ヒト神経芽腫細胞株における
新規ポリエチレングリコール誘導体 PEG-B による抗腫瘍効果の検討
日本大学大学院医学研究科博士課程 生理系機能生理学専攻
長崎瑛里
修了年 2019 年
指導教員 越永從道
目次
概要・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・1 略語一覧・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・9 緒言・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・11 対象と方法・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・15 結果・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・24 考察・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・30 まとめ・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・35 謝辞・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・36 利益相反の開示・・・・・・・・・・・・・・・・・・・・・・・・・・・・37 図・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・38 図説・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・56 引用文献・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・61 研究業績・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・65
1
【概要】
はじめに
化学療法は手術療法、放射線療法とともにがんの三大療法の 1 つであり、特 に小児においては、成人と比較して全身転移をきたして発見されるものが多い ことや、化学療法に感受性のある腫瘍が多いことから、がん治療の中心的役割を 担っているといえる。抗がん剤の最大の問題点は、腫瘍細胞だけでなく正常細胞 も障害するため、生体内で様々な副作用を生じることであるが、小児がんの治療 にあたっては、発育途上であることや治療後に長い余命が期待できることから、
極力副作用の少ない抗がん剤の使用が切望される。
センカファーマシー社の渡辺らは新規がん治療薬の開発を目的とした化合物 のスクリーニングを行い、子嚢菌類由来のポリエチレングリコール化合物およ
びその誘導体(Polyethylene glycol derivatives: PEG-B)が、培養系において各 種ヒト腫瘍細胞に対し強い細胞障害性を示す一方、正常細胞の生存率にはほと
んど影響を与えないことを確認した(特許第5364532号)。このことから、PEG- B は副作用の少ない新規の抗腫瘍薬として期待できると考えられるが、その詳 細な作用機序は未だ明らかでない。本研究では、特に小児がんで発症頻度が高い 神経芽腫における PEG-Bの抗腫瘍効果について検討し、PEG-B の細胞障害作
2
用の機序を解明することを目的として実験を行った。
目的
ヒト神経芽腫細胞株を用いて PEG-B の抗腫瘍効果について検討し、PEG-B の細胞障害作用の機序を解明する。
対象と方法
ヒト神経芽腫細胞株のNB9(MYCNコピー数増幅株)、SK-N-AS(MYCNコ ピー数非増幅株)、ヒト正常線維芽細胞HDFに0~50μMのPEG-Bを添加し、
72時間後の細胞生存率をWST (Water soluble Tetrazolium salts)-8 assayで解 析した。また、細胞増殖能については、NB9およびSK-N-ASに1μMのPEG- Bまたは溶媒のみを添加し、24時間毎1週間WST-8 assayを行い解析した。
PEG-B添加後の細胞周期の解析は、NB9およびSK-N-ASに0、0.1、1μM
の PEG-Bを添加後 24 または72 時間目に細胞を回収し、フローサイトメータ
ーを用いて核内Deoxyribonucleic acid (DNA)量を測定した。
グルコース欠乏下における細胞生存比については、NB9 および SK-N-AS に グルコース濃度の異なる培養条件下で 1μM の PEG-B または溶媒のみを添加 し、72時間後にWST-8 assayを行い解析した。グルコース濃度は、グルコース
3
なし、通常濃度の1/5の濃度、通常濃度の3段階に設定し、結果は、PEG-B非 添加・グルコース通常濃度下培養群を 1 とした細胞生存比で解析した。細胞死 様式については、NB9 および SK-N-AS にグルコース濃度の異なる培養条件下 で0.1、1μMのPEG-Bまたは溶媒のみを添加し、72時間後にAnnexin V-FITC apoptosis detection kitを用いて、アポトーシス細胞の比率を解析した。グルコ
ース欠乏下におけるAdenosine triphosphate (ATP)濃度については、NB9およ びSK-N-ASにグルコース濃度の異なる培養条件下で1μMのPEG-Bまたは溶 媒のみを添加し、12時間培養後、Intracellular ATP測定キットを用いて解析し た。
さらに詳細な PEG-B の作用点を絞り込むため、メタボローム解析を行い、
PEG-B 添加前後で変化する代謝産物を検討した。PEG-B によるミトコンドリ
アの呼吸活性の変化を検討するため、Flux Analyzerで酸素消費速度(Oxygen Consumption Rate: OCR)を計測した。PEG-Bによる呼吸鎖複合体Ⅰ~Ⅳの活
性変化について、MitoCheck Complex I , II/III, IV Activity Assay Kitを用いて 解析した。
結果
NB9および SK-N-AS に PEG-B を0~50μMの濃度で添加すると、濃度依
4
存的に細胞生存率が低下した(NB9では、PEG-B 10μM:*P=0.0162、20μM:
**P=4.23E-03、50μM:**P=3.72E-03、SK-N-ASでは、1μM:**P=4.70E-06、 10μM:**P=8.82E-05、20μM:**P=1.74E-05、50μM:**P=1.51E-05)。一 方、HDFでは高濃度のPEG-B添加でも50%以上の細胞が生存していた(PEG- B 0.1μM:*P=0.0183、1μM:**P=2.31E-04、10μM:**P=3.20E-03、20μ M:**P=2.88E-03、50μM:**P=3.57E-04)。細胞増殖能を解析すると、NB9
においてはPEG-B添加後3日目以降に増殖の抑制がみられるようになり、4日 目より非添加群との差が有意となった(PEG-B添加後4日目:**P=8.67E-03、 5日目:**P=1.86E-04、6日目:**P=3.04E-05、7日目:**P=3.22E-07)。SK- N-ASにおいては、PEG-B添加後に細胞増殖は全くみられず、添加後1 日目よ
り非添加群との間に有意な差を認めた(PEG-B添加後1日目:*P=0.0198、2日 目:**P=6.29E-07、3日目:**P=4.60E-06、4日目:**P=2.16E-04、5日目:
**P=5.83E-04、6 日目:**P=4.69E-04、7 日目:**P=3.86E-05)。細胞周期を
解析すると、PEG-B添加後24時間の時点では、両者ともにG0-G1期やG2/M 期の細胞の比率が増大したが、SubG1期の増加は認められなかった。一方、PEG- B添加72時間後においては、両者ともに SubG1 期の細胞の比率の増加、すな わち死細胞の増加が認められた。
グルコース欠乏実験では、NB9 、SK-N-ASともに、グルコース不含培地にお
5
いて特に、PEG-B添加による細胞生存比の顕著な低下を認めた(NB9では、グ ルコース濃度0g/L:**P=2.28E-07、0.4g/L:**P=4.52E-05、2g/L:**P=1.50E- 03、SK-N-ASでは、グルコース濃度0g/L:**P=9.19E-04、0.9g/L:**P=6.49E- 05、4.5g/L:**P=6.58E-06)。細胞死様式を検討すると、NB9 ではグルコース
