けるシナプス可塑性の役割
藤田 一寿
電気通信大学 情報システム学研究科 情報ネットワーク学専攻 博士 ( 工学 ) の学位申請論文
2007 年 3 月
プス可塑性の役割
博士論文審査委員
主査 中村 整 教授 委員 出澤 正徳 教授 委員 吉永 努 助教授 委員 阪口 豊 助教授
委員 樫森 与志喜 助教授
藤田 一寿
2007 年
tion relevant to localization behaviors
Kazuhisa Fujita Abstract
Stimuli received by an animal include several sensory information and noise. An an- imal has to extract behaviorally-relevant features from stimuli in order to know the situ- ation of enviroment. The problem is how efficiently synaptic plasticity of neural system is used to extract the features from stimuli including several infomation and noise. To investigate this issue, locarization behavior of some animals is focused, because stim- ulus relating with this behavior is relatively simple and there are many experimental studies of locarization behavior. In this paper, two locarization behaviors are discussed.
One is sound localization of barn owl. Another is electrolocation of a weakly electric fish. This study shows the roles of synaptic plasticity in extracting sensory informatiton relevant to locarization behavior using computer simulation.
Barn owl can detect the direction of a sound source in the dark using auditory cues alone, so called sound localization. Two characteristics of sound stimulus, ITD (inter- aural time diffrence) and ILD (interaural leval difference), are used to locarize a sound sound source. The characteristics exhibit complex changes depending on location of sound source. This study proposes a neural model for decoding the orientation of a sound source, in which ITD and ILD information are bound by synaptic plasticity based on Hebbian learning.
Weakly electric fish can detect an object by analyzing weakly modulated electric stimulus, so called electrolocation. This study presents three results. In the first re- sult, synapses which have optimal time constant between receptors and electrosensory lateral-line lobe (ELL) extract spike synchronicity of receptor afferents elicited by mod- ulated stimuli, and features of modulated stimuli are effeciently encoded by ELL neu- rons. In the second result, burst firing and feedback circuit of ELL enable the ELL
systems.
性の役割
藤田 一寿 概要
動物が受ける刺激の中には、複数の行動に関連する様々な情報がノイズと共に 含まれている。さらに、刺激は一定ではなく周囲の状況に依存して常に変化して いる。このような状況下で、その様な刺激の中から動物は行動に必要な情報を脳 内の様々な部位で適切に処理し、統合しなければならない。このような刺激処理の 神経メカニズムを探求するためには、動物の定位行動はよい研究対象となる。な ぜならば、定位行動では動物の様々な行動の中で刺激と行動の関係が比較的明瞭 なためである。そのような複雑な刺激の中から定位行動に必要な情報を抽出する には、脳の神経ネットワークは常に可塑的に変化しなければならない。そこで、神 経ネットワークのシナプスが刺激に応じて様々に変化することに着目し、そのシ ナプス可塑性が情報の抽出統合を実現するために重要な役割をしていると考えた。
本研究では、シナプス可塑性が動物の定位活動においてどのような機能的役割を 果たしているのかを調べるために、実験的知見の豊富な面フクロウの聴覚システ ムによる音源定位と弱電気魚の電気感覚システムによる電気定位を対象に、それ ぞれの神経ネットワークモデルを構築しシミュレーションを行うことで、定位行動 に必要な情報の抽出におけるシナプス可塑性の機能的役割について研究を行った。
面フクロウは音刺激から音源の方向を特定することが可能である。面フクロウ が受ける音刺激は、顔や耳の形状や材質により複雑に変化をする。そのため、音 情報は周波数ごとに異なった複雑な方向依存性を有する。その複雑な音刺激から 情報を抽出統合して、音源の方向を特定することができる。本研究では、音源定 位を実現するために、視覚からのトップダウン信号によるHebb則に基づいた長期 シナプス可塑性を用いて、音の位相差と音圧差情報を統合するモデルを構築した。
その結果、複雑な方向依存性を持つ音刺激から音源の方向を効果的に定位できる ことを示した。
弱電気魚は自らが生成した交流電場とその変調を電気刺激として電気受容器で
からの物体情報を抽出し電気定位を行っている。電気定位におけるシナプス可塑 性の役割を調べるために、電気感覚システムの解剖学的生理学的な知見に基づい た神経ネットワークモデルを構築し研究を行った。
まず、始めに電気刺激を処理する電気受容器から上位核である電気感覚側線葉 (ELL: electro sensory lateral line lobe)における神経メカニズムについて研究を行い 以下のことが分かった。電気受容器は物体による電気刺激の変調を受けると活性 が微少に増加する。その活性の微少増加により、物体が存在した場合ELLの電気 受容器層に対する受容野内の電気受容器の同期性は増加する。その発火同期性の 増加を適切な時定数を有するシナプスで処理することにより、物体により誘発さ れた微少な電気刺激の変調の抽出が可能となることを示した。
ELLには複数フィードバック経路が存在する。次に、ELLとそのフィードバッ ク回路により物体の特徴抽出がどのように行われるかそのメカニズムについて研 究をおこなった。その物体の特徴抽出において、フィードバック信号に依存して 変化する時間スケールの異なる2つのシナプス可塑性が重要な役割を果たす。時 間的変化の遅いバックグラウンド信号は長期シナプス可塑性により抑えられ、一 方、速い変化の短期シナプス可塑性により後脳の神経細胞の利得を制御する。そ れにより、後脳のバースト発火が安定に発生し、物体の距離と大きさの情報をこ のバースト発火領域とバースト間のインターバルにより効果的に抽出できること が分かった。
動物の受ける刺激には複数の行動情報を同時に含む場合がある。弱電気魚の場 合も定位する物体情報だけではなく、同種の魚とのコミュニケーションの情報が 同時に刺激に含まれることがある。このとき、各行動に対応した情報の分離、抽 出においてシナプス可塑性がどのような役割をしているのか研究を行った。その 結果、短期シナプス可塑性の促進、減衰のダイナミクスにより、物体情報とコミュ ニケーション情報をうまくよりわけることが可能であることを示した。これらの 結果は短期可塑性をもつダイナミックシナプスの機能的役割の理解に新たな知見 を与えるものである。
以上のように本研究では、刺激の特徴によりシナプスを可塑的に変化させ、神経 ネットワークの活動状態を調節することで、単一刺激の中に含まれる定位行動に 必要な情報を分離抽出することが可能であることを示した。本研究の成果は、定 位行動の感覚情報処理における神経ネットワークレベルでのシナプス可塑性の役
第1章 序論 1
第2章 面フクロウの音源定位行動に関係する情報抽出におけるシナプス可塑
性の役割 5
2.1 はじめに . . . 5
2.2 これまでの知見と問題 . . . 7
2.2.1 音の両耳到着時間差ITDの処理 . . . 7
2.2.2 両耳音圧差ILDの処理 . . . 7
2.2.3 音情報の複雑な周波数依存性 . . . 10
2.2.4 問題. . . 10
2.3 ICclsのITD、ILD統合モデル . . . 10
2.3.1 ICc lsシステムのニューラルネットワークモデル . . . 12
2.3.2 ICc lsモデルの数理的表現 . . . 12
2.3.3 結果. . . 14
2.4 ICxモデル . . . 16
2.4.1 結果. . . 17
2.5 結論 . . . 19
第3章 弱電気魚の電気感覚システムの概要とその様々な知見 20 3.1 弱電気魚とは . . . 20
3.2 弱電気魚の4種の行動にかかわる情報処理方法. . . 22
3.2.1 エレクトロロケーション行動 . . . 22
3.2.2 物体追尾行動 . . . 22
3.2.3 混信回避行動(Jamming Avoidance Response : JAR) . . . 22
3.2.4 エレクトロコミュニケーション行動 . . . 22
3.3 電気感覚システムの概要 . . . 23
3.4 弱電気魚が受ける電気刺激 . . . 26
. . . .
