ミツバチ科学 HoneybeeScience(2004)
セイ ヨウミツバチの攻撃的な働き蜂の
脳か ら同定された
Ka
k
u
g
o
ウイルス
藤幸 知子,竹内 秀明,久保 健雄
ミソバチの攻撃行動について
「ミツバチ科学」の読者の方々には,ミツバ チの生活の詳 しい紹介は不必要 とは思われ る が,私たちの研究の狙いをご理解いただ くため 仁,まずは私たちが注目しているミツバチの生 物学的特徴について簡単に述べさせていただき たい.ミツバチでは,雌が女王蜂 (生殖カース ト) と働き蜂 (労働カース ト)にカース ト分化 する.また働き蜂は,羽化後の日齢に応 じて巣 の掃除や育児等の巣内の仕事から,門番を経て, 花の蜜や花粉の採集 といった巣外の仕事に携 わる (齢差分業)(Winston.1987;Breed,etal, 1990).私たちは 1993年から,こうしたミツ バチの社会性行動を支える脳の分子的メカニズ ムを探るために,セイヨウミツバチ匝pl
'
sme
l
-1
1
'
f
e
r
a
L.)を用いて,脳の領野 (脳の部域)選択 的に発現する遺伝子や,脳で行動選択的に発現 す る遺伝子の網羅的な同定 と解析を進めてき た(Kamikouchi.eta1.,1998;Kamikouchi,etal, 2000;Takeuchi,eta12001;Takeuchi,eta1., 2002;Sawata,eta1.,2002,Park.eta1.,2003: Takeuchi,etal,2004).今回は,働き蜂の攻撃 行動に注目して行なった研究について,特に本 誌で紹介 したい.なお, ミツバチの脳で高次中 枢 (キノコ体)特異的に発現する遺伝子の解析 結果の一部については,2001年に,竹内らが 本誌ですでに紹介 しているのでご参照いただけ ると幸いである (竹内ら,200廿 最初にミツバチの攻撃行動がどのようなもの か復習 してみたい.ハチ頬の進化については諸 説あるが,一説によると,最初はキバチやハバ チのような単独性で植物食の仲間か ら出発 し, やがて肉食の寄生蜂が出現 した際に,宿主を麻 酔するために産卵管が針 として用いられるよう になったとされている.次いで.アシナガバチ やスズメバチのような社会性狩 りバチや,花の 蜜や花粉に全ての食料を依存する植物食のハナ バチが出現 した とされている (坂上,1983). 現在のミツバチは,最も高度な社会性を獲得 し たハチ頬 として,コロニーを形成 し,同じ巣の 中で共同 して幼虫や蛸を育てるとともに,花粉 や花の蜜を加工 したハチミツを食料 として貯え るという食料戦略をとっている. 巣に貯えられたハチミツに加えて,巣で育て られる幼虫や桶は,スズメバチや熊など,ミツ バチの天敵にとっては栄養価の高い食料になる ので,外敵からコロニーを守るためにミツバチ は高い攻撃性を有 している.この攻撃役を担 う のは働き蜂である.働き蜂は自分 自身の子孫を 残さず,女王蜂が産む子供 (働き蜂にとっては 妹)の世話をするので,その行動は全般的に 「利 他的」であるが,中でも攻撃行動は特に 「利他 的」である.セイヨウミツバチでは,働き蜂の 針は外敵に刺さると毒腺 とともに働き蜂の体か ら脱落するためにその個体は早晩死亡するとい うように,働き蜂が死 と引き換えに仲間を守る からである(Dade,1962;Wilson,1999).私た ちは,この攻撃性がコロニーの特定の働き蜂(門 番蜂)に分配されている点に注目し,ミツバチ の攻撃行動に関わる遺伝子候補 として,攻撃性 が高い門番蜂の脳で特異的に発現する遺伝子を 検索することにした.攻撃性を司る脳の分子的 メカニズムについては,ミツバチに限らず動物 一般について,現在なお明らかではない.攻撃 行動がコロニー内の一部の個体によって分業されるという,社会性昆虫の特徴を利用 したアプ ローチ法により,ミツバチひいては動物一般の 攻撃性の分子的基盤について,何 らかの手掛か りを得ることを期待 したのである.この研究の 大部分は,今年の2月に,米国のウイルス学 雑誌
(
J
o
u
r
n
a
lo
fVi
r
o
l
o
g
y
)
に出版されている が (Fu
j
i
y
u
k
i
.