不含培地においてのみ、PEG-B添加によりアポトーシス細胞の比率が顕著に増 加し、非添加群と比べて有意に高い値を示した(**P=2.04E-03)。一方、SK-N- ASではいずれの培地においてもPEG-B 添加群は非添加群と比べてアポトーシ ス細胞の有意な増加を示したが、特に低グルコース培地、グルコース不含培地に
おいてその差が顕著であった(グルコース濃度0g/L:**P=5.44E-03、0.9g/L:
**P=1.68E-03、4.5g/L:*P=0.0268)。さらに、細胞内ATP濃度を測定すると、
NB9ではグルコース不含培地においてのみ、PEG-B添加により細胞内ATP濃
度が有意に低下した(*P=0.0145)。一方、SK-N-ASではいずれの培地において
もPEG-B添加群は非添加群と比べて細胞内 ATP量の有意な低下を示したが、
特にグルコース不含培地においてその差が顕著であった(グルコース濃度0g/L:
*P=0.0217、0.9g/L:*P=0.0243、4.5g/L:**P=1.78E-03)。
PEG-B 添加後の細胞内代謝物をメタボローム解析により網羅的に調べると、
NB9においては、PEG-B添加群では非添加群と比較し、カルニチンの有意な低 下(*P=0.002)、 ア セ チ ル カ ル ニ チ ン の 増 加 傾 向 、Nicotinamide adenine
6
dinucleotide hydrate (NADH)の有意な増加(**P=0.001)が認められた。解糖
系およびTricarboxylic acid (TCA)回路については、一定の傾向は認められなか
った。SK-N-ASにおいては、PEG-B添加群では非添加群と比較し、解糖系の亢
進傾向、TCA 回路の抑制傾向、カルニチンの有意な低下(*P=0.029)、アセチル カルニチンの有意な増加(*P=0.044)、NADHの増加傾向が認められた。
ミトコンドリア呼吸活性の解析では、NB9、SK-N-AS いずれにおいても、
PEG-B添加後に顕著なOCRの低下が認められたが、このPEG-BによるOCR
の抑制はCarbonyl cyanide 4-(trifluoromethoxy)phenylhydrazone (FCCP)によ っても解除されなかった。さらに、PEG-B添加による呼吸鎖複合体Ⅰ~Ⅳの活 性変化の解析において、PEG-B は呼吸鎖複合体Ⅰに対してのみ、1μM 以上の 濃度での添加で有意に活性を抑制した(PEG-B 1μM:**P=9.17E-03、2μM:
**P=6.41E-03)。
考察
PEG-B は MYCN コピー数増幅の有無に関わらず、神経芽腫細胞に対し増殖
抑制効果を示した。正常細胞に対する効果は限定的であったことから、神経芽腫 に対する副作用の少ない治療薬として大いに期待できると考えられた。細胞周 期を解析すると、PEG-B による細胞死の誘導は添加後 24 時間目には認められ
7
ず、添加後 72 時間目で観察された。PEG-B を添加してすぐではなく時間が経 ってから死細胞が増加したことから、時間とともに培地の栄養が枯渇したため に細胞死が誘導されている可能性が示唆された。そのため、グルコース欠乏実験
を行ったところ、グルコース欠乏下において特に,PEG-B添加による顕著な細 胞生存率の低下とアポトーシス細胞の比率の増加を認め、さらに細胞内ATP濃 度の顕著な低下を認めた。以上の結果から、PEG-Bは解糖系以外のATP産生経 路を阻害し、細胞の増殖を抑制している可能性が示唆された。
メタボローム解析の結果、NB9、SK-N-ASともに、PEG-B添加により、カル ニチンの低下、アセチルカルニチンの増加、NADHの増加が認められたことと、
SK-N-AS において、解糖系の亢進傾向と TCA 回路の抑制傾向が認められたこ
とから、PEG-Bはミトコンドリア呼吸鎖複合体を阻害している可能性が考えら
れた。さらにミトコンドリア呼吸活性の解析結果から、PEG-Bの作用点の候補 として呼吸鎖複合体Ⅰ~Ⅳに絞り込むことができた。この結果を踏まえて、
PEG-B添加後の複合体Ⅰ~Ⅳの活性変化を検討したところ、複合体Ⅱ~Ⅳでは
PEG-Bの添加前後での活性変化は認めず、複合体Ⅰのみで活性低下を認めたた
め、PEG-Bの標的は複合体Ⅰであることが証明された。PEG-Bが腫瘍細胞に特
異的に効果を発揮する理由としては、腫瘍細胞はATPを大量生産・消費して増 殖するが、PEG-B により酸化的リン酸化による効率的な ATP 産生を阻害され
8
るからであると考えられた。
近年、がん細胞はその増殖・生存を維持する上で糖質への依存度が高いことに
着目し、糖質を制限したケトン食が欧米で注目されている。本研究では、PEG- B はグルコース欠乏下において顕著に効果を発揮したことから、ケトン食との 併用により相乗効果が期待できる。今後、動物実験において、通常食およびケト
ン食飼育下でPEG-Bを投与し、生体内におけるPEG-Bの抗腫瘍効果を検討す ることが必要であると考える。また、細胞間におけるPEG-Bの効果の差につい ての検討や、PEG-Bの複合体Ⅰに対する詳細な作用機序の検討については、今 後の課題となる。
結語
PEG-Bはミトコンドリア呼吸鎖複合体Ⅰを阻害することで、酸化的リン酸化
による効率的なATP産生を阻害する。このため、ATPを大量生産・消費して増 殖する腫瘍細胞に特異的に効果を表し、特にグルコース欠乏下においてPEG-B の効果が顕著になると考えられた。
9
【略語一覧】
2-OG: 2-Oxoglutaric acid 3-PG: 3-Phosphoglyceric acid ADP: Adenosine diphosphate ALCAR: O-Acetylcarnitine
AMPK: AMP-activated protein kinase ATP: Adenosine triphosphate
CE: Capillary electrophoresis DNA: Deoxyribonucleic acid F1, 6P: Fructose 1, 6-diphosphate F6P: Fructose 6-phosphate
FCCP: Carbonyl cyanide 4-(trifluoromethoxy)phenylhydrazone G6P: Glucose 6-phosphate
GAP: Glyceraldehyde 3-phosphate MT: Migration time
N.A.: Not Available.
NAD+: Nicotinamide adenine dinucleotide
NADH: Nicotinamide adenine dinucleotide hydrate
10
OCR: Oxygen Consumption Rate
PEG-B, PEG-X: Polyethylene glycol derivatives
本論文では、センカファーマシー社が発見、抽出、有機合成した化合物を
PEG-Xと総称し、そのうち本実験で使用した化合物をPEG-Bで表す。
PEP: Phosphoenolpyruvic acid PI: Propidium iodide
TCA: Tricarboxylic acid
TOFMS: Time-of-flight mass spectrometry WST: Water soluble Tetrazolium salts
11
【緒言】
がんの化学療法とその問題点
わが国における全部位のがん死亡数(2016年)は37.3万人であり(1)、総死亡 数の約 29%を占める。また、全部位のがん罹患数(2013 年)は約 85 万人と報告 されており、いまや2人に1人は一生のうちに何らかのがんに罹るとされる(1)。 小児がんは、年間約2100人と成人に比べて発生数は少ないが(2)、小児期の病 死原因としては最も頻度が高い(3)。化学療法は手術療法、放射線療法とともに がんの三大療法の 1 つであるが、特に小児においては、成人と比較して細胞増 殖速度が速いため、全身転移をきたして発見されるものが多いことや、化学療法 に感受性のある腫瘍が多いことから、全身療法である化学療法は治療の中心的
