九州大学学術情報リポジトリ
Kyushu University Institutional Repository
カイコセクロピンB抗菌性タンパク質の遺伝子解析と 誘導機構
谷合, 幹代子
https://doi.org/10.11501/3075497
出版情報:Kyushu University, 1993, 博士(農学), 論文博士 バージョン:
権利関係:
カイコセクロビンB抗菌性タンパク質の 遺伝子解析と誘導機構
谷合 幹代子
199 3
目 次
緒 言一一一一
第I章 カイコセクロビンBのcDNAの解析一一一一一一一一一一一一一一- 8 材料と方法一一一一一一一一一一一一一一一一一一一一一一一一10
結 果一一一一一一一一一一一一一一一一一一一一一一一一15
考 察一一一一一一一一一一一一一一一一一一一一一一一一26 摘 要一一一一一一一一一一一一一一一一一一一一一一一一30
第E章 カイコセクロビンBを誘導する因子の解析一一一一一一一一一一一一32 材料と方法一一一一一一一一一一一一一一一一一一一一一一一一34
結 果一一一一一一一一一一一一一一一一一一一一一一一一39
考 察一一一一一一一一一一一--・---一一一一一一一一一49 摘 要一一一一一一一一一一一…一一一--・--- -一一一一一---52
第E章 頼粒細胞のセクロピンB誘導への関与一一一一一一一一一一一一一---53 材料と方法一一一一一一一一一一一一一一一一一一一一一一一一54
結 果一一一一一一一一一一一一一一一一一一一一一一一---57
考 察一一
摘 要一一一一一一一一一一一一一一一一一一一一一一一一-66
第N章 カイコセクロビンB遺伝子の解析---ー一一一一---ー一一一一一67 材料と方法一一一一一一一一一一一一一一一一一一一一一一一一68
結 果一一一一一一一一一一一一一一一一一一一一一一一一73
考 察 一一一一一一一一一一一一一一一一一一一一一一一一91 摘 要 一一一一一一一一一一一一一一一一一一一一一一一一96
総 合 考察 一一一一一一一一一一一一一一一一一一一一一一一一一一一一一一-98
SUMMARY---一一一一一ー一一一一ーー一一一ーー一一一一一ーーーー一一ー一一ーー一一一ー一一ー104
謝 辞 一一一一一一一一一一一一一一一一一一一一一一一一一一一一一一一一107
引用 文献 一一一一一一一一一---四一一一一一一一一一一一一一一一一一一一108
-・・‘
点者
仁コ昆虫の生体防御 機構 は細胞性反応に基づくものと液性 反応に基づく
ものとの2種類に大別される。 細胞性防御 は? 血液細胞による異物の 捕食であり(Salt, 1970;和合, 1983; Lackie, 1988) , 血液細胞が異物の 周辺に集合して小癒を形成し 異物の分散を阻止する反応である。 大型 の異物の場合 は包囲化作用によりカプセルが形成 される。 一方4 液性 防御機構には, 異物の侵入によって体液中に常在する酵素が活性化す る反応および遺伝子が発現してタンパク質を合成する反応の2通りが
ある。 前者は,体液中のフェノール酸化 酵素前駆体が傷 または異物(生
物 および非生物〉の侵入により直ちに活性化型に変化し , これがチロ シン酸化 反応系を促進してメラニン 形成を導き, このメラニンによっ て傷を塞ぐと共に異物を封じ込める反応である(Ashida and Yamazaki,
1990)。 後者は, 細菌が侵入すると遺伝子発現により数時間のうちに 複数の抗菌性 タンパク質 またはレクチンなどが合成・分泌される反応 である(Boman and Hultma rkヲ1987)。 セン チニクバエSarcophaga
peregrlnaでは生体に傷を付与するか または細菌の注射による接種でレ クチンが誘導されることが明らかにされている(Komano et aJ., 1980)。
レクチンの生体防御における機能はまだ解明されていないが, 異物認 識のためのオプ、ノニン効果があるものと考えられている。
1
抗菌性タンパク質として, 体液中にリゾチームが存在することが以 前から知られていた(河原畑, 1970; Kawarabata, 1971; Powning and
