九州大学学術情報リポジトリ
Kyushu University Institutional Repository
カイコ体液プロフェノールオキシダーゼに関する研 究
山本, 幸治
九州大学農学研究科遺伝子資源工学専攻
https://doi.org/10.11501/3150654
出版情報:Kyushu University, 1998, 博士(農学), 課程博士 バージョン:
権利関係:
カイコ体液プロフェノールオキシダーゼに関する研究
山 本 幸 治
1998
目次
第一章 序論
参考文献 5
第二章 プロフェノールオキシダーゼの精製法および検出法の確立
第一節 緒言
7
第二節 実験材料
7
第一項 カイコ系統および体液採取法 第二項 皮膚採取法
第一項 試薬
第四項 PPO活性化因子の調製
第三節 実験方法
8
第一項 プロフェノールオキシダーゼの活性化
(1)
PPAEを用いたPPOの活性化 (2)界面活性剤を用いたPPOの活性化(3)
2-プロパノールを用いたPPOの活性化第二項 プロフェノールオキシダーゼ活性化後のPO活性の検出
(1)
PPAEを用いたフェノールオキシダーゼ活性の検出 (2) DBMAを用いたフェノールオキシダーゼ活性の検出 第三項 タンパク質の定量第四項 純度検定(SDS-PAGE)
第五項 未変性ポリアクリルアミド電気泳動(Native-PAGE) 第六項 プロフェノールオキシダーゼの検出(活性染色法)
第四節 実験結果
第一項 プロフェノールオキシダーゼの分離、 精製 第二項 プロフェノールオキシダーゼの活性化
第三項 DBMAとPPAEによるフロフェノールオキシダーゼの 活性化の比較
第四項 電気泳動後のフロフェノールオキシダーゼの検出
11
第五節 考察
13
第六節 小括 参考文献
4 5 1i 1i
第三章 プロフェノールオキシダーゼイソフォームの分離、 精製およびその 性質
第一節 緒言
26
第二節 実験材料
第一項 カイコ系統および体液採取法 第二項 試薬
26
第三節 実験方法
第一項 プロフェノールオキシダーゼの検出(活性測定法) 第二項 分子サイズ
第三項 等電点
第四項 基質特異性(ミカエリス定数、 最大速度および触媒効率の測 定)
26
II
第四節 実験結果
定)
第一項 プロフェノールオキシターゼイソフォームの分隊、 精製 第二項 プロフェノールオキシダーゼの性質
(1)分子サイズ
(2)等電点
(3)
pH安定性および至適pH(4)基質特異性(ミカエリス定数、 最大速度および触媒効率の測
2 7
第五節 考察
30
第六節 小括
31
参考文献
33
第四章 プロフェノールオキシダーゼcDNAのクローニングおよび 塩基配列
第一節 緒言
47
第二節 実験材料
47
第一項 血球
第二項 プラスミド
第三項 菌株 第四項 培地
第五項 制限酵素および核酸修飾酵素 第六項 その他の試薬
第三節 実験方法
第一項 トータノレRNAの調製
48
第二項
Reverse transcriptase-polymerase chain reaction (RT-PCR)
第三項
Polymerase chain reaction (PCR)
第四項アガロースゲル電気泳動
第五項 PCR生成物のサブクローニング
( 1)
PCR産物の制限酵素消化( 2)アガロースゲル電気泳動によるPCR産物の精製
(3)
Vector DNAの調製(4)ライゲーション
( 5
)コンビテントセルの調製(6
)トランスフォーメション(形質転換)( 7
)プラスミドDNAの精製第六項 塩基配列の決定
( 1
)シークエンス反応( 2)ポリアクリルアミドゲル電気泳動
第四節 実験結果
第一項 プロフェノールオキシダーゼcDNAのクローニング 第二項 プロフェノールオキシダーゼの塩基配列
第三項 アミノ酸配列の比較 第五節 考察
第六節 小括 参考文献
5 4
55
56
57
第五章 プロフェノールオキシダーゼの発現系の構築 第一節 緒言
第二節 実験材料
第一項 オリゴヌクレオチド 第二項 プラスミド
第二項 菌体 第四項 酵素
第三節 実験方法 第一項 PCR法
第二項 PCR産物のクローニング
第三項 発現ベクターへのライゲーション 第四項 PPOの発現
第五項 ウエスタンプロッティング 第六項 粗組み換えタンパク質の調製
第七項 粗組み換えタンパク質のDBMAとPPAEによる活性化
66
66
67
第四節 実験結果
69
第一項 PCRによる制限酵素部位の導入とクローニング
第二項 発現ベクターpET-11bへのライゲーション 第三項 PP01およびPP02の大腸菌内での発現 第四項 組み換えタンパク質の活性化
第五節 考察
70
第六節 小括
71
参考文献
第六章 カイコ変態過程におけるPPOの変動 第一節 緒言
第二節 実験材料 第一項 体液採取法 第二項 血球採取法
第三 節 実験方 法
第一項 体液中のプロフェノールオキシダーゼの検出 第二項 ポリクローナル抗体の作製
第三項 ELISA
第四項 ノーザンハイフリ夕、イゼーション
( 1
) トータルRNAの調製( 2
)変性アガロースゲル電気泳動(3
)プロッティング(4)ジコギシゲニン標識プローブの調製
( 5
)ハイブリダイゼーション(6)
DIG標識ハイブリダイズ体の検出 第五項 PPOmRNA発現量の測定第四節 実験結果
第一項 変態過程におけるフェノールオキシダーゼ活性の変動 第二項 変態過程におけるプロフェノールオキシダーゼ量の変動 第三項 変態過程におけるプロフェノールオキシダーゼ、mRNA の変動
72
79
79
79
84
第五節 考察 第六節 小括
参考文献
第七章 総括および考察 参考文献
第八章 要約 謝辞
85
86
88
93 98
100
103
第一章 序論
昆虫の変態とそれに伴う酵素系の変動については、 多数の研究があり、 そ の歴史は古い。 そのほとんどが酵素活性の変動を調べたもので、 その分子レ ベルでの制御機構について調べているものは少ない。 その中で、 イエバエ由 来のフェノールオキシターゼ系については、 今日まで多くの生化学者によっ て研究されている。 現在までに得られている知見のうち注目すべきものとし て(1)フェノールオキシダーゼは、 変態のある時期には不活性なプロエン ザイムとして存在していること、 (2)このフロエンザイムを活性化する因 子が存在すること、 (3)この活性化にはエクジソンが関与していることな どが挙げられる。
フェノールオキシダーゼ(PO:モノフェノール、 ジヒドロキシフェニルア ラニン:オキシゲンオキシドレダクターゼ; EC1.14.18.1)は、 別名チロシナ ーゼとも呼ばれ、 モノフェノールからoージフェノールへの水酸化そしてoージ フェノールからoーキノンへの酸化というこつの連続的な反応を触媒する。 昆 虫において、 POは、 メラニン合成、 表皮の硬化そして生体防御反応などに 関与していると考えられている。
昆虫の生育過程のある時期にフェノールオキシダーゼが不活性なプロエン ザイムの形で存在していることを最初に見いだしたのは、 Bodineらであるし 2)。 彼らは、 バッタ(Melanoplus differentia1is)の卵の解化過程における チロシナーゼの活性変動を調べ、 解化中の卵のチロシナーゼがプロチロシナ ーゼとして存在していることを明らかにした。 同時に、 このプロエンザイム に対し活性化因子(natural activator)が存在し、 その本体は脂溶性物質で あることを明らかにした3)。 さらにわvitroにおけるプロチロシナーゼの活性 化剤についても種々の検討を行い、 クロロホルム、 アセトン、 エチルウレタ ン、 尿素そしてオレイン酸ナトリウムのようなタンパク質の変性剤が活性化 に有効な物質であることを見いだした4)。 なおこの他に600C10分間の熱処 理、 激しい振塗によっても同様に活性化されることを見いだし5)、 活性化は
1
プロチロシナーゼの軽度の変性、 すなわち立体構造の微細な変化によって引 き起こされると考えた6,7)。
方、 イエバエ(Musca domestica)において、 変態、に伴うPO活性の変 動が詳細に研究されている。 船体、 稲葉らは8-10)、 イエバエ幼虫および!腕に おけるフェノールオキシダーゼ、の活性変動を調べ、 幼虫末期に高い活性を不 したフェノールオキシダーゼが姉化と同時に表面的な活性を全く消失し、 成 熟畑になると再び活性が出現することを観察した。 さらに、 焔化直後の姉 (前蜘)の抽出液にオレイン酸ナトリウムのような陰イオン界面活性剤また は成熟畑のホモジネートを加えると、 活性が出現することを見いだし、 フェ ノールオキシダーゼは前畑中で不活性なプロエンザイム、 すなわち、 プロフ ェノールオキシダーゼ(PPO)として存在していること、 成熟姉中にはプロ フェノールオキシダーゼを活性化する遊離のアクチベーター(natural activator)が存在していることを確認した8)。 次いで、 プロフェノールオキ シダーゼの活性化について種々検討した結果、 イエバエ前腕プロフェノール オキシダーゼ、のin vitro�こおける界面活性剤による活性化はオレイン酸ナト リウムやSDSのような陰イオン性界面活性剤によってのみ起こり、 陽イオン 性および非イオン|全界面活性剤では起こらないこと、 プロフェノールオキシ ダーゼとSDSがある一定量比関係におるときにのみ効果的な活性化が起こる ことを明らかにした9)。 さらに活性化のpH依存性、 温度依存性等を検討し、
活性化はプロチロシナーゼ分子の構造変化によるものであろうと推察した9,
10)。 このように変態過程においてPOは、 重要な役割を演じているにも関わ らずイエバエ以外では、 変態に関わるPOの役割が調べられていない。
カイコは、 (1)全生涯を通じて完全に人工的に制御できるので、 いつで も計画する時期に飼育して使用できる。 (2)入手、 飼育が容易である。
(3)生物
学的に発育段階の斉
一な材料を同時に多量に準備できる、 などの理由により昆虫生理学、 特に変態、に関する研究を行うには非常に適したモデ
ル動物である。 芦田らは、 カイコ体液より初めてPOの精製に着手し、 その
過程において体液中ではプロフェノールオキシダーゼとして存在することを
見いだした11)0 PPOは、 80kDaあたり2原子の銅を含む銅タンパク質である ことが確かめられた。 続いてカイコの幼虫外皮から活性化因子を精製した12,
13)。 この活性化因子は、 分子サイズ60 kDaでトリプシン用基質であるベン ゾイルアルギニンエチルエステルやトシルアルギニンエチルエステルの加水 分解を触媒するエステラーゼ活性を示すことを認めた。 さらにこの活性化因 子によりPPOを活性化すると一本のペフチドが遊離することを明らかにし た。 芦田らは、 カイコを用いた一連の実験において、 PPOの活性化因子によ る活性化の機構としてチモーゲンアクチベーションを考えた。 すなわち、
PPOは活性化因子によって限定加水分解を受けPOになるというものであ
る。 さらに、 彼らは、 カイコ体液中にカビの細胞壁成分(β-1,3-グルカン) あるいはバクテリアの細胞壁成分(ペフチドグリカン)が数十から数百ピコ グラム存在するとPPOが活性化されて最終的にメラニンが生じること、 そし てペプチドグリカンおよびか1,3-グルカン認識タンパク質が存在することを 明らかにし、 侵入異物の認識がPPO活性化システムの引き金を引くと提案し ている14)。 近年、 カイコ血球細胞由来のcDNAライブラリーから2種のPPO 遺伝子がクローニングされ、 そのアミノ酸配列が推定されている15)。 その結 果、 節足動物のヘモシアニンと相同なタンパク質であることが明らかとなっ た。 その他にも、 ザリガニ、 ショウジョウバエそしてタバコスズメガ由来の PPO遺伝子がクローニングされ、 その塩基配列が決定されている16-18)。 以
上述べたことがカイコのフェノールオキシダーゼについては調べられている が、 その生育過程におけるPOの変動および、その過程におけるPOの役割につ いては調べられていない。
カイコ変態過程におけるPOの役割を調べるにあたり、 著者はまずPPOの 精製法と検出法について検討を行った。 芦田によるPPOの精製法は、 1971 年に確立されている。 それから20年以上経た現在タンパク質の精製および分 析技術が飛躍的に進歩している。 さらに簡便かつ効率よくPPOを精製する方 法を確立する必要があると考えられる。 また、 PPOの検出に用いられている 活性化酵素は不安定であり、 繰り返し調製しなければならない。 この活性化
3
酵素によるPPOの活性化は、 高い塩濃度存在下で阻害されるため、 活性測定 的にサンプルの脱底をしなければならず、 操作が非常に煩雑である。 そこ で、 活性化酵:素を用いた活性測定法に変わる新たな測定法の開発を試みた。
この測定法を用いて活性染色を行ったところa80系統の体液中には3種のイ ソフォームが存在することが明らかとなった。 カイコにおけるPPOの生理学 的機能を調べるためには、 これらのイソフォームを分隊、 精製しその諸性質 を比較、 検討することが必要と考えられる。 そこで、 これらのPPOイソフォ ームを分離、 精製し、 その諸性質について調べた。 次に、 先に述べた新規活 性測定法によりカイコ生育時期!におけるPO活性の変動を調べた。 また、 精 製したPPOをマウスに対して免疫を行い抗PPO抗血清を作製し、 ELISA法に よりPPO合成量について検討を行った。 最後に、 このPPOの合成が転写レベ ルあるいは翻訳レベルにおいて制御されているのか調べるため、 PPOをコー
ドする遺伝子をクローニングし、 得られたcDNAをフローブとして、 ノーザ ンハイブリダイゼーション法を用いることによりmRNAの変動を検討した。
本論文は、 これらに関する結果をまとめたものである。
参考文献
1) J. H. Bodine and E. J. Boell, J. Cellular Comp. Physio1., 6,263 (1935).