役割を担っているといえる。薬剤の種類は、Deoxyribonucleic acid(DNA)鎖 内あるいは鎖間に架橋を形成して複製を阻害するアルキル化剤(4)や白金製剤
(5)、DNA合成に必要な葉酸の産生を阻害する代謝拮抗剤(6)、微小管形成を 阻害する植物アルカロイド(7)など様々な作用機序のものが存在する。
抗がん剤の最大の問題点は、腫瘍細胞だけでなく正常細胞も障害するため、生 体内で様々な副作用が生じることである。近年、腫瘍細胞に特異的な細胞特性を 規定する分子に作用する分子標的薬の開発が進められ、その有効性の報告が蓄
12
積されつつあるが、構造が類似した複数の標的にも阻害活性を示すため、やはり 副作用の問題は依然として残る。抗がん剤全般に共通する副作用として、骨髄抑 制による貧血、白血球減少による易感染性、血小板減少や消化器症状、などが挙 げられ、これらは時に患者の生命を脅かすこともあり得る。さらに小児がんの治 療にあたっては、成長・発育や妊孕性温存も含めた治療後晩期障害を考慮する必 要があるが、現時点では長期生存例におけるシスプラチンによる聴力障害や腎 障害、アントラサイクリン系薬剤による心不全、アルキル化剤による不妊症や二
次がんの発症などの問題がある(8)。このため、長期予後をも考慮した極力副作 用の少ない治療法の開発が喫緊の課題であると考える。
神経芽腫
神経芽腫は胎生期の神経堤細胞に由来する未分化な胎児性腫瘍であり(9)、副 腎や後腹膜に多く発生する(10)。脳腫瘍を除く小児悪性固形腫瘍で最も高頻度 に認められ(9)、本邦での新規発症数は年間150~200人程度である。治療は化 学療法、外科療法、放射線療法を組み合わせて行う。化学療法は、主にビンクリ スチン、シクロフォスファミド、シスプラチン、エトポシド、ドキソルビシンな どからなる多剤併用療法が行われる。予後は診断時年齢、臨床病期、生物学的因 子と強く関連し、中でもMYCN遺伝子は神経芽腫の発生、予後に関与する最も
13
重要な遺伝子であり、強力な予後不良因子である(11)。MYCN非増幅例の神経 芽腫患者の5年無再発生存率が76.4%、5年全生存率が86.1%であるのに対し、
MYCN増幅例の神経芽腫の5年無再発生存率は48.1%、5年全生存率は55.2% と有意に低く(12)、この予後不良群の治療の克服が課題となる。
PEG-B
これまでに、微生物を利用した医薬品製造は社会に大きな恩恵をもたらして きた。世界初の抗生物質であるペニシリンがアオカビから発見されたことは有
名である(13)。がんの分野においては、過去 70 年に全世界で承認された微生 物由来の小分子抗腫瘍薬は 189 種類にものぼる(14-16)。たとえば、小児にお いて腎芽腫や横紋筋肉腫の治療で頻用され治療成績の向上に寄与しているアク
チノマイシンDは、放線菌から発見された(17)。このように、微生物由来の化 合物はがん治療における創薬開発のリソースとして非常に有用であると考えら れる。
センカファーマシー社の渡辺らは、土壌から新種の微生物を単離し、その菌が ゾウリムシやミドリムシなどの他の微生物に対し毒性を持つことを確認した。
この微生物は真核生物であり、分類上コウジカビと同じ子嚢菌類に属すること が判明した。有効成分の抽出を試みたところ、菌の破砕物の非水溶性の画分に毒
14
性のある分子が含まれており、この分子は微生物だけでなく膵癌や肺癌などの ヒト腫瘍細胞に対しても強い細胞障害性を示した。有効成分を単離し構造決定 を行うと、有効成分はポリエチレングリコールの誘導体であり新規の化合物で あった。さらにこの化合物の構造類似体を有機合成し、いずれも腫瘍に対し細胞
障害性を示すことを確認した(特許第5364532号)。本研究ではこれらの化合物 を Polyethylene glycol derivatives: PEG-X と総称し、代表的な化合物として PEG-Bを図1 に示す(図1)。さらに興味深いことに、PEG-Bは正常細胞には
影響を与えず、腫瘍細胞に対してのみ障害性を示すことが確認された(図 2)。 このことから、PEG-Bは副作用の少ない新規の抗腫瘍薬として期待できると考 えられる。
しかしながらその作用機序は不明であり、また腫瘍特異的に効果を発揮する 理由もいまだ明らかではない。本研究では、特に小児がんで発症頻度が高い神経 芽腫における PEG-Bの抗腫瘍効果について検討し、PEG-B の細胞障害作用の 機序を解明することを目的として実験を行った。
15
【対象と方法】
1. 各種細胞株の培養
細胞株はヒト神経芽腫細胞株 NB9 および SK-N-AS、およびヒト正常線維芽 細胞HDFを使用した。NB9はRiken cell bank (Tsukuba, Japan)より購入し た。SK-N-AS および HDF は American Type Culture Collection (ATCC, Virginia)から購入した。NB9は15%ウシ胎児血清(Nichirei Bioscience, Tokyo, Japan)とPenicillin-Streptomycin-Glutamine (Nacalai Tasque, Kyoto, Japan)
含有のRPMI1640培地(Nacalai Tasque)で培養した。SK-N-ASおよびHDF は10%ウシ胎児血清およびPenicillin-Streptomycin-Glutamine含有のDMEM 培地(Nacalai Tasque)を使用した。すべての細胞株は37℃、5% CO2の培養 条件下で培養した。
上記細胞株のうち、NB9はMYCNコピー数増幅株であり、SK-N-ASはMYCN コピー数非増幅株である。
2. 細胞生存率および細胞増殖能の検討:WST (Water soluble Tetrazolium salts )-8 assay
NB9と HDFは 1.0×103個/100μl/wellの密度で、SK-N-AS は 2.0×103個
16
/100μl/wellの密度で96 well plateに播種し、24時間培養後に0~50μMの濃 度のPEG-Bを添加した培地に置換した。72時間後に発色試薬であるCell Count Reagent SF (Nacalai Tesque)を10μl添加し、1時間の呈色反応後、Wallac 1420 ARVO MX (Perkin Elmer, Waltham, MA)を用いて吸光度を測定した(WST-8 assay)。細胞増殖能については、NB9およびSK-N-ASを上記同様の密度で96 well plateに播種し、24 時間培養後に 1μM のPEG-B または溶媒のみを添加
した培地に置換した。その後24時間毎に1週間WST-8 assay を行った。測定 は各条件につき4 wellずつ行った。
3. 細胞周期の解析:FACS analysis
NB9は1.0×105個/dishの密度で、SK-N-ASは2.0×105個/dishの密度で10 cm dishに播種し、24時間培養後に0.1、1μMのPEG-Bまたは溶媒のみを添
加した培地に置換した。24、72 時間後に浮遊細胞および接着細胞を回収した。
細胞をPBS洗浄後、冷却した75%エタノールで一晩以上固定した。細胞をPBS 洗浄後、0.1% FBS、25μg/ml propidium iodide (PI)、200μg/ml RNase Aを添 加した。15 分間暗室で室温放置した後、フローサイトメーターFACSCallibur
(BD Biosciences, San Jose, CA)を用いて核内DNA量を測定した。PIで染色 されるDNAの含有量に応じて、DNAヒストグラムからSub-G1期、G0-G1期、
17
S期、G2/M期を同定し、それぞれの比率を算出した。なおSub-G1期の細胞は 死細胞に相当する。
6. グルコース欠乏下における細胞生存比の検討:WST-8 assay
NB9 は 2.0×103 個/100μl/well の密度で、SK-N-AS は 5.0×103個/100μ l/wellの密度で、それぞれ96 well plateに播種した。24時間培養後にグルコー
ス濃度の異なる培地に置換し、1μMのPEG-Bまたは溶媒のみを添加した。グ ルコース濃度はそれぞれ、NB9では0、0.9、4.5 g/l(通常濃度)、SK-N-ASで は0、0.4、2g/l(通常濃度)とした。72時間後にWST-8 assayを行った。測定 は各条件につき4 wellずつ行った。結果は、PEG-B非添加・グルコース通常濃 度下培養群(NB9は2g/L、SK-N-ASは4.5g/L)を1とした細胞生存比で解析 した。
