ß -aminopropionitrile
投与による鶏胚骨格異常の解析日本大学大学院歯学研究科歯学専攻
久保田 桜
(指導: 磯川 桂太郎 教授,山崎 洋介 助教)
目 次
概 要
……… 2
緒 言
……… 5
材料および方法
……… 7
成 績
………10
考 察
………13
結 論
………16
謝 辞
………18
文 献
………19
図 表
………22
基幹論文
Kubota S, Yuguchi M, Yamazaki Y, Kanazawa H, Isokawa K.
Highly reproducible skeletal deformities induced by administration of ß -aminopropionitrile to developing chick embryos.
Journal of Oral Science, in press
概 要
胚組織の骨格要素は,発生の進行に応じて種々の形態変化を遂げる軟骨や 骨であり,その主要な構成成分は細胞外のコラーゲン線維である。コラーゲン 性の骨格原基は,将来の生体各部の骨性骨格の雛形をなすが,その物性や性状 にはコラーゲンの分子内および分子間の架橋が大きく関わっている。これらの 架橋形成を担うのがリシルオキシダーゼであり,一方,この酵素の活性を選択 的に阻害するのが
ß -aminopropionitrile (BAPN)である。BAPNは,スイートピ
ーLathyrus odoratusから抽出,同定された物質であるが,スイートピーの種 子を摂取させた家畜では,骨の形態異常,解離性大動脈瘤,皮膚の弛緩など,ラチリズムlathyrismとよばれる諸症状が現れることが知られている。BAPNは,
こうしたラチリズムの原因物質であると判明しており,その投与によって,コ ラーゲン原線維内で分子間凝集が攪乱され,骨性ラチリズムが生じることが明 らかになっている。
本研究では,鶏胚を実験動物として用い,卵殻に開窓して卵内で発生中の 発生日齢(ED)3-9日目の鶏胚にBAPNを投与した。その後,発生を一定期間継続 させ,ED5-10となった鶏胚を取り出して固定し,薄切法を用いず胚全体として の骨と軟骨の二重染色および骨・軟骨以外の組織については透明化処理を施す という方法を用いて,BAPNの投与で骨格要素に生じる異常の特徴,異常形態の 再現性,出現経緯を精査した。さらに,注目すべき変形が高い再現性をもって 観察された骨をアルカリ法によって胚組織から取り出し,その微細構造的な変 化について,走査型電子顕微鏡による観察を行った。
まず,有精卵へのBAPN投与量の妥当性を,鶏胚の生存率と骨格系異常の出 現状況から検討した。その結果,卵内で発生中のED4の鶏胚に350 µg/eggの
BAPNを投与した場合,ED10での生存率は約50%であり,骨および軟骨で構成さ
れる骨格要素の変形異常は生存胚のほぼすべてで観察されることが明らかにな った。軟骨性骨格要素を青染,骨化した骨格要素を赤染させる二重染色による観察で,胚内の複数箇所の骨格要素に変形が認められ,それらには部位特異性 があることが判明した。すなわち,外篩骨,下顎,椎骨,椎肋骨あるいは後肢 の骨格要素(脛足根骨および足根中足骨)などで骨変形がみられ,前肢の骨格要 素,骨盤,大腿骨などでは変形はみられなかった。これら多数の変形の中で,
骨幹中央部が伸筋側に突出する特徴的な屈曲変形が高い再現性をもって出現す る後肢の脛足根骨tibiotarsusに,以後の解析の焦点をあてることにした。
脛足根骨の屈曲変形は,ED4でのBAPN投与後,経日的にED10までその屈曲度 が増大した。ED5-8に至るどの日齢での投与によっても屈曲変形は惹起された が,ED8での投与では,脛足根骨の近遠心にある骨幹端2箇所に屈曲が生じた。
これは非常に特異な所見であり,ED7以前に投与されたBAPNが惹起した屈曲変 形とは形態的に明らかな差異がある。本研究の知見からは,ED8は骨幹中央部 での屈曲変形が生じる条件が既に終了した時期だと考えられた。実際,ED9で
BAPN投与を受けた鶏胚では,ED8でみられた骨幹端2箇所も含めて,脛足根骨の
屈曲変形はもはや出現しなかった。脛足根骨の屈曲変形についての走査電顕的な観察では,屈曲変形の凹部(屈 筋側)に線維層板骨fibrolamellar boneの過形成が認められ,同部には活発な 骨基質形成を行う骨芽細胞の存在を示唆する多数の骨小窩が観察された。線維 層板骨は,BAPN非投与の屈曲変形のない脛足根骨では骨幹周囲を均等に覆って いるが,BAPN投与によって屈曲変形をきたした脛足根骨ではその分布に偏りが あり,とくに屈曲変形の凹部では過形成状態にあることが特徴的であった。一 方,屈曲変形の凸部(伸筋側)の骨面は平滑で線維層板骨がみられず,骨小窩も 乏しかった。また,BAPN投与胚の脛足根骨の内面(骨髄腔側)には,破骨細胞性 の明瞭な吸収窩が多数みられたが,非投与胚での屈曲をみない脛足根骨の内面 では,破骨細胞性吸収窩は不明瞭で数もわずかであった。
以上の結果は,BAPNの投与による骨格要素の変形異常とくに脛足根骨でみ られる屈曲変形は,高い形態的再現性をもって出現し,発生段階や部位的にも
格原基の強度減弱には,骨改造におけるバランスの異常も影響している可能性 があると示唆された。
な お , 本 論 文 は ,
Kubota S, Yuguchi M, Yamazaki Y, Kanazawa H, Isokawa K. Highly reproducible skeletal deformities induced by administration of ß -aminopropionitrile to developing chick embryos.