e
t
a
l,2004),原著論文をご覧いた だける機会は少ないと思 うので,ここでは,研 究の展開と結果をわか りやす く説明 したい.攻撃的な働き蜂の脳で発現する遺伝子の
検索 と
.Ka
k
u
go
ウイルスの同定
最初に行なったのは,ミツバチの働き蜂を, 攻撃性を指標に選別することであった.いろい ろと試 した結果,門番蜂にオオスズメバチ (ミ ツバチの天敵)を田として糸で吊るして提示す ると,オオスズメバチに飛びかかって くるもの がいたので,これ らを攻撃性が高い蜂 (攻撃蜂) として採集 した.一方,巣の中の巣板の上にい る働き蜂にオオスズメバチを提示すると,一部 の蜂が逃げまどう傾向があったので,これらを 逃避蜂 として採集 した.私たちにとってはそも そも,田のオオスズメバチを常に入手するのが 困難であったが,幸いにも,小野正人先生 (玉 川大学農学部)がど供与下さることにな り,今 日に至るまで,共同研究をさせていただいてい る.さ らには,J
.Vi
rol.誌 の掲載誌の表紙や, 私たちの研究成果が新聞報道された際には,小 野先生が撮影された 「天敵オオスズメバチを攻 撃するセイヨウミツバチの門番峰」の素晴 らし い写真を使わせていただいた (図1
). Kakugoタンパク質 図1 オオスズメバチを攻撃するミツバチ (撮影 :小野正人) それぞれ約 100匹程度の攻撃蜂 と逃避蜂の 脳からR
NA
を抽出し,d
i
f
f
e
r
e
n
t
i
a
一
d
i
s
p
l
a
y
法に より,逃避蜂に比べて,攻撃蜂の脳に特異的に 存在するR
NA
を検索 した・Di
f
f
e
r
e
n
t
i
a
ld
i
s
p
l
a
y
法は,複数のサ ンプルについて,いずれかの サンプルに選択的に存在する (発現 している)R
NA
を同定する方法である (L
i
a
n
ga
n
dP
a
r
d
e
e
, 1992). この方法については,2001年の総説 でも紹介させていただいたので詳細は省略する が (竹内ら,2001),逆転写反応 とPCR(ポ リ メラーゼ鎖反応)法,そしてシークエンスゲル 電気泳動を組み合わせることで,片方のサンプ ルのみに存在するR
NA
を検索できる.例えば, 攻撃蜂の脳で特異的に存在するRNAは,攻撃 蜂のレーンに特異的に検出されるバン ドとして 同定される.攻撃蜂の レーンに選択的に検出さ れた複数のバン ドの内,差が最も明瞭だったも のについて,ゲルから切 り出し,そこに含まれ るc
DNA
をサブクローニングした結果,1
つの キャプシドタンパク質 へリカーゼ
プロテアーゼ
RNA依存 RN
A
ポリメラー
ゼ サックブルー ドウイルス (ミツバチ) スキ ドクガピコルナ様ウイルス (ガ) 伝染性軟化病ウイルス (カイコ) A型肝炎ウイルス (ヒ ト) 口蹄疫ウイルス (ウシ) ポリオウイルス (ヒト) 急性蜂麻痔病ウイルス (ミツバチ) ショウジョウバエCウイルス (ハエ) (%) (%)(
%) 36 29 37 4 3 9 0 5 3 0 2 2 2 3 2 2 2 222
8
2 8 8 3 5 3 つ乙 つ乙 1 2 つん つん6
8
6
4
9
0
つL
2
つん
2
2
3 図 2 KakugoRNAのコー ドするタンパク質と他のピコルナ
(樵)ウイルスのタンパク質と相同性
の比 較 各酵素 ドメインにおけるKakugoタンパク質と他種ウイルスタンパク質との相同性を%で示
す
・
各ウイルスの宿主をウイルス名に続けて ()内に示す .遺伝子が同定され 私たちはそれを
Ka
ku
go
(栄 悼)と命名 した(
F
u
j
i
y
u
k
i
,
e
t
a
1
.