2) J. H. Bodine and T. H. Allen,斤oc. Soc. Exptl. ßio1. Med., 37,450 (1937).
3) J. H. Bodine and T. H. Allen, J. Cel1ular Comp. Physiol., 12, 71 (1938).
4) T. H. Allen and J. H. Bodine,庁oc. Nat1. Acad. Sci. U.S.A., 27,269 (1941).
5) T. H. Allen, A. B. Otis and J. H. Bodine, J. Gen. Physiol., 2 6, 151 (1943).
6) T. H. Allenむld J. H. Bodine, J. Gen. Physio1., 2 4, 99 (1940).
7) J. H. Bodine and L. D. Carlson, J. Gen. Physiol., 2 4,423 (1940).
8) M. Funatsu and T. Inaba, Agr. Biol. Chem., 26, 535 (1962).
9) T. Inaba, Y. Suetake and M. Funatsu, Agric. Biol. Chem., 27, 332 (1963).
10)稲葉
喬:九州大学農学部生物化学論文 第4号、1964.
11) M. Ashida, Arch. Biochem. Biophys., 1 4 4, 749-762 (1971).
12) K. Dohke, Arch. Biochem Biophys., 157, 203-209 (1973).
13) K. Dohke, Arch. Biochem. Biophys., 157,210-221 (1973).
14) M. Ashida and Y. 1. Yamazaki, 1n Molting and Metamorphosis (Edited by E. Ohnishi and H. Ishizald), 239-265. Japan Scientific Societies Press, Tokyo (1990).
15) T. Kawabata, Y. Yas山ara, M. Ochiai, S. Matsuura and M. Ashida,
Proc. Natl. Acad. Sci. USA, 92, 7774-7778 (1995).
16) A. Aspán, T. Huang, L. Cerenius and K. Söderhäll, Proc. Natl.
Acad. Sci. USA, 92, 939-943 (1995).
17) K. Fujimoto, N. Okino, S. Kawabata, S. Iwanaga and E. Ohnishi,
roc. Natl. Acad. Sci. USA, 92, 7769-7773 (1995).
18) M. Hall, T. Scott, M. Sugumaran, K. Söderhäll and J. H. Law,斤oc.
Natl. Acad. Sci. USA, 92, 7764-7768 (1995).
第二章 ブロフェノールオキシダーゼの精製法および検出法の確立
第一節 緒言
1971年、 芦田は1)、 カイコ体液PPOをディスク電気泳動的に均一な標品と して精製した。 それから20年以上経た現在、 タンパク質の精製および分析技 術が飛躍的に進歩している。 そこで本章では、 芦田の方法よりも簡便かつ迅 速なPPO精製法を確立することを目的とした。 また、 この目的のために要求
されるPPOの非酵素的活性化法について検討を行った。
第二節 実験材料
第一項 カイコ系統および、体液採取法
カイコ、 Bombyχmori (a80系統、 九州大学遺伝子資源センタ一家蚕遺 伝子部門)は、 桑葉を用いて飼育した。 体液は、 5齢4日の幼虫の腹脚を注射 針で突き刺し、 液体窒素中に滴下して採取した。 採取した体液は凍結乾燥 し、 使用時まで-300Cで保存した。
第二項 皮膚採取法
カイコ5齢4日の幼虫を氷冷した生理食庖水中にて解剖し、 皮膚を採取し た。 採取した皮膚は、 1%アスコルビン酸ナトリウム溶液を用いて洗浄し、
その後、 凍結乾燥し、 使用時まで-300Cで保存した。
第三項 試薬
ブチルトヨパール650Mは東ソ一社製を、 ヒドロキシアパタイトはナカラ イ社製を、 セフアクリルS-200、 Mono-QおよびセファデックスG-25はフ
ァルマシア社製を用いた。 ジメチルベンジルミリスチルアンモニウムクロリ ド (DBMA)は、 和光純薬より購入した。 その他の試薬は、 市販の特級のも のを用いた。
7
第四項 PPO活性化因子の調製
PPO活性化因子(prophenolòxidase activating en勾rme, PPAE)は、
Dohkeの方法に従い部分精製した2 )。 遠心分離は全て40C、 5,600Xgで行っ た。 第二項に述べた方法に従い採取した皮膚を緩衝液A(10 mMトリス塩
酸緩衝液(pH 9.0) /1 mMジエチルジチオカルバミン酸ナトリウム/0.5 M NaCl/10 mM CaC12)中で磨砕した。 遠心分離を行い、 上清をガーゼで滅
し、 凍結乾燥を行った。 凍結乾燥粉末300mgに20%硫酸アンモニウムを含 む10mMリン酸カリウム緩衝液(pH 7.5)を100ml力日え、 40Cで、1時間撹 持した。 その後、 15 分間遠心分離し、 上清に40%となるように硫酸アンモ ニウムを加え、 1晩40Cで撹枠した。 15分間遠心分離を行い、 沈殿を回収し 少量の10mMリン酸カリウム緩衝液(pH 7.5)に懸濁し、 遠心して沈殿を 除去した後、 同緩衝液で平衡化したセファデックスG-25カラム(1.6X21.5 cm)に供与した。 溶出は、 同緩衝液にて行った。第三節 実験方法
第一項 プロフェノールオキシダーゼの活性化 (1) PPAEを用いたPPOの活性化
活性測定に用いる試料は、 マイクロダイアライザー(アト一社製)により 10mMトリス塩酸緩衝液(pH 7.5)に対して12時間以上透析したものを用 いた。 PPAEは、 第二節で調製したものを用いた。 PPAE 50μ!と試料溶液50 μlを氷中で30 分間混合した後、 生じたPO活性を測定した。
(2)界面活性剤を用いたPPOの活性化
界面活性剤(SDS、 DBMA、 Trito沿(-100)を用たPPOの活性化において は、 界面活性剤を様々な濃度になるよう0.1 Mリン酸カリウム緩衝液(pH 6.5)に溶解し、 等量の酵素溶液と混合して室温に放置した。 混合液より50 μiずつ分取し、 生じたPO活性を測定した。
(3) 2-プロパノールを用いたPPOの活性化
2-プロパノールを用いての活性化は、 浅田らの方法3)を若干改良して行っ た。 すなわち、 2-プロパノールを段々な濃度になるように0.1 Mリン酸ナト リウム緩衝液に溶解し、 等量の酵素溶液と氷中で混合した。 混合液より50 μlず、つ分取し、 生じたPO活性を測定した。
第二項 プロフェノールオキシダーゼ活性化後のPO活性の検出 (1) PPAEを用いたフェノールオキシダーゼ活性の検出
活性化後のPO活性の検出は、 以下の方法で行った。 すべての操作は、