7. グルコース欠乏下における細胞死様式の検討:FACS analysis
NB9は1.0×105個/dishの密度で、SK-N-ASは2.0×103個/100μl/wellの密
度で10 cm dishに播種した。24時間培養後にグルコース濃度の異なる培地に置 換し、0.1、1μM の PEG-Bまたは溶媒のみを添加した。72 時間後、Annexin V-FITC apoptosis detection kit (BioVision, Inc., Milpitas, CA)を用いて、推奨
18
プロトコールに従い、アポトーシス細胞の比率を解析した。Annexin VとPIの 蛍光強度はフローサイトメーターで測定した。結果に描出された四分画のうち、
Annexin V 陽性細胞である右上と右下の分画をアポトーシス細胞として計測し
た。計測は各条件につき3回ずつ行った。
8. グルコース欠乏下におけるATP濃度の検討
NB9 および SK-N-AS を 5.0×103個/100μl/well の密度でそれぞれ 96 well
plate に播種した。24 時間培養後に上記同様グルコース濃度の異なる培地に置
換し、1μMのPEG-Bまたは溶媒のみを添加した。12時間培養後、Intracellular ATP測定キット(Toyo-b-net, Tokyo, Japan)を用いて推奨プロトコールに従い、
細胞内 ATP 濃度を解析した。抽出した ATP はホタル・ルシフェラーゼ発光法 の原理を利用してルミノメーターWallac 1420 ARVO MXで測定した。測定は 各条件につき4 wellずつ行った。
9. メタボローム解析
NB9およびSK-N-ASを2.0×103個/dishの密度で10 cm dishに播種した。
24時間後に1μMのPEG-Bまたは溶媒のみを含む培地に置換し、8時間後、細 胞内代謝物すべてをメタノールで抽出した。回収した抽出液を遠心分離し、上澄
19
みを限外濾過ユニット(Human Metabolome Technologies: HMT, Tsuruoka,
Japan)に移し、再度遠心分離を行うことでタンパク質を除去した。サンプルは
HMT社に委託し、下記の手法にてメタボローム解析を行った。キャピラリー電 気泳動(capillary electrophoresis: CE)-飛行時間型質量分析計(time-of-flight mass spectrometry: TOFMS)(CE-TOFMS)を用いてサンプル中の代謝物質を
解析した。測定は各条件につき3回ずつ行った。CE-TOFMSで検出されたピー クは、自動積分ソフトウェアのMasterHands ver.2.17.1.11(慶應義塾大学開発)
を用いて、シグナル/ノイズ(S/N)比が3以上のピークを自動抽出し、質量電荷比 (m/z)、ピーク面積値、泳動時間(Migration time: MT)を得た。得られたピーク面
積値は以下の式を用いて相対面積値に変換した:相対面積値=(目的ピークの面 積値)/(内部標準物質の面積値×試料量)。検出されたピークに対して m/zとMT の値をもとに、HMT代謝物質ライブラリ及びKnown-Unknownライブラリに 登録された全物質との照合・検索を行った。検索のための許容誤差は MT で±
0.5 min、m/zでは±10 ppmとした。
10.ミトコンドリア呼吸活性の検討:Flux Analyzer
Seahorse Bioscience XFp extracellular flux analyzer (Primetech corporation, Tokyo, Japan)を使用して細胞外培養液中の細胞による酸素消費速
20
度(Oxygen Consumption Rate:OCR)を計測し、ミトコンドリア呼吸の活性 を検討した。
NB9およびSK-N-ASを5.0×103個/100μl/wellの密度で FluxPak XFp 細
胞培養プレートに播種し、48時間培養した。FluxPak XFp センサーカートリッ ジはキャリブラントに浸し、CO2制御のない 37℃のインキュベーター内で 48 時間水和した。細胞の培地を解析培地XF Base Medium (11.1mM glucose, 1mM pyruvate, 2mM glutamine, pH 7.4)に置換し、CO2制御のない37℃のインキュ
ベーター内で 1 時間培養した。最終濃度が 1μM になるよう調整した PEG-B と 、 適 当 量 の 解 析 培 地 で 希 釈 し た ミ ト ス ト レ ス テ ス ト 試 薬 (Oligomycin, carbonyl cyanide 4-(trifluoromethoxy)phenylhydrazone (FCCP), Rotenone) をインジェクションポートにそれぞれ充填した。センサーカートリッジおよび
細胞培養プレートをXFp 本体へ設置し、解析を行った。測定は各条件につき3 回行った。なお、OligomycinはATP合成酵素(複合体Ⅴ)の阻害剤であり(18)、 Rotenoneは呼吸鎖複合体Ⅰの阻害剤である(19)。またFCCPは脱共役剤であ
り(20)、ミトコンドリア内膜にイオノフォアを形成することで膜電位の消失を 促すが、それに伴い、電子伝達系の回転速度が最大となる。
11. 呼吸鎖複合体Ⅰ活性の検討
21
ミトコンドリアにおける酸化的リン酸化反応系は、複合体Ⅰ(Nicotinamide adenine dinucleotide hydrate (NADH)脱水素酵素)、複合体Ⅱ(コハク酸脱水
素酵素)、複合体Ⅲ(シトクロムc酸化還元酵素)、複合体Ⅳ(シトクロムc酸化 酵素)、および複合体Ⅴ(ATP合成酵素)の5つの酵素複合体によって構成され ている(21)。MitoCheck Complex I Activity Assay Kit (700930, Cayman Chemical, Ann Arbor, MI)を用いて、推奨プロトコールに従い、PEG-B添加後 の呼吸鎖複合体Ⅰの活性を測定した。ポジティブコントロールとして、既知の呼
吸鎖複合体Ⅰ阻害剤である Rotenone 使用した。PEG-B を最終濃度が 0.2、1、 2μMになるように、またはRotenoneを最終濃度が2μMになるように、牛心
筋由来ミトコンドリアを含む試薬に添加した。NADH による 340nm の吸光度 を1分毎に4分間測定し、NADHの酸化率を算出することで呼吸鎖複合体Ⅰの 活性変化を解析した。測定は各条件につき3回行った。
12. 呼吸鎖複合体Ⅱ/Ⅲ活性の検討
MitoCheck Complex II/III Activity Assay Kit (700950, Cayman Chemical) を用いて、推奨プロトコールに従い、PEG-B添加後の呼吸鎖複合体Ⅱ/Ⅲの活性 を測定した。ポジティブコントロールとして、既知の呼吸鎖複合体Ⅲ阻害剤であ るAntimycinを使用した。PEG-Bを最終濃度が0.2、1、2μMになるように、
22
またはAntimycinを最終濃度が5μMになるように、牛心筋由来ミトコンドリ アを含む試薬に添加した。呼吸鎖複合体Ⅲは呼吸鎖複合体Ⅱより産生されたユ
ビキノールから電子を受け取り、シトクロムcを還元する。生成した還元型シト クロムcによる 550 nmの吸光度を30 秒ごとに15 分間測定し、シトクロムc の還元率を算出することで呼吸鎖複合体Ⅱ/Ⅲの活性変化を解析した。測定は各 条件につき3回行った。
13. 呼吸鎖複合体Ⅳ活性の検討
MitoCheck Complex IV Activity Assay Kit (700990, Cayman Chemical)を用
いて、推奨プロトコールに従い、PEG-B添加後の呼吸鎖複合体Ⅳの活性を測定 した。ポジティブコントロールとして、既知の呼吸鎖複合体Ⅳ阻害剤である
KCN を使用した。PEG-B を最終濃度が 0.2、1、2μM になるように、または KCNを最終濃度が100μMになるように、牛心筋由来ミトコンドリアを含む試