,
2
0
04
)
・名称は, 恰も 「覚悟」ができているような攻撃蜂から同 定されたことによる.Di
f
f
e
r
e
n
t
i
a
l
d
i
s
p
l
a
y
法 で 同定 され たKa
ku
go
のc
DNA
は断片 であったため,全長のKa
ku
go
c
DNA
を同定 し, どのよ うな タンパ ク質が コ ー ドされるか調べたところ,予想外なことに,Ka
ku
go
c
DNA
は色々な どコルナ様 ウイルスの タンパク質 と相同なタンパク質をコー ドするこ とがわかった (図2
)
.ウイルスゲノムにコー ドされるタンパク質は,ウイルス粒子の生成に 必要な複数のタンパ ク質を,一つの大 きな前 駆体 タンパ ク質 (ポ リプロテイ ン)の部分構 造 (ドメイン) として含むという特殊な構造を している.これらの ドメインとしては,ウイル ス粒子を構成するキャプシ ドタンパク質,ウイ ルスゲノムの複製に必要なへ リカーゼとポリメ ラーゼ,そしてこれらの ドメインを切断するプ ロテアーゼがある (図2
.3).Ka
ku
go
R
NA
の コー ドするタンパク質には,これらのタンパク 質 ドメインがすべて備わ っていた. ピコルナ 様ウイルスはゲノム としてDNA
ではな くR
NA
を持つので,宿主のゲノムDNA
中に挿入され る ことはない・実際,Ka
ku
go
R
NA
に対応す るDNA
も,ミツバチゲノム中には兄い出され なかった(
F
u
j
i
y
u
k
i
,
e
ta
1
.
,
2
0
04
)
.
このことは,Ka
kL
I
g
OR
NA
はミツバチのゲノムにコー ドされ るものではな く,新規 な どコルナ様 ウイルス(
K
a
k
u
g
o
ウイルス)のゲノムR
NA
そのもので あって,攻撃蜂の脳は,選択的にこのウイルス に感染 していた可能性を示唆 している,同じピ コルナ様 ウイルスに属するウイルスとしては, ミツバチのサックブルー ドウイルスや,ヒトのA
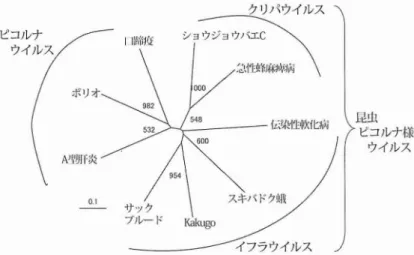
型肝炎ウイルス,ポリオ (小児麻疹)ウイル ス,ウシの口蹄疫病ウイルスなどがあるが,さ らに詳細な系統解析の結果,K
a
k
u
g
o
ウイルス は昆虫ピコルナ様ウイルスに属する新種のウイ ルスであることがわかった (図 4).昆虫 ピコ ルナ様ウイルスは,その名の通 り昆虫に感染す るが,ヒトを含めた哨乳類への感染は知 られて いない.
