富山大学大学院医学薬学研究部産科婦人科
総 説
制御性 T 細胞,制御性 NK 細胞からみた妊娠維持機構
齋 藤 滋,島 友 子,中 島 彰 俊
Regulatory T cells and regulatory NK cells play essential roles for maintenance of pregnancy
Shigeru SAITO, Tomoko SHIMAand Akitoshi NAKASHIMA Department of Obstetrics and Gynecology, University of Toyama
(Received August 10, 2012) summary
During pregnancy, semiallograftic fetus is allowed to grow without being rejected by the maternal immune sys-tem. Recent data show that paternal antigen speciˆc- or male antigen HY speciˆc-regulatory T cells (Treg) increase during pregnancy. Seminal plasma is necessary for the accumulation of Treg cells in uterine draining lymph modes just before implantation, and in uterus just after implantation. It has been reported that decreased number of Treg cells in peripheral blood or pregnant uterus in abortion or preeclampsia. In human and rodent pregnant uterus, NK cells are the major population (~80) of lymyhocytes, we have reported that CD25+NK cells increase in mouse pregnant
ute-rus, and these NK cells produce immunoregulatory cytokines such as IL10 and TGFb. They inhibit the expression of MHC class II antigen on dendritic cells and inhibit the cytotoxic T cell induction, suggesting that these NK cells have an ability for immunoregulation. Both Treg cells and regulatory NK cells play important roles for maintenance of pregnancy.
Key words―allogeneic pregnancy; CD25+NK cells; regulatory T cells; regulatory NK cells; seminal plasma
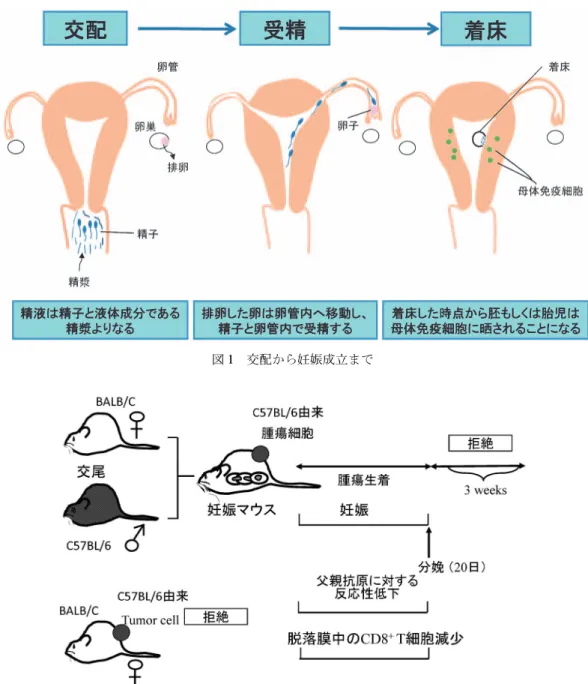
抄 録 Semiallograftである胎児を許容するために,妊娠時には父親抗原に対するトレランスが存在することが知られて いたが,その詳細な免疫学的機構は明らかではなかった.最近の研究により,妊娠時には父親抗原特異的もしくは 男性抗原である HY 抗原特異的制御性 T 細胞が増加していること,精漿のプライミングが父親抗原特異的制御性 T 細胞の誘導に重要であることが判明している.またヒトならびにマウスの流産や,ヒトでの妊娠高血圧腎症では 末梢血ならびに,妊娠子宮での制御性 T 細胞の減少が報告されている.妊娠初期子宮内膜では,特殊な NK 細胞 がリンパ球の主要な成分(約 80)を占める.我々は,マウス妊娠子宮では CD25+NK細胞が増加すること, CD25+NK細胞は IL10 や TGFb を産生すること,本 NK 細胞は樹状細胞上の MHC class II 抗原発現を抑制さ せ,細胞傷害性 T 細胞の誘導を抑制し,制御性 NK 細胞と呼べる性状を有することを見い出した. このように妊娠初期において,制御性 T 細胞と制御性 NK 細胞は協同的に作用し,胎児を母体免疫系の攻撃か ら守っている. 諸 言 妊娠は天与の移植成功例といえる.異物である胎 児が,妊娠子宮内で母体免疫細胞に攻撃されずにい ることは神秘的であるが,胎児許容機構がうまく働 かないと,流産や妊娠高血圧腎症(妊娠後半期に高 血圧と蛋白尿を呈する母子共に予後不良な疾患)が 生じる可能性も指摘されている. 腟内で射精された精子は子宮内に入り,さらに卵 管に辿り着き,卵と受精する(図 1).受精卵は胚 となり透明帯から孵化し(hatching),子宮内膜に 接着し,やがて子宮内に浸潤し(着床),胎盤を形 成する.子宮内は決して免疫学的に隔絶された臓器 ではなく,母体由来のリンパ球が多数存在する.し たがって,着床後より,胚もしくは胎児は,母体免 疫細胞に晒されることになる(図 1).では,どの ような機序で胎児は母体免疫細胞の攻撃を免れるの であろうか.
