断された患者数の情報 が報告されている。また,地方衛生研究所では,インフルエンザウイルス遺伝子検出やウイル ス分離を実施し,結果は中央感染症情報センターで集約され,インフルエンザウイルスの流行 予測および次年度ワクチン株選定に活用されている。また,地方衛生研究所と国立感染症研究 所では,抗インフルエンザ薬剤耐性株サーベイランスを実施している。 2012/13(2013 年 4 月から 8 月)および 2013/14(2013 年 9 月から 2014 年 3 月)シーズンの 新潟市におけるインフルエンザウイルスの薬剤耐性遺伝子の変異について調査したので結果を 報告する。材料と方法
1)材料
2013 年 4 月から 2014 年 3 月までに,病原体定点医療機関などでインフルエンザあるいはイ ンフルエンザ様疾患の患者より採取された咽頭ぬぐい液,鼻腔ぬぐい液,気管支吸引痰および 気管支肺胞洗浄液検体(111 検体)を材料とした。2)方法
A(H1N1)pdm09 亜型および B 型の薬剤耐性遺伝子の変異検出は,インフルエンザ診断マニュア- 16 -
ル(第 2 版)1)に準じて実施した。また,A/H3N2 亜型については,長島ら2)3)を参考に実施し
た。
シークエンス反応は,BigDye Terminator v3.1
Cycle Sequencing Kit(Applied Biosystems)を用い,Applied Biosysytems 3500 ジェネティッ クアナライザーを用いて塩基配列を決定した。
得られた塩基配列を基に遺伝子解析ソフト MEGA5 を用いて系統樹解析およびアミノ酸配列解 析を実施した。インフルエンザ診断マニュアル(第 2 版)1),Monto ら4)および Sheu ら5)を参
考に,A(H1N1)pdm09 亜型については,H275Y の薬剤耐性遺伝子の変異を,A/H3N2 亜型につい ては,E119V,D151E,D151V,R152K,R224K,Q226H,G248R,K249E,H274Y,E276D,R292K に ついて薬剤耐性遺伝子の変異の有無について確認した。また,B 型については,E119D,E119G, E119A,E119V,R152K,D198E,I222T について薬剤耐性遺伝子の変異の有無について確認した。
結果
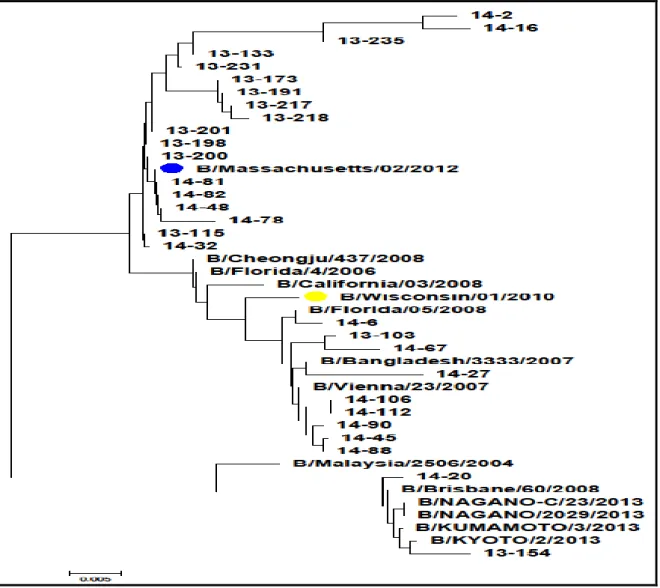
薬剤耐性遺伝子の変異を調査したところ,A(H1N1)pdm09 亜型で 46 検体中 1 検体(2.17%)に H275H/Y のアミノ酸の変異がみられた。A/H3N2 亜型 36 検体に E119V,D151E,D151V,R152K, R224K,Q226H,G248R,K249E,H274Y,E276D,R292K の薬剤耐性遺伝子の変異はみられなかっ た。また,B 型についても E119D,E119G,E119A,E119V,R152K,D198E,I222T の薬剤耐性遺 伝子の変異はみられなかった。 A(H1N1)pdm09 亜型,A/H3N2 亜型および B 型の NA 遺伝子系統樹を図 1,図 2,図 3 に示す(● は,2012/13 および 2013/14 シーズンのワクチン株,●は,2012/13 シーズンのワクチン株,● は,2013/14 シーズンのワクチン株)。
考察
新潟市の流行は,2012/13 及び 2013/14 シーズンの両期とも開始当初,A 型インフルエンザウ イルスの主流は A/H3N2 亜型であった。しかし,20013/14 シーズンは A(H1N1)pdm09 亜型と B 型の混合へと遷移した。この傾向は国内の流行状況と一致している。また,B 型インフルエン ザウイルスの国内流行状況は,Yamagata 系統と Victoria 系統の比率は 2:1 と言われているが6), 新潟市では,圧倒的に Yamagata 系統が多かった。 A/H3N2 亜型および B 型ともアミノ酸配列解析より薬剤耐性遺伝子の変異はみられなかった。 例年,培養上清を用いて実施すると 151 位のアスパラギン酸(D)からアスパラギン酸とグル タミン酸(G)の D151G/D 等の混合パターンがみられる検体がある。D151G/D や D151N 変異 は細胞培養中にも起こると言われているが,今回は臨床検体にて実施したことにより混合パタ ーンはみられなかった。その反面,泳動時に非特異バンドがみられる検体が多く,ゲル切出し による抽出検体数が増えた。これに対しては,DNase 処理を加えることにより改善されるか検 討する余地があると思われる。 