薬に添加した。還元型シトクロムcは呼吸鎖複合体Ⅳにより酸化される。還元型 シトクロムc による 550nm の吸光度を 30 秒ごとに15 分間測定し、還元型シ トクロム c の酸化率を算出することで呼吸鎖複合体Ⅳの活性変化を解析した。
測定は各条件につき3回行った。
23
13. 統計学的分析
細胞周期の解析を除いた全ての解析はそれぞれn≧3を用いて実験を行い、そ れらの平均値を解析した。解析における平均の差を Student’s t 検定で行い、
P<0.05を「統計学的有意差あり」とした。定量結果は平均値±標準偏差にて表
記した。
24
【結果】
1. 各種細胞株における細胞生存率および細胞増殖能の検討
NB9、SK-N-AS、HDFに 0~50μM の濃度のPEG-B を添加し、72 時間後
の細胞生存率をWST-8 assayで解析した。結果、NB9およびSK-N-ASにおい
て、PEG-Bは濃度依存的に細胞生存率を低下させた(図 3 左上のNB9 では、
PEG-B 10μM:*P=0.0162、20μM:**P=4.23E-03、50μM:**P=3.72E-03、 図 3 右上の SK-N-ASでは、1μM:**P=4.70E-06、10μM:**P=8.82E-05、 20μM:**P=1.74E-05、50μM:**P=1.51E-05)。一方、HDFでは高濃度のPEG- B 添加でも 50%以上の細胞が生存していた(図 3 左下、PEG-B 0.1μM:
*P=0.0183、1μM:**P=2.31E-04、10μM:**P=3.20E-03、20μM:**P=2.88E- 03、50μM:**P=3.57E-04)。また、NB9およびSK-N-ASにおいて、PEG-B 1μM 添加後の細胞増殖能を解析すると、NB9 においては PEG-B 添加後 3 日
目以降に増殖の抑制がみられるようになり、4日目より非添加群との差が有意と なった(図4 左、PEG-B添加後 4 日目:**P=8.67E-03、5 日目:**P=1.86E- 04、6 日目:**P=3.04E-05、7 日目:**P=3.22E-07)。SK-N-AS においては、
PEG-B添加後に細胞増殖は全くみられず、添加後1日目より非添加群との間に
有意な差を認めた(図4右、PEG-B添加後1日目:*P=0.0198、2日目:**P=6.29E-
25
07、3 日目:**P=4.60E-06、4 日目:**P=2.16E-04、5 日目:**P=5.83E-04、 6日目:**P=4.69E-04、7日目:**P=3.86E-05)。
2. 細胞周期の解析
PEG-B添加による細胞生存率の低下が、細胞周期の停止によるものか、それ
とも細胞死の誘導によるものかを検討するために、フローサイトメーターを用
いて核内DNA量を解析した。結果、PEG-B添加24時間後においては、NB9、 SK-N-ASともに、G0-G1期やG2/M期の細胞の比率が増大したが、Sub-G1期
の増加は認められなかった(図5上段)。一方、PEG-B添加72時間後において は、NB9、SK-N-AS ともに、Sub-G1 期の細胞の比率の増加、すなわち、死細 胞の増加が認められた(図5下段)。
3.グルコース欠乏下における細胞生存比の検討
細胞周期の解析で、PEG-B による細胞死の誘導は添加後 24 時間目には認め られず、添加後 72 時間目で観察された。PEG-B を添加してすぐではなく時間 が経ってから死細胞が増加したことから、時間とともに培地の栄養が枯渇した ために細胞死が誘導されている可能性が示唆された。そこで、グルコース欠乏実 験を行ったところ、NB9 、SK-N-AS ともに、グルコース不含培地において特
26
に、PEG-B添加による細胞生存比の顕著な低下を認めた(図6上のNB9では、
グルコース濃度0g/L:**P=2.28E-07、0.4g/L:**P=4.52E-05、2g/L:**P=1.50E- 03、図6下のSK-N-ASでは、グルコース濃度0g/L:**P=9.19E-04、0.9g/L:
**P=6.49E-05、4.5g/L:**P=6.58E-06)。
4.グルコース欠乏下における細胞死様式の検討
結果 3 を受けて、グルコース欠乏下における PEG-B 添加後の細胞死様式を Annexin V/PI二重染色法で検討した。結果、NB9ではグルコース不含培地にお
いてのみ、PEG-B添加によりアポトーシス細胞の比率が顕著に増加し、非添加 群と比べて有意に高い値を示した(図7-1、**P=2.04E-03)。一方、SK-N-ASで はいずれの培地においても PEG-B 添加群は非添加群と比べてアポトーシス細 胞の有意な増加を示したが、特に低グルコース培地、グルコース不含培地におい
てその差が顕著であった(図7-2、グルコース濃度0g/L:**P=5.44E-03、0.9g/L:
**P=1.68E-03、4.5g/L:*P=0.0268)。
5.グルコース欠乏下におけるATP濃度の検討
PEG-Bが細胞のATP産生に与える影響について検討するため、PEG-B添加
後のATP濃度を測定した。結果、NB9ではグルコース不含培地においてのみ、
27
PEG-B添加により細胞内ATP濃度が有意に低下した(図8上、*P=0.0145)。
一方、SK-N-AS ではいずれの培地においてもPEG-B 添加群は非添加群と比べ
て細胞内ATP濃度の有意な低下を示したが、特にグルコース不含培地において その差が顕著であった(図 8 下、グルコース濃度 0g/L:*P=0.0217、0.9g/L:
*P=0.0243、4.5g/L:**P=1.78E-03)。
6.メタボローム解析
これまでの実験結果から、PEG-Bは解糖系以外のATP産生経路を阻害し,細 胞の増殖を抑制している可能性があることが示されたため、PEG-Bの作用点を さらに絞り込むことを目的に、PEG-B添加後の細胞内代謝物をメタボローム解 析により網羅的に調べた。HMT代謝物質ライブラリに登録されている代謝物質 のうち、本実験で検出された物質を代謝経路に描画し、PEG-B添加群と非添加 群で明確な違いのみられた経路を図9に示した。結果、NB9においては、PEG- B添加群では非添加群と比較し、カルニチンの有意な低下(*P=0.002)、アセチル
カルニチンの増加傾向、NADHの有意な増加(**P=0.001)が認められた。解糖 系およびTCA回路については、一定の傾向は認められなかった(図9-1)。SK- N-AS においては、PEG-B 添加群では非添加群と比較し、解糖系の亢進傾向、
TCA 回路の抑制傾向、カルニチンの有意な低下(*P=0.029)、アセチルカルニチ
28
ンの有意な増加(*P=0.004)、NADHの増加傾向が認められた(図9-2)。
7.ミトコンドリア呼吸活性の検討
PEG-B がミ トコ ンドリ ア呼吸 に与 え る影響 を検討 する こ とを目 的に、
Seahorse Bioscience XFp extracellular flux analyzerを使用して、細胞外培養
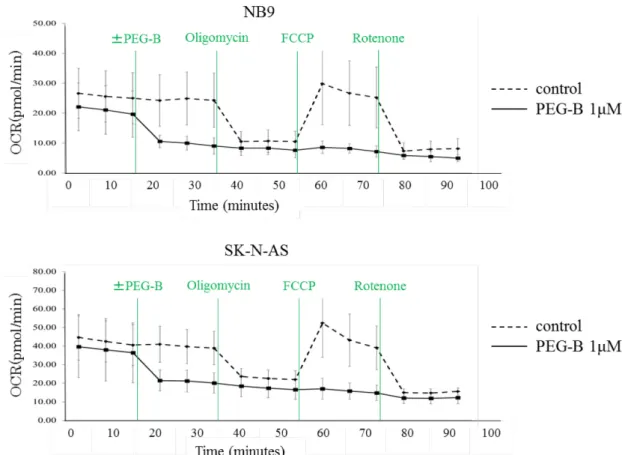
液中の細胞による酸素消費速度(OCR)を計測した。結果、NB9、SK-N-ASい ずれにおいても、PEG-B添加後に顕著なOCRの低下が認められた。このPEG- BによるOCRの抑制はFCCPによっても解除されなかった(図10)。
8.PEG-B添加後の呼吸鎖複合体Ⅰの活性変化
結果7より、PEG-Bは複合体Ⅰ~Ⅳのいずれかの機能を阻害している可能性
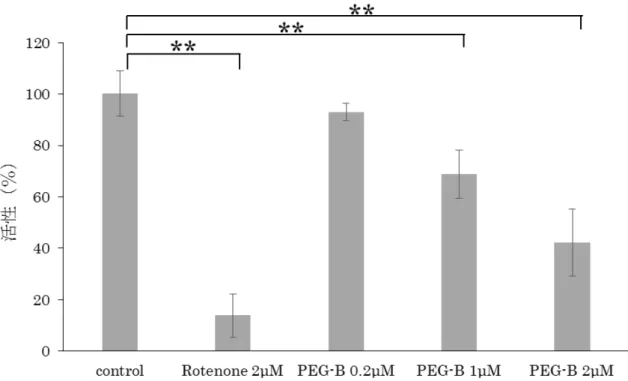
が示唆されたため、MitoCheck Complex I Activity Assay Kitを使用してPEG- B添加後の呼吸鎖複合体Ⅰの活性を測定した。結果、PEG-Bは1μM以上で有 意に呼吸鎖複合体Ⅰの活性を阻害した(図11、Rotenone 2μM:**P=8.76E-05、 PEG-B 1μM:**P=9.17E-03、PEG-B 2μM:**P=6.41E-03)。
9.PEG-B添加後の呼吸鎖複合体Ⅱ/Ⅲの活性変化
MitoCheck Complex II/III Activity Assay Kitを使用してPEG-B添加後の
29
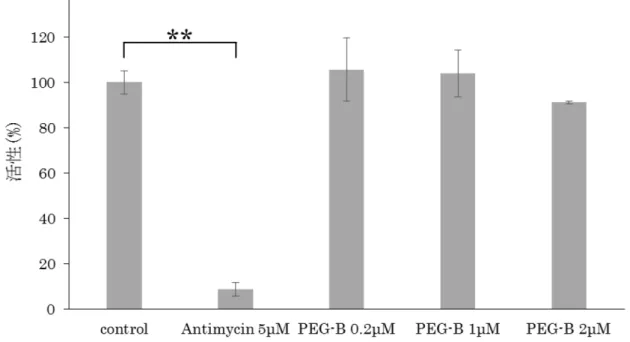