図 1 交配から妊娠成立まで 図 2 妊娠時には父親移植抗原に対して一過性にトレランスが生じる I. 妊娠時には父親抗原特異的なトレランスが一過 性に誘導される BALB/C ♀マウスに C57BL/6 ♂マウス由来の腫 瘍細胞を接種すると,腫瘍細胞は生着できずに拒絶 される.一方 BALB/C ♀マウスと C57BL/6 ♂マ ウスを交配して妊娠させてから,C57BL/6 由来の 腫瘍細胞を接種すると,腫瘍細胞は生着する.但 し,他のマウス由来の腫瘍細胞を妊娠 BALB/C マ ウスに接種しても,腫瘍細胞は拒絶されるため,こ の反応は父親抗原特異的トレランスの誘導を意味し ている1).一方,分娩後 2~3 週間以内に,生着し た C57BL/6 由来の腫瘍細胞は拒絶されるため1), 父親抗原特異的トレランスは妊娠しているときに限 局していることが判った(図 2). ヒ ト に お い て も 胎 児 の リ ン パ 節 中 に は , 微 量 (0.01~0.8)の母体リンパ球が存在し,胎児の制 御性 T 細胞は,母親由来 T 細胞に対して,特異的 に免疫反応を抑制し,また母親由来制御性 T 細胞 も,胎児抗原に対して特異的に免疫反応を抑制す る2).これらの成績は,マウスでもヒトでも妊娠時 には,母子間免疫寛容が存在することを示している. II. 母子間免疫寛容は,どのような機序で誘導さ れるのか 免疫寛容を誘導する細胞の本態が,CD4+CD25+ Foxp3+制御性 T 細胞であることは,Sakaguchi ら により明らかにされた3).2004 年になり,マウスな
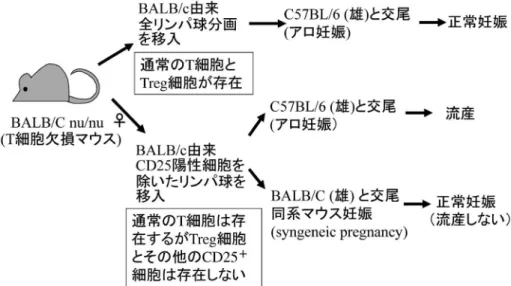
図 3 制御性 T 細胞はアロ妊娠維持には必要である らびにヒト妊娠維持に制御性 T 細胞が重要な役割 りを果たすことが報告された4,5).図 3 に示す如く, T 細胞欠損マウスである BALB/C nu/nu(♀)マ ウ ス に BALB / C 由 来 全 リ ン パ 球 を 移 入 し , C57BL/6(♂)マウスと交配させても,流産は生 じ な い . 一 方 , CD25 陽 性 リ ン パ 球 を 除 い た BALB/C 由来リンパ球を BALB/C nu/nu ♀マウス に移入した後に,C57BL/6 ♂マウスと交配させる と流産が生じる.一方,BALB/C ♂マウスとの同 系マウスの妊娠では,流産しない.Aluvihare らは CD25+細胞を除去しているので制御性 T 細胞が欠 損しているから,アロ妊娠では拒絶反応が生じると 考えた4).我々は,ヒト妊娠では,とくに子宮内膜 に制御性 T 細胞が増加すること,流産例では末梢 血,子宮内膜ともに制御性 T 細胞が減少すること を見出した5).さらに,妊娠中に高血圧,蛋白尿を 呈し,母子ともに予後不良となる妊娠高血圧腎症 (preeclampsia)においても,母体末梢血,妊娠子 宮で制御性 T 細胞が減少することを見出し6),これ らの病態に母子間免疫寛容の破錠が関与している可 能性を示した.さらに最近の研究により,原因不明 流産例や妊娠高血圧腎症では,Th17 細胞が増加 し,制御性 T 細胞が減少しており,制御性 T 細胞 への分化障害も,その病態に関与していることが明 ら か と な っ て い る7). ま た , 制 御 性 T 細 胞 を 抗 CD25 単クローン抗体で減少させた場合,着床期で はアロ妊娠では着床不全を引き起こし,妊娠初期で は流産を引き起こすが,妊娠後期では,全く異常を 示さなかったことより8),妊娠後期では胎児を攻撃 する細胞傷害性 T 細胞(CTL)は,clonal deletion を起こしているのかもしれない. これら,胎児もしくは父親抗原特異的制御性 T 細胞は,妊娠のいつから増加し,どのような機序で 増加するのであろうか.Robertson らは,精液中の 液体成分である精漿が父親抗原特異的 T 細胞の誘 導 に重 要な 役 割り を 果た して い るこ とを 報 告し た9).