Ka
ku
go
R
NA
がウイルスゲノムと考えられた ため,ウイルスとしての性状解析を,東京大学 大学院医学系研究科の野本明男教授,大岡静 衣助手 (微生物学) と共 同で進めた.詳細な 説明は省略するが,Ka
ku
go
R
NA
が検出される 働き蜂の組織の粗抽出液をショ糖密度勾配遠心 @ 図3
ピコルナ様ウイルスのゲノムR
N
A
やポリプロテインの構造と複製過程R
d
R
p;
R
NA
依存R
N
A
ポリメラーゼ図4 Kakugoウイルスとその他のピコルナ様ウイルスの分子系統樹 各ウイルスのRdRpドメインのアミノ腔配列データにもとづき,近隣 結合法によって作成したもの. ラップ値を示す.枝の長さはア にかけた ところ,遊離のKakugoRNAの他 に, ウイル ス粒子 の大 きさに相 当す る画分 か らも
KakugoRNAが 回収 され た こ とか ら,KaJqlgo
RNAが, ウイルス粒子 中にゲノムRNAとして 存在す るこ とが示 唆 され た.さ らに,Kakugo ウイルスの感染性を調べ る 目的で,Kakugoウ イルスに感染 した働 き蜂の組織粗抽出液を,罪 感染の働 き蜂の腹部に接種 した ところ,実験群 で は3日後 に顕著 なKakugoRNA量 の上昇が 見 られ た (図5).一方,対照実験 として,非 感染個体 の組織粗抽 出液や緩衝 液を注射 した 場合には,KakugoRNA量の増大は認め られな かった. この ことは,KakugoRNAが確かに感 染性 因子 の構成要素 であ るこ とを示 してい る (Fujiyuki,etal,2004)・ 次にKakugoウイルスが働 き蜂の攻撃行動 と どの桂度,関連 しているか調べる目的で,働 き 蜂を育児蜂,攻撃蜂,逃避蜂,採餌蜂に分けて 採集 し, それ ぞれ の脳 で のKakugoRNAの存 在量 を比較 した ところ,攻撃蜂 に特異的 に検 出された (図6).また,攻撃蜂の体の中では, 脳に強 く検出され,胸部や腹部では検出されな か った. この ことは,Kakugoウイルスの感染 と,働 き蜂の攻撃行動が密接に関連 しているこ とを示 している(Fujiyuki,etal.,2004)・実際に 攻撃行動を示 した働き蜂の どの くらいの割 り合 各分岐点における数値はブー トス ト ミノ酸置換数の推定値に比例している. いの個体がKakugoウイルスに感染 しているか は不明であるが, これまで本能的行動 と思われ ていた働き蜂の攻撃行動にウイルス感染が影響 する可能性が初めて指摘 された点で,本研究は 学術的意義をもつ と思われ る. 6 5 4 3 つL l 撃 萩
聖
e V N a Ogn
満
X *0
1 2 3 注射後日数 図5 Kakugoウイルスの感染性 採餌蜂の頭部に攻撃蜂由来細胞粗抽出液(黒), 採餌蜂由来細胞粗抽出液 (斜線),バッファー (灰)を注入し,按種後,それぞれの日数後に ミツバチを回収した.また,無処理 (白)の 個体を同時に回収した.回収 した個体の頭部 におけるKakugoRNAの墨をアクチンmRNA 塁で補正 した値を相対値として示 した.*;佃 のサンプルと有意差があることを示す(p< 0.05). 用いたサンプル数は,左から (0日後)=3.5, 4.5:1日後 =6,8,8.8.2日後 =3,3,3.2.3日 級-6,7,6.8匹.149 0
0
0
0 n V0
0 0 A .2
0
8 64.