呼吸鎖複合体Ⅱ/Ⅲの活性を測定した。結果、PEG-Bは0.2、1、2μMのいず れの濃度においても、control群と比較して有意な差を認めなかった(図12、 Antimycin 5μM:**P=1.07E-06)。
10.PEG-B添加後の呼吸鎖複合体Ⅳの活性変化
MitoCheck Complex IV Activity Assay Kitを使用してPEG-B添加後の呼吸
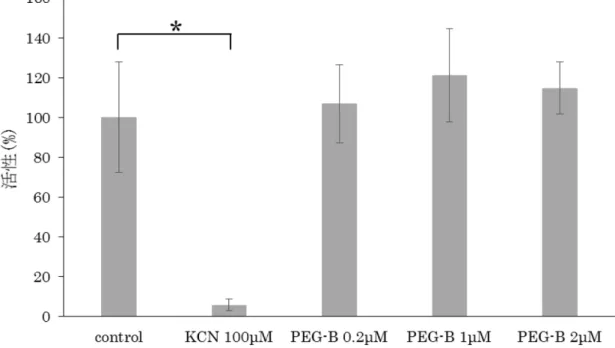
鎖複合体Ⅳの活性を測定した。結果、PEG-Bは0.2、1、2μMのいずれの濃度 においても、control群と比較して有意な差を認めなかった(図13、KCN 100 μM:*P=0.0268)。
30
【考察】
本研究ではPEG-BがMYCNコピー数増幅の有無に関わらず、腫瘍細胞に対 し増殖抑制効果を持つことが確認できた。正常細胞に対する効果は限定的であ
ったことから、PEG-Bは神経芽腫に対する副作用の少ない治療薬として大いに 期待できると考えられた。一方で、2細胞株間でのIC50の大きな違いが見られ たことや、メタボローム解析の代謝傾向の違いから、細胞間でPEG-Bの効果に 差があると考えられる。NB9とSK-N-ASの違いはMYCN増幅コピー数増幅の 有無の他に、NB9はTP53 wild type、SK-N-ASはTP53 null typeという違い がある。TP53は多くの成人癌で遺伝子変異がみられる代表的ながん抑制遺伝子 であり、乳癌や肺癌など複数の癌腫において TP53 の変異有りが予後と負の相 関を示すと報告されている(22、23)。変異型p53蛋白はエナジーセンサーであ るAMP-activated protein kinase (AMPK)経路を阻害することで腫瘍形成を促 進する(24)という報告があることと本研究結果から、TP53と低ATP感受性、
およびPEG-B感受性の関連が示唆される。細胞間におけるPEG-Bの効果の違
いについては、今後細胞株の種類を増やして詳細に検討する必要があると考え る。
PEG-B添加による細胞数の低下が、細胞周期の停止によるものか、それとも
31
細胞死の誘導によるものかを検討するために、フローサイトメーターを用いて
核内DNA量を解析した。結果、PEG-B添加24時間後の時点では、NB9、SK-
N-AS ともに、G0-G1 期や G2/M 期の細胞の比率が増大しており、細胞周期の
抑制が考えられたが、PEG-B添加72時間後になると、NB9、SK-N-ASともに、
Sub-G1期の細胞の比率が増大しており、細胞死が誘導されていると考えられた
(図5)。PEG-Bを添加してすぐではなく時間が経ってから死細胞が増加したこ
とから、時間とともに培地の栄養が枯渇したために細胞死が誘導されている可
能性が示唆された。そのため、次にグルコース欠乏実験を行ったところ、PEG- B 添加群では特にグルコース不含培地において、アポトーシスが誘導されて細
胞生存率が顕著に低下すること、および細胞内ATP濃度が顕著に低下すること がわかった(図6~8)。以上より、PEG-Bは解糖系以外のATP産生経路を阻害 し、細胞の増殖を抑制している可能性が示唆された(図14)。そこでメタボロー ム解析を行い、PEG-B添加前後で変化する代謝産物を調べた。結果、NB9、SK-
N-AS ともに、PEG-B 添加により、カルニチンの低下、アセチルカルニチンの
増加、NADHの増加が認められた。また、SK-N-ASにおいては、PEG-B添加 により、細胞のエネルギー産生において解糖系が優位になりTCA回路が抑制さ れている可能性が示唆された(図 9)。解糖系が亢進するとカルニチンは低下す る。これは解糖系の亢進および TCA 回路の抑制により増加したアセチル CoA
32
を消費するためにカルニチンが使用されるからである。この時にアセチル CoA がカルニチンにアセチル基を渡すため、アセチルカルニチンが産生される(25)。 一方で、TCA回路で産生されるNADH量は増加したことから、PEG-B添加に よりTCA回路自体は阻害されず、TCA回路で生じた NADHからATPを産生 する過程、すなわち、ミトコンドリア呼吸鎖複合体が阻害されている可能性が考
えられた(図15)。
ミトコンドリア呼吸においては、TCA 回路で得られた NADH 由来の電子を ミトコンドリア内膜上の酵素複合体間で伝達し、ATP を大量に産生するが、こ の過程で酸素が消費される。Flux Analyzer は細胞の酸素消費を酸素消費速度
(OCR)として、フォトルミネッセンス法により検出、指標とすることにより、細
胞レベルのエネルギー代謝状態を評価している。今回の実験で、Control群では Oligomycinの添加による OCR の低下、FCCP 添加による OCR の上昇、およ び Rotenone 添加による OCR の低下が確認されたが、PEG-B 添加群では、
OligomycinやRotenone添加時と同程度のOCRの低下を示した(図10)。この
結果から、PEG-Bはミトコンドリア呼吸を阻害していることが確認できた。さ
らに、PEG-BによるOCRの抑制はFCCP添加によっても解除されなかったこ
とから、複合体ⅤはPEG-Bの標的として除外できると考えられた。以上の結果
から、PEG-B は複合体Ⅰ~Ⅳのいずれかを阻害している可能性が示唆された。
33
(図16)。
この結果を踏まえて、PEG-B添加後の複合体Ⅰ~Ⅳの活性変化を検討したと ころ、複合体Ⅱ~ⅣではPEG-Bの添加前後での活性変化は認めず(図12、13)、 複合体Ⅰのみで活性低下を認めたため(図 11)、PEG-B の標的は複合体Ⅰであ ることが証明された。PEG-Bが腫瘍細胞に特異的に効果を発揮する理由として は、腫瘍細胞はATPを大量生産・消費して増殖するが、PEG-Bにより酸化的リ ン酸化による効率的なATP産生を阻害されるからであると考えられた。
これまでに、酸化的リン酸化を治療標的とした研究はいくつか報告されてい
る。2-デオキシグルコースとメトホルミンによる腫瘍細胞におけるエネルギー産 生経路の2重の阻害が、食道癌や乳癌において抗腫瘍効果を認めたことや(26)、 複合体 I を標的とした酸化的リン酸化阻害剤が、脳腫瘍モデルマウスにおいて 強力な抗がん作用を示したこと(27)、さらに、進行がん患者における酸化的リ ン酸化阻害剤の臨床試験が行われた(28)などの報告が存在する。このように、
がんにおける酸化的リン酸化を標的とした治療法は非常に有望であると考えら れるが、未だ商品化に至った化合物は存在しない。また、神経芽腫において酸化 的リン酸化を標的とした治療法の研究報告は認めない。このことから、本研究で は、新規化合物のPEG-Bが酸化的リン酸化を阻害する作用を有することを証明 し、神経芽腫においても酸化的リン酸化が治療標的になりうることを示した点
34
で意義があると考える。
近年、がん細胞はその増殖・生存を維持する上で糖質への依存度が高いことに
着目し、糖質を制限したケトン食が欧米で注目されている(29、30)。日本でも、
ステージⅣ進行再発大腸がん、乳癌を対象としたケトン食による安全性と有効
性を評価するpilot studyが行われており、抗がん剤との併用でがんの縮小効果 を認めたと報告している(31)。本研究では、PEG-Bはグルコース欠乏下におい て顕著に効果を発揮したことから、ケトン食との併用により相乗効果が期待で
きる。今後、動物実験において、通常食およびケトン食飼育下でPEG-Bを投与 し、生体内におけるPEG-Bの抗腫瘍効果を検討することが必要であると考える。
また本研究では、PEG-Bがどのように複合体Ⅰに作用しているかについては 検討を行っていない。ミトコンドリア呼吸活性の検討実験より、PEG-B は Rotenoneと同様に、添加後速やかにOCRを低下させたことから、PEG-Bの作 用機序は遺伝子を介するものではなく、すみやかに活性中心に結合して効果を 表す反応と推測されるが、その証明については今後の課題となる。
35
【まとめ】
PEG-Bはミトコンドリア呼吸鎖複合体Ⅰを阻害することで、酸化的リン酸化
による効率的なATP産生を阻害する。このため、ATPを大量生産・消費して増 殖する腫瘍細胞に特異的に効果を表し、特にグルコース欠乏下においてPEG-B の効果が顕著になると考えられた。