300Cにて行った。 石英セル中にて0.5mlの10mM L-3-(3. 4-ジヒドロキシ フェニル)アラニン(L-ドーパ)を含む12mM HClと0.5ロ吐の10mMリン 酸カリウム緩衝液(pH 6.5)を30秒撹伴、 混合した。 続いて第一項(1)の 方法で活性化された溶液50μlを添加し、 475nmにおける吸光度の増加を測 定した。 PPAEを用いて活性化する場合は、 5分間に吸光度を0.01憎加させ る酵素量を1ユニットと定義した。
(2) DBMAを用いたフェノールオキシダーゼ活性の検出
DBMAによるPPOの活性化におけるpHの影響を検討する場合は、 PPO 溶 液と等量の 0.4mMDBMA を含む各pHで調製した広域緩衝液を混合し、
300Cで保温した。 この反応液より50μl分取し、 生じたPO活性を測定した。
温度の影響を検討する場合は、 PPO溶液と等量の0.4mMDBMAを含む0.2 Mリン酸カリウム緩衝液(pH 6.5)を混合し、 各温度にて保温した。 この 反応液より50μl分取し、 生じたPO活性を測定した。
DBMAを用いたフェノールオキシダーゼ活性の検出は、 以下の方法で行っ た。 すべての操作は、 300Cにて行った。 石英セル中にて0.5mlの10mM L
ドーパを含む12mM HClと0.5mlの10mMリン酸カリウム緩衝液(pH 6.5)を30秒撹伴、 混合した。 DBMAにより活性化されたプロフェノールオ キシダーゼ溶液0.05mlを添加することにより反応を開始させ、 475nmの吸
9
光度変化を5分間測定した。 1ユニットは、 ドーパクロムのモル吸光係数 3,715 M-lcm -lを用いてl分あたり1μmolのドーパクロムを生産する酵素量
と定義した。
第三項 タンパク質の定量
タンパク質の定量は、 牛血清アルブミンを標準タンパク質としてLowηrら の方法4)に従い測定した。
第四項 純度検定(SDS-PAGE)
SDS-PAGEは、 Laemmliの方法5)に従って行った。 すなわち、 泳動は、
SDS-トリスーグリシン系(泳動pH 8.8)を用いスラブ型10 %分離ゲルにて 行った。 試料溶液は、 1%SDS、 1%2-メルカプトエタノール、 0.01%ブロ モフェノールブルー、 20%グリセロール、 10mMトリス塩酸緩衝液(pH
6.8)になるように調製し、 1000Cで3分間処理によりSDS化後PAGEに供与 した。 PAGE後のタンパク質の検出は、 0.25%クマシーブリリアントブルー R-250 (CBB)染色または銀染色にて行った。第五項 未変性ポリアクリルアミド電気泳動(Native-PAGE)
未変性ポリアクリルアミド電気泳動 (Native -PAGE)は、 Davisの方法6) に従った。 すなわち、 泳動は、 トリス-グリシン系を用いスラブ型4.75%お よび10%分離ゲ、ルにて行った。 試料溶液は、 50%スクロース, 0.1%ブロモ フェノールブルー溶液と等量混合して供与した。 PAGE後のタンパク質の検
出は、 銀染色にて行った。
第六項 プロフェノールオキシダーゼの検出(活性染色法)
電気泳動後のプロフェノールオキシダーゼの検出は、 以下の方法で行っ た。 すなわち、 ポリアクリルアミド電気泳動の後、 ゲルを5m ML-ドーパと 0.2mMDBMAを含む0.1M リン酸カリウム緩衝液 (pH 6.5)溶液に室温
で浸した。 活性化されたPPOによりLードーパはメラニンとなり黒色のバンド
として生じる。 適当な濃さのバンドが得られた後、 蒸留水でゲ、ルを洗浄して 基質を取り除いた。
第四節 実験結果
第一項 プロフェノールオキシダーゼの分郎、 精製
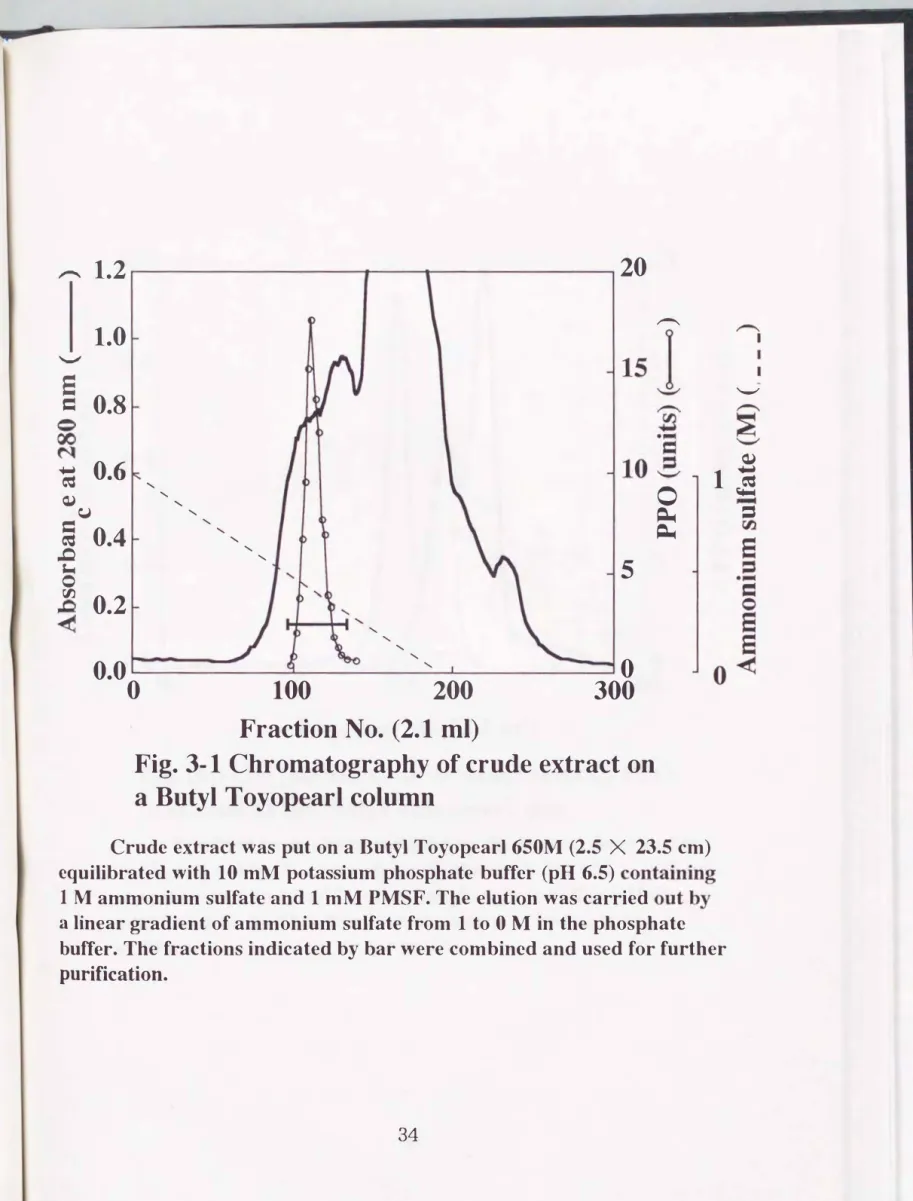
フロフェノールオキシダーゼは、 以下の操作によって精製した。 特に断 らない限り、 精製操作はすべて 4tで行った。 凍結乾燥した体液17gを1mM PMSFおよび4M NaClを含む10mMリン酸ナトリウム緩衝液(pH 7.0)に 溶解した。その後5,6 00X gで30分間遠心分縦を行い上清を粗抽出液とし、 同 緩衝液で平衡化したブチルトヨパール650Mカラム(2.2 X 30 cm)に供与 した。 同緩衝液で カラムを洗浄後、NaCl濃度を 4Mから1Mに直線的に下げ、
吸着した蛋白質を溶出し、活性画分 (2 M付近)を集めた(Fig. 2-1)。 得 られた活性画分を限外癒過膜(HI膜、 旭化成社製)を用いて減圧濃縮し、 l MNaClを含む10mMリン酸ナトリウム緩衝液(pH 7.0)で平衡化したセフ
アクリルS-200カラム(2 X 140.5 cm )に供与した。 同緩衝液で溶出し、
活性画分(No. 217-247)を集めた( Fig.2-2)。 得られた活性画分を1M NaClを含む10mMリン酸ナトリウム緩衝液(pH 7.0)で平衡化したヒドロ
キシアパタイト (1.2 X 7 cm )に供与した。 同緩衝液で洗浄後、 緩衝液中 のリン酸ナトリウム濃度を10mMから100mMまで直線的に上昇させ、 吸着
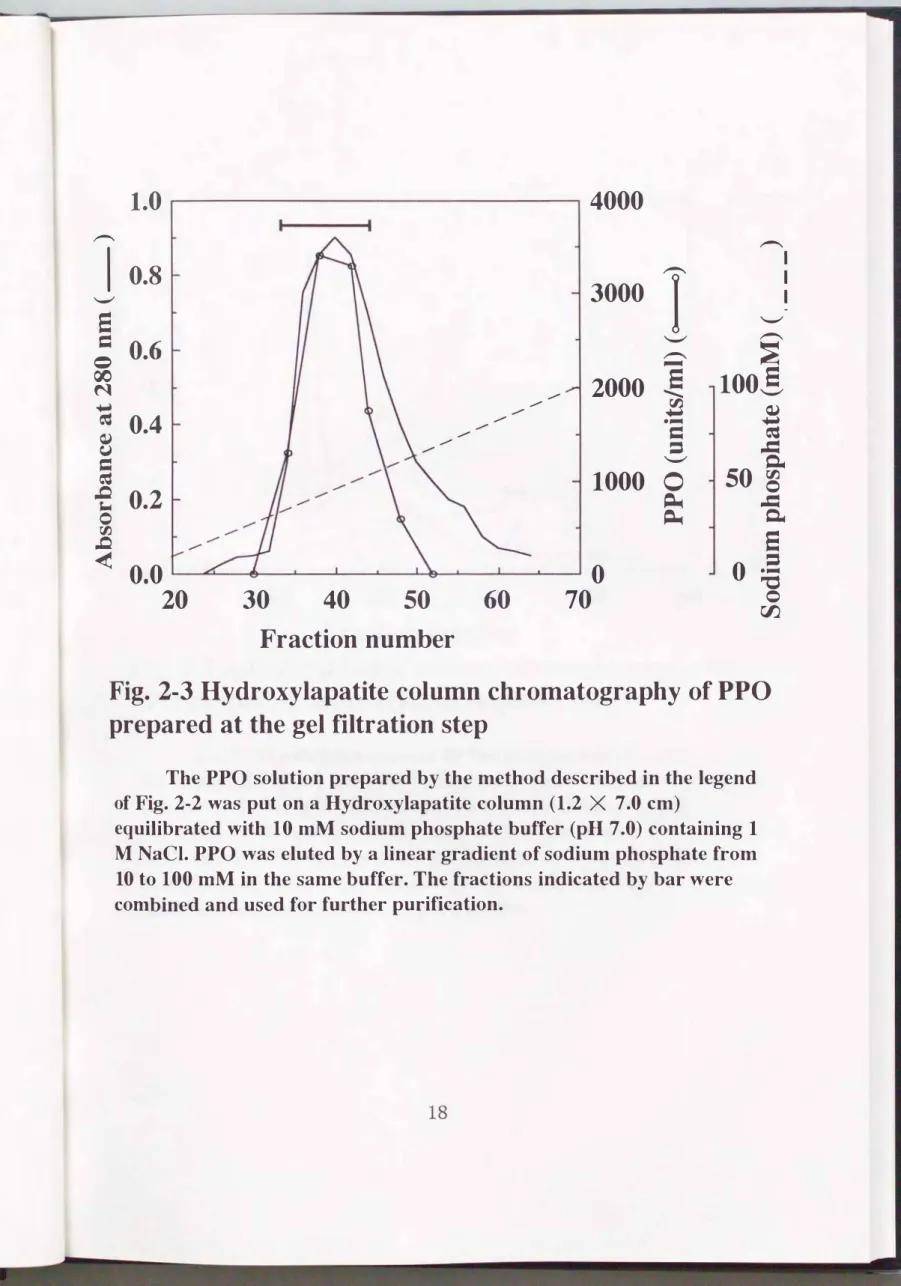
したタンパク質を溶出し、 活性画分(50mM付近)を集めた(Fig.2-3)。
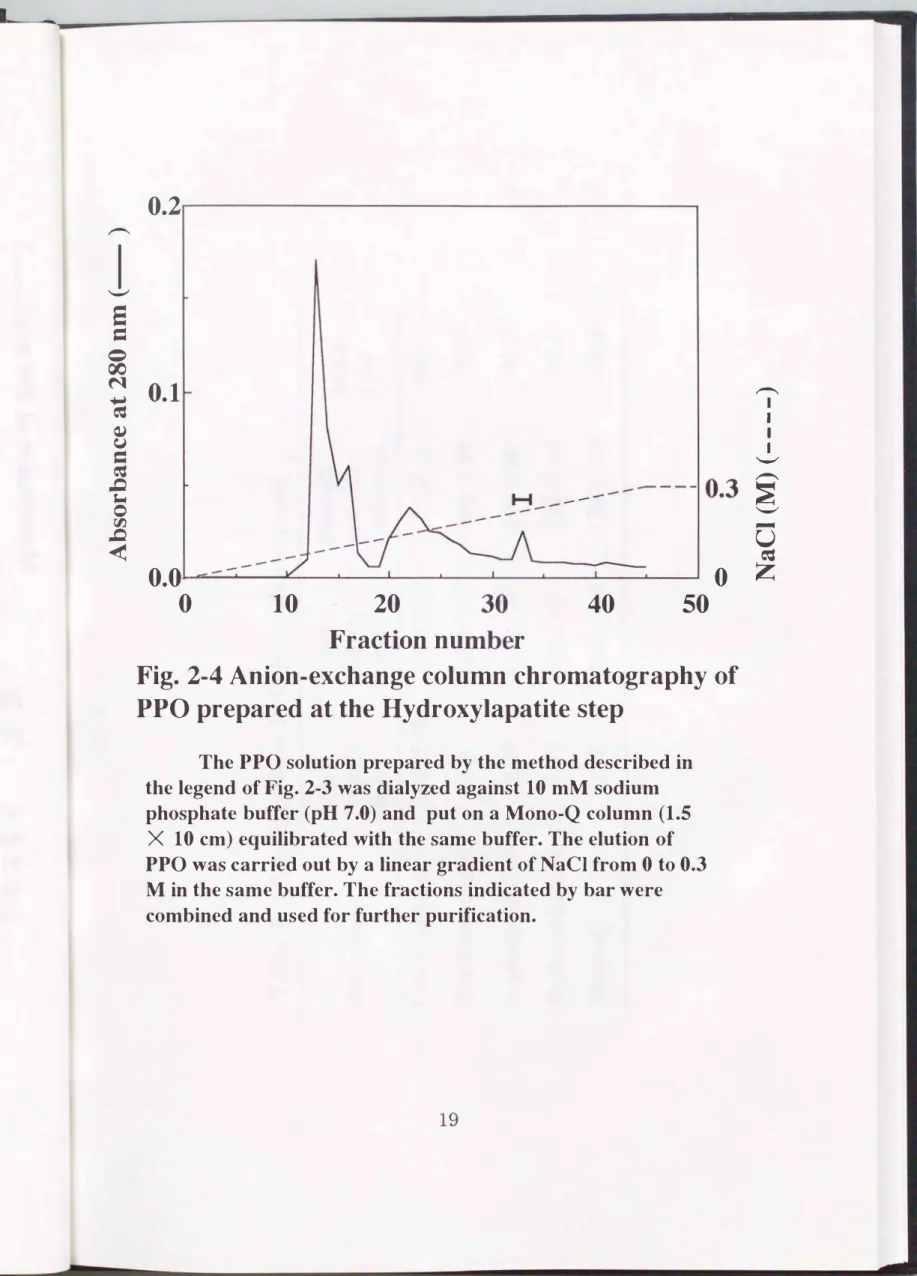
得られた活性画分を10 rnMリン酸ナトリウム(pH 7.0)に対して1晩透析 した後、 同緩衝液で平衡化したFPLC Mono-Q HRカラム(1.5 X 10 cm )
に供与した。 同緩衝液でカラムを洗浄後、 緩衝液中のNaCl濃度をOMから
0.3Mまで直線的に上昇させ、 吸着したタンパク質を溶出した(Fig.2-
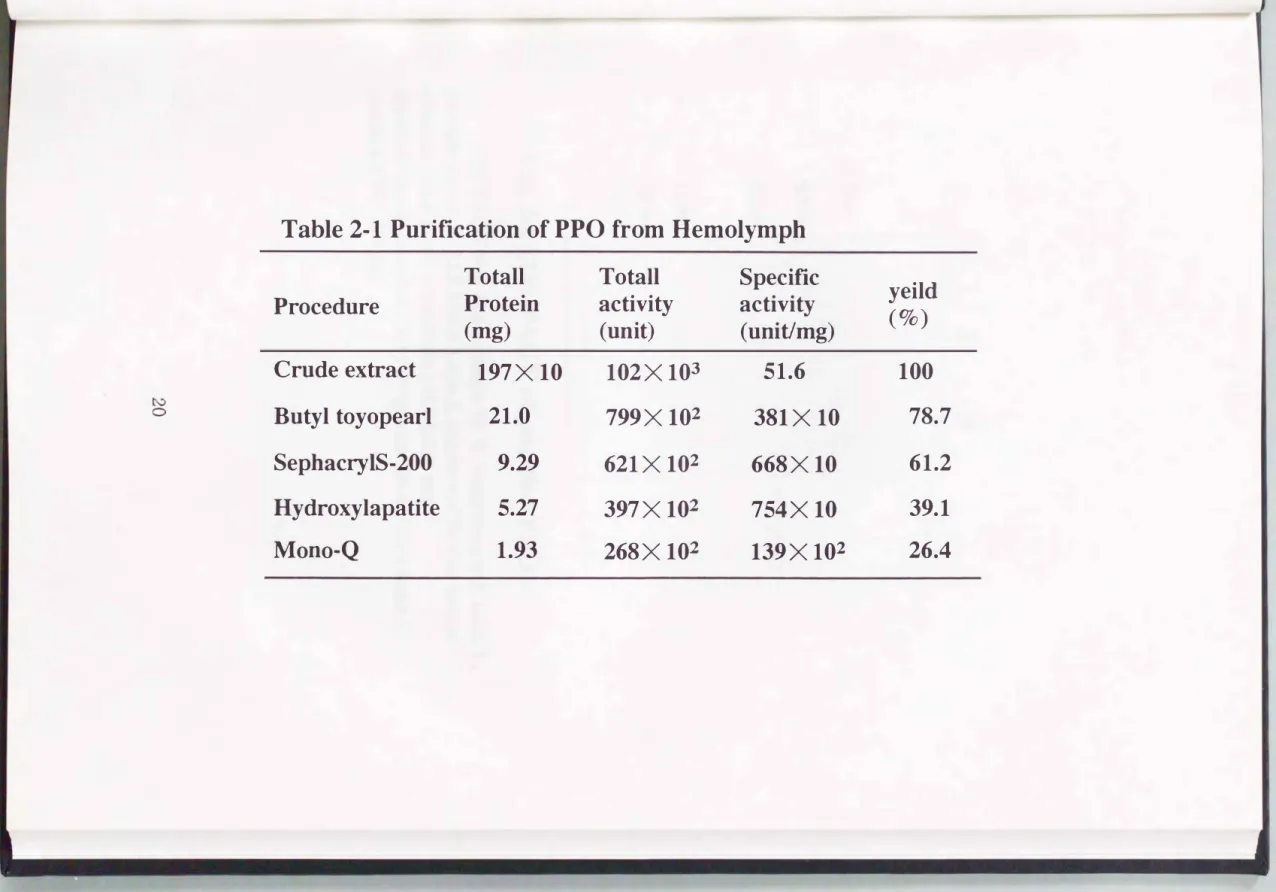
4)。 得られた活性画分( 0.25mM付近)を精製標品とした。 以上述べた精 製段階における酵素活性、 タンパク質量およびその比活性をTable 2-1に ま とめる。 精製標品を SDS-
PAGEに供したところ分子サイズ75 kDaの移動度
を示す位置にl本のバンドが検出された(Fig.2- 5)。
第二項 プロフェノールオキシダーゼの活性化
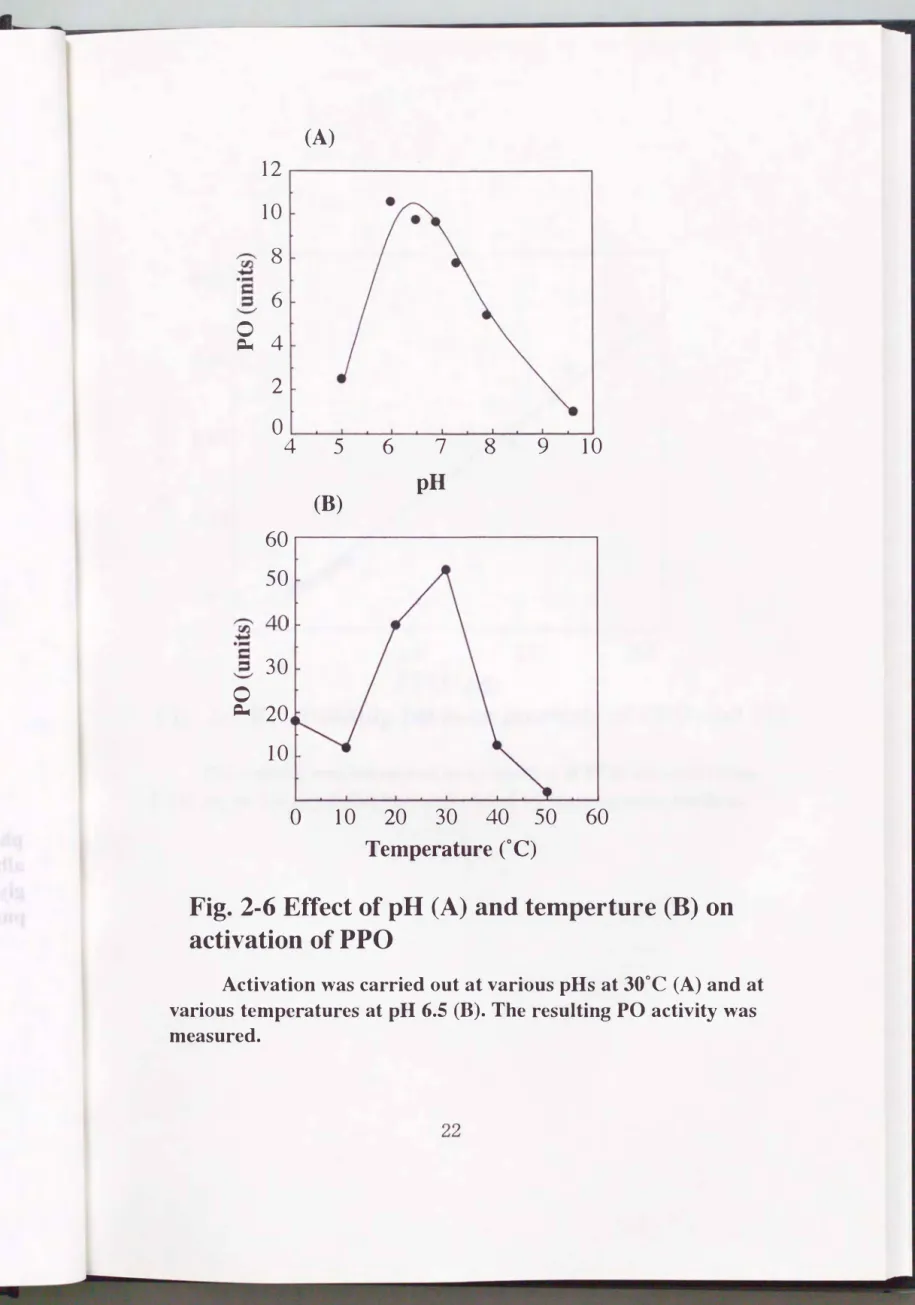
第一項で精製したPPOを用いてT吋to心仁一10 0、 SDS、 DBMAそして2ープ ロパノールによる活性化について検討した。 その結果、 カイコPPOは、
SDS、 2-propanolそしてDBMAにより活性化されたがTrito沿(-10 0では活 性化されなかった。 DBMAを用いた場合において、 カイコPPOは最も活性化 されることがわかった。 そこで、 DBMAによるPPOの活性化におけるpHお よび温度の影響について調べた(Fig.2-6)。 その結果、 pH 6.5および300C において最も活性化されることがわかった。 この条件下において、 活性測定
系に用いたPPO量と生じたPO量の関係について検討したところ、 PPO量が 0.047μgから2.8 μgの範囲において測定可能であった(Fig.2-7)。
第三項 DBMAとPPAEによるプロフェノールオキシダーゼの活性化の
比較
フロフェノールオキシダーゼをDBMAで活性化した場合とPPAEにより活 性化した場合に生じるPO活性の経時的変化について調べた。 結果をFig.2- 8に示す。 DBMAで活性化した場合、 PPOは直ちに活性化され1分後に最大 活性を示し、 その後徐々に減少した。 PPAEで活性化した場合、 活性化は DBMAに比べて遅く、 15分後にDBMAで活性化した場合の最大活性と同程 度まで活性化され、 その後ほぼ一定とな った。