2(
㌔)
] J 宗 空 e r kiVN
∝ 育児蜂 0 0 0 0 0 0 2 0 8 6 4 2 ( % ) ヨ 頁 空 G 7 咽 V N 出攻
撃蜂 採集蜂 脳 頭部 胸部 腹部 図6 K
a
k
u
g
o
ウイルスの分布(
A
)
攻劉 箕でのR
N
A
ftiを1
0
0
%
とし,脳内の相対的なKa/alsoR
NA
芸を行動別に比頗 した・(
B)
脳でのR
NA
,L.i.を1
0
0
%
とし,攻碓削祭の休lノ、Jの相対的R
N
A
塁を比暇した.檎 子はKaku80R
NA
,無地はアクチンmR
NA
星を示す.ウイルス感染と攻撃行動の関連に
関する考察
現在の ところ,Ka
k
u
g
o
ウイルスの感染が ミ ツバチの攻撃行動 とどのよ うに関わ るかは不 明である.最 も面 白い可能性の一つは, ウイ ルス感染が,働 き蜂の攻撃行動を誘発す る と い うものであろう.これまでの実験結果では,Ka
k
u
g
o
ウイルスが感染 していないコロニーで も門番蜂の攻撃行動は観察されているので (藤 幸 ら,未発表),Ka
k
u
g
o
ウイルスの感染が攻 撃行動の発動に必須なわけではないのだろう. む しろウイルス感染は,元々攻撃性が高 くない 働き蜂に攻撃性を賦与 し,門番蜂の割合を増や している可能性が考えられる.もう一つの可能 性は,Ka
k
u
g
Oウイルスは元々,非常に低い レ ベルで普通の コロニー にも感染 してお り,働 き蜂が分業 し,門別 蚤になる過程でKa
k
u
g
o
ウ イルスの増殖に適当な生理条件がもたらされ, 脳でKa
k
u
g
o
ウイルスが増殖するというもので ある.いずれに しても攻撃蜂の捨て身の攻撃行 動は, 自身の死 と引 き換 えに, コロニーか ら ウイルスを排除する作用をもつ可能性があ り, ウイルス と宿主の機能的相互作用を考える上 で興味深い.なお, ご く最近,ダニ によって 媒介されるd
e
f
o
r
me
dwi
n
gv
i
r
u
s
(縮れた麹に なるミツバチの病気の,推定上の原 因ウイル ス)のc
DNA
の塩基配列がd
a
t
a
b
a
s
e
に登録され,Ka
k
u
g
Oウイルス とごく近縁のウイルスである ことが判明 しているが(
F
u
j
l
y
u
k
i
,
e
ta
l,2004),K
a
k
u
g
o
ウイルス との関連については現在の と ころ,不明である. ウイルス感染が動物の行動を変える例 として は,狂犬病ウイルス (攻撃的になる)やカイコ の核多核体病ウイルス (高い枝に登って死亡す る)などがあるが(
Mo
o
r
e
,2002),既知の例が いずれ も病理的な作用 と思われるのに対 して,Ka
k
u
g
o
ウイルスの感染 は本能的な攻撃行動に 関連する可能性が示された点で興味深い.小児 麻疹の原因ウイルスであるポ リオウイルスは, 感染 した運動神経を破壊することで麻痔を起 こ させ る (野本,1999). この ことか ら頬推する と,Ka
k
u
g
o
ウイルスが攻撃性に関連す るメカ ニズムの一つ として,Ka
k
u
g
o
ウイルスが感染 する神経細胞は元々,攻撃性の抑制に働 く神経 であ り,これをKa
k
u
g
o
ウイルスが破壊するこ とで,感染個体の攻撃性を高めるとい う仮説が 考 えられる.Kak
ugo
ウイルスの.ヒ トおよび
ミツバチにおける病原性について
先述 したように,Ka
k
u
g
o
ウイルスが属する 昆虫 ビコルナ様 ウイルスには哨乳類への感染性 が知 られているものは今の ところ存在 しない. 新聞報道 (読売新聞平成 16年 3月 5日付)で,Ka
k
u
g
o
ウイルスが ヒ トA
型肝炎 ウイルスと塩基配列上の類似性があることが短 く報 じられた ため,ミツバチが産生するローヤルゼ リーやハ チ ミツを食べると
A
型肝炎になるのではない か と危供された読者か ら筆者のところに2件, 「ミツバチ科学」には5件のお問い合わせをい ただいたそうだが,その懸念は恐 らくは杷憂で あろう.確かに,Kakugoウイルスが属する昆 虫 ピコルナ様 ウイルスは ヒ トA