第4章 ノイズを伴った刺激からの微小差異抽出におけるシナプス時定数の効
果 40
4.1 始めに . . . 40
4.2 これまでの知見と問題 . . . 40
4.3 モデル . . . 41
4.3.1 電気刺激のモデル . . . 41
4.3.2 電気受容器のモデル . . . 42
4.3.3 シナプスのモデル . . . 43
4.3.4 ELLの錐体細胞のモデル . . . 43
4.4 結果 . . . 43
4.4.1 発火同期性の検証 . . . 47
4.5 結論 . . . 48
第5章 電気定位の物体情報の抽出におけるシナプスダイナミクス 49 5.1 はじめに . . . 49
5.2 今までの知見と問題 . . . 49
5.3 モデル. . . 50
5.3.1 錐体細胞のモデル . . . 51
5.3.2 NP、EGPのモデル . . . 52
5.4 結果 . . . 53
5.4.1 ELLの応答. . . 53
5.4.2 バーストの特徴 . . . 56
5.4.3 シナプスの可塑性の役割 . . . 56
5.5 結論 . . . 57
第6章 刺激特徴抽出における短期シナプス可塑性の機能的役割 60 6.1 はじめに . . . 60
. . . .
6.3.5 TSの神経細胞のモデル . . . 65
6.4 結果 . . . 66
6.4.1 弱電気魚が受ける刺激 . . . 66
6.4.2 各ネットワークの応答 . . . 67
6.4.3 シナプス後電位の応答 . . . 70
6.4.4 単一細胞の応答 . . . 71
6.4.5 コミュニケーション情報の抽出 . . . 71
6.4.6 物体のEOD変調のみの時の応答 . . . 73
6.5 結論 . . . 74
第7章 結論と議論 75 7.1 結論 . . . 75
7.1.1 面フクロウ . . . 75
7.1.2 弱電気魚 . . . 75
7.2 議論 . . . 77
一般用語の省略形
ISI inter spike interval インタースパイクインターバル
PSP post synaptic potential シナプス後電位 PSTH peri stimulus time histgram PSTヒストグラム
メンフクロウの用語の省略形
IC inferior colliculus 下丘
ICc central nucleus of inferior colliculus 下丘中心核 ICcls lateral shell of central nucleus of inferior colliculus 側面下丘中心核 ICx external nucleus of inferior colliculus 下丘外側核
ILD interaural lebel difference 両耳音圧差
IPD interaural phase difference 両耳位相時間差
ITD interaural time difference 両耳到着時間差
NA nucleus anguralis 角状核
NL nucleus laminaris 層状核
NM nucleus magnocellularis 大細胞核
OT optic tectum 視蓋
VLVp nucleus ventralis lemnisci lateralis pars posterior 後外側毛帯核
bp cell basilar pyramidal cell 錐体細胞 EGP eminentialis granularis posterior 顆粒隆起後部 ELL electrosensory lateral line lobe 電気感覚側線葉
EOD electric organ discharge 発電器官の発電
EOD AM EOD amplitude modulation EODの振幅変調 JAR jamming avoidance response 混信回避応答
NP nucleus praeeminetialis 隆起前域核
OT optic tectum 視蓋
TS torus semicircuralis 半円隆起
動物の受ける刺激の中には、複数の行動に関連する様々な情報がノイズと共に 含まれている。さらに、刺激は時空間的に一定ではなく、周囲の状況に依存して 常に変化している。このような状況下で、動物はその中から行動に必要な感覚情 報を、脳内の様々な部位で適切に処理し統合しなければならない。このような刺 激処理の神経メカニズムを探求するためには、動物の定位行動はよい研究対象と なる。なぜならば、定位行動は動物の様々な行動の中で刺激と行動の関係が比較 的明瞭なためである。そのような複雑な刺激の中から定位行動に必要な情報を抽 出するには、脳の神経ネットワークが常に可塑的に変化しなければならない。そ こで、神経ネットワークのシナプスが刺激に応じて様々に変化することに着目し、
そのシナプス可塑性が情報の抽出統合を実現するために重要な役割をしていると 考えた。本研究では、シナプス可塑性が動物の定位活動においてどのような機能 的役割を果たしているのかを調べるために、実験的知見の豊富な面フクロウの聴 覚システムによる音源定位と弱電気魚の電気感覚システムによる電気定位を対象 に、それぞれの神経ネットワークモデルを構築しシミュレーションを行った。そ の結果、定位行動に必要な情報の抽出におけるシナプス可塑性の機能的役割につ いて様々な知見が得られた。
第2章では面フクロウの音源定位について言及する。
メンフクロウは音源から発せられた音刺激から音源の方向を特定することがで きる。その能力は音源定位と呼ばれている。面フクロウの音処理システムでは、音 情報を音の両耳到着時間差(interaural time difference: ITD)と両耳音圧差(interaural level difference: ILD)とに分けて処理する[34]。それぞれの情報は神経経路のいく つかの段階まで周波数ごとに並列に処理される。ITD、ILD情報は側面下丘中心核 (lateral shell of Central nucleus of inferior colliculus: ICcls)で周波数ごとに統合され
に複雑な変化を示す[6]。その周波数によるITD、ILDの依存性は複雑で、単純な 加算統合では必ずしも位置が特定できるとは限らない。音源の方向に対する正し い情報を得るには、OTからの視覚情報に基づいて、ICclsとICxの間でシナプス 可塑性による学習が起こらなければならないと考える。なぜならば、成長や環境 の変化により生じる、ITDとILDの方向に対する複雑な依存性の変化に対応する ためには、音情報と視覚情報との整合性を常に確認し、音情報の方向依存性が変 化しても適応しなければならないからである。本研究では、この考え方に基づき、
ICclsとICxの神経ネットワークモデルを構築し、視覚情報と聴覚情報を統合する
際の周波数特異的な音情報処理におけるシナプス可塑性の役割を提案する。
以降の章では弱電気魚の電気定位について言及する。第3章では、弱電気魚の 電気感覚システムの概要について言及し、電気刺激の中に物体の情報がどの様に 含まれるかについて提案する。弱電気魚は自らが生成した交流電場(electric organ
discharge: EOD)とその変調を電気刺激として電気受容器で受け、その情報を脳で処
理することで様々な情報を得ることが出来る。電気感覚により得た情報から、物体 の位置や大きさなどの認識(電気定位)や他の同種の魚とのコミュニケーションなど の行動を行うことができる。そのEODはまず電気受容器によりスパイクにコード される。その情報は第一次処理核である電気感覚側線葉(electrosensory lateral line
lobe: ELL)で処理される。そのELLは時間的特性と受容野の異なる4つのセグメ