第四項 電気泳動後のプロフェノールオキシダーゼの検出
凍結乾燥体液をリン酸カリウム緩衝液に懸濁し、 とれを粗抽出液とし、
10% native-PAGEに供与し、 銀染色および活性染色を行った(Fig.2- 9)。 銀染色によって、 多くのバンドが検出された。 活性染色を行った場 合、 精製したPPOと同じ移動度に1本のバンドが検出された。 一方、 粗抽出 液を4.7 5% Native-PAGEに供与し活性染色を行った場合、 3本のバンドが得
られた。
第五節 考察
芦田は、 硫安分画、 熱処理、 DEAE-セルロース陰イオン交換カラムクロ マトグラフィー、 ヒドロキシルアパタイト吸着カラムクロマトグラフィー、
フェニルセフアロースCL-4B疎水カラムクロマトグラフィーの4段階により カイコ体液よりPPOを精製している1)。 本章において著者は、 ブチルトヨパ ール疎水カラムクロマトグラフィー、 セフアクリルS-200ゲル滅過カラムク ロマトグラフィ一、 ヒドロキシアパタイト吸着カラムクロマトグラフィーそ してFPLCシステムを用いたMono-Q陰イオン交換カラムクロマトグラフイ ーの4段階でPPOを精製する方法を確立した。 この精製法により、 熱処理な
どの煩雑な操作を省略することができた。 また、 FPLCシステムを用いるこ とにより迅速にPPOを精製することが可能となった。
PPOは、 表皮中に存在する活性化因子(PPAE)により活性化されること が知られている7-9)。 しかし、 この活性化因子は、 不安定であり活性測定の 標準化が困難であるため、 繰り返し調製しなければならない。 また、 活性化
因子によるPPOの活性化反応が高い塩濃度により阻害されるため、 例えば PPO精製|時の活性測定に際して多数のサンプルを脱塩をしなければならな い、 などの欠点がある。 一方、 非酵素的な活性化は、 これまでに、 ショウ ジョウバエPPOは、 2-プロパノールにより活性化されることが3)、 またイエ バエPPOは、 SDSにより活性化されることが報告されている10)。 しかし、
カイコPPOについては、 非酵素的な活性化について調べられていない。 本章 において、 カイコPPOをDBMAで活性化した場合とPPAEで活性化した場 合、 PPOから誘導されるPO活性を比較すると、 その最大活性はほぼ同値で あった。 そこでDBMAによる活性化を利用して新規な活性測定法を開発し た。 この方法では、 0.047μgから2.8μgまでのPPOが検出可能であった。 本 章で確立した検出法を用いることにより、 以下の問題点を解決することがで きた。 (1)高塩濃度存在下においてもPPOは活性化され、 PPOから誘導さ
れるPO活性を測定することが可能となった。 すなわち、 精製時など多数の サンプルを脱息する操作を省くととができた。 (2)PPAEを用いた活性測 定法と比較して、 活性測定の標準化が容易となった。 すなわち、 繰り返し PPAEを調製する操作を省くことができた。 PPOは、 さまざまな無脊椎動物 より精製されているが、 その諸性質を詳細に検討しているものは少ない。 こ れは従来行われていたPPAEを用いてのPPO検出法が煩雑なためと考えられ る。 木章で篠立したPPO検出法を用いてPPOの諸性質を検討した結果を第三
章に記載した。
第六節 小括
(1)凍結乾燥したカイコ(a80系統)体液よりブチルトヨパール疎水カラ ムクロマトグラフィー、 セフアクリルS-200ゲル鴻過カラムクロマトグラフ ィ一、 ヒドロキシアパタイト吸着カラムクロマトグラフィーそしてMono-Q 陰イオン交換カラムクロマトグラフィーを用いてPPOを電気泳動的に均一に 精製する方法を確立した。
(2)凍結乾燥した体液17 gより比活性139
X
102ユニット/mgの酵素標品 を得た。(3)カイコ由来のPPOは、 SDS、 DBMAそして2-プロパノールにより活性 化されることがわかった。 また、 DBMAで活性化した場合とPPAEで活性化
した場合、 PPOから誘導されるPO活性を比較すると同程度であった。
(4) DBMAによるPPOの活性化反応の至適条件は、 pH 6.5および300Cで あった。 この条件において0.047μgから2.8μgまでのPPOが検出可能で、あっ た。
(5)本章において確立した活性染色法によりカイコa80系統体液中には、 3 種のPPOイソフォームが存在することが明らかとなった。
参考文献
1) M. Ashida, Arch. Biochem. Biophys., 144,749-762 (1971).
2) K. Dohke, Arch. Biochθm Biophys., 157,203-209 (1973).
3) N. Asada, T. Fukumitsu, K. Fujimoto and K. Masuda, Insect Biochem. Molec. Biol., 23, 515-520 (1993).
4) O. H. Lowry, N. J. Rosebrough, A. L. F訂r and R. J. Randall,よBiol.
Chem., 19 3, 265-275 (1951).
5) U. K. Laemmli, Nature ,227, 680-685 (1970).
6) B. J. Davis, Ann . N. Y. Acad. Sci., 1 2 1,404-427 (1964).
7) M.
Yonemura, K. Kasat釘lÎ,N.
Asada訂ldE.
Ohnishi,Zoo1. ScÍ., 8,
865-867 (1991).
8) A.
Asp組,J. Sturtevant, V. J. Smith and K. Söderhäll,
Insect Biochem., 20,709-718 (1990).9) K. Dohke, Arch. Biochem. Biophys., 15 7, 210-221 (1973).
10) T. lnaba and M. Funatsu, Agric. Bio1. Chem., 28,206-215 (1964).
(lili--)(宮)む偲Z
組岨T 今,M
AO
3000
S
�
州Z
白A A
2000
. .
・ .
‘、、 、“ ‘、. ‘、 句、. ー、、 “‘ 、、. ‘『
6
zd
4 3 2 1
(ll)gz。∞三偲ωωga旬。msd刊
oo ov 。
120 100
80 60
40 20
Fractioo oumber
Fig.2・ 1 Chromatography of crude extract 00 a Butyl Toyopearl columo
The crude extract was put on a Butyl Toyopearl 650M (2.1 X 30 cm) equilibrated with 10 mM sodium phosphate buffer (pH 7.0)
containing 1 M NaCI and 1 mM
P島'ISF.The elution was carried out by a
linear gradient of NaCI from
4to 1 M in the same buffer. The fractions
indicated by bar were combined and used for further purification.
3000
圏、ν (?ll) (-g\切羽田ロ)。角宮 2000
1000
-
. .
4
3
2
