第14 15回
本日の学習のポイント
• ナイーブT細胞が樹状細胞(抗原提示細胞)に出会う過程での接着
因子とケモカインの役割
• ナイーブT細胞の抗原認識における補助刺激分子B7とCD28の役割
• 感染に対するエフェクターT細胞(キラーT細胞,Th1細胞,Th2細胞)
の役割
• Th1・Th2バランスの偏りとハンセン病の病態の変化
• Th1細胞/Th2細胞上のCD40リガンド(CD40L)と、マクロファージ/B
細胞上のCD40との結合の意義
• 莢膜保有菌ワクチンでの莢膜抗原へのキャリアタンパク付加の意義
自然免疫の特徴
• 微生物に特有の分子・構造を認識する(パターン認識)。
• 自然免疫が感知した病原体の情報を、獲得免疫に伝える。
マクロファージ
好中球
樹状細胞
ナイーブ
T細胞
CD8
CD4
ナイーブ
T細胞
自然免疫
獲得免疫
キラー
T細胞
ヘルパーT細胞
MBL
補体
樹状細胞は末梢組織で抗原を補足し
、
二次リンパ組織で外来抗原をナイーブT細胞へ提示する
図
8.2
組織における
樹状細胞の形態変化
赤:リソソームタンパク質
MHC分子へのペプチド抗原の結合過程
図
5.20
ウイルスなど
細菌・真菌
タンパク質など
MHC クラスⅡ分子
MHC クラスⅠ分子
樹状細胞による抗原の処理と提示様式
図
8.5
図
8.4
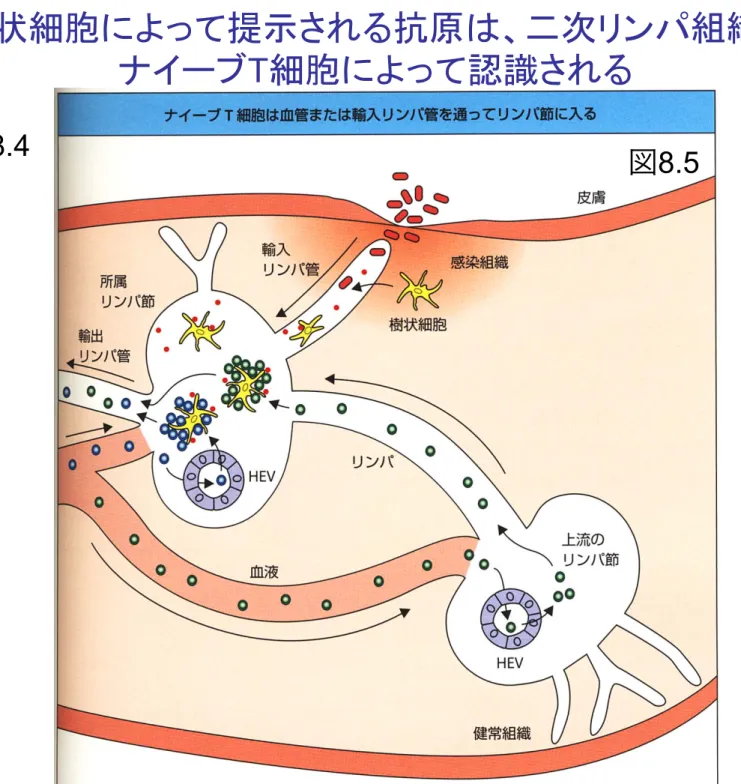
樹状細胞によって提示される抗原は
、
二次リンパ組織で
ナイーブT細胞によって認識される
ナイーブT細胞は接着因子を介して高内皮小静脈に
結合し
、
樹状細胞から放出されたケモカインの濃度勾配
に従って二次リンパ組織へと移動する
CCL21, CCL19
(樹状細胞から)図
8.6
図
8.7
CCR7(ケモカイン受容体)
L-
セレクチン:
CD62L
ナイーブT細胞と樹状細胞は
、
接着因子を介し
て強固に結合し
、
T細胞受容体と「抗原-MHC複合体」との結合を
誘導する
図
8.8
図
8.9
樹状細胞やマクロファージがナイーブT細胞に抗原認識させて増殖と
分化を誘導するためには
、
B7
と
CD28
を介する刺激も必要である
図
8.10
補助刺激分子(
B7
)
と
CD
28
との結合を介さないナイーブT細胞の抗原認識は
、
T細胞の不応答
(anergy)
をもたらす
図
8.18
T細胞の抗原認識と活性化には、T細胞受容体とMHC classII分子との結合による
第1シグナル(①)に加え、抗原提示細胞上の
B7(