2014 年 4 月 28 日現在,全国的に 4.5%の割合で H275H と H275H/Y 株が報告されている。当 所においては,1 検体(図 2 中 14-51)に H275H/Y のヒスチジン(H)とチロシン(Y)のアミ ノ酸の混合パターンがみられ 2.2%の割合であり,全国と同じ傾向であった。H275H/Y は,治 療中および治療後の患者からみられることが有り,今回の事例もべラミビル投与後の検体であ った。NA 遺伝子の系統樹解析により今シーズン,札幌市等でみられている H257Y 株とは違う クラスター群を示した。また,国立感染症研究所で薬剤感受性試験を実施し,オセルタミビル・ べラミビルの両薬剤に対し感受性の低下が確認された。 日本は、世界最大の抗インフルエンザ薬使用国である。薬剤耐性遺伝子の変異を迅速に把握 することは,公衆衛生上重要でありワクチンや薬剤の有効性などの動向を探る上でも,保健所 感染症対策室と連携しながら調査をしていく必要がある。- 17 -
謝辞
検査材料の採取にご協力いただきました各医療機関および施設等の方々に深謝いたします。文献
1)国立感染症研究所:病原体検出マニュアル,インフルエンザ診断マニュアル(第 2 版) 平成 24 年 3 月. 2)長島真美,他:東京都健康安全研究センター,研究年報,60,61-66,2009. 3)長島真美,他:東京都健康安全研究センター,研究年報,62,57-63,2011.4)Monto,A.S,McKimm-Berschkin,J.L.,et al.:Antimicrobial Agents and Chemotherapy., 50,2395-2402,2006.
5)Sheu,T.G.,Deyde,V.M.,et al.:Antimicrobial Agents and Chemotherapy.,52,3284-3292,2008. 6)国立感染症研究所:今冬のインフルエンザの発生動向 (2013/14 シーズン) http://www.nih.go.jp/niid/images/idsc/disease/influ/fludoco1314.pdf 7)国立感染症研究所:抗インフルエンザ薬剤耐性サーベイランス 2014 年 4 月 28 日 http://www.nih.go.jp/niid/ja/influ-resist/4604-flu-dr20140428.html
図 1 A/H3N2 亜型 NA 遺伝子の分子系統樹
- 18 -
図 2 A(H1N1)pdm09 亜型 NA 遺伝子の分子系統樹
- 19 -
図 3 B 型 NA 遺伝子の分子系統樹
- 20 - 調査研究報告(6)
下水における下痢症ウイルス遺伝子の検出状況について
(第 2 報)
桃井拓也 齋藤哲也 足立玲子 南波裕太概 要
当所では平成24 年度より,下水における下痢症ウイルスの遺伝子を調査している。 今年度は,昨年度と異なり1年を通してノロウイルスGⅠ,サポウイルスが検出された。 下水を調査した結果,市内においてノロウイルス及びサポウイルスは発症者が増加する冬季 だけではなく,1年を通して注意しなくてはいけない病原体であることがわかった。 下水を調査することは市内における流行状況の把握や地域に存在しているウイルスを継続的 に監視することができ,流行や季節変動を把握するためには非常に有用な検体であるといえる。1.はじめに
嘔吐や下痢を主症状とする急性胃腸炎の原因として,細菌,ウイルス,原虫などがあり, 今日の傾向としてウイルス性による症例報告が多い1)。食中毒事例や集団感染事例の原因と されるノロウイルスなどの下痢症ウイルスはその事例に係る吐物や便と共に下水処理場へ 流れ込む。平成25 年における新潟市内の下水道普及率は 82.8% (http://www.city.niigata.lg.jp/kurashi/jyogesuido/gesui/ keikaku/fukyuu.html)であり, この処理場に入り込んでくる汚水から感染症のリスク評価および市中に循環する下痢症ウ イルスが把握できる可能性がある。 今回,感染症発生動向調査における感染性胃腸炎患者の定点あたりの報告数と下水中の 下痢症ウイルスの遺伝子検出状況の関連性について検討した。また,下水を用いた感染症 のリスク評価を行った。2.材料と方法