Journal of Oral Science, in press
を 基 幹 論 文 と し , さ ら にß -
aminopropionitrileの投与量と鶏胚生存率との関係についての新たなデータを
加えることによって総括している。緒 言
胚組織の骨格要素は,発生の進行に応じて種々の形態変化を来す軟骨や骨 であり,その主要な構成成分は細胞外のコラーゲン線維系である。これらのコ ラーゲン性の骨格原基は,個体発生の過程で直接あるいは軟骨を介しての骨化 を遂げて,将来の生体各部の骨性骨格構造に移行する(1,2)。部位によって は,軟骨性原基の一部が骨化することなく,例えば関節軟骨などとして成体の 骨格構造の一部となる場合もある。したがって,成体の骨格構造の雛形をなす コラーゲン性原基の質的不良やそれを惹起する形成不全は,充分な強度をもつ 骨格構造が将来形成される上での深刻な障害となる。雛形となるコラーゲン性 の骨格原基に求められる物性や性状には,コラーゲンの分子内および分子間の 架橋が大きく関わっている(3-5)。この架橋構造の形成には,アミンオキシダ ーゼの一種であるリシルオキシダーゼ(LOX, EC1.4.3.13)が関わる。すなわ ち,コラーゲンのペプチド鎖中のリシン残基を
LOX
がアリシンへ変換するた め,生じたアリシン2
残基間で,共有結合性のコラーゲン分子内あるいは分子 間の架橋構造が形成される(6,7)。一方,スイートピーLathyrus odoratus の種子を摂取させた家畜で,骨の 形態異常,解離性大動脈瘤,皮膚の弛緩などが生じることは古くから知られて おり,それらの諸症状はラチリズム
lathyrism
とよばれていた。その後,スイ ートピーの種子から抽出されたß -aminopropionitrile (BAPN)がラチリズムの
原因物質lathyrogen
であると同定され(8,9),実際,このBAPN
の投与によっ て骨の形態異常も引き起こされることが示された(9-11)。症状の現れ方によっ て,ラチリズムは神経ラチリズムと骨性ラチリズムに大別されるが(11,12),ニワトリおよびその胚ではどちらのタイプのラチリズムもスイートピー摂取で 生じるが,マウスやラットでは骨性ラチリズムが生じないとの報告がある
(12)。また,当初,諸説が入り乱れた BAPN
の作用対象についても,研究者のてコラーゲン原線維内でコラーゲンの分子間凝集が攪乱されることを明らかに した。
本研究では,卵内で発生を継続させることができる鶏胚を実験動物とし,
薄切法を用いず胚全体として骨と軟骨の二重染色をし,同時に他の胚組織を透 明化することで(14, 15),骨格要素の可視化と出現する形態異常の
3
次元的な 観察を行った。BAPN の投与量と胚の生存率の関係を検討した後,投与量を一定 にした実験で,胚内各部での骨格系の形態異常をマッピングした。続いて,脛足根骨
tibiotarsus
に注目し,同部の形態異常の再現性および形成過程をBAPN
の投与時期,作用期間の観点から経日的に精査した。さらに,特徴的な骨変形 が生じる脛足根骨の微細構造的な変化を走査型電子顕微鏡(SEM)で検討した。
材料および方法
1. 鶏卵の開窓とインキュベート
白色レグホン
Gallus gallus の有精卵(約 70 g, 大畑シェーバー孵化場)を
温度勾配恒温器(EYELA MTI-201A; 東京理科)を用いて39℃でインキュベートし
た。インキュベート開始後3-9
日目に,卵殻にサイズ約10 × 15 mm
の開窓を 施し(16),さらにピンセットを用いて,開窓部の直下の卵殻膜と卵黄膜を除去 した。開窓部からの実体顕微鏡(SZ61; オリンパス)による観察で異常の認めら れなかった発生中の鶏胚に,一卵あたり100 µl