Davidson, 1973)。 誘導性の抗菌性タンパク質として最初に報告され たのはセクロピアサンHyalophora cecropiaのセクロビンとアタシンで ある。 Enterob acter cloacaeを注射したセクロピアサン踊からセクロビ ンA, Bが精製され(Hultmark et al., 1980) , それらのアミノ酸配列 が明らかにされた(Steiner et al., 1981)。 その後ヲ 昆虫の抗菌性タン パク質の研究は活発かっ急速に進められ 双麹目, 鱗麹自, 膜題目,
鞘麹目および脈麹目から様々な分子種が発見された。 昆虫の抗菌性タ ンパク質は構造上の特徴からいくつかのファミリーに分類されている (Kimbrell, 1991)。 まず, セクロビンファミリー〈分子量約4ヲ000) に属するタンパク質がカイコBombyx mori (Shiba et al., 1983) , タノく コスズメガManduca sex臼(Dickenson et al., 1988) , サクサン
Antheraea pernyi (Qu et al., 1982) , センチニクノくエ(Kanai and
Natori,
1989) , ショウジョウノくエDrosophila melanogaster (Kylsten et al.,
1990) , ツェツェノくエGlossina morsitans morsitans (Kaaya et al., 1987) から分離された。 セクロビン類はA, B, CおよびD型に分けられて おり, 通常これらのうち2つ以上が1種類の昆虫から精製されている。
セクロビン類の抗菌スペクトルは広く, 特にAおよびB型はグラム陽 性, 陰性菌の両方に幅広く効果がある。 これらタンパク質は2個のα
ヘリックスを持つ両親媒性の陽イオンタンパク質で, C末端の長い疎
水性部位が細菌の細胞膜の脂質に結合してイオンチャンネルを形成す
ることが人工膜を用いた実験により明らかにされた。 セクロビン類は 真核生物の細胞に対して毒性を有しないことが報告された(Steiner et
al., 1988; Christensen et al., 1988)。 次に, アタシンファミリー(分子 量2 0,000�2 3,000)と称する1群のタンパク質はグリシンに富むGド メインを持つことが特徴である。 セクロピアサンから酸性および塩基
性アタシンが精製され(Hultmark et al., 1983) , センチニクバエから アタシンに類似のザルコトキシン11(分子量24,000)が分離された (Ando et al・ヲ1987)。 アタシンおよびザルコトキシン11は主として増
殖期の大腸菌に効果があり, 外膜タンパク質の合成阻害により菌の形 態異常を引き起こすものとされている(Carlsson et al., 1991)。 アミ ノ酸配列は異なるが同じくGドメインを持つタンパク質として, ニク
パエPhorrnia terra刀ovaeから分子量9,000のディプテリシン(Dimarcq et al., 1988)およびゴミムシダマシ類の一種, Zophobas atratusから分 子量 8,000のコレオプテリシン(Bulet et al., 1991)が精製された。 コ レオプテリシンの抗菌スペクトルは非常に狭く, 若干のグラム陰性菌 と1種類のグラム陽性菌に効果がみられるのみであった。 また, 日甫乳 類のマクロファージが持つ抗菌ペプチドであるディフェンシン
(Lehrer et al., 1991)と相向性のあるペプチドが昆虫にも存在するこ とが明らかになった。 すなわち分子量4,000の昆虫ディフェンシンが
ニクバエから(Lambert et al., 1989) , 同じく分子量4,000のザーペシ ンがセンチニクバエから(Matsuyama and Natori, 1988) , 分子量
10,500のヒメノプタエシンがミツバチApismelliferaの成虫から
(Casteels
etal., 1993) , 分子量5,5 00のロイヤリシンがミツバチのロ イヤルゼリーから(Fujiwara
etal., 1990)それぞれ精製されている。
その他,ゴミムシダマシ類(Bulet