1) 下水の採取 2013 年 4 月から 2014 年 2 月の間,18 週( 4 月) 22 週( 5 月) 26 週( 6 月) 31 週( 7 月) 35 週( 8 月) 39 週( 9 月) 43 週( 10 月) 48 週( 11 月) 52 週( 12 月) 4 週( 1 月) 8 週( 2 月) において市内の下水処理施設(計画処理人口:約 275,000 人,処理方法:標準活性汚泥 法)から約 2L の流入水を採取した。 2) 検査項目 ノロウイルス,サポウイルス,アストロウイルス,A 群ロタウイルス,C 群ロタウイル ス,アデノウイルス,エンテロウイルス,大腸菌群数,大腸菌数 3) ウイルスの濃縮方法 3,000rpm×30 分遠心後,上清約 1L に対し,ポリエチレングリコール 6000 を 10%, 1MNaCl を添加し(以下,PEG 濃縮),4℃で一昼夜保管後,10,000rpm×30 分遠心した。 その沈査に滅菌蒸留水2ml を加え,12,000rpm×20 分遠心後上清に 30%ショ糖溶液を重 層し,40,000rpm×120 分遠心した。その沈査を滅菌蒸留水 400μl で溶解し濃縮検体とし た。- 21 - 4) ウイルスの検出方法
3)の検体から RNA 抽出は QIAamp Vial RNA Mini Kit(QIAGEN)を使用し,DNaseⅠ処 理を行った。ノロウイルス,サポウイルス,アストロウイルスおよびエンテロウイルスの 検体はrandom primer を用いて逆転写し,Super Script Ⅱ(invitrogen)を用い cDNA を作 製した。 A 群ロタウイルスおよび C 群ロタウイルスについては,DNaseⅠで処理した後,ジメチ ルスルホキシド(和光)を 10%添加し,97 ℃ 5 分後急冷したものに,SuperScriptⅡ (invitrogen)を用い,cDNA を作製した。 アデノウイルスについては,RNA 抽出液を 98℃10 分処理したものを使用した。 各ウイルスの検出方法は病原体検査マニュアルおよび通知等に準じ実施した。 5) 遺伝子解析
検出した増幅産物は,シーケンサーABI 3500 Genetic Analyzer(Applied Biosystems)を 用い塩基配列の解析を実施した。
3.結果
2013 年 18 週から 2014 年 8 週に採取した下水 11 検体のウイルス検索の結果を表 1 に示 した。 ノロウイルス遺伝子については,GⅠ遺伝子・GⅡ遺伝子ともに採取した週全てに検出さ れた。検出されたウイルス量はGⅠでは最大 1.6×103 [copies/5μl],最小 4.0 [copies/5μl] で,GⅡでは最大 2.2×104 [copies/5μl] ,最小 1.1×102 [copies/5μl]であった。 ノロウイルス遺伝子の検出状況と感染症発生動向調査に係る感染性胃腸炎の定点報告数 との関連は,ノロウイルスGⅡ遺伝子のウイルス量が増加した 52 週にかけて,その報告数 に増加傾向が認められた。(図1)。 また,検出されたノロウイルスの遺伝子塩基配列を解析した結果,各週において 1 つず つの遺伝子型がみられ,複数の型を認めた週はなかった。GⅠでは GⅠ.1 , GⅠ.3 , GⅠ.4 及 びGⅠ.6 が検出され,その中でも GⅠ.4 及び GⅠ.6 の遺伝子型が多く検出された。一方 G ⅡではGⅡ.2 , GⅡ.3 及び GⅡ.4 が検出され,9 検体が GⅡ.4 であった。この GⅡ.4 の塩基 配列を解析した結果,48 週は変異株 Hu/GII.4/New Orleans1805/2009/USA に近縁で,22 週 , 35 週 , 52 週 , 4 週及び 8 週は変異株 Hu/GII.4/Sydney/NSW0514/2012/AU に近縁で あった。 2 月及び 3 月に発生した食中毒事例で検出されたノロウイルスの塩基配列と 8 週に採取し た下水から検出されたノロウイルスの塩基配列との相同性を比較した結果,8 週に採取した 下水と食中毒事例で検出されたものの塩基配列は一致した(図2)。 サポウイルスは採取した週全てで検出された。また検出された遺伝子型はGⅠ.1 , GⅠ.2 であった。 アストロウイルスは22 週 , 26 週及び 31 週で検出された。また検出された遺伝子型はⅠ- 22 - 及びⅡ型であった。 A 群ロタウイルスは 18 週 , 43 週 , 48 週 及び翌年の 4 週 , 8 週で検出された。また検出 された遺伝子型はG1 , G3 及び G4 であった。 アデノウイルスは22 週 , 26 週 , 35 週 , 39 週 , 43 週及び翌年の 4 週 , 8 週で検出され た。また検出された遺伝子型は31 型及び 41 型であった。 C 群ロタウイルス及びエンテロウイルスは検出されなかった。 大腸菌群数および大腸菌数を検査した結果,大腸菌数は全ての検体で 1.5×103 [cfu/ml] から8.6×104 [cfu/ml],大腸菌群数は全ての検体で 2.0×104 [cfu/ml] から 2.0×105 [cfu/ml] の値であった。