ントに分かれている。それらで処理された情報は半円隆起(torus semicircuralis: TS) で統合される。本章では、定位する物体の側方距離と大きさの情報が魚の周りの 電場のどのような特徴によって表されるか調べるために、魚の周囲の電場計算モ デルを作成した。そのモデルを用いて、物体の距離や大きさの情報がEOD変調の 最大振幅と半値幅という特徴に含まれることが分かった。
第4章ではEODを処理する電気受容器から上位核である電気感覚側線葉(ELL) における神経メカニズムについて言及する。電気受容器は、通常は魚が発する自身 のEODを受けて、不規則に発火している。この不規則さは刺激のノイズや受容器 の感度に由来している。電気受容器は物体によるEOD変調を受けると活性が微少 に増加する。その活性の微少増加により、物体が存在した場合ELLの電気受容器 層に対する受容野内の電気受容器からのスパイク出力の同期性はわずかに増加す る。その発火同期性の増加を適切な時定数を有するシナプスで処理することによ り、物体により誘発された微少な電気刺激の変調の抽出が可能となることを示す。
第5章では電気感覚システムにおいて時空間的に変化する電気刺激からどの様 に物体の情報を抽出し表現するか提案する。動物がどのようにして刺激の時空間
的変化から行動に必要な情報を抽出しているのかは重要な問題である。特に定位 行動においては、定位されなければならない物体は相対的に静止している物体で あることは少なく、多くの場合では物体や動物自体が移動していると考えられる。
このような移動によって、動物が受ける刺激は時空間的に変化する。このように、
時空間的に変化する刺激をうまくエンコードするためには神経細胞やネットワー クのダイナミックな特性をうまく調節しなければならない。神経細胞のダイナミッ クな特性がどのようにしてその刺激から物体の特徴を取り出しているのか問題で ある。この問題を具体的に考えるために、我々は弱電気魚の電気感覚システムに 注目した。弱電気魚は発電器官を用いEODを自身の回りに形成することができ る。そして、物体により変調した電場から物体の場所を特定することが可能であ る。我々は第一次処理核であるELLの神経ネットワークモデルを構築した。本章 では、物体の距離と大きさに依存して生じたEOD変調の特徴がELLのバースト 発火によりコードされることを示す。これらの特徴はバーストの発火範囲とその インターバルによって表される。その同期発火は、ELLへのフィードバックを介 したシナプスの興奮、抑制の短期変化によりELLの神経細胞の利得がコントロー ルされ、安定にEOD変調の特徴をELLで抽出することが可能となることを示す。
第6章では複数の行動に対応する情報を含んだ刺激から短期シナプス可塑性を 用いることで各行動に対応する情報を分離することが可能になることを示す。動 物は単一の行動に関する情報のみを受けている場合よりも、むしろ複数の行動に 関する情報を同時に受容している場合が多い。動物は複数の行動に関する情報を 含む刺激を同時に受けた場合、その中から特定の行動に必要な情報のみを抽出し なければならない。神経ネットワークでどのようにして複数の行動に由来する情 報を含む刺激から特定の行動に関連した情報を抽出するのか問題である。この問 題を研究するにあたり、弱電気魚はよいモデル系である。電気魚は電気刺激から 様々な情報を得ることができる。そして、弱電気魚は同時に物体定位、物体追尾、
混信回避、他の魚とのコミュニケーションなどの行動に関連した情報を同時に受 け取る。このとき、電気刺激であるEODにはこれらの行動に対応した情報が同時 に含まれる。本章において、物体の情報とコミュニケーションの情報が刺激のな かに同時に含まれる状況を考える。なぜならば、弱電気魚は必ずしも、物体から
ワークモデルを作成し、短期シナプス可塑性により、ELLでエンコードしたEOD 刺激の異なる時間情報をTSにおいて物体あるいはコミュニケーション情報のみを 抽出できることを提案する。
以上のように本研究では、刺激の特徴によりシナプスを可塑的に変化させ、神経 ネットワークの活動状態を調節することで、単一刺激の中に含まれる定位行動に 必要な情報を分離抽出することが可能であることを示した。本研究の成果は、定 位行動の感覚情報処理における神経ネットワークレベルでのシナプス可塑性の役 割を解明する上で、重要な知見を与えるものと考えられる。
関係する情報抽出におけるシ ナプス可塑性の役割
2.1 はじめに
定位行動に関連した情報は、複数の感覚刺激からもたらされる。我々人間も五 感から様々な刺激を受けている。特に、視覚と聴覚からの情報を用い、物体の定 位を行うことが出来る。その能力は人間に限ったものではなく、本章で取り扱う 面フクロウは聴覚による定位能力が特に優れている。面フクロウは聴覚情報を処 理することで高精度な音源の定位をすることが可能である。面フクロウの音源定 位は、耳で受け取る音の2つの特徴量を処理することで行われる。それは、両耳 に入る音源からの音の両耳到着時間差(ITD: interaural time difference)と両耳音圧 差(ILD: interaural level difference)である[34]。ITDとILDは音源の方向によって 変化する。おもに、ITDは音源の水平方向を表し、ILDは音の垂直方向を表すと 考えられている[34]。内耳(inner ear)に入った音刺激は、周波数選択性を持つ受容 器により周波数ごとに分解され、スパイクに変換される。その変換された情報は 図2.1のように、音の周波数ごとにITDとILDで別々の経路で並列に処理される。
別々に処理されたITD、ILDは側面下丘中心核(ICcls:lateral shell of Central nucleus of inferior colliculus)へ送られ、周波波数ごとに統合される。ICclsの情報は下丘外 側核(ICx: external nucleus of inferior colliculus)でさらに統合処理され、視蓋(OT:
optic tectum)に送られ、視覚情報と統合される。ICxとOTでは音源方向のマップ
が形成されている。本章ではICc coreとVLVpがどの様に周波数ごとに統合され、
さらに周波数ごとに処理された情報がどの様に統合されるのか提案する。
図2.1: 面フクロウの音情報の処理経路。ICc (central nucleus of inferior colliculus), VLVp (nucleus ventralis lemnisci lateralis pars posterior), ICcls (lateral shell of Central nucleus of inferior colliculus), ICx (external nucleus of inferior colliculus), OT (optic tectum)。
IC inferior colliculus 下丘
ICc central nucleus of inferior colliculus 下丘中心核 ICcls lateral shell of central nucleus of inferior colliculus 側面下丘中心核 ICx external nucleus of inferior colliculus 下丘外側核
ILD interaural lebel difference 両耳音圧差
IPD interaural phase difference 両耳位相時間差
ITD interaural time difference 両耳到着時間差
NA nucleus anguralis 角状核
NL nucleus laminaris 層状核
NM nucleus magnocellularis 大細胞核