のBAPN
溶液あるいは等量の滅 菌水を投与し,後者は対照群とした。卵内の乾燥や胚の脱水防止のため,開窓 部は透明なプラスチックテープで封鎖し,発生日齢(ED; embryonic days)4-10 日に至るまで,さらにインキュベートを継続した。なお,ED4-10 の鶏胚は,Hamburger
とHamilton (17)の形態的基準によれば発生ステージ 24-36
に相当 する。2. BAPN
投与量の検討有精卵一つあたりの
BAPN
投与量をとくに骨格要素の形態異常の出現と胚の 生存率の観点から検討するために,12 段階のBAPN
溶液,すなわち0.25, 0.5, 1.0, 2.5, 3.0, 3.5, 4.0, 4.5, 5.0, 12.5, 25.0
および50.0 mg/ml
を滅菌水 で調製した。ED4の鶏胚(n = 62)に開窓を施し,12段階いずれかの濃度のBAPN
溶液
100 µl
を投与し,恒温器内でED10
まで発生を継続させた。3. BAPN
投与のスケジュール前項に記載の検討で得られた結果から,一卵あたりの
BAPN
量を350 µg
(3.5 mg/ml
溶液を100µl
投与)とし,次の3
系統の実験を行った。いずれの場 合も,対照群とする鶏胚には等量の滅菌水を投与した。第
1
の系統の実験(Fig. 1A)では,ED4 の鶏胚にBAPN
を投与し,発生を継 続させてED10
で生存率を求めた。生存胚については,固定後にwhole mount
の骨・軟骨二重染色(n = 14)を施し,骨格要素の形態異常に関しての評価を行 った。また,ED10での生存胚の一部(n = 7)はSEM
観察に供した。第
2
の系統の実験(Fig. 1B)では,ED3-9 の鶏胚にBAPN
を投与し,発生を 継続させてED10
で生存率を求めた。生存胚(n = 83)については,固定後にwhole mount
の骨・軟骨二重染色を施し,投与時期の差異にもとづく骨格要素の形態異常の観察を行った。
第
3
の系統の実験(Fig. 1C)では,ED4 の鶏胚にBAPN
を投与し,発生を継続させて
ED5-10
で生存率を求めた。生存胚(n = 120)については,固定後にwhole mount
の骨・軟骨二重染色を施し,作用期間の差異にもとづく骨格要素の形態異常の観察を行った。
4. whole mount
骨・軟骨二重染色全胚まるごとの骨・軟骨二重染色は,Kellyと
Bryden (14)によるプロトコ
ールを基本にYamazaki
ら(15)による改変を加えて実施した。すなわち,観 察,評価を行う日齢にまで発生した鶏胚を,まず,リン酸緩衝生理食塩水(pH7.35; PBS)で調製した 4%パラフォルムアルデヒドに 2
時間,4℃で浸漬し,固定 を 行 っ た 。 続 い て , 固 定 液 を
PBS
で 洗 浄 し た 鶏 胚 を ア ル シ ア ン 青(AB;
alcian blue 8GX, C.I.74240; 和光純薬)の染色液に一晩浸漬して軟骨の染色
を行った。AB 染色液は,95%エタノール80 ml
と99.7%酢酸 20 ml
を混和した溶液に
AB
を10 mg
溶解させて調製した。AB 染色後の鶏胚は,直ちに1%水
酸化カリウム(KOH)-70%エタノール溶液に移し,そのまま一晩浸漬して,AB 染 色によって胚内に生じていた低
pH
環境の中和を図った。その後,下降エタノ ー ル 系 列 を 通 し ,0.002
%(w/v)
ア リ ザ リ ン 赤(AR; alizarin red S,
C.I.58005; 和光純薬)の染色液に 24
時間浸漬して骨の染色を行った。AR 染色後の鶏胚は, 2%KOH溶液に室温下で