etal., 1991)および、マダラヤンマ
Aeschna cyanea
(Bulet
etal., 1992) からも同様なタンパク質が精製さ れている。 ディフェンシンはグラム陽性菌に効果を有する。 また, プ
ロリンに富む(Pドメインをもっ〉特徴的なペプチドがミツバチから 2種類, ショウジョウバエから1種類精製されている。 すなわち分子 量2,000のアピダエシン(Casteels
etal., 1989) , 分子量4ヲ000のアバ
エシン(Casteels
etal., 1990)および分子量2,400のドロソシン(Bulet
et
al., 1993)などである。 いずれも非常に短いペプチドであり, アピ ダエシンはグラム陰性菌に, アバエシンは数種のグラム陽および陰性
菌双方に作用する。 ドロソシンは糖鎖を有しており, 糖鎖を除くと抗 菌活性が低下する。
以上の抗菌タンパク質のうち若干についてはcDNAが単離され,
シグナルペプチドを含めた前駆体構造が明らかにされた。 cDNAを プローブとしたノーザンブロッテイングにより, 個々のタンパク質に ついて合成器官および誘導のタイムコース(Kato
etal., 1993) が調べ られ, 体液中の抗菌活性と比較して, はるかに高い感度で誘導が証明 された。 このような実験において, 抗菌性タンパク質には生体防御に 働くのみならず発育段階特異的な遺伝子発現を行うものがいくつか認 められている。 ショウジョウバエのセクロビン(Tryselius
et al., 1992),
ディプテリシン(Reichhart
etal., 1992) , センチニクバエのザーペシ
ン(Komano
etal., 1991)などである。 この事実は2重機能タンパク質 として発生における形質発現制御および機能分化の面から興味が持た れる。
抗菌性タンパク質はし1ずれも細菌を注射すると極めて短時間のうち に当該遺伝子が発現するため, 晴乳類のc-反応性タンパク質, 血疑 アミロイドAなどと同様に生体防御関連急性期タンパク質と称するこ とができる。 晴乳類ではヲ リポポリサッカライドまたはペプチドグリ カンの刺激に基づき, 白血球などから分泌されるサイトカイン類をメ
デ、イエーターとして急性期タンパク質遺伝子が発現する(Heinrich
etal., 1990)。 一方, 昆虫ではサイトカイン類似タンパク質はし、まだ見 出されていない。 それにもかかわらず, アタシン, ディプテリシンな どの遺伝子解析において? 当該遺伝子の上流に晴乳類急性期タンパク 質遺伝子の発現制御に働く核タンパク質結合モチーフすなわちN F
κB結合部位(Sen and Baltimore, 1986 ) が認められた(Sun and Faye,
1992)。 この事実から, 昆虫における細胞性反応と液性反応とが互い に関連を有する可能性が考えられる。 また? 抗菌性こそ認められない が, 細菌の注射によって抗菌性タンパク質類と共に誘導されるタンパ ク質ヘモリンがセクロピアサンから精製され, このタンパク質は遺伝 子解析の結果? イムノグロプリンスーパーファミリーに属することが
明らかにされた(Sun
etal., 1990)。 イムノグロプリンスーパーファ ミリーとしてはこれまで多数の細胞表面認識タンパク質が知られてお
り7 昆虫ではショウジョウバエのamalgam遺伝子産物(Seeger
etal.ラ
1988) , バッタのFasciclin遺伝子産物(Harrelsonand Goodman, 1988) など佐発生・神経成長に関わるタンパク質があるが, 生体防御関連タ ンパク質については初めての報告である。 ヘモリンは異物の認識に働 くと推定されている。 昆虫は抗原抗体反応を持たないため, 昆虫の生 体防御機構は脊椎動物のものと全く異なっていると信じられていたが,
上述のように晴乳類との類似性が報告された結果, 共通の機構の存在 も考慮に入れる必要を生じた。 この情報は遺伝子解析により得られた ものであり? この方法によって異なる昆虫の生体防御関連タンパク質 の遺伝子とその発現を調べることが急務である。
このような観点に立って, 本論文はカイコのセクロビンをコードす る遺伝子の構造と発現誘導機構について研究することを目的とした。