4.考察
2013/14 シーズンにおいて下水から下痢症ウイルス検索を試み,2012/13 シーズンと同様 に全ての週でウイルス遺伝子を検出した。ノロウイルスについて,下水から検出した copy 数は,GⅠと GⅡとを比較すると,GⅡの方がより多かった。このことは GⅠより GⅡが人 の腸管で増殖しやすい及び接触の可能性が高いと考えられる。また,その下水から検出さ れたノロウイルス GⅡのウイルス量が増加すると,医療機関からの感染性胃腸炎患者数の 報告が多くなる傾向が見られた。このことは,集団事例や多くの散発事例が発生したこと によるものと思われる。 G Ⅱ の 遺 伝 子 型 は G Ⅱ .4 が 多 く 検 出 さ れ 、 塩 基 配 列 を 解 析 し た 結 果 、 多 く が Hu/GII.4/Sydney/NSW0514/2012/AU に近縁であった。冬季だけでなく 2013 年 22 週や 2013 年 35 週でも検出されたことから,市内にノロウイルスの変異株が 1 年を通して存在 していることが考えられる。市内に存在しているGⅡ.4 の株は 2013 年 52 週から流行を引 き起こしていた可能性が高く、2014 年 8 週に検出した塩基配列はその後に発生した 2 件の 食中毒事例の塩基配列と相同性が認められた(図 2)。 サポウイルスは昨年度と異なり,1 年を通して検出された。遺伝子型は GⅠ.1 及び GⅠ.2 が検出されている。したがって,市内にはサポウイルスが 1 年を通して存在していること が考えられる。特に GⅠ.2 は既に国内や海外で食中毒を引き起こす遺伝子型として報告さ れている2),3)。このことからノロウイルスが検出されない感染事例が発生した場合,サポウ イルスを疑う必要があると考えられる4)。 アストロウイルスは2013 年 22 週から 31 週まで検出されたことから,この時期に市内で 散発的に発生したと考えられる。 検出されたA 群ロタウイルスの遺伝子型は人から検出される遺伝子型の一つ 5)であるた め,この時期に市内でロタウイルスによる感染性胃腸炎の発生が多くあったことが考えら れる。一般的にロタウイルスは春季に流行するウイルスだが,43 週 , 48 週に検出されたこ とから春季以外の季節においても十分に考慮しなくてはいけない。 大腸菌群数および大腸菌数を検査した結果,大腸菌群数は採取した全ての週で 2.0×104- 23 - [cfu/ml] から 2.0×105 [cfu/ml] の値であった。このことから雨水は下水中のウイルスの濃 度には大きく影響しないと思われる。 このように,下水中から下痢症ウイルスの遺伝子の検出を調査することは市内における 感染症事例の発生の予測できる。今後とも下水を調査し、感染症対策室と情報を共有する ことで感染症予防に寄与していきたい。
5.文献
1) 厚生労働省医薬食品局食品安全部監視安全課長通知:ノロウイルスによる食中毒の発生 予防について.平成 25 年 1 月 11 日,食安監発 0111 第 2 号(2013) 2) 小林 慎一 他: 給食弁当を原因としたサポウイルスによる大規模食中毒事例―愛知県, 病原微生物検出情報, 31(11)322-323,2010 3) 岡 智一郎 他: 愛知県と川崎市の食中毒事例から検出されたサポウイルス GI/2 の塩基 配列の比較,病原微生物検出情報, 31(11)324-325,2010 4) 山﨑 俊治 他:ノロウイルスが検出されない下痢症患者におけるアストロウイルス,サ ポウイルス,アイチウイルスの調査(第 2 報).平成 16 年度福岡市保健環境研究所報 5) 小林 宣道:全遺伝子配列に基づくヒトロタウイルスの遺伝子型別とその世界的な動向. 病原微生物検出情,35(3)66-67,2014 18週 22週 26週 31週 35週 39週 43週 48週 52週 4週 8週 NV GⅠ ●GⅠ.6 ●GⅠ.6 ●GⅠ.6 ●GⅠ.4 ●GⅠ.3 ●型別不能 ●GⅠ.1 ●GⅠ.4 ●GⅠ.4 ●GⅠ.6 ●GⅠ.4 NV GⅡ ●GⅡ.2 ●GⅡ.4 ●GⅡ.4 ●GⅡ.4 ●GⅡ.4 ●GⅡ.3 ●GⅡ.4 ●GⅡ.4 ●GⅡ.4 ●GⅡ.4 ●GⅡ.4 Sapo ●GⅠ.2 ●GⅠ.2 ●GⅠ.2 ●GⅠ.1 ●GⅠ.2 ●GⅠ.1 ●GⅠ.1 ●GⅠ.1 ●GⅠ.1 ●GⅠ.1 ●GⅠ.1 Astro ●Ⅰ ●Ⅰ ●Ⅱ A rota ●G1 ●G3 ●G4 ●G3 ●G1 C rotaAdeno ●type31 ●type41 ●type41 ●type41 ●type41 ●type41 ●type41 Entero