4
時間浸漬して浸軟処理を施し,その後は0.5%KOH
とグリセロールの3:1, 1:1
そして1:3
の混和溶液に順に8-24
時間ず つ浸漬して組織の透徹を行った。二重染色が完了した鶏胚は,50%グリセロー ル中に静置して,デジタルカメラ(Digital SIGHT DS-5M-L1; ニコン)を備えた 実体顕微鏡で観察,記録を行った。5. 変形骨の単離と SEM
観察第
1
の系統の実験でBAPN
の投与を受けた鶏胚(Ad4-Ev10, Fig. 1A)をPBS
で調製した2.5%グルタールアルデヒドに 2
時間,4℃で浸漬し,固定を行っ た。固定後の鶏胚試料を37℃に加温した 4N KOH
に1-3
日間浸漬して軟組織を すべて溶解させ,骨性の骨格要素を取り出した。BAPN 投与群の鶏胚から単離さ れた後脚部の変形した骨性骨格要素と,対照群の相当する骨性骨格要素とを併 置して実体顕微鏡下で比較観察した。続いて,変形した脛足根骨と対照群の脛 足根骨を上昇エタノール系列で脱水し,臨界点乾燥器(HCP-2; Hitachi)で処理 した。得られた乾燥試料に四酸化オスミウムで20
秒間のコーティング(HPC-1S;
真 空 デ バ イ ス 社)
を 施 し た 後 , 電 界 放 射 型 走 査 電 子 顕 微 鏡(S-4300;
Hitachi)を用いて,加速電圧 15 kV
で観察,記録を行った。成 績
有精卵への
BAPN
投与量の妥当性を,鶏胚の生存率(Fig. 2)と骨格系異常の 出現状況から検討した結果,100 µg/egg 以下の投与量(1 mg/ml 以下のBAPN
溶 液で100 µl)では骨格系異常が出現せず,400 µg/egg
以上の投与量(4 mg/ml 以 上の溶液で100 µl)ではほぼ全例が致死になると判明した。一方, 250-350 µg/egg の投与(2.5-3.5 mg/ml
の溶液で100 µl)を受けた鶏胚は,生存率が 50%以上で,骨格系の異常も高い頻度で観察された。このため,以降の本研究
の実験では,有精卵一つあたりのBAPN
投与量を350 µg
とした。ED4
でBAPN
投与を受けたED10
の鶏胚では,胚内の複数箇所の骨格要素(AB で青染する軟骨性要素およびAR
で赤染する骨化した骨格要素)に変形が観察さ れた。実体顕微鏡下で認知し易い変形は,後肢の軛脚部zeugopodial segment
および下顎部に認められた(Fig. 3)。顕微鏡下での精査の結果,軛脚部の脛足 根骨に最も著しい変形が観察され,それは骨幹中央部が伸筋側に突出する顕著 な屈曲変形であった。腓骨fibula
や足根中足骨tarsometatarsals
も軽度ある いは中等度の屈曲変形を示したが,大腿骨femur
や趾骨phalanges
には変形が 認められなかった(Fig. 4A, B)。下顎部においては(Fig. 4C-F),メッケル軟 骨に捻じれをともなった重度の屈曲変形が両側性に観察されたが,隣接部に発 生中の下顎骨には明確な異常は認められなかった。観察された後肢骨格要素,下顎部およびその他の部位における変形を
BAPN
投与鶏胚14
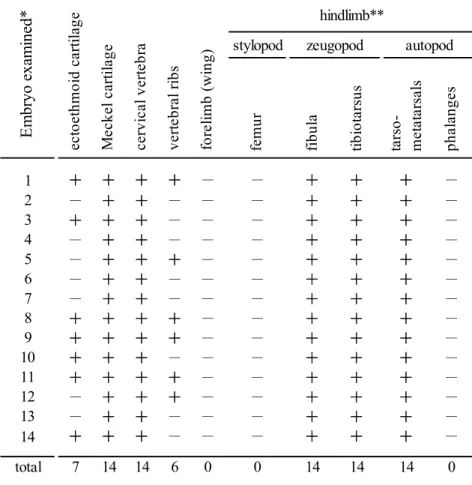
例で評価した(Table 1)。後肢の軛脚部,足根中足骨および下顎部 のメッケル軟骨の変形は,出現の頻度および異常の形態的再現性が非常に高か った。頸椎の軟骨性原基における変形(図示せず)もすべての生存胚で観察され た。しかし,翼となる前肢の骨格要素,骨盤,大腿骨では変形はみられなかっ た。外篩骨ectoethmoid (Fig. 4E, F)および椎肋骨 vertebral ribs (Fig.