精嚢腺を除去すると精漿がなくなるが,精子 は存在し,妊娠は可能である.一方,精管を結紮す ると精子は消失するが,精漿は精液中に含まれるこ とになる.精嚢腺除去マウスでは,♂由来腫瘍細胞 を妊娠♀マウスに接種しても,♂由来腫瘍に対する トレランスは誘導されなかったが,精管結紮マウス と交尾した♀マウスでは♂由来腫瘍に対してトレラ ンスを誘導した.我々は別の系で,精嚢腺除去マウ スでは,活性化され増殖している父親抗原特異的制 御性 T 細胞が子宮所属リンパ節や妊娠子宮に増加 しないことを報告した10).また,精漿のプライミン グにより,着床前に父親抗原特異的制御性 T 細胞 が子宮の所属リンパ節に集まり,増殖マーカーであ る Ki67 を発現していること,着床後,すみやかに 子宮内に同制御性 T 細胞が集積することを見出し た10). III. 妊娠時における NK 細胞の役割 哺乳類の妊娠子宮には,NK 細胞が増加する.こ れらの NK 細胞が免疫寛容に関与するかについて は,明らかでなかった.我々はマウス白血病細胞 WEHI 3Bを BALB/C マウスに輸注し,白血病モ デルを作製すると,1 ヶ月後に末梢血中,骨髄中に CD25+NKが増加することを見出した11).この NK
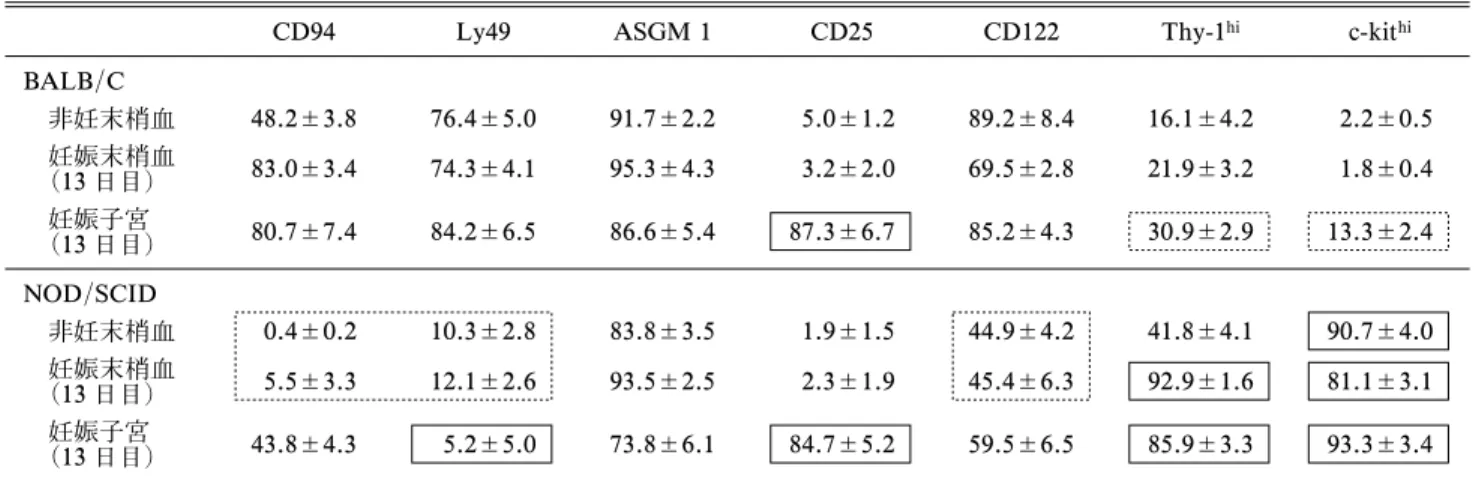
表 1 妊娠時における NK 細胞分画の変化
CD94 Ly49 ASGM 1 CD25 CD122 Thy-1hi c-kithi
BALB/C 非妊末梢血 48.2±3.8 76.4±5.0 91.7±2.2 5.0±1.2 89.2±8.4 16.1±4.2 2.2±0.5 妊娠末梢血 (13 日目) 83.0±3.4 74.3±4.1 95.3±4.3 3.2±2.0 69.5±2.8 21.9±3.2 1.8±0.4 妊娠子宮 (13 日目) 80.7±7.4 84.2±6.5 86.6±5.4 87.3±6.7 85.2±4.3 30.9±2.9 13.3±2.4 NOD/SCID 非妊末梢血 0.4±0.2 10.3±2.8 83.8±3.5 1.9±1.5 44.9±4.2 41.8±4.1 90.7±4.0 妊娠末梢血 (13 日目) 5.5±3.3 12.1±2.6 93.5±2.5 2.3±1.9 45.4±6.3 92.9±1.6 81.1±3.1 妊娠子宮 (13 日目) 43.8±4.3 5.2±5.0 73.8±6.1 84.7±5.2 59.5±6.5 85.9±3.3 93.3±3.4 (Lin Y., Saito S., et al. Fertil Steril. 2009)12) は 11)の論文で制御性 NK 細胞として共通の性格を有する細胞表面マーカー
細胞は他に Thy1, 2 が強陽性で c-kit 弱陽性で, CD94と Ly49 が 陰性 であ った. この NK 細 胞は NK 活性 が弱 く, TGFb を産 生し ,樹 状細 胞と CD25+NK細胞を共培養すると樹状細胞上の MHC class II 発現を減少させ,また,細胞傷害性 T 細胞 (CTL)誘導を阻害し,免疫抑制活性を有してい た.妊娠時にこれらの CD25+NK細胞が増加する かを調べたところ,表 1 のように CD25+NK 細胞