カイコはショウジョウバエと並んで遺伝学的, 生理・生化学的蓄積が 豊富である上, 産業的な要請から病理学の研究も進展している。 その 成果の1っとしてカイコでは無菌飼育が可能となったことの意義は大 きい。 このようにカイコは生体防御研究にとって著しく有利な昆虫で あると考えられる。
まず, 第I章ではセクロビンBのcDNAクローンの単離を行った。
これはカイコの抗菌性タンパク質遺伝子としては最初の報告である。
続いてセクロビンB誘導の基礎的研究を行った。 第E章でセクロビン
Bを活性化する因子について追究し, 第E章で細胞性防御反応との関
わりを検討した。 細菌の細胞壁構成成分が食細胞によって遊離すると
いう報告は? 昆虫ではこれが初めてである。 さらに第W章ではゲノム
ライブラリーを用いてセクロビンB遺伝子の解析を行い, 同遺伝子の 多重化とそれに関わるトランスポゾン様因子および遺伝子上流域の特 徴について検討した。
本研究の結果, 細胞性防御反応と液性防御反応とは独立したもので なく, 一連の反応であることが確認できた。 またセクロビンB遺伝子 上流には晴乳類イムノグロブリン遺伝子で報告されている菌体リポポ リサッカライド応答性の制御配列が見出され, 昆虫においても脊椎動 物の免疫機構におけるものと類似した遺伝子発現様式が存在するもの
と推察した。
第I章 カイコセクロビンBのcDNAの解析
カイコセクロビン類の発見と確認の経緯は以下のとおりである。 ま ず, 大腸菌を注射したカイコ幼虫から 35個のアミノ酸残基から成る
2種類の抗菌性タンパク質が分離され, レピドプテランと命名された (Shiba
eta1., 1983)。 いずれもセクロピアサンのセクロピンBとよく 類似しており, アミメ酸5'"'-'8残基が異なるのみであった。 その後,
同様にして合計3種類のレピドプテランが分離され, それぞれレピド プテランA, B, Cとして報告された(Teshima
eta1., 1986, 1987)。
別の研究グループによってもこの3種と同ーのタンパク質が精製され,
それらはセクロビンB2, B 1ヲ A1として発表されている
(Morishima
eta1., 1990)。 この3種に加えて, カイコからアミノ酸 35 残基から成るレピドプテランタイプの抗菌性タンパク質CM IV
(Zhang
eta1., 1985)およびアミノ酸36残基から成りセクロピアサン のセクロビンDに類似したCM2 P h 1 (Tu
eta1., 1989)が精製され,
それぞれアミノ酸配列が報告された。 以上のようにカイコでは誘導性 の抗菌性タンパク質として少なくとも5種類のセクロビンが確認されヲ アミノ酸配列が解析されており, さらに他の種類の抗菌性タンパク質 の存在も考えられる。
カイコにおける液性防御反応をセクロビンの発現・誘導機構を指標
8
として研究するに当たり, 抗菌活性のみを指標として誘導の検定を行 った場合には複数の異なる抗菌タンパク質の存在によって解析が困難 となると思われる。 特定のタンパク質の挙動を把握することが有効な 知見を得るために必須であり, c DNAのクローニングおよび遺伝子 解析はそのための方法としても有効である。 しかし, カイコのセクロ ビンについてはcDNA ・遺伝子レベルの研究は全く行われていない。
そこで, 本章においてカイコからセクロビンcDNAのクローニング を試みた(遺伝子解析については第W章で述べる〉。
カイコ5齢幼虫を大腸菌で免疫し脂肪体からcDNAライブラリー
を作製した。 一方, セクロビンBのアミノ酸配列に基づいてセクロビ
ン遺伝子の1部分に対応する塩基配列をDNA合成装置で作製し 得
られた断片をプライマーとしてpolymerase chain reaction (P C R )を
行いテンプレートとして用いたcDNAの一部を増幅させた。 この部
分配列をプローブとしてライブラリーから4種類のcDNAクローン
を得た。 この4個のクローンの塩基配列を決定し, これまでに報告さ
れている他の昆虫のセクロビン類とのアミノ酸配列および塩基配列と
の比較を行った。
材料と方法
カイコおよび細菌