4G, H)においては,特徴的な変形が観察されたが,これらの出現頻度は 40-
50%程度であった。
次に
BAPN
投与の時期をED3-9
に変化させたED10
の鶏胚(Fig. 1B; 生存率 をFig. 5
に示す)において,脛足根骨に生じた変形を精査した。ED3でBAPN
投 与を受けた鶏胚は全例死亡であった。ED4-7でBAPN
投与を受けた鶏胚の生存例 ではすべて,脛足根骨の骨幹中央部が伸筋側にアーチ状に突出した屈曲変形を 示した(Fig. 6A-D)。対照的に,ED8 でBAPN
投与を受けた鶏胚では,脛足根骨 の近遠心の骨幹端すなわち2箇所に屈曲変形が観察された(Fig. 6E)。2箇所 に屈曲が現れる特徴的な変形は,ED8でのBAPN
投与胚に特異的でその再現性も 高かった。一方,ED9でBAPN
投与を受けた鶏胚では,脛足根骨に変形は生じな かった(Fig. 6F)。ED4
でのBAPN
投与後,発生をED5-10
まで継続させた鶏胚(Fig. 1C; 生存率を
Fig. 7
に示す)において,脛足根骨に生じた変形を精査した。ED5 の鶏胚では,軟骨性骨格原基の存在を示す
AB
陽性像が後肢軛脚部に未だ現れなかっ た (図示せず)。ED6-9 の鶏胚でそれぞれの脛足根骨が示す伸筋側へのアーチ状 の屈曲変形(Fig. 8)は,ED6-7 では未だ軽度であったが,ED8-9 では顕著とな り, ED10 で観察される屈曲変形(Figs 3A, 4A および6A)の状態に至ることが
判明した。ED4
でBAPN
投与を受けたED10
の鶏胚(Figs 3A, 4A および6A
に相当)の後 肢から単離した骨性骨格要素とBAPN
非投与の対照群からの骨性骨格要素を実 体顕微鏡下で比較観察した(Fig. 9)。いずれの骨格要素も中空性の骨殻bone
collar
とその表面に綿毛状構造が観察された。綿毛状構造は,大腿骨ではほぼその全表面に,脛足根骨では中央
2/3
に,足根中足骨では中央1/3
にみられ,腓骨では観察されなかった。BAPN 非投与群と比べて投与群で特異であった点 は,脛足根骨と足根中足骨の屈曲変形と,これらの屈曲変形で生じた屈筋側凹 部での綿毛状構造の集積であった。光学顕微鏡下では白く不透明にみえる綿毛 状構造は,脛足根骨の屈曲変形の凹部に豊富で,足根中足骨の屈曲変形の凹部 では量的なばらつきはみられるが少量であった。単離された腓骨の形態は,
の腓骨とは異なっていた。この差異は,BAPN 投与群での腓骨形成が,投与群よ りもやや遷延していることを示す。
脛足根骨の屈曲変形凹部の綿毛状構造は,SEM 観察の結果,BAPN 非投与群 の 脛 足 根 骨 等 で も そ の 表 面 を 覆 っ て い る 綿 毛 状 構 造 と 同 様 な 線 維 層 板 骨
fibrolamellar bone
であることが判明した(Fig. 10)。言い換えれば,屈曲変 形の凹部では,そうした多孔性の線維層板骨の過形成が生じていることにな る。脛足根骨の屈曲変形の凸面側(伸筋側)では,骨面は比較的平滑であり,
BAPN
非投与群の脛足根骨であれば存在しているはずの骨面表層の線維層板骨が 欠如していた。線維層板骨を欠く屈曲変形の凸面には骨小窩が乏しく,この部 位においては骨膜最深層に位置する骨芽細胞数が乏しいことを反映すると考え られた。対照的に,屈曲変形の凹部(屈筋側)では,線維層板骨の過形成部の表 面に多数の骨小窩が観察された。BAPN
投与を受けた鶏胚の脛足根骨の内表面は,非投与の対照群とは明らか な差異が認められた(Fig. 11)。投与群の脛足根骨内表面には,活性型の破骨 細胞の存在を示す多数の境界明瞭なハウシップ窩がみられた。一方,対照群の 脛足根骨内表面には,浅くサイズも小さい吸収窩様の構造がわずかに散在する のみで,それら吸収窩様の構造と周囲骨面との境界も不明瞭であった。考 察
本研究で得られた知見は,BAPN の投与による骨格要素の変形が発生段階や 部位的に特異な様式で生じることを示している。BAPN を投与された鶏胚では,
屈曲あるいは捻じれなどの変形が,外篩骨,下顎,椎骨,肋骨あるいは後肢骨 格などの特定の骨格要素において生じたが,前肢を含むその他の骨格要素では 異常はとくに観察されなかった。観察された骨格系の変形の中で,メッケル軟 骨の変形は形態的に独特で甚だしかった。しかし,下顎骨については,メッケ ル軟骨とは独立に膜内骨発生で生じることが知られ,その異常や経日的変化が 本研究で扱った発生期間内では顕著でなかった。一方,脛足根骨でみられた屈 曲変形は,それが発生段階や部位的に特異な変形であることが捉え易い状況で あった。部位的な特異性は,2つの点から明瞭であった。まずは,屈曲変形 が,体幹から趾骨へ至る後肢近遠心軸に沿う一連の骨格要素の中で,軛脚部の 骨格(とくに脛足根骨)と足根中足骨に特異的に生じた点である。もうひとつ は,屈曲変形が軛脚部の骨格および足根中足骨それぞれの骨幹中央部で特異的 に生じた点である。これらの変形の部位的出現パターンは,明らかにこれらの 骨格要素の形成時期に関わる時間的出現パターンと関連している。