型肝炎 ウイル スと同じ仲間 (ピコルナ様ウイルス)に分類さ れるが (図 4),これは遺伝子の塩基配列や構 造が似ているためであ り,同じ仲間には,病原 性も性質も異なる様々なウイルスが含まれてい る.Kakugoウイルスは, ミツバチや哨乳類へ の病原性を指標 として単離 された訳ではな く, 研究の過程で偶然,同定されたウイルスである. また,Kakugoウイルスが最近,ミツバチに顕 著に伝播 しているという訳でもない.これまで にローヤルゼ リーやハチミツを感染源とした病 原性ウイルスの報告例が全 くないことを考える と,Kakugoウイルスはミツバチに感染する多 くのウイルスと同様に,ヒ トにとっては無毒で あ り,また,古 くからミツバチに感染 している ウイルスの一つなのであろう. 一方で,病理的なものであれ,生理的なもの であれ,Kakugoウイルスが ミツバチの攻撃性 に関わるとすると,様々な展望が拓けて くる. 特に,北米では現在,攻撃性が非常に高いアフ リカ蜂化 ミツバチ (キラー ビー)の勢力の拡 大が深刻な問題になってお り (Guzman-Novoa,eta1.,1994),米国が現在,ミツバチのゲノム 計画を推進する主な動機の一つが,アフ リカ 蜂化ミツバチの問題への対処法の開発とされて いる.仮 に,アフ リカ蜂化 ミツバチにおいて Kakugoウイルスの感染が頻繁に見 られるよう であれば,Kakugoウイルスの駆除がアフリカ 蜂化ミツバチへの対処法 として浮かび上がって くる.また,Huntらは,遺伝学的な解析から, 独自に働き蜂の攻撃性に関わる遺伝子座を同定 してお り,これ らの遺伝子 と,Kakugoウイル スの感染への感受性 との関連も興味深い (Hunt,
eta1.,1998;Lobo,etalH2003)・Kakugoウ イ
ルスの今後の研究が,学術的にも産業的にも意 義深いものになることを期待 したい. 謝辞 本文でも述べたように,本研究は玉川大学農 学部 小野正人助教授,同 ミツバチ科学研究施 設 佐々木哲彦助教授,東京大学大学院医学系 研究科 野本明男教授,大岡静衣助手 との共同 研究 として行なわれました.また本研究は一部, 生物系特定産業技術研究支援センター,文部科 学省,(財)テルモ科学技術振興財団,内藤記 念科学振興財団からの研究助成によって行ない ました.この場をお借 りして深 く感謝申し上げ ます.最後に,「ミツバチ科学」誌上で私 ども の研究を紹介させていただく機会を下さった中 村純先生に,深甚なる感謝の意を表 します. (〒 113-0033文京 区本郷7-3-1 東京大学大学院理学系研究科) 引用文献
Breed,M D.CERobinsonandJREPage1990
Behav.Ecol.SoclOblOl.27(6):395-401
Dade,H A・1962・AnatomyanddlSSeCtionorthe honeybeelBRA,Cardiff164PPI
Fujiyuki.T・,H Takeuchi,M・Ono.Sl0hka.T・Sasaki. A.Nomoto.andT.Kubo.2004JVirol.78 (3):
1093-1100.
Guzman-Novoa.E・andR・E・Page・1994・Am・BeeJ・
134:101-106.
Ilunt,C.J..E.Cuzman-Novoa,M K.FondrkandJ.R. EPage1998Genetics148(3)1203-1213
Kamikouchi,A.H Takeuchi.M Sawata,K Ohashi, SNatoriandT・Kubo・1998・Biochem・Biophys・ Res.Commun.242(け 181-186.
Kamikouchl.A..H.Takeuchl,M.Sawata.S.Natorl, andT.Kubo.2000∫Comp Neuro1417 (4)
50ト510.
Liang.P andA B Pardee・1992Science257
(5072).967-971.
Lobo,N.F..LQTon,C.A.Hill,C.Emore.JRomer o-Severson.G.J.HuntandF.H Collins2003.