1
(ll)g図。∞SmwωωSG旬。msd刊
。 270 170 190 210 230 250
Fraction number (1.0 ml) 0
150
Fig. 2-2 Gel flltration chromatography of PPO prepared at the Butyl Toyopearl step
The PPO solution prepared by the method described in the Iegend of Fig. 2・1 was fiItered through a SephacryI S-200 (2 X 141 cm). The column had been equilibrated and was developed with 10 mM sodium phosphate buffer (pH 7.0) containing 1 M NaCI. The fractions indicated by bar were combined and used for further purification.
17
(ill・)(冨g)S2昏sagE句。∞ 00 0v AHv zd 咽I (?ll )(-g\お宮ロ )。AA
4000
3000
2000
1000
/ / / / / / / /
- E
. .
1.0 0.8 0.6 0.4 0.2
(ll)g図。∞23ωgf。msd刊
0 。 70 0.0
20 30 40 50 60
Fraction number
Fig. 2・3 Hydroxylapatite column chromatography of PPO prepared at the gel filtration step
The PPO solution prepared by the method described in the legend of Fig. 2・2 was put on a Hydroxylapatite column (1.2 X 7.0 cm)
equilibrated with 10 mM sodium phosphate buffer (pH 7.0) containing 1
M NaCI. PPO was eluted by a linear gradient of sodium phosphate from
10 to 100 mM in the same buffer. The fractions indicated by bar were
combined and used for further purification.
0.2
(iiXE)む匂Z
--一一一10.3
ト-1___---
0.1
(111) g図。∞N二dωω国史官。m m--d刊
0 50 0.0
0 40
V且
hp
e 割引 帥 u n n 0 4E' ρ、 a v且 F 20
10
Fig. 2・4 Anion-exchange column chromatography of PPO prepared at the Hydroxylapatite step
The PPO solution prepared by the method described in the legend of Fig. 2-3 was dialyzed against 10 mM sodium
phosphate buffer (pH 7.0) and put on a Mono-Q column (1.5
X 10 cm) equilibrated with the same buffer. The elution of PPO was carried out by a linear gradient of NaCI from 0 to 0.3
M in the same buffer. The fractions indicated by bar were
combined and used for further purification.
Table 2・1 Purification of PPO from Hemolymph
Totall Totall Specific
yeild Procedure Protein activity activity
(mg) (unit) (unitJmg) (%)
Crude extract 197X 10 102X 103 51.6 100
ぞ
。、コ
Butyl toyopearl 21.0 799X 102 381X 10 78.7
SephacηIS・200 9.29 621X 102 668X 10 61.2
Hydroxylapatite 5.27 397X 102 754X 10 39.1
Mono-Q 1.93 268X 102 139X 102 26.4
1 2 3
(一〉
(kDa) 97・-
66� � PPO
45 ...
36�
(+) Fig.2・5SDS圃PAGE ofpurified PPO
SDS・P AGE was done using a 10 % separating gel. Lane 1,
phosphprylase b (92.5 kDa); Lane 2, mixture of bovine serum albumin (66.0 kDa), ovalubmin (45.0 kDa) and
glyceraldehydephosphate dehydrogenase (36.0 kDa); Lane 3,
purified PPO (1.0 pg).
(A)
12 10
冨、g., 〆
8 6
� 4
2
。
4 5 6 7
8
9 10
60 50
・
.Fロ、冨ロ ,,
司,
40 30 C E』20
10
(B) pH
o 10 20 30 40 50 60
Temperature CC)
Fig. 2-6 Effect of pH (A) and temperture (B) on activation of PPO
Activation was carried out at various pHs at 300C (A) and at
various temperatures at pH 6.5 (B). The resulting PO activity was
measured.
0.08
0.06
、 JーEE E
・〆0.04
。� 0.02
。
。 1.0 2.0 3.0
PPO(μg)
Fig.2・7 Relationship between amounts of PPO and PO
PO activity was measured as a function of PPO amount (from
0.047 pg to 2.8 pg). Solid line: calculated by least-squares method.
23
0.25 ハU ζJ
ハυ ζJ
うん 11
11 nv
ハU ハV ハU
ハU (EE\的ト寸〈伺)kCτ沼ω〈 ハUnu AU
40 10
Activation time (min) 30 20
。
Fig.2・8 Relationship between activation time and resulting PO avtivity
PPO was mixed with an equal volume of either 0.4 mM DBMA (口)
or
PP AE (163 pg/ml) (e) in 0.2 M potassium phosphate buffer (pH 6.5).