CD80/CD86)
とT細胞上のCD28との
結合による第2シグナル②が必要である。
第2シグナルがなければ、T細胞は
アナジー
となる(末梢での自己免疫の回避に重要)。
アナジー(anergy)
=免疫不応答ともいう。 単に活性化されないのみならず、再び同じ抗
原に出会ったときにも反応できない状態をいう。
B7
CD28
CD28
B7
CD28
末梢性免疫寛容
図
8.13
微生物由来抗原(LPSなど)は
、
それを感
知するToll様受容体などを介して
、
抗原提
示細胞上に補助刺激分子を発現させる
。
マクロファージ
樹状細胞
NK細胞などに発現
図
2.22
MyD88!
IRAK!
TRAF
!
NFκB!
JNK/p38!
ペプチドグリカン
TLR4
LBP!LPS
リポテイコ酸
細菌性(CpG)-DNA
CD14!リポ蛋白
TLR1/2/6!
TLR5
細菌性
フラジェリン
dsRNA
ヒト
Toll
様レセプター
(TLR)
による微生物成分の認識
TLR3
TLR7/9
ssRNA
TNF-α
+
補助刺激
分子の発現
微生物の構成成分はTLRを介して適応免疫を増強する
アジュバント
として作用する
。
タンパク質だけで免疫しても免疫応答はほとんど起こらない
TLR4
LBP!LPS
リポテイコ酸
細菌性(CpG)-DNA
CD14!リポ蛋白
TLR1/2/6!
TLR5
細菌性
フラジェリン
dsRNA
TLR3
TLR7/9
ssRNA
ヒト
Toll
様レセプター
(TLR)
による微生物成分の認識
ペプチドグリカン
不活化ワクチンでは
、
主に抗体による体液性免疫(中和抗体)のみが誘導される
。
標準微生物学 免 疫 応 答 強生ワクチンと不活化ワクチンとの比較
プロフェッショナル抗原提示細胞によって
活性化されたナイーブT細胞はIL-2 を分泌し
、
オートクリン(autocrine)作用で増殖する
ナイーブCD4陽性T細胞は
、
プロフェッショナル抗原提示細胞が提供
する環境(サイトカイン環境)によってTh1細胞やTh2細胞へと分化する
図8.20
図8.19
サイトカイン
転写因子
CD4
ナイーブ
T細胞
Th1
免疫での役割
疾患との関連
B細胞
Th2
Th17
Treg
自己免疫疾患
アレルギー性疾患
自己免疫疾患
自己免疫(
Treg↓で)
悪性疾患(
Treg↑で)
自己免疫疾患
マクロファージの
活性化
(細胞性免疫)
B細胞の分化
(液性免疫)
抗菌ペプチドの産生誘導
好中球の動員
→真菌・細菌感染防御
免疫応答の抑制
?