36
【謝辞】
本研究を実行するにあたりまして、懇切丁寧な研究のご指導を賜りました日 本大学医学部内科学系総合内科学分野 藤原恭子助教に深謝いたします。
本研究に関して、ご指導・ご校閲を賜りました日本大学医学部外科学系小児外 科分野 越永從道教授、上原秀一郎准教授に深謝いたします。
本研究を行うにあたりまして、多大なるご協力をいただきました日本大学医 学部内科学系総合内科・総合診療医学分野 相馬正義教授、日本大学医学部総合 医学研究所難治疾患治療開発グループの皆様、ならびに日本大学医学部外科系 小児外科分野の医局員の皆様に深謝いたします。
37
【利益相反の開示】
本研究に関連し、開示すべき利益相反関係にある企業は以下の通りです。
受託研究・共同研究費:センカファーマシー社
38
【図】
図1:PEG-Bの構造 特許第5364532号
ポリエチレングリコールの誘導体およびそれを有効成分として含む抗腫瘍剤 株式会社 センカファーマシー社
39
図2:PEG-Bの腫瘍細胞および正常細胞に対する作用
PEG-Bのヒト表皮角化細胞に対する作用
PEG-Bの子宮頸癌Hela細胞に対する作用 特許第5364532号
ポリエチレングリコールの誘導体およびそれを有効成分として含む抗腫瘍剤 株式会社 センカファーマシー社
40
図3:各種細胞株におけるPEG-B添加後の細胞生存率(濃度変化)
-50 0 50 100 150
細胞生存率(%)
PEG-B濃度(μM)
NB9
* ** **
200 4060 10080 120
細胞生存率(%)
PEG-B濃度(μM)
HDF
* ** ** ****
0 50 100 150
細胞生存率(%)
PEG-B濃度(μM)
SK-N-AS
**** ** **
41
図4:NB9およびSK-N-ASにおけるPEG-B添加後の細胞増殖能
0 5 10 15 20 25 30 35 40 45 50
0 1 2 3 4 5 6 7
細胞増殖能
day
NB9
PEG-B (-) PEG-B(+)
** ** ** **
0 2 4 6 8 10 12 14 16 18 20
0 1 2 3 4 5 6 7
細胞増殖能
day
SK-N-AS
PEG-B (-) PEG-B (+)
** ** ** ** ** **
*
42
【PEG-B添加24時間後】
【PEG-B添加72時間後】
図5:細胞周期の解析
0%
20%
40%
60%
80%
100%
0μM 0.1μM 1μM
NB9
0%
20%
40%
60%
80%
100%
0μM 0.1μM 1μM
SK-N-AS
0%
20%
40%
60%
80%
100%
0μM 0.1μM 1μM
NB9
0%
20%
40%
60%
80%
100%
0μM 0.1μM 1μM
SK-N-AS
43
図6:グルコース欠乏下における細胞生存比
0 0.2 0.4 0.6 0.8 1 1.2
1 2 3 4 5 6
細胞生存比
NB9
0 0.2 0.4 0.6 0.8 1 1.2
1 2 3 4 5 6
細胞生存比
SK-N-AS
グルコース濃度 (g/L) PEG-B (1uM)
2
0 0.4
- + - + - +
**
** **
グルコース濃度 (g/L) PEG-B (1uM)
4.5
0 0.9
- + - + - +
** ** **
44
図7-1:グルコース欠乏下におけるアポトーシス細胞の割合①(NB9)
-5 0 5 10 15 20 25 30 35 40
0 0.4 2
アポトーシス細胞の比率(%)
グルコース濃度(g/L)
NB9
PEG-B (-) PEG-B (+)
**
45
図7-2:グルコース欠乏下におけるアポトーシス細胞の割合②(SK-N-AS)
0 5 10 15 20 25
0 0.9 4.5
アポトーシス細胞の比率(%)
グルコース濃度(g/L)
SK-N-AS
PEG-B (-) PEG-B (+)
**
** *
46
図8:グルコース欠乏下における細胞内ATP濃度
0 200 400 600 800 1000 1200 1400 1600 1800
1 2 3 4 5 6
細胞内ATP濃度(nM)
NB9
0 200 400 600 800 1000 1200 1400 1600 1800
1 2 3 4 5 6
細胞内ATP濃度(nM)
SK-N-AS
**
グルコース濃度 (g/L) PEG-B (1μM)
2
0 0.4
- + - + - +
*
グルコース濃度 (g/L) PEG-B (1μM)
4.5
0 0.9
- + - + - +
* *
47
図9-1 メタボローム解析(NB9)
48
図9-2 メタボローム解析(SK-N-AS)
49
図10:ミトコンドリア呼吸活性の検討
50
図11:呼吸鎖複合体Ⅰの活性変化
51
図12:呼吸鎖複合体Ⅱ/Ⅲの活性変化
52
図13:呼吸鎖複合体Ⅳの活性変化
53
図14:PEG-Bの標的―解糖系以外のATP産生経路
54
図15:PEG-Bの標的―ミトコンドリア呼吸鎖複合体
55
図16:PEG-Bの標的―呼吸鎖複合体Ⅰ~Ⅳのいずれか
56
【図説】
図1:PEG-Bの構造
新規がん治療薬の開発を目的とした化合物のスクリーニングを行い発見され た、子嚢菌由来のポリエチレングリコール化合物の誘導体PEG-Bの構造。
図2:PEG-Bの腫瘍細胞および正常細胞に対する作用
PEG-Bを添加すると腫瘍細胞である子宮頸癌Hela細胞の生存率(破線、四
角いプロットの曲線)は強く抑制されるが、正常細胞であるヒト表皮角化細胞
(実線、四角いプロットの曲線)に対しては10μM 以下では効果がなく、そ れ以上の高い濃度で初めて細胞生存率の抑制が認められた。
図3:各種細胞株におけるPEG-B添加後の細胞生存率(濃度変化)
NB9、SK-N-AS、HDFに0~50μMの濃度のPEG-Bを添加し、72時間後 の細胞生存率をWST-8 assayで解析した。結果、NB9およびSK-N-ASにお いて、PEG-Bは濃度依存的に細胞生存率を低下させた(NB9では、PEG-B 10 μM:*P=0.0162、20μM:**P=4.23E-03、50μM:**P=3.72E-03、SK-N- ASでは、1μM:**P=4.70E-06、10μM:**P=8.82E-05、20μM:
**P=1.74E-05、50μM:**P=1.51E-05)。一方、HDFでは高濃度のPEG-B 添加でも50%以上の細胞が生存していた(PEG-B 0.1μM:*P=0.0183、1μ M:**P=2.31E-04、10μM:**P=3.20E-03、20μM:**P=2.88E-03、50μ M:**P=3.57E-04)。
図4:NB9およびSK-N-ASにおけるPEG-B添加後の細胞増殖能
NB9およびSK-N-ASにおいて、PEG-B 1μMもしくは溶媒のみを添加後、
24 時間毎に 1 週間 WST-8 assay を行い、細胞増殖能を解析した。グラフは、
PEG-B 添加前(day0)を 1 とした増殖曲線で示している。結果、NB9 におい
てはPEG-B添加後3日目以降に増殖の抑制がみられるようになり、4日目より
非添加群との差が有意となった(左図、PEG-B添加後4日目:**P=8.67E-03、 5日目:**P=1.86E-04、6日目:**P=3.04E-05、7日目:**P=3.22E-07)。SK- N-ASにおいては、PEG-B添加後に細胞増殖は全くみられず、添加後1 日目よ り非添加群との間に有意な差を認めた(右図、PEG-B添加後1日目:*P=0.0198、 2日目:**P=6.29E-07、3日目:**P=4.60E-06、4日目:**P=2.16E-04、5日 目:**P=5.83E-04、6日目:**P=4.69E-04、7日目:**P=3.86E-05)。
図5:細胞周期の解析
57
NB9およびSK-N-ASにおいて、PEG-Bを添加して24、72時間後の細胞を 回収し、フローサイトメーターを用いて核内DNA量を解析した。結果、PEG- B添加24時間後においては、NB9、SK-N-ASともに、G0-G1期やG2/M期 の細胞の比率が増大したが、Sub-G1期の増加は認められなかった(左上図:
NB9、右上図:SK-N-AS)。一方、PEG-B添加72時間後においては、NB9、
SK-N-ASともに、Sub-G1期の細胞の比率の増加、すなわち、死細胞の増加が