は妊娠子宮に増加し,Thy1highckit+の NK 細胞は,
NOD/SCID 妊娠マウス子宮に多く存在することが 判 っ た12). 表 面 マ ー カ ー か ら CD25+Thy 1highc kit+NK細胞を制御性 NK 細胞と仮に仮定すると, BALB/C 妊娠子宮の 13程度が制御性 NK 細胞で あり,NOD/SCID マウスでは 80以上の NK 細胞 が制御性 NK 細胞であることになる.これらの NK 細胞は 20前後が TGFb を産生し,66~81が IL-10を産生していた12). 通常,妊娠マウスに poly (1 : C)や LPS を投与 すると流産が誘導されるが,NOD/SCID マウスで は poly (1 : C)や LPS 投与によっても流産は増加 せず,NK 活性の活性化も認められなかった12).こ
れらのことより,CD25+Thy1highckit+NK細胞は
免疫制御活性を持ち,TLR を介した刺激に対し て,過剰に反応しないよう調節していると考えられ る.我々は NOD/SCID マウスに CD25 陽性細胞を 除いた BALB/C マウスのリンパ球を移入した際, C57BL/6 ♂マウスと交配すると高率に流産が生じ るが,妊娠した NOD/SCID 由来もしくは BALB/ C 由来の CD25+NK 細胞を輸注すると,流産率は 著しく減少することを認めている.これら CD25+ NK 細胞は,ヒト妊娠子宮に存在することは,すで にわれわれにより報告されている13).しかしながら CD25+NK 細胞はヒト流産例で著明に減少しない ことより,ヒトでは別の subpopulation の NK 細胞 が,免疫抑制活性を持つのかもしれない.今後の検 討が必要であろう. 結 論 制御性 T 細胞のみならず,制御性 NK 細胞も妊 娠子宮で増加し,妊娠維持に重要な役割を果たす, 精漿のプライミングは父親抗原特異的制御性 T 細 胞の子宮内への集積に重要な役割を果たしている. また,これらの母子免疫寛容がうまく働かないと, 着床不全や,流産を引き起こし,妊娠高血圧腎症の 病態の一因となっている可能性がある. 文 献
1) Tafuri A, Alferink J, M äoller P, et al. : T cell awareness of paternal alloantigens during preg-nancy. Science. 270 : 630633, 1995.
2) Mold JE, Micha äelsson J, Burt TD, et al. : Maternal alloantigens promote the development of tolerogenic fetal regulatory T cells in utero. Science. 322 : 15621565, 2008.
3) Sakaguchi S. : Regulatory T cells : key con-trollers of immunologic self-tolerance. Cell 101 : 454458,2000.
4) Aluvihare VR, Kallikourdis M, Betz AG. : Regulatory T cells mediate maternal tolerance to the fetus. Nat Immunol. 5 : 266271, 2004. 5) Sasaki Y, Sakai M, Miyazaki S, et al. : Decidual