OT optic tectum 視蓋
VLVp nucleus ventralis lemnisci lateralis pars posterior 後外側毛帯核 表2.1: メンフクロウの用語の省略形
2.2 これまでの知見と問題
2.2.1 音の両耳到着時間差 ITD の処理
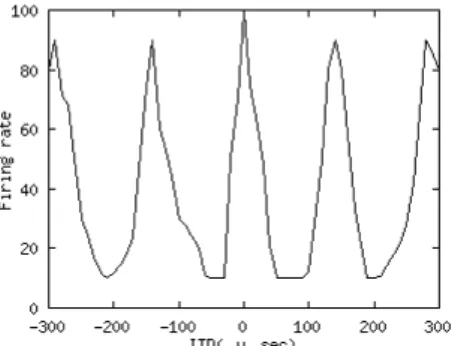
ITDは左右の耳に到着する音の位相のずれである。ITDは主に左右の耳の音源 からの距離の差で生じる。その時間差は左右の蝸牛で生じたスパイクの神経細胞 への到達時間の差を検出をすることでITDを特定している。その一致を検出する 神経細胞によりITDはICc coreで最終的に表現される。ICc coreでは特定のITD を表現する神経細胞がその表現するITDの値の昇順に並んでいる。周波数ごとに 処理されそのITDは音の周期ごとにピークを持つ(図2.2)。それを位相多義性とい う。そしてそのピークはITDにあわせてシフトする。ICc coreのシミュレーション 実験においても同様に、ITDが変化すると形を保ちながらシフトする(図2.3、
図2.4)。
図2.2: ITDに対するICc coreの実際の応答。横軸は神経細胞が表すITD。[20]
2.2.2 両耳音圧差 ILD の処理
図 2.3: 7kHzの音のITD=0µsecのと
きのICc coreの応答のシミュレーショ
ン結果。縦軸は発火率、横軸は神経細 胞が表すITD。
図2.4: 4kHzの音のITD=100µsecのと
きのICc coreの応答。縦軸は発火率、
横軸は神経細胞が表すITD。
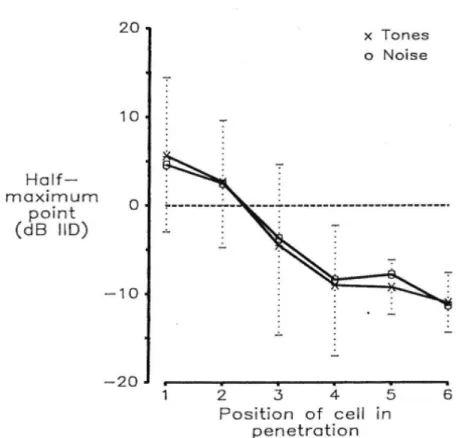
に対する応答がS字曲線になる。(図2.5)。ILDを変えていくとそのS字曲線の半 値の位置がシフトしていく(図2.6)。その実験的知見に基づき、神経ネットワーク モデルを構築するすることで左右のVLVpの発火率のギャップとして音圧差を抽出 することができることを以前の研究により示している[22, 30]。そのネットワーク モデルによるシミュレーション結果から、図2.7, 2.8のようにILDの変化に対し、
左右のVLVpの発火ギャップがシフトする。それは、片方の耳の音圧があがるた め、対側のVLVpにかかる抑制が大きくなるからである。
図2.5: ILDに対するVLVpの神経細胞の応答。横軸はILD、縦軸は発火率。Aは
背側の細胞の応答を表し、Cは腹側の細胞の応答を表す。実線、破線点線は耳に入 る音圧の平均を変えている。グラフのIID(interaural intensity defference)はILDと 同意。
図2.6: VLVpの神経細胞の場所とその応答の半値の関係。場所が異なると応答に 必要なILDの値がほぼ線形に変化する。
2.2.3 音情報の複雑な周波数依存性
内耳で周波数毎に分離され、処理されてきた音情報は、下丘中心核シェル(ICc ls)から下丘外側核(ICx)へ投射される際に統合される。統合された情報は視蓋(OT) へ送られる。OTでは、視覚情報とICxからの音情報が統合される。Brainardらに よりOTにおけるITDとILDのマップが計測された(図2.9) [6]。その実験で、OT
におけるITD、ILDマップは、各周波数で方向に対する応答特性が大きく異なる
ことが判明した。このような、同じ音源の方向に対して、各周波数で大きくITD、
ILDが異なり、さらに方向に対するITD、ILDの変化も各周波数で大きく異なる。
このような、周波数ごとに大きく異なる方向依存性を、単純な情報の重ね合わせ では、一意的に決定するのは難しいと考えられる。
2.2.4 問題
問題1 過去の研究によりILDが左右のVLVpの発火のギャップにより表現可能で あることが示された。しかし、このようにギャップとして表現されたILDと ITDがどの様にICclsで統合されるのか分かっていない。
問題2 面フクロウが基底膜で受け取るITD、ILDは音源の方向によって不規則か つ複雑に変化する。その結果OTにおけるITD、ILDのマップは周波数ごと に非常に異なり、不規則なものになっている[6]。その複雑なITD、ILDをど のように統合するのかはまだ明らかになっていない。
本章では、ICcにおけるITDとILD情報の統合メカニズムについて、神経モデ ルを構築し検証した。
2.3 ICcls の ITD 、 ILD 統合モデル
問題1に対しモデルを構築しシミュレーションをおこなった。別々の経路で検 出されてきたITDとILDは、ICc lsで初めて統合される。この統合における効果 的なメカニズムを解明するために、本研究ではPenaと小西の実験的事実[43]をも とに以下のモデルを提案する。
図 2.9: メンフクロウのOTにおけるIPD、ILDマップ。横軸は横方向の音源の方 向、縦軸は縦方向の音源の方向。IPD(interaural phase difference)は両耳位相差を表
2.3.1 ICc ls システムのニューラルネットワークモデル
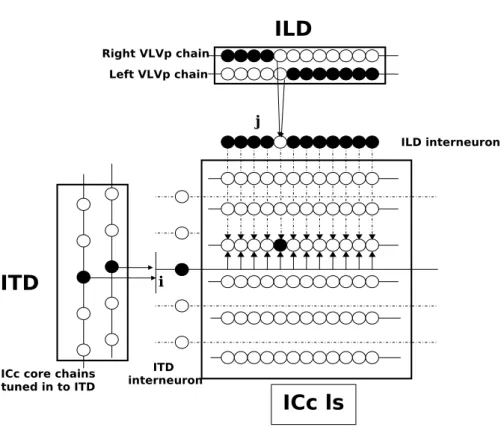
Penaと小西の実験により、ITD経路からの入力は、下丘中心核コア(ICc core)か ら興奮性として、ILD経路からの入力は、後外側毛帯核(nucleus ventralis lemnisci lateralis pars posterior: VLVp)から抑制性として入力されることが分かっている[43]。
図 2.10の左の縦に並ぶニューロン列はICc coreのITD検出ニューロン列を表す。
図2.10の黒塗りのニューロンは発火していることを示す。ICc coreからのインター ニューロンを介した興奮性入力は、ICc lsの碁盤目状に並んだニューロン列の横一 列全てに入力される(図2.10の矢印付き実線)。またこのとき同時に、図2.10の上 部に位置する左右VLVpからはインターニューロンを介してICc lsのニューロン 列に抑制性の信号が入力される(図2.10の矢印付き破線)。しかし、左右VLVpの ニューロン列には発火ギャップ(左右VLVpに存在する無発火のポジション)が存在 するので、そのギャップ内に位置するインターニューロンを介したICc lsのニュー ロンは、抑制されずに発火する。それによって、ICc lsでは、音刺激のITD, ILD の値に対応したニューロンのみが発火する。このように、ICc lsへの入力を全てイ ンターニューロンを介したものにすることで、Penaと小西の指摘する乗算的統合 を実現している。
ICc coreのインターニューロンから興奮性の入力が、横一列のICc lsのニューロ
ンに入力される。それと同時に、VLVpのインターニューロンからくる抑制性入 力によってICc lsのニューロンは抑制される。この結果、R-VLVpとL-VLVpの重 ならないギャップ内のインターニューロンを介したICc lsニューロンのみが抑制さ れずに発火する。ICc lsモデルではこのようにして、音源の水平方向の情報をもつ ITD情報と、音源の垂直方向の情報をもつILD情報が方向マップ上で統合される。
2.3.2 ICc ls モデルの数理的表現
ITDインターニューロン出力Prob(UITD(i;t)= 1)は、
Prob(UITD(i;t)=1)= 1
1+exp[−(VITD(i;t)−Vthr)/hvr], (2.1)
UITD(i;t)=
1 rand≤ Prob(UITD(i;t)=1) 0 otherwise
(2.2) と表す。
図2.10: 音源定位のモデル。矢印付き実線は興奮性接続を表す。矢印付き点線は抑 制性接続を表す。
ILDインターニューロン出力Prob(UILD(i;t)= 1)は Prob(UILD(i;t)= 1)= 1
1+exp[−(VILD(i;t)−Vthr)/hvr] (2.3)