認められた(左下図:NB9、右下図:SK-N-AS)。 図6:グルコース欠乏下における細胞生存比
NB9およびSK-N-ASにグルコース濃度が異なる培養条件下で1μMのPEG- B または溶媒のみを添加し、72 時間後に細胞生存比を WST-8 assay で解析し た。グラフは、PEG-B 非添加・グルコース通常濃度下培養群(NB9 は 2g/L、 SK-N-ASは4.5g/L)を1とした細胞生存比で示している。結果、NB9 、SK-N- AS ともに、グルコース不含培地において特に、PEG-B 添加による細胞生存比 の顕著な低下を認めた(上図の NB9 では、グルコース濃度 0g/L:**P=2.28E- 07、0.4g/L:**P=4.52E-05、2g/L:**P=1.50E-03、下図のSK-N-ASでは、グ ルコース濃度0g/L:**P=9.19E-04、0.9g/L:**P=6.49E-05、4.5g/L:**P=6.58E- 06)。
図7:グルコース欠乏下におけるアポトーシス細胞の割合
NB9およびSK-N-ASにグルコース濃度の異なる培養条件下で0.1、1μM のPEG-Bまたは溶媒のみを添加し、72時間後にAnnexin V-FITC apoptosis
detection kitを用いて、アポトーシス細胞の比率を解析した。四分画の図にお
いて、縦軸はPI、横軸はAnnexin Vの蛍光強度を表しており、Annexin V陽 性細胞である右上と右下の分画をアポトーシス細胞として計測した。結果、
NB9ではグルコース不含培地においてのみ、PEG-B添加によりアポトーシス 細胞の比率が顕著に増加し、非添加群と比べて有意に高い値を示した(図7- 1、**P=2.04E-03)。一方、SK-N-ASではいずれの培地においてもPEG-B添 加群は非添加群と比べてアポトーシス細胞の有意な増加を示したが、特に低グ ルコース培地、グルコース不含培地においてその差が顕著であった(図7-2、 グルコース濃度0g/L:**P=5.44E-03、0.9g/L:**P=1.68E-03、4.5g/L:
*P=0.0268)。
図8:グルコース欠乏下における細胞内ATP濃度
NB9およびSK-N-ASにグルコース濃度の異なる培養条件下で1μMのPEG- Bまたは溶媒のみを添加し、12時間培養後、Intracellular ATP測定キットを用
58
いて細胞内ATP濃度を測定した。結果、NB9ではグルコース不含培地において のみ、PEG-B添加により細胞内ATP濃度が有意に低下した(上図、*P=0.0145)。
一方、SK-N-AS ではいずれの培地においてもPEG-B 添加群は非添加群と比べ
て細胞内ATP濃度の有意な低下を示したが、特にグルコース不含培地において その差が顕著であった(下図、グルコース濃度 0g/L:*P=0.0217、0.9g/L:
*P=0.0243、4.5g/L:**P=1.78E-03)。 図9:メタボローム解析
メタボローム解析を行い、NB9およびSK-N-ASにおいて、PEG-B添加前 後で変化する代謝産物を解析した。HMT代謝物質ライブラリに登録されてい る代謝物質のうち、本実験で検出された物質を代謝経路に描画し、PEG-B添 加群と非添加群で明確な違いのみられた経路を示した。結果、NB9において
は、PEG-B添加群では非添加群と比較し、カルニチンの有意な低下
(*P=0.002)、アセチルカルニチンの増加傾向、NADHの有意な増加
(**P=0.001)が認められた。解糖系およびTCA回路については、一定の傾向
は認められなかった。SK-N-ASにおいては、PEG-B添加群では非添加群と比 較し、解糖系の亢進傾向、TCA回路の抑制傾向、カルニチンの有意な低下 (*P=0.029)、アセチルカルニチンの有意な増加(*P=0.044)、NADHの増加傾 向が認められた。
G6P: Glucose 6-phosphate, F6P: Fructose 6-phosphate, F1, 6P: Fructose 1, 6-diphosphate, GAP: Glyceraldehyde 3-phosphate, 3-PG: 3-Phosphoglyceric acid, PEP: Phosphoenolpyruvic acid, 2-OG: 2-Oxoglutaric acid.
図10:ミトコンドリア呼吸活性の検討
PEG-B によるミ トコンドリ アの呼 吸 活性の変化 を検討 す るため、Flux Analyzerで酸素消費速度(OCR)を計測した。結果、Control群ではOligomycin の添加によるOCR の低下、FCCP 添加によるOCR の上昇、およびRotenone 添加による OCR の低下が確認されたが、PEG-B 添加群では、Oligomycin や Rotenone添加時と同程度のOCRの低下を示した。またPEG-BによるOCRの 抑制はFCCP添加によっても解除されなかった(上図:NB9、下図:SK-N-AS)。 図11:呼吸鎖複合体Ⅰの活性変化
PEG-B による呼吸鎖複合体Ⅰの活性変化について、MitoCheck Complex I Activity Assay Kitを用いて解析した。ポジティブコントロールとして既知の阻 害剤であるRotenoneを使用した。結果、PEG-Bは1μM以上で有意に呼吸鎖 複合体Ⅰの活性を阻害した(Rotenone 2μM:**P=8.76E-05、PEG-B 1μM:
59
**P=9.17E-03、PEG-B 2μM:**P=6.41E-03)。 図12:呼吸鎖複合体Ⅱ/Ⅲの活性変化
PEG-Bによる呼吸鎖複合体Ⅱ/Ⅲの活性変化について、MitoCheck Complex II/III Activity Assay Kitを用いて解析した。ポジティブコントロールとして既 知の阻害剤であるAntimycinを使用した。結果、PEG-Bは0.2、1、2μMの いずれの濃度においても、control群と比較して有意な差を認めなかった
(Antimycin 5μM:**P=1.07E-06)。 図13:呼吸鎖複合体Ⅳの活性変化
PEG-Bによる呼吸鎖複合体Ⅳの活性変化について、MitoCheck Complex IV Activity Assay Kitを用いて解析した。ポジティブコントロールとして既知の 阻害剤であるKCNを使用した。結果、PEG-Bは0.2、1、2μMのいずれの濃 度においても、control群と比較して有意な差を認めなかった(KCN 100μ M:*P=0.0268)。
図14:PEG-B標的―解糖系以外のATP産生経路
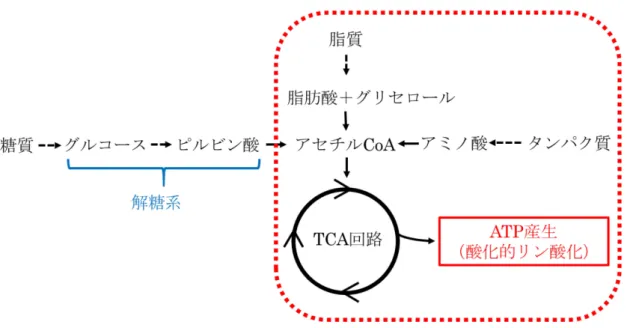
グルコース欠乏実験の結果から、PEG-Bの標的は図の赤破線枠内に示され る、解糖系以外のATP産生経路である可能性が考えられた。
図15:PEG-Bの標的―ミトコンドリア呼吸鎖複合体
メタボローム解析の結果から、PEG-B添加により、細胞のエネルギー産生に おいて解糖系が優位になりTCA回路が抑制されている可能性が示唆された。カ ルニチンは、解糖系の亢進およびTCA回路の抑制により増加したアセチルCoA を消費するのに利用されるため低下する。この時にアセチルCoAがカルニチン にアセチル基を渡すため、アセチルカルニチンが増加する。一方、TCA回路で 産生されるNADH量は増加していることから、PEG-B添加によりTCA回路自 体は阻害されず、TCA回路で生じたNADHからATPを産生する過程、すなわ ち、ミトコンドリア呼吸鎖複合体が阻害されている可能性が考えられた。
NAD+: Nicotinamide adenine dinucleotide.
図16:PEG-Bの標的―呼吸鎖複合体Ⅰ~Ⅳのいずれか
メタボローム解析の結果から、PEG-Bはミトコンドリア呼吸鎖複合体を阻害 している可能性が考えられた。またミトコンドリア呼吸活性の検討実験におい
て、PEG-B はミトコンドリア呼吸を阻害していることが確認できた。さらに、
PEG-B による OCR の抑制は FCCP 添加によっても解除されなかったことか
60
ら、複合体ⅤはPEG-Bの標的として除外できると考えられた。以上より、PEG- Bは呼吸鎖Ⅰ~Ⅳのいずれかを阻害すると考えられた。
ADP: Adenosine diphosphate.