After the mixture containing DBMA and PP AE were incubated for
various times at 300C and on ice, respectively, 50 pl aliquot was assayed
f
or PO activity. PO activity was plo“ed as a function of activation time.
1 2 3 4
(ー)
(+)
Fig.2・9 Native-PAGE ofPPO and hemolymph
Crude hemolymph was subjected to native PAGE on a
10 % separating gel, and proteins were visualized by silver staining (Lane 1). Purified PPO (Lane 2) and crude
hemolymph (Lane 3) were subjected to native-P AGE on 10 % separating gel, and PPO were detected by activity staining.
Crude hemolymph was subjected to native PAGE on 4.75 % separating gel, and PO and/or PPO were detected by activity staining (Lane 4).
25
第三章プロフェノールオキシダーゼイソフォームの分隊、 精製およびその 性質
第一節緒言
カイコa80系統の体液中には3種のPPOイソフォームが存在することが前 章の実験より明らかとなった。 PPOの生化学的および生理学的性質を調べる ためには、 まずこれらのイソフォームを分離、 精製しその諸性質を調べるこ とが必要不可欠である。 本章では、 カイコa80系統よりPPOイソフォームを 分離精製し、 その諸性質について検討した。
第二節実験材料
第一項 カイコ系統および体液採取法 前章の方法により、 体液を採取した。
第二項 試薬
Q-セフアロースは、 ファルマシア社製を用いた。 その他の試薬は、 市販 の特級のものを用いた。
第三節 実験方法
第一項 プロフェノールオキシダーゼの検出(活性測定法)
前章の方法に従い、 フロフェノールオキシダーゼをDBMAにより活性化し た後に生じるフェノールオキシダーゼ活性を利用し、 検出した。
第二項 分子サイズ
SDS-P AGEは、 前章の方法に従って行った。 ゲル瀦過は、 Andrewsの方 法1)に準じてSuperose 12HR 10/30カラムを用いて行った。 すなわち、
0.15 M塩化カリウムを含む10mMリン酸カリウム緩衝液(pH 6.5)で平衡 化し、 同緩衝液でタンパク質を溶出した。 標準タンパク質は、 カタラーゼ
(240 kDa)、 アルドラーゼ(158 kDa)、 牛血清アルブミン(68
kDa)、 卵白アルブミン(45 kDa)そしてキモトリプシノーゲンA(25 kDa)を用いた。
第三項 等電点
等電点は、 Svenss on の方法2)�こ従って等電点電気泳動を用いて測定し た。 すなわち、 pH 3.5-
10の2%アンフォラインを含む5%ポリアクリルアミ
ドゲルを用い、 40C、 200 Vで12時間泳動し、 さらに400 Vで1時間泳動し た。 PPOは、 活性染色法にて検出した。 ゲ、jレのpHは、 ゲルを2mmごとにス ライスし、 蒸留水(1ml)でアンフォラインを抽出し、 pHメーターを用い て測定した。
第四項 基質特異性(ミカエリス定数、 最大速度および触媒効率の測 定)
速度論的特性(ミカエリス定数、 最大速度そして触媒効率)を
Lineweaver-Burk plotを用いて求めた。 基質は、 L-ドーパ、 ドーパミン、
N-アセチルドーパミン、 Lーチロシンそしてヒドロキノンを0.476-3.810 mMの濃度範囲で使用した。 また、 L-ドーパ、 ドーパミンそしてLーチロシン
は475nm 、 N-アセチルドーパミンは400nrn そしてヒドロキノンは440 nrn の吸光度で前章第三節の方法で、活性測定を行った。
第四節 実験結果
第一項 プロフェノールオキシダーゼイソフォームの分離、 精製
プロフェノールオキシダーゼは、 以下の操作によって精製した。 特に断ら ない限り、 精製操作はすべて4'Cで行った。 凍結乾燥した体液3gを1M硫酸 アンモニウムおよび0.1mMPMSFを含む10mMリン酸カリウム緩衝液 (pH 6.5) 200 rnlに溶解した。その後5,600X gで30分間遠心分離を行い上 清を粗抽出液とし、 同緩衝液で平衡化したブチルトヨパール650Mカ ラム
(2.5 X 23.5 cm)に供与した。 同緩衝液でカラムを洗浄後、硫酸アンモニ ウム濃度を 1 MからOMに直線的に下げ、吸着した蛋白質を溶出し、活性画分
(0.4 M付近)を集めた(Fig.3-1)。 この活性国分を塩析により濃縮後、0.5 M KClを含む10mMリン酸カリウム緩衝液 (pH 6.5)で平衡化したセ フアクリルS-200カラム(2.2
X
140 cm)に供与した(Fig. 3-2)。 得られ た活性画分を塩析により濃縮し、 平衡化緩衝液に対して透析後、 再び同条件 にて同カラムに供与した(Fig. 3-3)。 得られた活性画分を10mMリン酸カリウム緩衝液(pH 6.5)に対して透析を行い、 同緩衝液で平衡化したQ
セフアロースカラム(0.8
X
28.5 cm)に供与し、 I(Cl濃度をOMから0.3 Mに直線的に増加させることにより溶出を行った(Fig. 3-4) 0 KCl濃度 0.25 M付近に活性画分が溶出された。 このタンパク質の溶出と活性の溶出パ ターンは一致していなかったので、 各フラクションをNative-PAGEにて分 析した。 活性染色および銀染色を行ったところ、 単一な成分を含む画分を得 ることが可能であり、 溶出JII貢にフラクション200-202をQ1,フラクション 210をQ2そしてフラクション226をQ3とした(Fig. 3-4, inset)。Q1、 Q2そしてQ3画分をNative-PAGEに供与し、
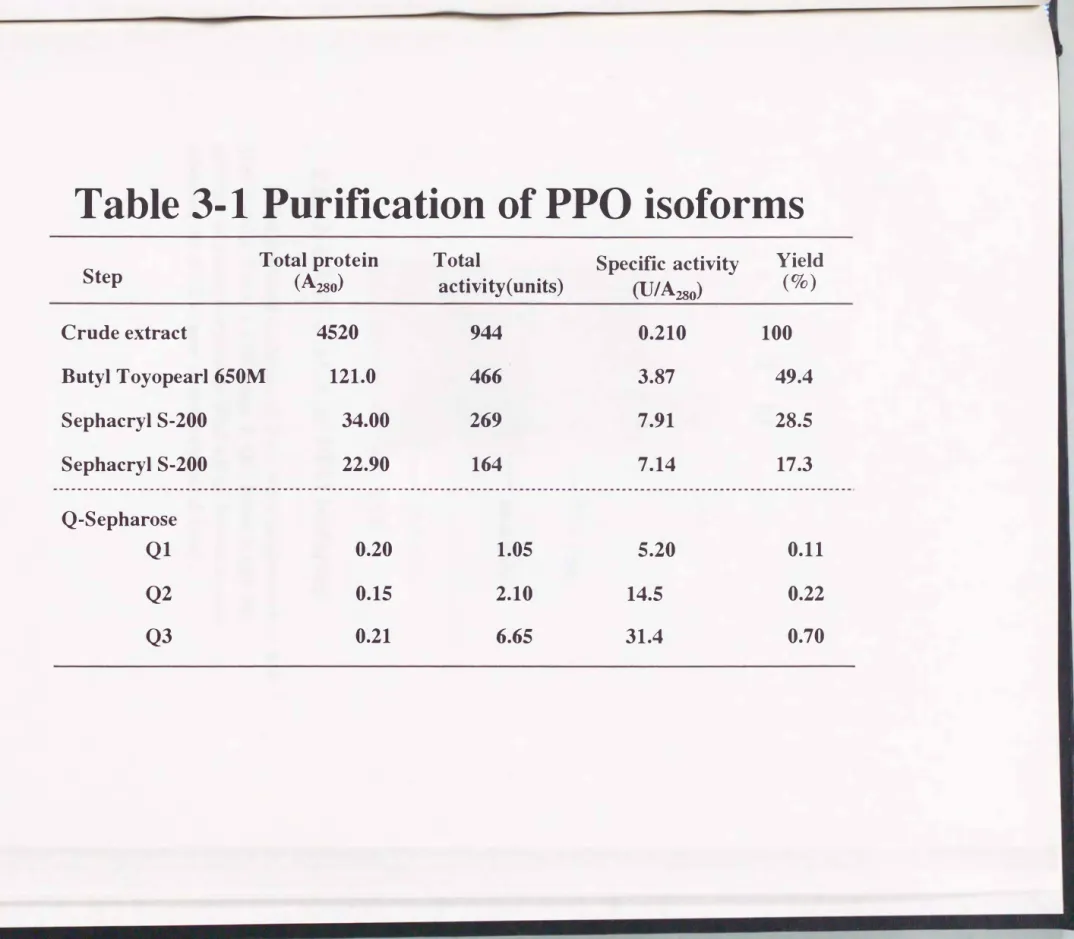
活性染色および、銀染色を行った結果をFig. 3-5に示した。 それぞれQ1,Q2そしてQ3は、 それぞれ 移動度の異なる1本のバンドを示した。 これらのバンドの移動度は、 カイコ 体液を泳動した時に生じる3本のバンドの移動度に相当していた。 この結果 より、 体液中の3種のプロフェノールオキシダーゼイソフォームが単一に精 製されたことが示された. これらの精製過程の各段階における酵素活性、 タ ンパク量および比活性を測定し、 Table 3-1にまとめる。 PPOイソフォー ム、 Q1,Q2そしてQ3は、 それぞれ粗抽出液より24.8,69.0 そして150倍ま で精製できた。 また、 比活性は5.2,14.5 そして31.4U/A280であった。
第二項 プロフェノールオキシダーゼ の性質 (1)分子サイズ
精製したそれぞれのイソフォームをSDS-PAGEに供与した結果を、 Fig.