UILD(i;t)=
1 rand≤ Prob(UILD(i;t)=1) 0 otherwise
(2.4) と表す。
ICc ls (i, j)ニューロンの膜電位は τm
dVs(i, j;t)
dt =−(Vs(i, j;t)−Vrest)+EITD(i;t)−IILD(j;t)) (2.5) と表す。
Vs(i, j;t) ICc core (i, j)ニューロン の膜電位 Vrest ニューロンの静止電位
EITD(i;t) ICc coreithインターニューロン の興奮性入力 IILD(j;t) VLVp jthインターニューロン の抑制性入力 τm 500に設定する
ICc ls(i, j)ニューロンの出力Us(i, j;t)の式:
Us(i, j;t)= 1
1+exp[−(Vs(l, j;k)−Vthr)/hvr]. (2.6) ICc lsモデルの発火頻度Fls(ω,ITD,ILD;i, j)の式:
Fls(ω,ITD,ILD;i, j)= 1 nmax
nmax
∑
n=1
Us(i, j;t). (2.7)
2.3.3 結果
いくつかの周波数、ITD、ILDの組み合わせについて以下に例を挙げる。各組み 合わせの図は、左からICc coreからの入力、VLVpからの入力、ICc lsにおける出 力の順に並べている。全てのICc lsの結果についてITD情報に起因する位相多義 性はまだ存在したままで、本来音源が存在するポジション以外のニューロンも高 い発火頻度を示している。この位相多義性はICxにおける各周波数の情報統合の 際に解消されることが知られている。
図2.11: 4kHz, ITD=100µsec, ILD=0dBの場合のICc lsの出力
図2.12: 7kHz, ITD=0µsec, ILD=8dBの場合のICc lsの出力
2.4 ICx モデル
本モデルはICxとICclsで構成される(図2.13)。ICclsは周波数ごとに層をもつ。
その層には各周波数ごとのITDとILDが図 2.10のように入力される。そして、
ICclsの神経細胞はITDとILDの両方が入力されている場合発火する。このこと
は、小西らの実験によるICclsはITDとILDが積算的統合されるという結果に基 づいた。ICclsは周波数ごとに情報を処理し、本モデルでは3種類の周波数を処理 するため、3層とした。
図2.13: 音源定位のモデル。ICclsは二次元の格子状に神経細胞が配置され、その
神経細胞はそれぞれ周波数ごとのITDとILDを表す。ICxでも神経細胞が二次元 に配置され、その神経細胞は物体の方向を表す。
ICxの神経細胞のモデルはleaky integraterを用いた(式(2.8))。
τdV
dt =−V +Input. (2.8)
ICxの神経細胞iの入力Inputiは、神経細胞iがICclsの神経細胞jと接続してい るとすると、
Inputi =w(t)Xj, (2.9)
となる。ICxとICcls間、そしてICcls層間のシナプス結合強度w(t)はHebb則に基 づいた学習を行う。神経細胞iと神経細胞 jとの間のw(t)は
τdw(t)
dt = −w(t)+λXiXj, (2.10)
となる。λは学習率。 Xは出力であり、次の式で決まっている。
X= 1
1+exp(−V−ϑε ). (2.11)
2.4.1 結果
図2.14は物体の方向(0, 0)のときの単純にICclsの出力を加算統合結果である。
位相多義性が消えず、ICxの出力が一意的に決まらない。これは、Hebb則による 学習を行わない状態では、音の情報だけでは物体の方向が決定できないことをあ らわす。
図2.14: ICclsの出力を単純にあわせたもの。左はICclsの周波数ごとの層の応答、
右はICxの応答。ICxにおいて一点に場所が決まらない。
次に3つの位置の3つの周波数について、ネットワークに位置を学習させた。学 習後、物体の方向(0, 0)の入力を加えると図2.15のように一点にきまる。その時、
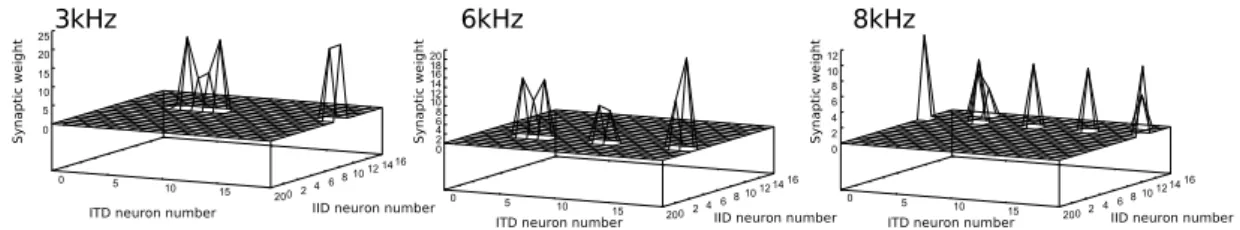
ICxの発火した神経細胞に対するICclsの結合強度は図2.16のように成長する。
図2.15:学習後、入力(0, 0)を与えた結果。左はICclsの周波数ごとの層の応答、右 はICxの応答。ICxの応答が一点でおこり、ICx上で音源の方向が一点にきまる。
図2.16: 発火したICxの神経細胞へのICclsとの結合強度。
2.5 結論
VLVpからICclsへの抑制性接続とインターニューロンを介在させることにより、
左右のVLVpの発火ギャップとしてコードされたILDとITDを統合することが可 能となることを示した。
Knudsenらはプリズムを用い視覚情報をシフトさせることで、ICxとICcls間で
接続が変わることを示した[37]。これは視覚情報と聴覚情報が不一致を起こした
ときICxとICcls間でその不一致を補正するということである(図2.17)。本モデル
はプリズムによる視覚情報の変化への適応と、ITD、ILDの周波数特異性の解決の メカニズムを説明することが可能である。本研究の結果、ICxとICcls間の可塑性 は、ICclsではITD、ILDが周波数ごとに異なった音源の方向依存性の解決の役割 をはたしていることを説明した。この可塑性により環境の変化や成長によりITD、
ILDの方向への依存性が変わっても対応できることを示した。
音源定位の実験では、音源の方向はITDとILD情報をフィードフォワードの流 れに従って階層的な処理によって決められていると考えられてきた。本研究の成果 は、むしろ音源の方向は視覚と聴覚の整合性によって決まるもので、それがITD、
ILDの複雑な周波数特異性を解消するのに役立っていることを示したことである。
本研究の成果は視覚と聴覚を用いた外界認識の問題に対して重要な知見を与える ものである。
図2.17: 実線の視覚情報が点線に変わったとき、接続もそれにより変化する
の概要とその様々な知見
3.1 弱電気魚とは
生物は生きていく上で必要な情報を得るために様々にその器官を進化させてき た。人間は視覚、聴覚、嗅覚、味覚、触覚を発達させてきた。しかし、生物の中 には、人間の持つ五感とは別の感覚器官を発達させたものもいる。弱電気魚は、
濁った川などに生息しているため視覚情報だけでは、十分に周囲の環境情報を得 る事はできない。そのため、周囲の状況を知るために弱電気魚は電気感覚を発達 させた。弱電気魚は、発電のしかたにより、ミリ秒単位の短い電気パルスをだす
パルス種(エレファントノーズフィッシュなど)と、連続的に周期波を出すウェーブ
種(アインゲンマニアなど)に分けられる。アマゾンに住むアイゲンマニア(図3.1)
や、アフリカに住むエレファントノーズフィッシュなどの弱電気魚は、細長くのび た尾の部分に興奮性の細胞を連ねた発電器官を持っており、ここから数ミリボル トの電気を発生させる(EOD: electric organ discharge)。アイゲンマニアの場合、発 生する電気は200–600Hzぐらいのサイン波型の交流電流で、通常は一定の周波数 で発電している。本研究では特にウェーブ種の弱電気魚を取り扱う。弱電気魚は、
発電器官により発生させた電気を用い、周囲に電界を形成する。その電気刺激か ら弱電気魚は様々な情報を得ることができる。例えば、魚の体の周辺に水とは異 なる電気抵抗の物体が存在した場合、物体によって電場が変調する。弱電気魚は その変調を感じ取り、それを脳内で処理することによって、視界の効かない暗闇 や濁った環境でも周囲の物体を検知できる。この能力をエレクトロロケーション (Electrolocation)という。
図3.1: ウェーブタイプの電気魚(アイゲンマニア)。サイン波型の電気を発する。
図 3.2: 弱電気魚は、尾部にある発電器官によって発せられる交流電流EODによ