カイコ(東海×朝日〉は人工飼料(日本農産工〉によっておOCで飼 育した。 Escherichia coli (大腸菌 ) K12株由来のDH1株(Low,1968) はLB培地(Maniatis et al., 1982)中で370Cで培養し,170 gで10分間
遠心分離して沈澱させ, 昆虫用生理食塩水(1 P S : 150 mM NaCl, 5 mM KCl, pH 6.8)で、懸濁・洗浄し再度遠心した沈澱を少量の1 P Sで 浮遊させ, 40Cで保存した。
cDNAライブラリーの作製
5齢5日幼虫5頭に大腸菌を1頭当たり4
x106個注射にて接種し,
9時間後に解剖して脂肪体を集めた。 0.56 gの脂肪体からQuick Prep mRNA Purification Kit (Pharmacia)を用いてmRNAを精製し, その
うち8μgからcDNA Synthesis Kit (Pharmacia)を用いてcDNAを 調整した。 cDNAの両末端にリガーゼ(160C, 15時間〉によって EcoRIリンカーを付着させ, えg t 10ベクター(Murray et al., 1977) のEcoRI部位に組み込み, Gigapack 11 Gold Packaging Extract
(Stratagene)を用いたin vitroノミッケージ法によってファージを構築
して大腸菌NM514に感染させ形質転換させた。
プローブの 作製
カイコセクロビンBのアミノ酸配列(Teshima
eta1., 198 6, 1987;
Morishima
eta1., 1990)をもとに, 合成DNA装置(AB 1 )により合 成プライマーを作製した。 5'プライマー(18塩基〉は成熟タンパク質 におけるアミノ酸残基第2番トリプト ファンから第7番リジンまでを もとにして5'四TGGAA(A/G)AT (A/C/T)TT(C/T)AA(A/G)AA(A/G)♂と した。 3 'プライマー(14塩基〉は第 24番プロリンから第28番ノくリン までをもとにして5'-AC(T/C)TC(NG!f)AT(A/G!f/C)GC(A/G/T/C)GG- 3'とした。 このプライマーをそれぞれ 1μM 用いてPCRを行った。
そのため40 ngのcDNAをテンプレートとして用い, 10 mM dNTP, 2.5単位Taq D N Aポリメラーゼ(Promega)および緩衝液 (50 mM KCl, 10 mM Tris-HCl緩衝液, pH 9.0, 1 %トリトンX-100,
1.5 mM MgC12)を混合し温度制御装置PC -700(Astec)中で900C 1分,
400C 2分, 720C 3分を1サイクルとして30サイクル反応させた。
PCR産物を4%低融点アガロース(Seakem)ゲルで電気泳動により 分離し 増幅が認められたDNAバンドを切り出して以下の方法で
DNAを精製した。 ゲル片をTE (10 mM Tris-HCl緩衝液, 1 mM
EDTA, pH 8.0)を加えて650Cで融解した後, 等量の TE飽和フェノ ールを加え 混合(Vortexを使用〉し, 6,000 g で5分間遠心分離し上清 を得た。 この操作( フェノール抽出〉をさらに2回行い? 上清をフェ ノール/クロロホルム(1 : 1, v/v) , クロロホルム/イソアミルアル
コール(24 : 1, v/v)で それぞれl回ずつ処理した後, 1/10量の3
M酢酸ナトリウムと2.5倍量の100%エタノールを加えて-800Cに15 分間放置し6,000 g で10分間遠心して沈澱を得た。 これを少量のTE
で溶解し精製DNAとして回収した。 このDNAをKlenowフラグメ ントにより末端を平滑にした後, 制限酵素即応11で、クローニングサイ トを切断したM13m
p18 ベクターに組込み, ダイデオキシ法(Sanger
et a1.,
1977)により塩基配列を決定した。 得られた結果がセクロビン
Bのアミノ酸をコードする配列と一致することを確認した。
プラークハイブリダイゼーション 5 -p-
cDNAライブラリー1
x10プフ ークを Nytranメンブレン
(Schleicher and Schuell)に転写し, 5
XSSPE (Maniatis