後肢の骨格系は,体幹部の骨盤から後肢遠位端に位置する趾骨へ,発生プ ログラムが規定する順に従って連続的に形成が進行する(1, 2, 18)。このた め,脛足根骨に特異的に出現した屈曲変形は,この異常が比較的狭い時間的な ウィンドウの中で生じることを示唆している。ウィンドウ枠の開始点は,軟骨 性原基の可視化を本研究では
AB
染色性に依拠しているため,明確にはできな かった。つまり,脛足根骨の原基はED6
以降でAB
陽性を示すと考えられるが(19-21),発生プログラムが間葉中で脛足根骨の原基を規定するのはそれより
も早いのである(1)。一方,ウィンドウ枠の終了点はED8
だと考えられる。な ぜならば,ED8でBAPN
投与を受けた鶏胚で生じる脛足根骨の変形は独特で,近 遠心の2箇所にある骨幹端で屈曲が生じ(Fig. 6E),それ以前の時期に投与さ生じる屈曲変形については植松による研究報告(22)にも記載を認めるが,その 解釈はなされていない。本研究の知見からは,ED8 は骨幹中央部での屈曲変形 が生じる条件が終了した時期だと考えることができる。実際,ED9で
BAPN
投与 を受けた鶏胚では,ED8 でみられた骨幹端2
箇所も含めて,脛足根骨の屈曲変 形はもはや出現しなかった(Fig. 6F)。本研究の結果に関しての前段までの解釈や説明は,BAPN 投与によるコラー ゲン分子の架橋構造の減少が骨格要素の強度(硬さ
stiffness)を 25%以上減弱
させるとの研究報告(3-5)に基づいている。この意味においては,軟骨および 骨のいずれもが,特定の発生段階において協調的あるいはそれぞれに独立し て,骨格系の異常形成に関わっているといえる。鶏胚後肢における長管骨形成 の過程はこれまでにもその詳細が報告されている(2, 23-25)。すなわち,個々 の骨格要素の原基としての桿状の軟骨がまず形成され,その中央部から,桿状 の軟骨の表面を覆うように鞘状(円筒状)に骨形成が進行する。後者は骨殻ある いは骨性シリンダーの形成と表現される。この骨殻で覆われた内部では,やは りその中央部から骨端側へと軟骨組織の侵食が進行する(2, 23, 25)。これら 一連の知見は,脛足根骨が当初桿状の軟骨として存在し,その後,軟骨とそれ を包む骨殻という二重構造を示し,最終的にはシリンダー状のみになることを 意味する。後肢骨格について,BAPN の投与による機械的強度の減弱と骨変形と の因果関係を考える場合,これら骨格要素の3つの様態およびそれらの移行期 の様態を考慮しなければならないであろう。骨性骨格要素では,骨の改造現象がその機械的強度に影響を及ぼす可能性 も高い。本研究では,BAPN を投与した鶏胚の脛足根骨の内面(髄腔面)に,非投 与の対照群よりも境界明瞭で多くのハウシップ窩が分布することを認めた。こ れは,破骨細胞数やその活性が増大していることを示している。一方,発生中 の骨殻の外表面には多数の骨小窩が観察され,骨基質形成中の骨芽細胞が石灰 化した骨表面に埋入されようとしている状態と考えられた。そうした骨小窩の 数は,屈曲変形の凸面(伸筋側)で乏しく,凹部(屈筋側)で豊富であった。こう
した所見は,凹部を覆う骨膜の骨芽細胞では基質形成が活発なために凹部の線 維層板骨が過形成状態に至っていること,また,凹部骨面に豊富な骨小窩は,
急速で多量の基質産生を担った骨芽細胞が基質に埋入されて骨細胞へ移行する 像だと考えられる。
本研究では,発生中の筋が生む力が骨の変形を増悪させる可能性について 実験的な検証せず考察もしていない。しかし,外界から遮断された静穏な卵内 においては,発生中の微小な筋であっても,BAPN によって機械的強度が低下し た骨の変形を惹起する可能性はある。鶏胚における筋発生と成長の時期(26),
卵内の筋における収縮活動の亢進時期(27)などが,本研究で観察された骨変形 の出現タイミングと一定の整合性を示す可能性がある。
結 論
卵内の鶏胚にBAPNを投与した後に発生を一定期間継続させ,骨格要素に生 じる異常の特徴,異常形態の再現性,出現経緯,微細構造を精査したところ,
以下の結果と結論を得た。
1.