GenomeRes13(12)2588-2593
Moore.J 2002Par.aslteSandthebehaviorofani
-mals.OxfわrdUnlV.Press,NewYork.332pp. 野 本 明 男.1999.ピコルナウイルスと感染症.標準
微 生 物 学 第7版 (平松啓一 ・山西弘一編 ).pp
418-425.
Park.IM..T.KuniedaandT.Kubo.2003.1.Biol.
坂上昭一. 1983 ミツバチの 世界.岩波書店,東京. 221pp
Sawata,M,D YoshlnO,H Takeuchi,A.Kamikouchi,
K OhashiandT Kubo2002RNA8(6):772-785
竹 内秀 明 ・上川内あづ さ ・鹿毛枝 里子 ・久保健 雄
2001.ミツバチ科学22(3).113-120
Takeuchi.H.E Rage.M Sawata.A Kamlkouchi,
K Ohashi.M・Ohara,T FuJlyukl.T Kunleda.K・
Sekimizu,S NatoriandT.Kubo.T.2001.Insect
MoIBiol10(5)487-494
Takeuchi.H.T FuJlyuki,K・Shlrai.Y・Matsuo,A・
Kamikouchi.Y FujlnaWa.A Kato.A・Tsujimoto
andT.Kubo.2002 FEBSLeft513(213)・230-234.
Takeuchi.H..A.Yasuda.Y.Yasuda-Kamatanl,M.
Sawata.Y Matsuo,A Kato,A TsLU1mOtO.T
Naka-」imaandT.Kubo 2004 CellTISSueRes 316(2)
281-293.
wilson,E 0 1999.社会生物学 (合本版),新思索社,
東泉 1341pp
Winston,M L・1987・Thebiologyofthehoneybee,
HarvardUnivPress.Cambridge281pp
ToMOKOFUJtYUKl,HIDEAKITAKEUCHIandTAKEOKuBO
Kakugovirusidentifiedinthebralr10ftheaggressIVe
workerhoneybee.HoneybeScience(2004)25(4)
145-151DepartmentoFBiologlCalSciences,Gr
adu-ateSchoolofScience.TheUniversityofToky071311,
Hongo.Bunkyo-ku,Tokyo.113-0033Japan・
WorkersoftheEuropeanhoneybee(ApISme/
-IIFel-aL.)show altruisticaggressivebehaviorsto
defendthelrCOlonyfrom naturalenemies,suchas
hornetsThemolecularmechanism underlylngthe
aggressivenessoFthehoneybeesinthebrai
n,how-ever.remainsmostlyunknown Recentlyweldenti一
RedanovelRNA.termedKakL/gO,thatwasdetected
exclusivelylnthebrainsoH hehoneybeeworkers
thathadattackedaglanthornet(I/
e
spamandarJ'nJ'ajaponlCa)ThenucleotldesequenceoftheKakugo
RNA hadslgnIFicanthomologytothoseOFRNA
genomesofvarlOuSPICOrna-likevir
uses.espe-ciallysacbroodvirus.WhlChInfectsthehoneybeesI
KakL/gORNALnthetissuelysaleofinfectedworkers
wasseparatedatthesamesedimentationcoefficient
withvirionpartlCleofpjcorna-likevirusFurther一
more.whenthetissuelysateoftheinfectedworkers
waslnJeCtedintonon-infectedworkers,theKakLlgO
RNA wasamplifiedjn vJ'vo.Theseresultsdeml
onstratethatKakugoRNAisagenomeofanovel
picornalIikevirus(KakugovlruS)KakugoRNAwas
detectedinthebrainsoFtheaggressiveworkersbut
notinnursebeesorroragers,Suggestingthebrains
oftheaggressiveworkersareselectivelyInfectedby
thisnovelvirusHerewesummarizetheprocessto
identifyKakugovirusanddiscusspossiblerelation
betweenKakugoviralinfectioninthebrainandthe
aggressIVebehaviororthehoneybeeworkers We
alsodiscussposslblepathogenicityoFKakugovirus