2013 2014
- 24 - FD_2014.2 ATGAAGATGGCGTCGAGTGACGCCAACCCATCTGATGGGTCCGCAGCCAACCTCGTCCCA FD_2014.3 ATGAAGATGGCGTCGAGTGACGCCAACCCATCTGATGGGTCCGCAGCCAACCTCGTCCCA sewage_2014.2 ATGAAGATGGCGTCGAGTGACGCCAACCCATCTGATGGGTCCGCAGCCAACCTCGTCCCA ************************************************************ FD_2014.2 GAGGTCAACAATGAGGTTATGGCTCTGGAGCCCGTTGTTGGTGCCGCCATTGCGGCACCT FD_2014.3 GAGGTCAACAATGAGGTTATGGCTCTGGAGCCCGTTGTTGGTGCCGCCATTGCGGCACCT sewage_2014.2 GAGGTCAACAATGAGGTTATGGCTCTGGAGCCCGTTGTTGGTGCCGCCATTGCGGCACCT ************************************************************ FD_2014.2 GTAGCGGGCCAACAAAATGTAATTGACCCCTGGATTAGAAATAATTTTGTACAAGCCCCT FD_2014.3 GTAGCGGGCCAACAAAATGTAATTGACCCCTGGATTAGAAATAATTTTGTACAAGCCCCT sewage_2014.2 GTAGCGGGCCAACAAAATGTAATTGACCCCTGGATTAGAAATAATTTTGTACAAGCCCCT ************************************************************ FD:食中毒事例株 sewage:下水由来株 図2 下水由来株及び食中毒事例株の塩基配列の相同性 0 5 10 15 20 25 30 35 40 0 5000 10000 15000 20000 25000 30000 35000 40000 18週 22週 26週 31週 35週 39週 43週 48週 52週 4週 8週 2013 2014 [copies/5μl] 図1 ノロウイルスcopy数と定点報告数 定点報告数 NoV GⅠ NoV GⅡ [人]
- 25 - 調査研究報告(7)
市販二枚貝からの下痢症ウイルス遺伝子の検出状況について
(第 2 報)
南波裕太 桃井拓也 齊藤哲也 足立玲子概 要
平成24 年より市販二枚貝について下痢症ウイルスおよび A 型肝炎ウイルス(HAV)を 調査しており,平成25 年の夏季については,下痢症ウイルス遺伝子および A 型肝炎ウイル ス遺伝子の検出はなかったが,冬季にはノロウイルス(NV),サポウイルス(SaV)および アイチウイルス(AiV)を検出した。また,それぞれのウイルスの検出率は NV35.9%, SaV2.6%および AiV28.2%であった。1.はじめに
NV・SaV・アストロウイルス(AstV)などの下痢症ウイルス1)は,人の体内に侵入する と急性胃腸炎を引き起こす。特にNV はカリシウイルス科に属し,冬季に多発するウイル ス性食中毒の主な原因のウイルスである2)。ウイルスゲノムを比較することにより, GenogroupⅠ(GⅠ)と GenogroupⅡ(GⅡ)の遺伝子群に分類され3),4),さらにそれぞれ に多くの遺伝子型が存在する5)。また,NV 同様カリシウイルス科に属する SaV を原因と する胃腸炎が増加しており,特に乳幼児に多く認められる。SaV には GenogroupⅠ~Ⅴが あり,人に感染が確認されているのは,GenogroupⅠ,Ⅱ,Ⅳ,Ⅴである。 一方,HAV はピコルナウイルス科に属し,経口感染により発熱や下痢を伴った急性肝炎 を発症させる6)。 NV や HAV は食中毒の原因物質に指定されており,ウイルス性食中毒において注意しな ければいけないウイルスである。また,二枚貝はその中腸線にウイルスが蓄積されるとい う性質上,ウイルス性食中毒の原因となり易い7)。 今回,市販二枚貝(生カキ・アサリ・シジミ)について,NV などの下痢症を引き起こす ウイルス,HAV およびエンテロウイルス(EnV)の汚染実態を調査したので,報告する。2.材料と方法