卵内で発生中のED4の鶏胚に350 µg/eggのBAPNを投与した場合,ED10での生 存率は約50%であり,骨および軟骨で構成される骨格要素の変形異常は生 存胚のすべてで観察された。2.
変形異常の発生には部位特異性があり,外篩骨,下顎,椎骨,椎肋骨ある いは後肢の骨格要素(脛足根骨および足根中足骨)などで観察され,前肢の 骨格要素,骨盤,大腿骨などではみられなかった。3.
脛足根骨では,骨幹中央部が伸筋側に突出する特徴的な屈曲変形が高い再 現性をもって出現した。ED4でのBAPN投与後,屈曲度は経日的にED10まで増 大した。ED5-8での投与によっても屈曲変形は生じた。但し,ED8での投与 では,脛足根骨の近遠心にある骨幹端2箇所に屈曲が生じた。ED9での投与 では屈曲変形は生じなかった。4.
脛足根骨の屈曲変形の凹部には線維層板骨の過形成が認められ,同部には 活発な骨基質形成を行う骨芽細胞の存在を示唆する多数の骨小窩が観察さ れた。屈曲変形の凸部の骨面は平滑で線維層板骨はみられず,骨小窩も乏 しかった。5. BAPN投与胚の脛足根骨内面には,破骨細胞性の明瞭な吸収窩が多数みられ
たが,非投与胚の屈曲をみない脛足根骨内面では,破骨細胞性吸収窩は不 明瞭で数もわずかであった。以上の結果は,BAPN の投与による骨格要素の変形異常とくに脛足根骨でみ られる屈曲変形は,高い形態的再現性をもって出現し,発生段階や部位的にも 特異性がみられることを示している。また,骨変形の基盤である
BAPN
による骨格原基の強度減弱には,骨改造におけるバランスの異常も影響している可能 性が示唆された。
謝 辞
本研究の遂行にあたり,数々のご協力とご助言を賜りました本学部解剖学 第Ⅱ講座 磯川桂太郎教授および同講座の皆さまに謹んで心より感謝申し上げ ます。
文 献
1. Isokawa K, Krug EL, Fallon JF, Markwald RR (1992) Leg bud mesoderm retains morphogenetic potential to express limb-like
characteristics (“limbness”) in collagen gel culture. Dev Dyn 193, 314-324
2. Namba Y, Yamazaki Y, Yuguchi M, Kameoka S, Usami S, Honda K,
Isokawa K (2010) Development of the tarsometatarsal skeleton by the lateral fusion of three cylindrical periosteal bones in the chick embryo (Gallus gallus). Anat Rec 293, 1527-1535
3. Oxlund H, Barckman M, Ortoft G, Andreassen TT (1995) Reduced
concentrations of collagen cross-links are associated with reduced strength of bone. Bone 17, 365S-371S
4. Viguet-Carrin S, Garnero P, Delmas PD (2006) The role of collagen in bone strength. Osteoporos Int 17, 319-336
5. Saito M, Marumo K (2010) Collagen cross-links as a determinant of bone quality: a possible explanation for bone fragility in aging, osteoporosis, and diabetes mellitus. Osteoporos Int 21, 195-214 6. Eyre DR, Paz MA, Gallop PM (1984) Cross-linking in collagen and