et a1.,1982) , 0.1 mg/ml サケ精子DNA, 50%ホルムアミドを含む5
XDenhardt's
液中で420C4時間プレハイブリダイゼーションを行った。 次に, DN Aラベリングキット(ニッポンジーン〉を用いて[α_32p]dC TPに
よりランダムラベルしたプローブをプレハイブリダイゼーション液に 加えて420Cで16時間ノ\イブリダイゼーションを行った。 その後メン ブレンを室温で2回ヲ 2
XSSC (Maniatis
et a1.,1982)により5分間 洗浄じ さらに600Cで2回, 0.2
XSSC (0.1% SDS含有〉で30分間 洗浄を行い, 乾燥後X線フィルムに感光させた。
えファージDNAの調製
プラークハイブリダイゼーションによるスクリーニングを3回繰り
返して得られた陽性ファージをSM緩衝液( 1,000 mlのH20にNaCl 5. 8 g, MgS0 4.η-I20 2.0 g, Tris 6.05 g, 2%ゼラチ ン5 mlを溶解し,
5 →←
pHをHClで7.5に調整 〉 に懸濁し, 10--' pfuを大腸図NM 5 1 4に吸着
させ, 4 mlのLBソ フトアガー(0.75%アガーを含むLB培地〉と 混合して9cm径のペトリ皿に固めたLBアガープレート(1.5%アガ ーを含むLB培地〉の表面に重層した。 370Cで一夜放置後, 4 mlの SM緩衝液を加え, テープでシールし, 40Cで 2�3時間緩やかに振 とうした。 この液を遠心管に移して40Cで200 gにて10 分間遠心分離
し 上清を20%ポリエチレングリコーノレ6000 と2M NaClを含む SM緩衝液と等量 混合した後, 氷上で1時間インキュベートした。 続 いて170 gで20分間遠心分離し沈澱をLB培地に浮遊させ, LB培
地に懸濁したD E 5 2-セルロース(Whatman)を加え 遠心分離して上清 を取る操作を2回行った。 この上清に50μg/mlプロテ イナーゼ、Kと 0.5% S D Sを室温で5分間反応させてファージ外皮タンパク質を溶解
し, 3M酢酸カリウム液を加えて8 80Cで 20 分間処理後, 10分間氷冷,
6,000 gで10分間遠心分離してタンパク質を除いた。 上清を等量のイ ソプロパノールと混合して-800Cに15分間置き6,000 gで10 分間遠心 分離し沈澱( DNA)を乾燥後少量のTEに浮遊させ-200Cで保存し
た。
塩基配列の決定
えファージ DNAからEcoRIによってインサートを切り出し, 1%
アガロースゲルで電気泳動をしてサイズを確認後, ゲルを切り出して
DNAフラグメントを精製した後, M13mp18ベクターにサブクロー ニングした。 得られたリコンビナントファージをXL-1-blue大腸菌に 感染させ形質転換させ, えファージと同様の方法でDNAを複製させ 調 製した。 SequenaseJ.'- Version 2.0 DNA Sequencing Kit (United States
RBiochemical Corp.)を用いてダイデオキシ法により塩基配列を決定し た。 このため, まずM13Sequencing Primer(-20) (TOYOBO)を用い,
このプライマーによって読みとった配列に基づいて合成DNAプライ
マーを作製し, さらに先を解析する方法を採用した0 32pでラベルさ れたDNA断片を6%アクリルアミドゲル上で電気泳動によって分離 後, ゲルをドライヤー〈アトー〉で乾燥させX線フィルムに感光させ
7こO
cDNA配列のコンビュータ解析
塩基配列について, SDS-Genetyxコンビューターソフト, すなわち
タンパク質の疎水性解析, 塩基配列のAT含量分布図作製, ハープロ
ット解析およびアミノ酸ホモロジー比較の各フ。ログラムを用いて解析
を行った。 タンパク質の疎水性・親水性解析においてはKyte and
Doolitle (1982)のパラメータを用いてアミノ酸5個ずつの平均値を計
算した。 AT含量分布図は核酸50bpごとにATの出現頻度を計算し
作成した。 ハープロット解析ではセクロピアサンおよびショウジョウ
バエのセクロビンBのcDNAとの比較を15塩基配列につき10塩基