and peripheral blood CD4+CD25+ regulatory
T cells in early pregnancy subjects and spon-taneous abortion cases. Mol Hum Reprod. 10 : 347353, 2004.
ual- CD4+CD25bright regulatory T cells in
pre-eclampsia. Clin Exp Immunol, 149 : 139145, 2007.
7) Saito S, Nakashima A, Ito M, Shima T. : Clini-cal implication of recent advances in our under-standing of IL-17 and reproductive immunolo-gy. Exp Rev Clin Immunol. 7 : 649657, 2011. 8) Shima T., Sasaki Y., Itoh M., et al. : Regulato-ry T cells are necessaRegulato-ry for implantation and maintenance of early pregnancy but not late pregnancy in allogeneic mice. J. Reprod Im-munol 85 : 121129, 2010.
9) Robertson SA, Guerin LR, Bromˆeld JJ, et al. : Seminal ‰uid drives expansion of the CD4+
CD25+T regulatory cell pool and induces
toler-ance to paternal alloantigens in mice. Biol Reprod. 80 : 10361045, 2009.
10) Shima T, Inada K, Nakashima A, et al. :
Pater-are increased just before the implantation by seminal ‰uid priming. J Reprod Immunol 94 : 12, 2012.
11) Ebata K, Shimizu Y, Nakayama Y, et al. : Im-mature natural killer cells suppress dendritic cell functions during the development of leukemia in a mouse model. J Immunol. 176 : 41134124, 2006.
12) Lin Y, Zhong Y, Saito S, et al. : Characteriza-tion of natural killer cells in nonobese diabetic/ severely compromised immunodeˆcient mice during pregnancy. Fertil Steril. 91 : 26762686, 2009.
13) Nishikawa K. Saito S, Morii T, et al. : Accumu-lation of CD16-CD56+natural killer cells with
high a‹nity interleukin 2 receptors in human early pregnancy decidua. Int Immunol 3 : 743 750, 1991.