elastin. Annu Rev Biochem 53, 717-748
7. Kagan HM (1986) Characterization and regulation of lysyl oxidase.
In Regulation of Matrix Accumulation, Mecham RP ed, Academic Press, New York, 321-398
8. Schilling ED, Strong FM (1954) Isolation, structure and synthesis of a lathyrus factor from L. Odoratus. J Am Chem Soc 76, 2848 9. Dasler W (1954) Isolation of toxic crystals from sweet peas
(Lathyrus odoratus). Science 120, 307-308
10. Wawzonek S, Ponseti IV, Shepard RS, Wiedenmann LG (1955) Epiphyseal plate lesions, degenerative arthritis, and dissecting aneurysm of the aorta produced by aminonitriles. Science 121, 63-65
11. Selye H (1957) Lathyrism. Rev Can Biol 16, 1-82
12. Belanger LF (1959) Observations on the manifestations of
osteolathyrism in the chick. J Bone Joint Surg Br 41B, 581-589
ß -aminopropionitrile (lathyrus factor). J Exp Med 110, 771-790 14. Kelly WL, Bryden MM (1983) A modified differential stain for
cartilage and bone in whole mount preparations of mammalian fetuses and small vertebrates. Stain Technol 58, 131-134
15. Yamazaki Y, Yuguchi M, Kubota S, Isokawa K (2011) Whole-mount bone and cartilage staining of chick embryos with minimal
decalcification. Biotech Histochem 86, 351-358
16. Korn MJ, Cramer KS (2007) Windowing chicken eggs for developmental studies. J Vis Exp 8, 306
17. Hamburger V, Hamilton HL (1951) A series of normal stages in the development of the chick embryo. J Morphol 88, 49-92
18. Rowe DA, Fallon JF (1982) The proximodistal determination of skeletal parts in the developing chick leg. J Embryol Exp Morphol 68, 1-7
19. Linsenmayer TF, Toole BP, Trelstad RL (1973) Temporal and spatial transitions in collagen types during embryonic chick limb
development. Dev Biol 35, 232-239
20. Thorogood PV, Hinchliffe JR (1975) An analysis of the condensation process during chondrogenesis in the embryonic chick hind limb. J Embryol Exp Morphol 33, 581-606
21. Yamazaki Y, Mikami Y, Yuguchi M, Namba Y, Isokawa K (2012)
Development of collagen fibres and lysyl oxidase expression in the presumptive dermis of chick limb bud. Anat Histol Embryol 41, 68-74 22.
植松 健 (1965) 軟骨の変性および老化に関する研究. その3
ß-aminopropionitrile
の鶏胚長管骨に対する影響について. 横浜医学 16,1-16
23. Caplan AI, Pechak DG (1987) The cellular and molecular embryology of bone formation. In Bone and Mineral Research 5, 1987, Peck WA ed, Elsevier Science Publishers B.V. , Amsterdam, 117–183
24. Bruder SP, Caplan AI (1989) First bone formation and the dissection of an osteogenic lineage in the embryonic chick tibia is revealed by monoclonal antibodies against osteoblasts. Bone 10, 359-375 25.
難波 祐一 (2010) 形成過程の長管骨骨殻の破骨細胞性穿孔について. 日大歯学 84, 41-49.
26. Kardon G (1998) Muscle and tendon morphogenesis in the avian hind limb. Development 125, 4019-4032
27. Mikic B, Isenstein AL, Chhabra A (2004) Mechanical modulation of cartilage structure and function during embryogenesis in the chick.
Ann Biomed Eng 32, 18-25
Table 1. The observed skeletal deformities induced by BAPN
stylopod
femur fibula tibiotarsus tarso- metatarsals phalanges
1 + + + + - - + + + -
2 - + + - - - + + + -
3 + + + - - - + + + -
4 - + + - - - + + + -
5 - + + + - - + + + -
6 - + + - - - + + + -
7 - + + - - - + + + -
8 + + + + - - + + + -
9 + + + + - - + + + -
10 + + + - - - + + + -
11 + + + + - - + + + -
12 - + + + - - + + + -
13 - + + - - - + + + -
14 + + + - - - + + + -
total 7 14 14 6 0 0 14 14 14 0
** Hindlimb skeleton distal to girdle consists sequentially of 3 segments; stylopodium (femur), zygopodia (fibula and tibiotarsus) and autopodia (3 tarsometatarsals and more distal phalanges).
Em br yo e xa m ine d*
forelimb (wing)* Embryos were treated by BAPN as in Fig.1A
hindlimb**zeugopod autopod
ectoethmoid cartilage Meckel cartilage cervical vertebra vertebral ribs
Fig. 1
Dosage schedule of BAPN. Three sets of experiments (A, B, C) were performed in this study. The day of BAPN administration is denoted by a closed circle or
‘Ad’ plus ‘embryonic day’, and the day of evaluation (based on specimens stained doubly with AB and AR) is by an open square or ‘Ev’ plus ‘embryonic day’. Therefore, ‘Ad4-Ev10’ stands for a schedule in which BAPN was administered at ED4 and the embryo evaluated at ED10. Please note that ‘Ad4- Ev10’ in parenthesis in B) and C) is basically the same schedule with A).
3 4 5 6 7 8 9 10
Embryonic Days Ad4-Ev10
Ad4-Ev5 Ad4-Ev6 Ad4-Ev7 Ad4-Ev8 Ad4-Ev9 Ad3-Ev10 (Ad4-Ev10)
Ad5-Ev10 Ad6-Ev10 Ad7-Ev10 Ad8-Ev10 Ad9-Ev10
(Ad4-Ev10) A)
B)
C)