1)検査項目NV GⅠ・GⅡ,SaV,AstV,A 群・C 群ロタウイルス(RotaV groupA・groupC),アデ ノウイルス(AdV),AiV,HAV,EnV
2)材料
- 26 - を検査材料とし,生カキについては3 個の中腸線,アサリ・シジミについては 5 個の中腸 線を1 検体とし,合計で 39 検体の二枚貝(生食用生カキ 8 検体・加熱調理用生カキ 17 検 体・アサリ10 検体・シジミ 4 検体)を調査した。また,生カキは冬季,アサリは 6 月及び 1 月,シジミは 9 月に採取し調査した。 PBS(-) 45 mL で乳剤を作成し,10,000 rpm × 20 min 冷却遠心を行い,上清にポリエ チレングリコール(Mw = 6,000)を 10%, NaCl を濃度が 1M になるように添加し,4℃で一 昼夜静置した。その後,10,000 rpm × 20 min 冷却遠心を行い,上清を捨て,沈渣を滅菌 蒸留水2 mL に浮遊させ,30%ショ糖に重層させた後,40,000 rpm × 120 min 冷却遠心 を行った。上清を捨て,沈渣をPBS(-)で洗浄した後,滅菌蒸留水 200 L 中に浮遊させ検体 とした。 3)方法 平成24 年同様に,NV については厚生労働省通知8)に準じて行った。他の下痢症ウイル スおよびHAV については厚生労働省通知9)および国立感染症研究所のウイルス性下痢症診 断マニュアル第3 版に準じて行った。 陽性となったPCR 産物の遺伝子解析は,各種プライマー10)を用いて遺伝子領域の一部を 増幅し,得られた増幅産物をQIAquick PCR Purfication Kit (QIAGEN)により精製し, BigDye® Terminator v3.1 Cycle Sequencing Kit (Applied Biosystems)を用い,
3500GeneticAnalyzer (Applied Biosystems)でダイレクトシーケンス法により塩基配列を 決定し,遺伝子解析ソフト MEGA5 を用いて系統樹解析を行った。
3.結果
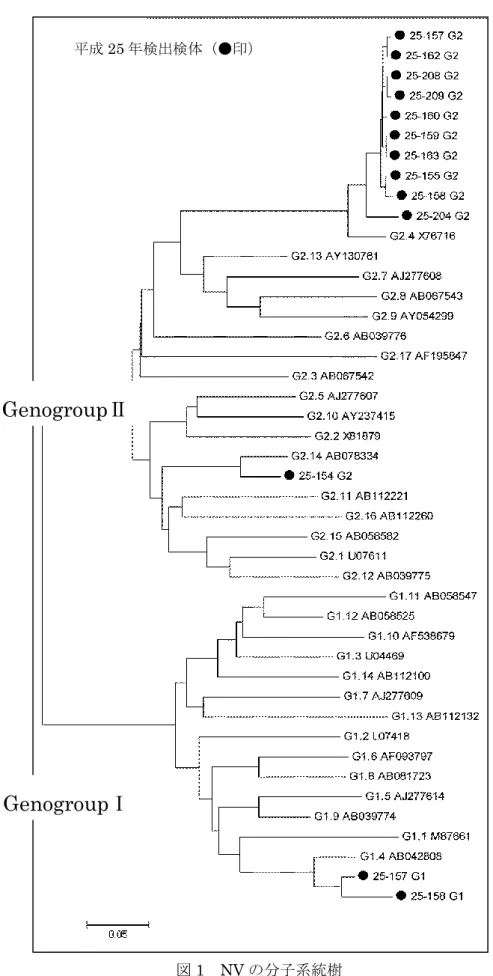
市販二枚貝について下痢症ウイルス遺伝子及びHAV 遺伝子を調査した結果,生カキ 3 検 体からNV GⅠ遺伝子,生カキ 14 検体から NV GⅡ遺伝子,生カキ 11 検体から AiV 遺伝 子,生カキ1 検体から SaV 遺伝子が検出された。また,これらのウイルス遺伝子はある 1 検体から重複して検出される場合もあった。ウイルス遺伝子の検出パターン及び陽性数を 表1 に示す。 表1 二枚貝における各種ウイルス遺伝子の検出状況 生カキにおいてAstV,RotaV(groupA 及び C),HAV,EnV については検出されなかっ た。また,アサリ・シジミからは下痢症ウイルス遺伝子およびHAV 遺伝子は検出されなか 検出パターン 陽性率[%] NV GⅡ 12.8 NV GⅠ/GⅡ 2.6 NV GⅡ,AiV 15.4 NV GⅠ/GⅡ,AiV 2.6 NV GⅠ/GⅡ,AiV,SaV 2.6 AiV 7.7 陽性数/検体数 5/39 3/39 1/39 1/39 1/39 6/39- 27 - った。 今回調査した生カキについて生食用および加熱調理用の陽性数を表2 に示す。また,今 回検出されたウイルス遺伝子および平成24 年に調査した結果を表 3 に示す。 表2 生カキにおける各種ウイルス遺伝子の検出状況 表3 平成 24 年及び 25 年の検出状況 NV GⅠ及び NV GⅡ(計 17 件)のうち塩基配列を決定できた 13 件について系統樹解析 した結果,GⅠ.4(2 件),GⅡ.4(10 件),GⅡ.14(1 件)に分類された(図 1)。
AiV11 件について塩基配列を決定し系統樹解析した結果,11 件すべてを AiV genotypeA に分類された(図2)。 SaV1 件について塩基配列を決定し系統樹解析した結果,SaV GⅠ.2 に分類された(図 3)。
4.考察