マクロファージ
ヘルパーT細胞の多様性
抑制
IL-12
IFN-γ
IL-4
TGF-β
IL-6
TGF-β
IFN-γ
IL-4, IL-5
IL-17, IL-21
TGF-β, IL-10
図8.28
図8.21
ハンセン病ではTh1細胞応答vsTh2細胞応答どちらが
優位に働くかによって異なる病像(病態)を呈する
CD8陽性ナイーブT細胞は3つの様式で活性化される
図8.22
CD8陽性ナイーブT細胞が一度補助刺激分子を介してエフェクターT細
胞に分化すると
、
IL-2によってオートクライン的に分化増殖し
、
補助刺
激分子を介せずにclass I分子上で提示される抗原を認識して感染細
胞を殺傷する
図8.24
図
8.7
図
8.4
T細胞の活性化に伴い
、
細胞表面分子の発現は変化する
• ヒト全身性エリテマトーデス(SLE)のモデルマウス
• 症状
リンパ節腫脹、耳介/皮膚の壊死, 関節炎、糸球体腎炎(ループス腎炎)
(24週までに約半数が腎不全にて死亡)
• 血清学的特徴
ヒトSLE同様に、dsDNAや核タンパク成分に対する抗体(自己抗体)が陽性
抗核抗体
抗dsDNA抗体
抗Sm抗体
抗RNP抗体
24 weeks old!
Molano, et al, clinical immunol, 2003
細胞表面接着因子L-セレクチン(CD62L)とCD44の発現を調べることで
CD4陽性T細胞の活性化の状態を知ることが出来る
61.9" 15.7" 4.8" 17.8"CD44
CD62L
6週齢"
12週齢"
24週齢"
31.9%" 7.6" 37.7%" 22.8" 68.2%" 17.7" 8.1" 6.0" 32.1" 43.0" 22.0" 2.9" 29.1" 50.7" 18.8" 1.4" 1.0%" 90.5%" 4.7" 3.8"ナイーブ
メモリー
エフェクター
SLEモデルマウス
(MRL/
lpr
系)
健常マウス
(C57BL/6系)
図8.27
パーフォリン
グランザイム
キラーT細胞の細胞傷害
キラーT細胞の細胞傷害
ウイルス感染細胞
細胞傷害性
T
細胞が特異抗原を認識すると
、
サイトトキシンが直接標的細胞に向けられる
図8.27
LG: lytic granule
パーフォリン
グランザイム
などを含む傷害顆粒
図8.36
図8.34
Th1細胞によるマクロファージの活性化機構
CD40LとCD40との結合は
、
マクロファージをIFN-γ
活性化Th1細胞によるマクロファージ小胞内細菌感染への応答
図8.35
LT:
リンホトキシン
GM-CSF:
顆粒球
-
マクロファージコロニー刺激因子
CXCL2:
ケモカイン
LT LT図8.36
図8.34
Th1細胞によるマクロファージの活性化機構
CD40LとCD40との結合は
、
マクロファージをIFN-γ
に対して反応性にするために重要である
。
乾酪性壊死
活性化Th2細胞はナイーブB細胞を刺激して増殖・分化させる
B細胞表面のCD40とTh2細胞表面のCD40Lとの結合は
体細胞高頻度変異
と
クラススイッチ
に必須であり
、
重要である
図8.37
B7
CD28
CD28
B7
CD28
図8.34
図8.18
①B7とCD28の結合は抗原を認識するエフェクターT細胞に分化さ
せるために重要である
②CD40とCD40Lとの結合はエフェクターT細胞としての機能を発揮
させるために重要である
図8.37
まとめ
• B細胞のクラススイッチ(CSR)や体細胞高頻度変異(SHM)の過程
で必要な遺伝子の突然変異の結果、クラススイッチができないため
に血清IgMが高値となる疾患。"
• クラススイッチに関与する遺伝子は複数あり、いくつかの原因遺伝
子が報告されている(HIGM Type 1~6)。"
"
• クラススイッチに障害があるために中和やオプソニンに重要なIgG
、粘膜免疫に重要なIgA値が低値となり、易感染性(呼吸器感染、
膿皮症、中耳炎など)となる(主に液性免疫不全となる)。"
• 生後2年以内に診断される事が多い。"
• ガンマグロブリン定期補充療法(1回/3 4週)が治療の主軸。"
高
IgM症候群
HIGM Type 1! ! ! ! HIGM Type 2! ! ! HIGM Type 3! ! ! HIGM Type 4! ! ! HIGM Type 5! ! ! HIGM Type 6! ! CD40-L " (X-linked R)" " " AID (AR)! " " CD40 (AR)" " " Unknown (AR?)" " " UNG (AR)" " " NEMO " (X-linked R)" (NFκB essential modulator)"
Mutation (genetic inheritance)"
A co-stimulatory molecule on T cells. Altered T cell-B cell
interaction. Without this co-stimulation, B cells do not receive the needed signal to undergo SHM and CSR. "
"
An essential factor for SHM and CSR. Clinically similar to HIGM5." "
A co-stimulatory molecule on B cells. Altered T cell-B cell interaction. Similar to HIGM1 patients, HIGM3 patients fail to undergo SHM and CSR."