3-6 に示す。 それぞれのイソフォームQ1, Q2そしてQ3は、 いずれも73kDa および74kDaの2本のポリペプチドより補成されていた。 Native�犬態の分子 サイズをSuperose 12HRカラムを用いたFPLC により推定した。 その結果、
Q3の分子サイズは、 122kDaと推定された(Fig. 3-7)。
(2)等電点
等電点電気泳動を用いて精製されたPPOイソフォームの等電点を調べた結 果をFig. 3-8 に示すo PPOイソフォームQ1、 Q2そしてQ3は、 それぞれ単一 のバンドを示し、 pH勾配によりその等電点は、 Q1が5.55、
Q2が5.63そし
てQ3が5.58であった。 分離されたそれぞれのイソフォームを再び混合し、等電点電気泳動に供与したところ、 それぞれ等電点5.55、 5.58そして5.63の ところ に移動した。
(3) pH安定性および、至適pH
Q1, Q2そしてQ3を異なるpHで40C、 24時間放置後、 残存活性を測定し た。 結果を、 Fig.3-9に示す。 いずれのイソフォームもpH6-10の範囲で安 定であり、 pH 6以下およびpH 10以上で不安定であった。 DBMAによる PPOの活性化反応の至適pHの結果をFig.3-10 (A)に示す。 いずれのイソフ ォームもpH 6.5付近で最大活性を示した。 また、 活性化されたPPOの触媒 反応における至適pHを測定した結果をFig. 3-10 (B)に示す。 至適pHは、
Q1が5.5から7.5、 Q2が5.5から7.5そしてQ3が5.5から6.5であった。
(4)基質特異性(ミカエリス定数、 最大速度および触媒効率の測
定)
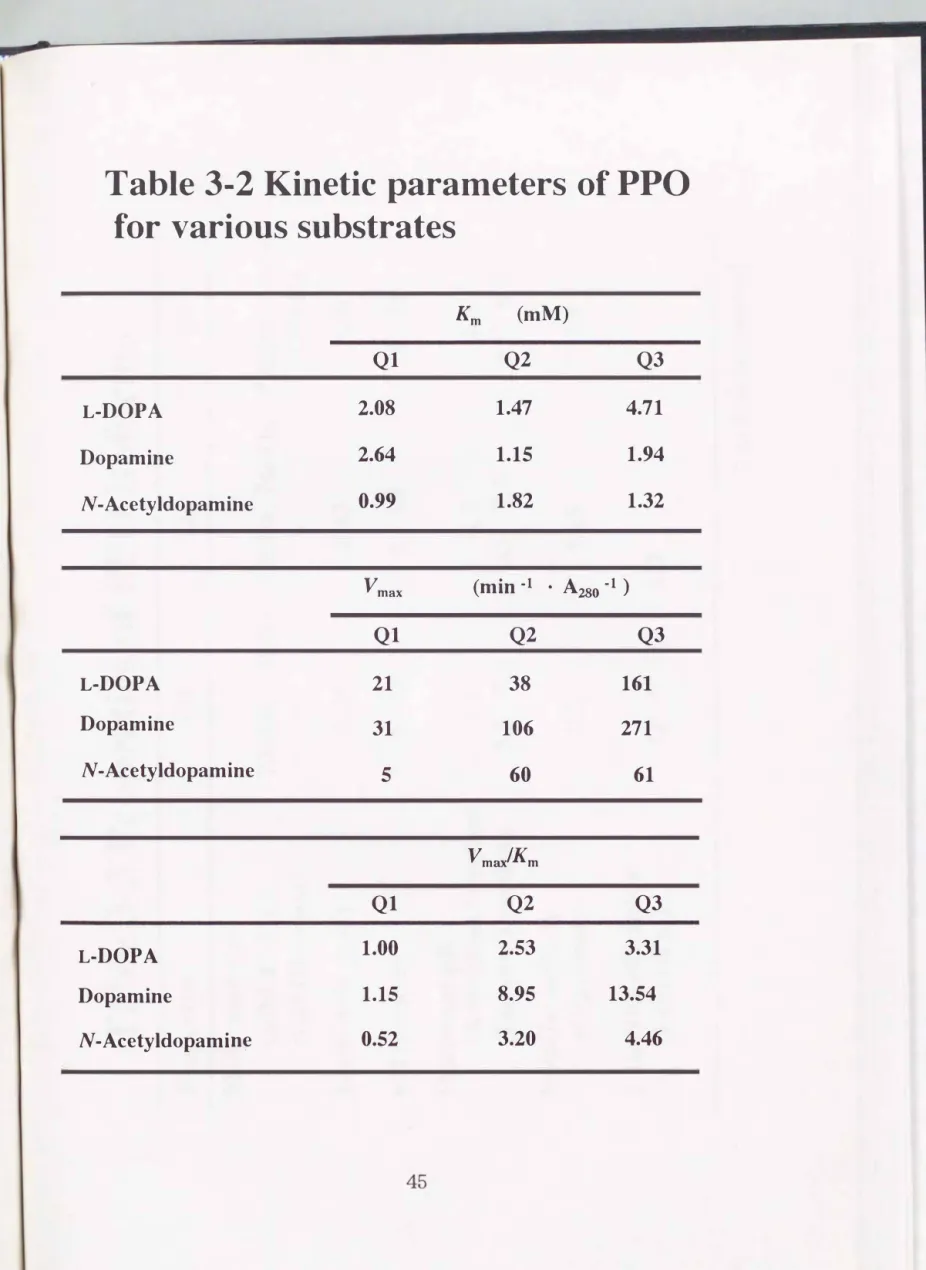
L-ドーパ、 ドーパミン、 Nーアセチルドーパミン、 Lーチロシンそしてヒド ロキノンを基質としてQ1、 Q2そしてQ3の基質特異性を調べたo また、
Lineweaver-Burlζplotを用いてミカエリス定数(Km)、 最大速度 (Vmax)そして触媒効率(Vmax/Km)を求めた(Table 3-2)。 その結
29
果、 いずれのイソフォームもL-ドーパ、 ドーパミンそしてNーアセチルドー パミンに対してのKm値は、 0.99'"'-'4.71 mMの範間にあった。 その触媒効率 は、 Q3> Q2 > Q1であった。 いずれのイソフォームもドーパミンを最も良
い某質としていた。 Lーチロシンおよびヒドロキノンの酸化は、 触媒しなかっ た。
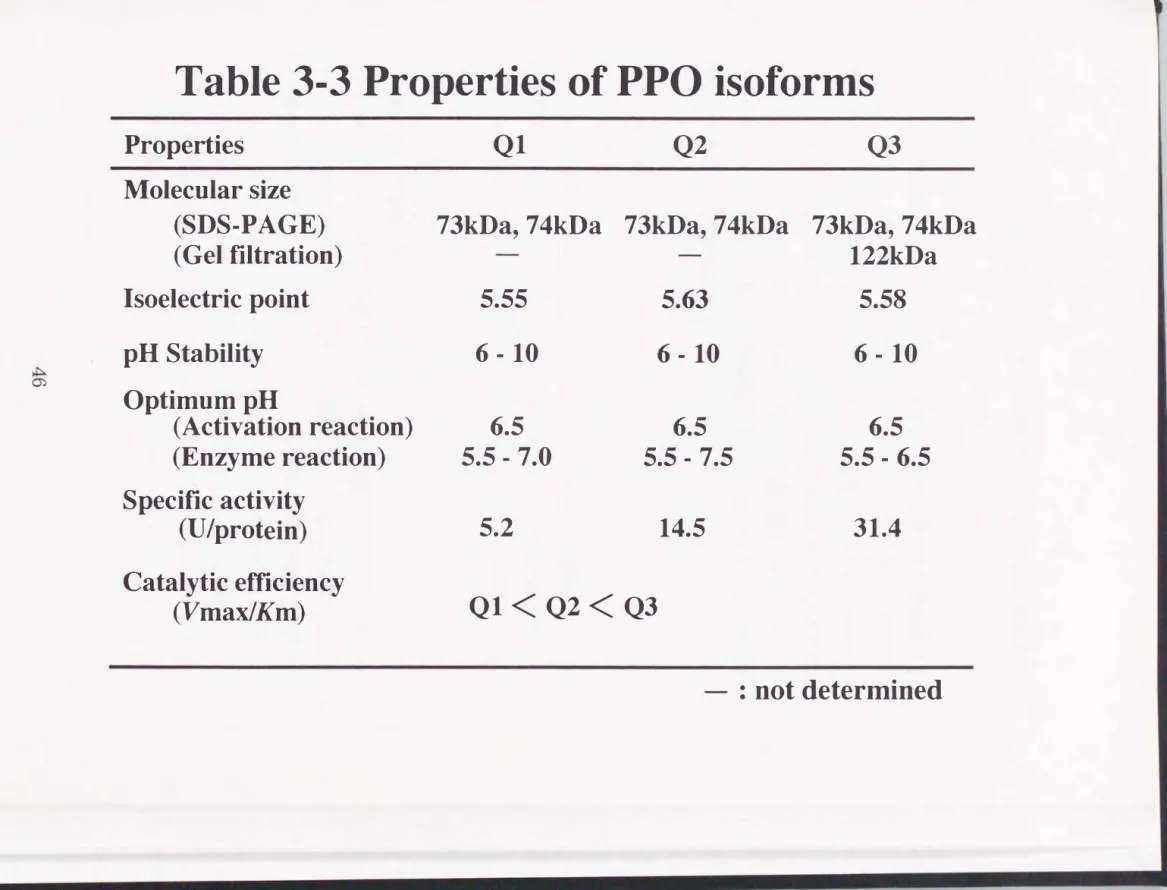
以上、 3E性質を調べた結果をTable 3-3にまとめた。
第五節 考察
カイコPPOのイソフォームの分離、 精製に初めて成功し、 Q1、 Q2および Q3の精製標品を得た。 SDS-PAGEの結果、 Q1、 Q2およびQ3は、 いずれの イソフォームにも73kDaおよび74kDaのポリペプチドが含まれることがわ かった。 Q3の分子サイズは、 ゲル瀦過を用いて調べたところ122kDaであ り、 Q3は73kDaおよび74kDaのヘテロダイマーであることが予想される。
安原らは、 錦秋×鐘和系統由来のPPOは、 71.5kDaおよび71kDaの2つの ポリペプチドより構成され、 native状態の分子サイズは、 126kDaであるこ とを報告している3)。 この結果から、 彼らは、 2つのポリペプチドは、 ヘテ ロダイマーを形成していると示唆している。 そして、 錦秋×鐙和系統の体液
中には、 2種類のヘテロダイマーの存在を報告している。 しかし、 これらの ヘテロタイマーすなわちイソフォームは、 Mono-Qやhydroxylapatiteを用 いても分離不可能であることを報告しており3)、 彼らが調べたPPOの諸性質 は、 2種類のイソフォームが混合した状態で検討したものである。 その他の 無脊椎動物において、 SDS-PAGEの結果、 ショウジヨウバエ(Drosophi1a melanogaster) PPOの分子サイズは、 78kDaおよび77kDa 4)、 ザリガニ
(Pacifastacus leniusculus)の分子サイズは76kDa 5)、 タバコスズメガ (Manduca sexta)の分子サイズは、 77kDaおよび71kDa 6)であり、 カイ コPPOの分子サイズと近い。
Q1、 Q2そしてQ3の等電点(pl)は、 5.55、 5.58そして5.63であった。 そ の他の無脊椎動物由来のPPOのpIは、 錦秋×鎧和系統のpIが、 それぞれ4.95
そして4.98であり3)、 ザリガこでは、 5.45)、 ショウジョウパエでは、 5.8お よび6.7である4)。 すなわち、 報告されている無脊椎動物由来のPPOは、 いず れも酸性タンパク質である。
前章において確立したDBMAを用いての活性測定法で基質特異性について 検討した。 いずれのイソフォームもoージフエノールの酸化を触媒したがpージ、
フエノールおよびモノフエノール類の酸化は触媒しなかった。 これに対し て、 ショウジョウバエにおいては、 モノフエノール類を酸化するものとジフ エノール類を酸化するイソフォームの存在が報告されている4)。 次にKm Vm以そしてVmax/KmをL-ドーパ、 ドーパミンそしてN-アセチルドーパミ
ンに対して測定したo Km{1直は、 0.99"'-'4.71mMの範囲にあり、 ほとんど同 じであった。 タバコスズメガにおいては、 体液のPPOと皮膚のPPOのドーパ に対するKm{1直は、 それぞれ4.15そして8.4mMであった6)。 また、 ドーパミ ンに対する両酵素のKm{1直は、 0.71および0.91であり、 ドーパに対してはカ イコ由来のPPOの方が親和性が高く、 ドーパミンに対してはタバコスズメガ 由来の方が親和性が高かった。
第六節 小括
(1)カイコa80系統の体液から、 ButylToyopearl 650M疎水クロマトグラ フィー、 SephacrylS-200ゲル癒過クロマトグラフィー、 Q-Sepharose�
イオン交換カラムクロマトグラフィーを用いて3種のPPOイソフォーム、
Q1、 Q2およびQ3を分離、 精製した。
(2)いずれのイソフォームもSDS-PAGEの結果、 73kDaおよび:74kDaの ポリペプチドより構成されていた。 ゲル滅過の結果、 Q3の分子サイズは、
122 kDaであった。
(3)等電点は、 Q1が5.55、 Q2が5.63そしてQ3が5.58であった。
(4)いずれのイソフォームもpH6-10の範囲で安定であり、 DBMAによる 活性化の至適pHは、 6.5であった。 また、 活性化されたPPOの触媒反応にお
ける至適pHは、 Q1および、Q2がpH 5.5-7.5そしてQ3がpH 5.5-6.5であっ
31
た。
(5)いずれのイソフォームもLードーパ、 ドーパミンそしてN-アセチルドー パミンの酸化を触媒した。 一方、 L-チロシンおよびヒドロキノンの酸化は、
触媒しなかった。
(6)
Lードーパ、 ドーパミンおよびN-アセチルドーパミンを用いてミカエリ ス定数(Km)、 最大速度(Vm(L'{)および触媒効率CVmax/Km)を求めた ところ、 いずれのイソフォームも同程度の親和性を示した。Vmaxは、
いず れの基質に対しでもQ3> Q2 > Qlであり、 触媒効率も、 Q3> Q2 > Qlで あった。参考文献
1) P. Andrew, Bíochem. J., 9 1, 222-223 (1964).