今回の調査でウイルス遺伝子が検出された検体は生食用または加熱調理用の生カキのみ で,アサリ・シジミからの検出はみられなかった。 流通している生カキには生食用と加熱調理用の区別があり,生食用においては食中毒を 防止するために成分規格などの規格基準が定められているが,ウイルスに関する基準は定 められていない。そのためウイルスによる食中毒が広く認識された現在,生産者は出荷前 の浄化洗浄や自主検査などによりウイルスに汚染されている生カキの市場への提供を自粛 している。しかし,今回の調査ではNV の陽性率は生食用カキで 62.5%,加熱調理用カキ で53%と決して低いとは言えない数値となった。また,検出された各ウイルス遺伝子にお いて平成24 年に調査した結果よりも高い陽性率を示した。今回の結果から平成 25 年はカ キの採取海域における糞便汚染が示唆されたが,平成24 年・25 年に調査した生カキは無作 為に採取し,産地における偏りはなく,検査数も大差はなかった。また,平成24 年・25 NV GⅡ 37.5 11.8 NV GⅠ/GⅡ 12.5 0 NV GⅡ,AiV 0 35.3 NV GⅠ/GⅡ,AiV 12.5 0 NV GⅠ/GⅡ,AiV,SaV 0 5.9 AiV 0/81/8 12.5 1/172/17 11.8 1/8 0/17 0/8 6/17 1/8 0/17 生食用 陽性数/検体数 陽性率[%] 加熱調理用 陽性数/検体数 陽性率[%] 3/8 2/17 検出パターン 平成24年 平成25年 NV GⅠ 0/42(0) 3/39(7.7) NV GⅡ 1/42(2.4) 14/39(35.9) SaV 0/42(0) 1/39(2.6) AiV 2/42(4.8) 11/39(28.2) ウイルス 陽性数/検体数(陽性率[%])- 28 - 年における全国的なNV の検出状況においても大きな違いはみられなかった。今後,カキ の採取海域などを限定した調査や継続的なデータの収集が望まれる。 生カキからはNV 以外にも SaV,AiV が検出されており,AiV は胃腸炎の原因ウイルス で,1989 年の愛知県で集団発生した胃腸炎患者からはじめて分離されたウイルスである11)。 遺伝子型はA,B,C の 3 種類があり12)13),今回の調査では検出したすべてのAiV が genotypeA に分類された。AiV は他の下痢症ウイルスと比較して検出頻度が低く,病原性 を疑問視されている。また,WHO は腸管内に感染を起こすウイルスとして分類したが,食 品媒介ウイルス感染症としての重要性は低いと評価している。しかしながら,AiV が多く の場合がNV と同時に検出されていることや NV 等と同様に腸管内に感染するウイルスで あることから,二枚貝の生育海域の糞便汚染調査における一つの指標となり得ると考えら れる。 また,近年SaV を原因とする食中毒が増加しており,SaV は NV と同様に糞便中に排泄 されるため,環境や食品の汚染が危惧される。今回の調査においても1 検体から検出して おり、SaV についても注意が必要である。 今回の調査では,生食用・加熱調理用生カキからNV,AiV,SaV を検出した。これらの 中には3 種のウイルスが重複して検出された検体もあり,このカキが採取された海域が下 痢症ウイルスによって汚染されていた可能性を示唆した。また,これらの結果を行政を通 じて広く還元することで市内におけるカキを原因とする食中毒防止に寄与することができ ると思われる。
- 29 -
図1 NV の分子系統樹
GenogroupⅠ
GenogroupⅡ
- 30 - 図2 AiV の分子系統樹 図3 SaV の分子系統樹
genotype A
genotype B
Bovine Kobuvirus
genotype C
平成25 年検出検体(●印) 平成25 年検出検体(●印)- 31 -
文献
1) 牛島廣治:ヒト急性胃腸炎の起因ウイルス,総合臨床,51:2923-2927,2002 2) 国立感染症情報センター.病原微生物検出情報(IASR)2003; 31: 312-314
3) Ando T, Noel JS, Fankhauser RL. Genetic classification of “Norwalk-like viruses”. J Infect Dis 2000; 181 (Suppl 2): 336-348.
4) Katayama K, Shirato-Horikoshi H, Kojima S, Kageyama T, Oka T, Hoshino FB, et al. Phylogenetic analysis of the complete genome of 18 Norwalk-like
viruses. Virology 2002; 299: 225-239.