"
Intact SHM but altered CSR; It is supposed that the mutation in HIGM4 patients occurs in a gene active downstream of AID." "
Responsible for the cleavage of cytosines that have been deaminated by AID. Clinically similar to HIGM2."
"
Mutated NEMO is unable to phosphorylate IκB. Therefore,
transcription cannot be initiated by NFκB, and SHM and CSR are blocked."
Type"
高
IgM症候群
HIGM Type 1! HIGM Type 3! HIGM Type 6! HIGM Type 2! HIGM Type 5! HIGM Type 4!
Etzioni A and Ochs HD Pediatr Res 2004" κ
κ
高IgM症候群の原因遺伝子
"
T
細胞
"
補体
食細胞
感染後の経過時間
時間
0
6
12
8
10
12
14
16
CD8 ナイーブ T細胞 感染細胞 殺傷 キラーT細胞ナイーブ
T細胞
CD4
日数
樹状細胞
麻疹ウイルス感染における免疫応答 抗体
自然免疫
獲得免疫
B細胞
補体
食細胞
感染後の経過時間
時間
0
6
12
8
10
12
14
16
CD8 ナイーブ T細胞 感染細胞 殺傷 キラーT細胞CD4
日数
樹状細胞
麻疹ウイルス感染における免疫応答 抗体
自然免疫
獲得免疫
B細胞
ナイーブ
T細胞
補体
食細胞
感染後の経過時間
時間
0
6
12
8
10
12
14
16
CD8 ナイーブ T細胞 感染細胞 殺傷 キラーT細胞ヘルパーT細胞
日数
麻疹ウイルス感染における免疫応答 抗体
自然免疫
獲得免疫
B細胞
補体
食細胞
感染後の経過時間
時間
0
6
12
8
10
12
14
16
CD8 ナイーブ T細胞 感染細胞 殺傷 キラーT細胞分化
麻疹ウイルス感染における免疫応答 抗体
自然免疫
獲得免疫
B細胞
ヘルパーT細胞
日数
体細胞高頻度変異
、
クラススイッチ
補体
食細胞
感染後の経過時間
時間
日数
0
6
12
8
10
12
14
16
CD8 ナイーブ T細胞 感染細胞 殺傷 キラーT細胞形質細胞
麻疹ウイルス感染における免疫応答 抗体
自然免疫
獲得免疫
ヘルパーT細胞
分化
補体
食細胞
感染後の経過時間
時間
0
6
12
8
10
12
14
16
CD8 ナイーブ T細胞 感染細胞 殺傷 キラーT細胞日数
記憶B細胞
麻疹ウイルス感染における免疫応答 抗体
自然免疫
獲得免疫
形質細胞
ヘルパーT細胞
分化
Th2細胞は
、
同一抗原を認識するB細胞のみを活性化する
ハプテン:抗原性があるが
、
免疫原性がない物質
(
免疫応答を誘導できない物質
)
一般に
、
多糖や脂質
、
核酸などの分子量数百以下の低分子
ハプテンに対する抗体を産生させる機序
図8.37
小児髄膜炎の起因菌
Hibと肺炎球菌のワクチン抗原には
キャリアタンパク
を結合させている(1)
•
インフルエンザ菌b型(Hib)
(Haemophilus influenzae: Hi
、グラム陰性桿菌
)