3.2 弱電気魚の 4 種の行動にかかわる情報処理方法
弱電気魚が発生した放電電流EOD(Electric Organ Dsicharge)は、周囲の状況に よって変調される。変調されたEODにはエレクトロロケーション、物体追尾行動、
エレクトロコミュニケーション、混信回避などの行動に関する情報が含まれる。
3.2.1 エレクトロロケーション行動
エレクトロロケーションは、水と異なる電気抵抗をもった物体によってEODの 振幅が変調され、その歪みを捕らえる事で感知できる。この行動によって弱電気 魚は、視界の効かない暗闇や濁った環境でも、餌や障害物などの位置や大きさな どを検知することができる。
3.2.2 物体追尾行動
弱電気魚は、普段は岩陰や水草の間などの狭い場所に身を潜めている。そのよう な狭い場所に安定して身を潜めるために、弱電気魚は電気感覚で物体によるEOD 変調を受けようと物体の移動に伴って移動する。このような行動を物体追尾行動 という。
3.2.3 混信回避行動 (Jamming Avoidance Response : JAR)
ウェーブ種の電気魚では、電気定位のために常に発電を続けるが、個体によっ てそれぞれ発電の周波数が決まっていて、発電周波数は通常は変化しない。しか し、発電周波数の近い2尾が出会うと、互いに発電周波数を変更し相手の周波数 との差を広げる行動をとる。これは、発電周波数の近い相手との間で周囲に形成 される電場の干渉が起き、電気定位の能力がそこなわれるのを避けるために起こ る。この行動を混信回避行動(jamming avoidance response: JAR)と呼ぶ。
3.2.4 エレクトロコミュニケーション行動
オスとメスの2匹の弱電気魚が求愛行動をする場合やオス同士が縄張りに入って きた相手を威嚇する場合などに、自分のEODに高い周波数の一過性のInterruption を作り出し、仲間の魚とコミュニケーションをとる。
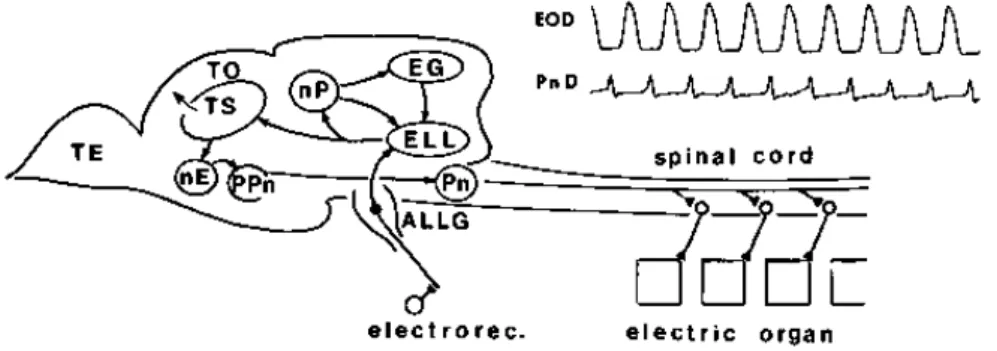
3.3 電気感覚システムの概要
弱電気魚がEODの振幅変調を処理し物体との距離を検知するための電気感覚シ ステムの構造については、解剖学的、生理学的研究によって多くの実験データが 得られている。図3.3に示すように、EOD振幅変調を体表面上に分布した電気感 覚受容器で受け取り、スパイクに変換する。EOD振幅変調は物体の魚からの距離、
大きさ、形、材質などによって異なり、電気感覚受容器で受ける電気刺激も異な る。電気受容器で変換された情報は、求心性神経線維を伝わり、後脳の電気感覚 側線葉(ELL: Electro sensory lateral-line lobe)へ送られる。ELLは4つのセグメン トに分かれている。それぞれのセグメントは電気受容器に対する受容野の大きさ と時間的特性が異なっている。ELLでは求心性神経線維より送られてきたスパイ クを受け取る。ELLでは、体表面の各位置に置かれた物体によって引き起こされ るEODの振幅変調を、体表面上の位置に一対一に対応する(ソマトトピカルな)脳 内マップで処理する。左右のELLはその出力を半円隆起(TS: Torus semisurcularis) に送り、電位の振幅変化や位相変化をそれぞれ処理し、更に上位の視蓋(OT: Optic
Tectum)でひとつに統合される(図3.4)。体表面上で受容した情報はソマトトピカ
ル(位置保存的)に上位の神経核に送られる(図3.4)。
図3.3: 弱電気魚の脳の断面図(各器官の空間的位置関係を示す)。発電器官によっ て集められたEOD AM情報は、後脳にある電気感覚側線葉(ELL)に集められる。
そこで処理された情報は半円隆起(TS)に送られて物体の位置が認識される。
bp cell basilar pyramidal cell 錐体細胞 EGP eminentialis granularis posterior 顆粒隆起後部 ELL electrosensory lateral line lobe 電気感覚側線葉
EOD electric organ discharge 発電器官の発電
EOD AM EOD amplitude modulation EODの振幅変調 JAR jamming avoidance response 混信回避応答
NP nucleus praeeminetialis 隆起前域核
OT optic tectum 視蓋
TS torus semicircuralis 半円隆起
表3.1: 弱電気魚の用語の省略形
3.4 弱電気魚が受ける電気刺激
3.4.1 弱電気魚の電気刺激の実験的知見
弱電気魚のアイゲンマニアは、常に尾部の発電器官より一定の交流電流を発生 し、周囲に一定の電場を作っている。その電場に物体があるときEODの振幅変調 (EOD AM : EOD Amplitude Modulation)が起こる(図 3.5)。J. Bastianによる実験 では、弱電気魚の近くに直径12mm、長さ5mmの筒状の物体を配置し、そのとき のEOD の振幅変調を測定している(図 3.6A-C) [3]。さらに、物体の半径を4, 8, 12mmと変えたときのEODの振幅変調の様子を測定している(図3.6D-F)。それら
のBastianの実験結果は、物体の半径が大きくなるにつれて、EOD振幅変調も大
きくなっていくことを示している。つまり、魚は物体の大きさをEODの振幅変調 の大きさとして捕らえていることが分かる。次に魚の側面から一定の距離(1, 3, 6,
10mm)で物体の直径を徐々に大きくしていった場合の電位の変化を図3.7Aで分か
るに魚からの距離に関係なく、物体の直径が大きくなるにつれて、EODの振幅変 調も大きくなっていくのが分かる。また、魚と物体の距離が近づくにつれてポテ ンシャルの変化も大きい。次に物体の直径は一定として、(4, 8, 12mm)、魚の側面 からの距離を徐々に大きくしていった場合のポテンシャル変化を図3.7Bに示す。
物体が、魚から離れるにつれて、徐々にポテンシャルは小さくなっていく。また、
Von der Emdeらの実験では、物体の大きさを様々に変えたところEOD変調の最大
勾配により表されると示唆している[47]。
図3.5: EODの振幅変調の様子。自ら発したEODの振幅の変調に様々な情報が含
まれる。
図3.6: 物体によるEODの振幅変調
図 3.7: (A)物体の半径と電位の変化の関係。物体の半径が大きくなるにつれて電
3.4.2 弱電気魚が受ける電気刺激の数値計算
弱電気魚は、発電器官により電気を発生し周囲に電界を作る。生体実験では、実 際の水槽内のすべての位置におけるポテンシャルを測定することは不可能である。
そのため、Heiligenberg [28]、Hoshimiyaら[31]によって、二次元平面における電 気魚の周囲のポテンシャルの計算が行われた。本研究では、彼らのモデルを参考 にし、三次元空間に拡張した。
弱電気魚がつくるEODによるポテンシャルを計算するために、本研究では、次 のようなモデルを提案する。図3.8に示すように弱電気魚がいる3次元の水槽を考 える。水槽内を多数の小さな区画に区切る(図3.9)。その水槽の中央に電気魚を配 置する。現実には、魚は流線型の形をしているが、計算を簡略化にするためここ では長方形としている。このモデルでは長さの単位として1L=0.1cmとしている。
弱電気魚は、内部と体表面とでそれぞれ異なる電気抵抗を持っており、また表皮の 電気抵抗は式(3.4)であらわされるように、頭部から尾部に行くにしたがって、抵 抗が小さくなっている。ここで、ρskinは皮膚の電気抵抗、ρfishは体内の電気抵抗を 示す。電気魚の発電器官として、(X,Y,Z)= (182,50,15)に−100p.u.(p.u.=potential unit)(X,Y,Z) = (209,50,15)に+100p.u.の双極子を配置する(1p.u. = 0.125mv)。周 囲の水は、3800Ωの電気抵抗を持っている[31]。また、シミュレーションの目的 によって、魚の周囲に物体を配置し、その物体にも電気抵抗ρobjを与える。