2) H. Svensson, Acta. Chem. Scand., 1 6,456-466 (1962).
m e
'n c
o m
h c k
a AU h cρ A
M Au m
γム勾b c .m 別
u u z (
0 3 k t y . 凶 a
人M n
・a-円\U
U qJ市n 紅引ω vh∞
m
4) K. Fujimoto, K. Masuda, N. Asada and E. Ohnishi, J. Biochem., 1 1 3,
285-291 (1993).
5)
A. Asp組and K.Sりderh札Insect. Bíochem., 2 1,363-373 (1991)
6) Y. Aso, K. J. Kr紅ner, T. L. Hopkins and G. L. Lコokhart, Insect Biochem. , 1 5, 9-17 (1985).
33
,.,-.. 1.2 20
〆-._ 〆圃""'
1.0
、国〆
g
'-"'ロ0.8
笥。令b3h
�
105 FJ回 2
-、� 0.6 1 5
開白聞則自ω
\
。
色 白岡
ω ω
冨\
\ \
\
何0.4
\ \�
\ \。� 5
回
4Sぞ 0.2
\ \
\
、。
\0.0
\。
。 100 200 300
Fractioo No. (2. 1 ml)
Fig. 3・ 1 Chromatography of crude extract 00
a Butyl Toyopearl column
Crude extract was put on a Butyl Toyopearl 65 0M (2.5 X 23.5 cm) equilibrated with 10 mM potassium phosphate buffer (pH 6.5) containinf;!
1 M ammonium sulfate and 1 mM PMSF. The el山n was carried out b J
a ilnear gradient of ammonium sulfate from 1toOM in the phosphate
bufTer.The fractions indicated by bar were combined and used for further
purification.
(111 )(£富田)。鳥島
20
15
10
5 0.8
0.6
0.4
0.2
,....、、
wg図。∞N芯ωω国史官。sd刊
200 0
ov ov ov
100
Fraction No. (2. 1 ml)
Fig.3・2 Gel filtration chromatography of PPO prepared at the Butyl Toyopearl step
The PPO preparation obtained by thc method described in the legend of Fig. 3・1 was put on a Sephacryl S-200 (2.2 X 140 cm) equilibrated with 10 mM potassium phosphate buffer (pH 6.5)
containing 0.5 M KCI. The fractions indicated by bar were combined and used for further purification.
35
(?lj) (お何回忌)。島内回一 25
20
15
10
,hd
200 0 0.6
0.2
00 00 00
0.4
〆ー、
)g図。∞N沼ωωロ祭τ。sd刊
100
Fraction No. (2.0 ml)
Fig.3・3 Rechromatography of PPO prepared at the gel filtration step
The PPO solution obtained by the method described in the legend of Fig. 3-2 was rechromatographed on the
Sephacryl column; the chromatography was done under the same conditions as those described in the legend. The
fractions indicated by bar were combined and applied to a
final chromatography.
�
J:に u 凶
。
.---
、ー〆
15十
10S
ロロ
。 A
zd A
1.0
o. 190
1.0
0.5 (ll) 自由 。∞N おωωロ.祭主。ωsd刊
210 200
。帥W ミd
100 200
Fraction No. (0.6 ml)
が勺 00
Figふ4 Anion圃exchange column chromatography of PPO prepared at the gel filtration step
The PPO preparation obtained by the method described in the legend of Fig. 3・3 was chromatographed on a Q
Sepharose column equilibrated with 10 mM potassium phosphate buffer (pH 6.5)・The elution was carried out by a Iinear gradient of KCI from 0 to 0.3 M in the same buffer.
(Inset) Fractions only containing an isoform were shaded. Ql,
Q2, and Q3 were corresponding to two fractions (No・200-201),
one fraction(No・210), and one fraction (No. 226), respectively.
37
1 2 3 4 5 6 7
(
一)
(+)
Fig.3・5 Native-PAGE ofPPO isoforms
PAGE was done on a 4.75 % separating gel, and the gel was submitted to activity (Iane 1・4) and silver (Iane 5・6) stainings. Lane 1; Lyophilized hemolymph (5 mg) was dissolved with 1 ml of 0.1 M potassium
phosphate buffer (pH 6.5), and 20pl of the solution was electrophoresed. An aliquots (total A280=2.0 ) of Ql, Q2 and Q3 described in the legend of Fig. 3-4 were
electrophoresed in lane 2 and 5 (Ql), lane 3 and 5 (Q2),
and lane 4 and 7 (Q3), respectively.
Table 3-1 Purification of PPO isoforms
Step Total protein
Crude extract
Butyl Toyopearl 650M Sephacryl S-200
Sephacryl S-200
Q-Sepharose Q1
Q2 Q3
(A280)
4520 121.0
34.00 22.90
0.20 0.15 0.21
Total
activi句r(units)
944 466 269 164
1.05
2.10 6.65
Specific activi勿
(U/A280)
0.210 3.87
7.91 7.14
5.20
14.5 31.4
Yield (0/0 )
100 49.4 28.5 17.3
0.11
0.22
0.70
1 2 3 M
(ー)
一-
97.4 kDa
ー-
66.0 kDa
ー-
45.0 kDa
(+)
Fig. 3-6 SDS・PAGE ofPPO isoforms
The purified isoforms (2.5 pl) were subjected to 7.5 %
SDS-PAGE. Lane 1, Ql; Lane 2, Q2; Lane 3, Q3; M,
mixture of phosphorylase b (92.5 kDa), bovine serum
albumin (66.0 kDa) and ovalubmin (45.0 kDa).
1α)()
Column: Supcrosc 12HR Buffer: 10 mM K-P04 (pH 6.5) • O.15MKCl
Catalase Q
g 1∞
お
2回起
。"-
2何 [ 122 kDa
10
22 24 26 28 30 32
Elution volume (ml)
Figふ7 Determination of molecular size of
Q3
by gel filtration
PPO and standard proteins were separately chromatographed on a Superose 12HR column. The standard proteins were catalase (245 kDa), aldolase
(158 kDa), BSA (66.2 kDa),。、叫bumin (45 kDa) and chymotrypsinogen A (25 kDa). Arrow and numerals indicate the elution position and estimated molecular size of Q3, respectively.41
1 2 3 4
(+) (pI) - 5.55 - 5.58
- 5.63
(
ー) Fig.3・8 Isoelectric focusing of PPO isoforms
Lane 1, Ql (0.4 pg); Lane 2, Q2 (0.4 pg); Lane 3, Q3 (0.2 pg);
Lane 4, mixture of Ql, Q2 and Q3
100
4 40 D且 H QO
10 12 5
0 5 0
2 7 5
2 (ぶ)kmwτ判明》吋
Fig.3・9 pH-Stability of PPO isoforms
PPO isoform was incubated at various pHs at 4 U C for 24 h. After activation, resulting PO activity was measured. Symbols: 0, Ql; .ð., Q2;
口, Q3 .
43
(A) 「一 100
_.-...
容75
'-'"を ω 吋 5 25 0
。
2 4 6 8 10
pH
(B)
_.-...
100
さ75 菅吋 ‘ω
伊 ー
通 5 0
2S
。
2 4 6 8 10
pH
Fig.3・10 Effects of pH on activation (A) and en勾Tme reaction (B)
Activation was carried out at various pHs and activity
was measured at pH 6.5. Symbols: 0, Ql;ム, Q2;口, Q3.
Table 3-2 Kinetic parameters of PPO for various substrates
L-DOPA Dopamine
N-Acetyldopamine
L-DOPA Dopamine
N-Acetyldopamine
L-DOPA Dopamine
N-Acetyldopamin�
Km (mM)
Ql Q2 Q3
2.08 1.47 4.71
2.64 1.15 1.94
0.99 1.82 1.32
V max (min -1 ・ A280・1 )
Ql 21 31 5
Ql 1.00 1.15 0.52
Q2
38 106
60
Vma1Km
Q2 2.53 8.95 3.20
Q3 161 271 61
Q3 3.31 13.54
4.46
Table み3 Properties of PPO isoforms
Properties Q1 Q2 Q3
Molecular size
(SDS-PAGE) 73kDa,74kDa 73kDa,74kDa 73kDa, 74kDa
(Gel filtration) 122kDa
Isoelectric point 5.55 5.63 5.58
pH Stability 6 - 10 6 - 10 6 - 10
Optimum pH
(Activation reaction) 6.5 6.5 6.5
(Enzyme reaction) 5.5 - 7.0 5.5 - 7.5 5.5 - 6.5 Specific activity
(U/protein) 5.2 14.5 31.4
Catalytic efficiency
Ql< Q2< Q3 (VmaxlKm)
一:
oot determioed
第四章 プロフエノールオキシダーゼcDNAのクローニングおよび塩基配列
第一節 緒言
カイコ変態過程におけるプロフエノールオキシダーゼ遺伝子の制御機精を 調べるためには、プロフエノールオキシダーゼをコードする遺伝子をクロー ニンク令することが不可欠である。 前章において、3種のPPOイソフォーム (Q1, Q2そしてQ3)は、いずれも73kDaおよび、74kDaのポリペプチドよ り構成されていることが明らかとなった。 そこで本章では、2種のポリペプ チドをコードする遺伝子をカイコ血球細胞よりクローニングし、その塩基配 列を決定することを目的とした。
第二節 実験材料 第一項 血球
カイコ(a80系統)5齢4日の体液をフェニルチオ尿素を含む遠心管中に氷 上で採取し、40C、3,000 rpmで15分間遠心分離を行い血球を沈殿させた。
沈殿した血球を氷冷した生理食境水で2回洗浄し、使用時まで-800Cにて保 存した。
第二項 プラスミド
Vieira1)らによって開発されたpUC18(宝酒造社製)を用いた。 このプラ スミドは、pBR322由来のアンピシリナーゼ遺伝子とDNAの複製開始点およ び大腸菌のlacZ'遺伝子から構成されており、lac領域には制限酵素認識部位 が連なったマルチクローニング部位があるため、種々の制限酵素DNA断片 が挿入できるという特徴を持っている。 また、pUC18をlac-大腸菌
(JMI09)に形質転換すると、コロニーの色の違いによりDNA断片の挿入 を判定できる。