19世紀末、欧州でインフルエンザが大流行した際、患者の喀痰から発見されたことから、
インフルエンザの原因菌と考えられていた(インフルエンザウイルスは
1933年に発見された)。
Hiは
多糖体(ポリサッカライド)からなる莢膜
の有無によって大別されるが、莢膜を有する
ものは、
a~fの6つの血清型に分類され、このうちb型(Hib)は病原性が強く、肺炎や喉頭蓋炎
、中耳炎、髄膜炎をひきおこし、毎年
500人以上の乳幼児がHib髄膜炎に罹患している。
•
肺炎(レンサ)球菌
(Streptococcus pneumoniae:
グラム陽性球菌
)
莢膜を有し、肺炎の起因菌として重要であり、2才以下の乳幼児や高齢者が重篤な症状
をひきおこす。中耳炎や、髄膜炎をひきおこす起因菌としても重要であるが、特に生命予後
にかかわる髄膜炎では、インフルエンザ菌とともに2大起因菌として重要である。
肺炎球菌の
莢膜
厚生労働省 新興・再興感染症研究事業(H22 新興 一般 013)より引用
メチレンブルー染色
厚生労働省 新興・再興感染症研究事業(H22 新興 一般 013)より引用
Hibと肺炎球菌のワクチン抗原には
キャリアタンパク
を結合させている(2)
• Hib、肺炎球菌の両者とも
莢膜
を有し、親水性の付加により
マクロファージや
好中球からの捕獲から逃れ
、また、
補体の結合による溶菌からも回避
できる。
• 両者を排除するためには、莢膜に特異的なIgG抗体を誘導し、
オプソニン作用
によるマクロファージや好中球による貪食が効果的
である。
オプソニン作用
莢膜非保有菌
莢膜保有菌
溶菌される
溶菌されない
標準微生物学第11版より改変抗莢膜多糖体抗体
Hibと肺炎球菌のワクチン抗原には
キャリアタンパク
を結合させている(3)
• 莢膜抗原は多糖体であるため、MHC class II 分子上にペプチド抗原として提示
されない。
→多糖体成分のみの免疫では莢膜抗原特異的Th2細胞を誘導する
ことは困難。
• その結果、B細胞のクラススイッチができないため、オプソニン抗体(IgG)産生
が誘導できない。
• Th2細胞が莢膜多糖体を抗原として認識出来るように、ペプチド抗原となる
キャ
リアタンパク
を付加させる必要がある。
Hibと肺炎球菌のワクチン抗原には
キャリアタンパク
を結合させている(4)
多糖体抗原のみでの免疫
多糖体抗原
+
キャリアタンパク
での免疫
キャリアタンパク
MHC class II分子上に抗原
を提示できないため、
Th2
細胞から刺激を受けない。
MHC class II分子上に抗原を
提示し、
Th2細胞から刺激を受け
て
IgGにクラススイッチする。
IgM
IgM
IgG
クラススイッチを誘導
IgM
Hibと肺炎球菌のワクチン抗原には
キャリアタンパク
を結合させている(5)
多糖体抗原のみでの免疫
多糖体抗原
+
キャリアタンパク
での免疫
キャリアタンパク
MHC class II分子上に抗原
を提示できないため、
Th2
細胞から刺激を受けない。
MHC class II分子上に抗原を
提示し、
Th2細胞から刺激を受け
て
IgGにクラススイッチする。
IgM
IgM
IgG
クラススイッチを誘導
IgM
肺炎球菌ワクチン:プレベナー®(
キャリア:ジフテリアトキソイド
)
Hibワクチン:アクトヒブ®(
キャリアタンパク:破傷風トキソイド
)
第一製薬「ワクチンインフォーメーション」Vol.2、No.12から引用。