5) Kageyama T, Shinohara M, Uchida K, Fukushi S, Hoshino FB, Kojima S, et al. Coexistence of multiple genotypes, including newly identified genotypes, in
outbreaks of gastroenteritis due to Norovirus in Japan. J Clin Microbiol 2004; 42: 2988-2995. 6) 入谷展弘, 勢戸祥介, 春木孝祐, 西尾治, 久保英幸, 改田厚, 他.市販生カキからのノロ ウイルスおよびA 型肝炎ウイルスの検出. 生活衛生 2005; 49: 279-287. 7) 食品ウイルス性胃腸炎集団発生実態調査研究班 : 最近 5 年間の食品媒介ウイルス性胃腸 炎集団発生全国実態調査総合報告書 . 食品媒介ウイルス性胃腸炎集団発生実態調査研究班 8) 厚生労働省医薬局食品安全部監視安全課長通知:ノロウイルスの検出法について.平成 15 年11 月 5 日,食安監第 1105001 号(2003). 9) 厚生労働省医薬局食品保健部監視安全課長通知:ふん便及び食品中の A 型肝炎ウイルス の検査法について.平成 14 年 8 月 16 日, 食監発第 0816001 号(2002). 10)齊藤哲也, 足立玲子, 桃井拓也. 国産および輸入二枚貝からの下痢症ウイルス遺伝子の 検出状況について. 新潟市衛生環境研究所 年報(調査研究報告) 2013; 37: 7-9
11) Yamashita T, Kobayashi S, Sakae K, Nakata S, Chiba S, Ishihara Y, Isomura S.: Isolation of cytopathic small round viruses with BS-C-1 cells from patients with gastroenteritis. J Infect Dis 164: 954-957, 1991.
12)Yamashita T et al.: Application of a Reverse Transcription-PCR for Identification and Differentiation of Aichi Virus,aNew Member of the Picornavirus Family Associated with Gastroenteritis in Humans. J. Clin. Microbiol. 38 : 2955-2961 , 2000
13)K. Ambert-Balay et al. : Prevalence and Genetic Diversity of Aichi Virus Strains in Stool Samples from Community and Hospitalized Patients. J. Clin. Microbiol. 46 : 1252-1258 , 2008
- 32 - 調査研究報告(8)
新潟市河川における水生底生生物実態調査(第 12 報)
猪股秀子 岡田裕美 立川正幸 松田哲明概 要
当所では,平成 17 年度の水生底生生物実態調査基本計画に基づき,新潟市の河川環境につい て毎年調査を行っている。平成 21 年度までの 5 年間で,市内 13 河川,33 地点の 1 回目の調査 が終わった。平成 22 年度からは調査地点を信濃川,西川,阿賀野川の 3 河川にしぼり,経年 変化や河川環境をより詳しく調べることにした。今年度は阿賀野川の 4 地点について調査を実 施し,底質など基礎資料を収集するとともに,水質変化との関連等を前回(平成 22 年度)と 比較しながら,河川環境を総合的に評価した。1 はじめに
新潟市の河川について水環境を評価するため、平成 14 年度から水生底生生物を実施している。 平成 25 年度は、平成 22 年度に調査した早出川・阿賀野川の 4 地点について調べ、比較検討した ので報告する。2 調査方法
(1)調査日 平成 25 年 5 月 29 日 (2)調査地点(図 1) St1 早出川上流(五泉市水戸野) St2 早出川中流(五泉市太田善願橋) St3 早出川下流(新潟市秋葉区新郷屋) St4 阿賀野川下流(新潟市江南区小杉大阿賀橋) 図 1 調査地点 新郷屋 大阿賀橋 善願橋 水戸野 阿賀野川 早出川 日本海 St 1 St 2 St 3 St 4 信濃川- 33 - (3) 調査項目 水生底生生物を採集し,科レベルの分類を行った。また気温,水温,透視度, pH,溶存酸素,化 学的酸素要求量(以下 COD と表示),生物化学的酸素要求量(以下 BOD と表示),全窒素,全りん, クロロフィルa,電気伝導率,底質の含水量,強熱減量の理化学調査を行った。 (4) 河川環境の評価方法 水生生物による河川環境の評価はスコア法,水質階級法の 2 法を用いた。 ① スコア法 (「生物等による水環境評価マニュアル」 環境庁水質保全局より) 62 の科レベルの 指標生物に 1~10 までのスコアが設定され,きれいな川に住む生物ほど数字が大きい。地点毎に, 採集された指標生物の平均スコア値(以下 ASPT 値という)を算定し,ASPT 値が 10 に近いほど汚濁 の程度が少なく自然環境に近い人為的影響の少ない河川環境であると評価する方法。 ② 水質階級法 (「川の生き物を調べよう」 環境庁水質保全局 建設省河川局編より) 30 種類の指 標生物を,きれいな水(水質階級Ⅰ),ややきたない水(水質階級Ⅱ),きたない水(水質階級Ⅲ), とてもきたない水(水質階級Ⅳ)の 4 階級に分類し,多くの種類が採集された生物の階級で河川環 境の評価を行う方法。