電場の計算方法
弱電気魚が発生する交流電場により周囲に一定の電場が生じる。水槽内の(X,Y,Z) にあるセルの静電ポテンシャルU(X,Y,Z)を求める。図3.10に水槽内の一部のセ ルを示す。セルの中心部のポテンシャルを計算する場合を考える。中央のセルの ポテンシャルは、隣接するセル(合計6個)のポテンシャルより計算される。各セ ルには、水・魚(内部・皮膚)・物体といった種類があり、そのセルの抵抗や電気 容量はそれぞれ異なる。図3.10に示す回路のよって求められる電流保存則(キル ヒホッフの法則)により、隣接する各セルから中央のセルに流入する電流の総和は 0になる。これをもとに、中央のセルと隣接する6個のセルの回路により、中央 のポテンシャルを求める。中央のポテンシャルは、式3.1、3.2、3.3によりもとめ られる。EODは時間的に変化する電場を与えるので、セル間には変異電流も流れ ることになる。ここで、U(X,Y,Z)は(X,Y,Z)に中心があるセルのポテンシャル、
図3.8: EODの計算に用いた系。水槽の奥行きX、横Y、縦Zはそれぞれ200mm、
100mm、30mmとした。その中の魚の長さ、横幅、縦幅はそれぞれ80mm、8mm、
16mmとした。
z y
fish tank head
tail fish
object
R(X,Y,Z)、C(X,Y,Z)はそのセルが持つ電気抵抗と電気容量である。(X, Y)セルが 水、物体、魚の内部・皮膚によってR(X,Y)、C(X,Y)の値は決まる。この方程式を 以下のような環境状況のもとで、初期解を設定してセルフコンシテントに解くこ とによりU(X,Y,Z)を求める。
図3.10: 中央のセルのポテンシャルを電流保存則により求めるための図。ここで、
U(x,y,z)は、(X,Y,Z)に中心があるセルポテンシャル、R(x,y,z)、C(x,y,z)は、そ のセルが持つ電気抵抗と電気容量である。(x,y,z)セルが水、物体、魚の体内、皮 膚によってR(x,y,z)、C(x,y,z)の値が決まる。
∑
Z=z−1,z+1 X=x,Y=y
∑
Y=y−1,y+1 X=x,Z=z
∑
X=x−1,x+1 Y=y,Z=z
(IC+IR)=0 (3.1)
IC = (C(X,Y,Z)+C(x,y,z))× d
dt(U(X,Y,Z,t)−U(x,y,z,t)) (3.2)
IR = 1
R(X,Y,Y)+R(x,y,z,t) ×(U(X,Y,Z,t)−U(x,y,z,t)) (3.3) 水槽の境界は魚の電場はそこまで影響しないと考え、水が無限に広がっていると いうことを仮定して、境界条件U(X,Y,Z)= 0を用いる。また、EODの発生源であ
る、双極子の場所(182, 50, 15), (209, 50, 15)のセルのポテンシャルは、U(x,y,z)= +100p.u.、U(x,y,z) = −100p.u.とする。アイゲンマニアの皮膚の電気抵抗ρskinは
Hoshimiyaらが用いたものと同様で式3.4 で与えられる [31]。水と魚体内の抵抗
ρwaterとρfishの値は、ρwater =3.8×102[Ω·cm]とρfish =10.0[Ω·cm]である。
ρskin =
12000 (60< x<108) 6150−45×x (108< x <132) 1000 (133< x <140),
(3.4)
シミュレーション結果
弱電気魚は物体などによるEODの変調を処理することで周囲の状況を知ること ができる。そこでまず、物体が存在しない状況と、弱電気魚の側方から4mmの場 所((X,Y,Z)= (80,58,15))に抵抗ρobj = 3.8×10−4Ωの一辺の大きさ4mmの立方体 の物体を配置した状況において計算を行った。そのときのEODの様子を図3.11、
3.11に示す。物体が存在しない場合(図3.11)と、物体が存在する場合(図3.11)と を比較すると、物体の付近のEODは歪んでいることが分かる。
図 3.13は魚の体表面z = 15における場所xに対するEOD強度の変化である。
x= 60が魚の先頭を表し、x = 140が魚の末端を表す。図3.13の破線と実線の値 の差が、物体によるEODの変調である。図3.14は物体により生じたEOD変調の 強度の体表面状における分布である。EODの変調は物体の位置に伴って移動して いく(図3.14,図3.15)。
-2 0 -4 -6
-8
[p.u.]
図 3.11: 物体が存在しない場合の電気
魚が生成するEODの様子。
0 -4 -2 -6
-8
[p.u.]
図3.12: 物体が存在する場合のEODの
様子。物体周囲のEODは歪んでいる。
dV [p.u.]
x [mm]
with no object with one object
図3.13: 魚の体表面におけるEODの強度。実線は物体が無い場合、破線は物体が
有る場合。この差が物体によるEODの変調である。弱電期魚はこの小さな変調を 電気感覚システムにより抽出している。
60 65 70 75 80 85 90 95 100 8
10 12 14 16 18 20 22
60 65 70 75 80 85 90 95 100 8
10 12 14 16 18 20 22
0 0.6 1.2 1.8
60 65 70 75 80 85 90 95 100 8
10 12 14 16 18 20 22
ZOO
[OO
ZOO
[ O O
ZOO
[ OO
図3.14: 魚が受けるEOD変調の体表面状での強度分布。上図は物体がx=70に存
在する場合。中央の図は物体がx=80に存在する場合。下図は物体がx=90に存
65 70 75 80
85 90 95 100 location x (mm) 0
100 200
300 400
500 600
700 800
time (mse 0 c)
0.4 0.8 1.2 1.6
EOD AM (p.u)
図3.15: 物体が5cm/sで移動するときの位置xにある受容器が受けるEOD振幅変
調の時間変化。
物体の情報とEOD変調の特徴 EOD変調は物体の側方距離や大きさ(図3.16)に 依存し変化していく[3]。本モデルを用い、様々に物体の大きさと側方距離を変え、
シミュレーションを行った。まず、一辺6mmの立方体がx= 75に存在するとき、
魚からの側方距離を2–12mmまで2mmごとに変えて計算を行った。その結果、物 体が魚の近くにあるときは、魚が受けるEOD変調は大きくなり、物体が遠ざかる につれてEODの振幅変調は小さくなっていくことが分かった(図3.17)。次に側方 距離6mmに物体を配置し、一辺の長さを2–12mmに2mmごとに変えて計算を行っ た。この場合、物体が大きければ大きいほどEOD変調は大きくなることが分かっ
た(図3.18)。これらの結果は、Bastianなどが測定した結果に合致する。さらに、こ
れらのEOD変調の半値幅と物体の特徴の関係を調べた。図3.19、図3.20はEOD 変調の半値幅を比べやすくするため、EOD変調を規格化したものである。その結 果、側方距離が大きくなるほどEOD変調の半値幅は大きくなる(図3.19)。また、
物体の大きさが大きくなるほど半値幅も大きくなる(図 3.20)。様々な側方距離と 物体の大きさでシミュレーションを行い、図3.21に距離と最大振幅、半値幅の関 係を、図3.22に大きさと最大振幅、半値幅の関係を示す。物体により引き起こさ れたEOD変調は、側方距離を大きくしていくと最大振幅は小さくなり、半値幅は 大きくなる(図3.21)。また大きさを大きくしていくと、EOD変調の最大振幅は大 きくなり半値幅も大きくなる(図3.22)。
EOD AM [p.u.]
x
distance=2mm distance=4mm distance=6mm distance=8mm distance=10mm distance=12mm
図3.17: 一辺6mmの立方体が位置75に有る場合に側方距離を変えた場合のEOD
変調の変化。物体が魚の近くにあるときは、EOD変調は大きくなり、徐々に物体 が遠ざかるにつれて、EOD変調は小さくなっていく。
EOD AM [p.u.]
x
size=2mm size=4mm size=6mm size=8mm size=10mm size=12mm
図 3.18: 側方距離6mm、位置75に有る場合に物体の大きさを変えた場合のEOD
変調の変化。物体が大きければ大きいほどEOD変調は大きくなる。
normalized EOD AM
x
distance=2mm distance=4mm distance=6mm distance=8mm distance=10mm distance=12mm
図3.19: 一辺6mmの立方体が位置75に有る場合の側方距離と規格化したEOD変
調の関係。
normalized EOD AM
x
size=2mm size=4mm size=6mm size=8mm size=10mm size=12mm
0.8 1 1.2 1.4 1.6 1.8 2
3 3.5 4 4.5 5 5.5 6 6.5 7 10 12 14 16 18 20
maximum amplitude Em (p.u.) half-maximum width σ (mm)
distance (mm)
Emσ
図 3.21: 一辺4mmの立方体により生じるEOD変調の最大振幅と半値幅の側方距
離に対する変化。
0.8 1 1.2 1.4 1.6 1.8 2
2 3 4 5 6 7
10 12 14 16 18 20
maximum amplitude Em (p.u.) hal f- maximum w idt h σ (mm)
size (mm)
Emσ
図 3.22: 側方距離4mmにある立方体により生じるEOD変調の最大振幅と半値幅
の大きさに対する変化。
まとめ
我々のシミュレーションの結果を解析することにより、物体の特徴はEOD変調 の最大振幅と半値幅で表されることが分かった。この結果は、これらのEODのシ ミュレーション結果はBastian [3], Von der Emdeら[47]の測定と合致するものであ る。特に、EOD変調がガウス分布関数で表されるとした場合、半値幅はVon der Emdeらが提唱した最大勾配という指標と、等価なものである。
しかし、一般的に物体によって生じる電場の変調は極めて小さく実験的に正確 に測定することは難しい。本章で述べたような数理モデルによるアプローチは、魚 の周囲に形成される電場の様々な知見を与えてくれる。また、本章で示したよう な物体の距離や大きさを反映するEODの振幅変調の特徴は、電気定位の神経メカ ニズムを解明する上で重要な手がかりを与えるものと考える。
本章で述べた魚が受けるEOD変調は元のEODの振幅に対し数%程度であり、そ の微小な変調をどのように抽出するかは問題である。さらに、物体により生じる EOD変調の特徴である最大振幅と半値幅をどのように抽出するか問題である。以 降の章でこれらのについて言及していく。
小差異抽出におけるシナプス 時定数の効果
4.1 始めに
動物は常にノイズを伴った刺激を受けている。その中から必要な情報を抽出し ている。弱電気魚は自ら発したEODの数%の変調から周囲の状況を認識すること が出来る。弱電気魚の電気受容器は、通常自らの生成した電気刺激に対し不規則に 発火している。この不規則さは刺激のノイズや受容器の感度に由来している。本 章では、移動物体により引き起こされた小さな電気刺激の変調により誘発された 電気受容器の微少な活性の上昇を電気感覚システムによりどのようにしてを抽出 するのか議論する。
4.2 これまでの知見と問題
物体が魚の近傍にある場合などで生じるEOD AMは、自ら発するEODにくらべ わずか数%と非常に小さい。その電気刺激を処理する電気受容器は体表面上に分布 する。その電気受容器はEODの振幅情報を処理するP-typeと位相情報を処理する
T-typeの2種類存在する。本研究で扱う受容器は、振幅情報を処理するP-type受容
器である。その電気受容器は電気刺激の強度に対し確率的な応答を示す。Ratram
とNelsonは、電気受容器の応答の実験データの解析により、電気受容器は刺激の
過去の履歴を用いることでEODの微少な変調をコードすることを示した。電気受 容器の応答はマルコフ過程であることを、RatnamとNelsonは示している[44]。さ らに、BrandmanとNelsonにより電気受容器の活性に依存した閾値を取り入れた 電気受容器モデルが提案された。そのモデルは、マルコフ過程的な応答を示し、電 気受容器の生理実験結果と合致することが示されている[8]。物体により誘発され
た活性の向上は、電気受容器の発するスパイクのペアで表現されるとする研究も 報告されている[10]。電気受容器とELLの錐体細胞とを繋ぐシナプスは興奮性と 抑制性とで時定数が異なっている[7]。また、ELLの錐体細胞が電気受容器層に対 し中心興奮周辺抑制的な受容野を有する[4]。
しかし、確率的に発火する電気受容器の繁雑なスパイク列から物体により生じ たEOD変調を抽出するネットワークメカニズムはよくわかっていない。本章では ネットワークメカニズムという観点から議論する。
4.3 モデル
本章において、弱電気魚が受けるEOD変調は、頭部から尾部に移動する物体に より生じるとする。このときの物体の移動速度は5cm/secとした。弱電気魚は頭部 から尾部へ移動する物体により生じたEOD変調を処理すると想定する。このとき の、EODの振幅変調は第3章の図3.15のように変化する。
本章では図4.1のような電気感覚システムのモデルを構築した。本モデルネット ワークは、電気受容器層とELLからなる。電気受容器、錐体細胞は2次元格子状 に並んでおり、体表面上の位置を保存しながら各層で電気刺激が処理される。電 気受容器層は83×33個の受容器で構成され、i番目の受容器とi+1番目の受容器 が表す体表面上での位置の差は0.5mmである。ELLは35×10個の錐体細胞で構 成され、i番目の錐体細胞とi+1番目の錐体細胞が表す体表面上での位置の差は 1mmである。ELLは電気受容器層に対し中心興奮周辺抑制的な結合を有する。
4.3.1 電気刺激のモデル
受容する電気刺激は、魚自身が発するサイン波型の交流の振幅VEOD0、前章で示 したような物体による電場の歪みVEODAM(t)、そしてホワイトノイズVNoise(t)で表 す。よって、電気受容器が受ける電気刺激VEOD(t)は、
VEOD(t)= (VEOD0 +VEODAM(t)) sin(2πf t)+VNoise(t) (4.1)