配列が一致した場合に1個のドットを描く条件を設定して行った。
結 果
cDNAライブラリーおよびスクリーニング
合成プライマーによって増幅されたDNAは約80 bpの大きさを有
していた(Fig. 1)。 このDNAをM13m
p18 にクローニングして塩 基配列を解読したところ, 成熟タンパク質の第2番から第28番のアミ ノ酸配列に相当するものであった。 この大きさと配列はプライマーを 設定するに当たり予想されたものと完全に一致していた。
免疫したカイコ5齢幼虫の脂肪体から得られたcDNAライブラリ
6
_ r_ _ I _ -r:. -J- J.._ ...f"\5
ーのタイターは3.72
x10 v pfu I μgであった。 10 -' pfuを上記プロー
ブでスクリーニングしたところ, 3次スクリーニングでは約1ヲ000個 のプラークがポジティブであった。 従って陽性プラークは約1%に相 当した。 これらのポジティブプラークのうち30個を選んでDNAを 精製し, EcoRIで切断した。 インサートDNAのサイズは400�600 bpで, このうち約半分が約500 bp前後のものであった (典型例を
Fig.2に掲げる〉。 約500 bpのフラグメント4個(Fig. 2 のNo. 3, 19ヲ 21,24)を選び7 ゲルを切り出してDNAフラグメントを精製し, M 13m
p18ベクターにサブクロ ーニングしダイデオキシ法により全塩基
配列を決定した。
2 3
4トー
Fig. 1. Gel electrophoresis of amplified DNA by
polymerase chain reaction method.
DNA was analyzed by 4% agarose gel electrophoresis. Lane1:
Molecular size marker (ØX174 phage DNA digested with HincII) with the fragments of 1,057, 770, 612, 495, 392, 345,341, 335 and 297 bp. Lanes 2 and 3: DNAs ampli白ed by PCR.1μg of DNA was loaded per lane. Arrow indicates the position of 80 bp.
16
.J〆(ご・=一=一→rF一七ν--1J一一ご一一/-一{一ニ(}()『一三JJ〆三=υ{{一泊三{'一-一LJ'二:一三υ一(一二一/乞{{一U{一}一CU\-rLUニ 一 Z一戸-一rLIKK一一(Zv/ニχ-cFLFX乙Zuyニご一λ一J一〕一H4ムJ一ヲフ //一ピ~ニコJミ}.. Z イ目u-“・-一Iz--N.SFdd--』z-m吋一AZI-.-rz-umこzu一3y2ιと5回んJC//‘(一.角よ目んdlぜこzdAX一戸ZAV 三二)=ごU\)「一C」ち〈c一一之三JJ』三一(}Eυ={ぷ』一泊コ・ノ乙功υUコ一一寸ト二一/乞)54・一三一八く子三一=υν. 三一二U叫ヅムC/一一d功とよハJ5っ三一ミ一(一υ~と.
の門寸内内向cmmN∞N。NmNマN門内
r
p、J戸FFOF。NOF。FAF寸戸内
c) αコ
(v)
N∞向山仇寸llNF匂
。ドドヘ門的OF
芝
cDNAの塩基配列 および推定アミノ酸配列
塩基配列決定の結果およびそれより 推定したアミノ酸配列 をFig.3 に掲げる。 選んだ4個のcDNAはいずれもオープンリーデイングフ レームを含んでおり, アミノ酸配列 に翻訳すると全 て完全にセクロビ ンBをコードする ものであった。 そこで, これらをそれぞれBmcecB3,
BmcecB19, BmαcB21, BmcecB24 と命名した。 これら4種はアミノ
酸に影響しない部分の塩基配列 において互いに異なっており, Bmcec3 を基準に するとBmcec19 ,Bmcec21 , Bmcec24の順に それぞれ8,15フ30箇
所が異なっていた。 またBmcec24は,323-328と345 -348に他の3つの cDNAにはない挿入配列が見られた。 これらの 4種のcDNAのサイ ズはそれぞれ416, 420, 412, 402 bpであり ,翻訳開始コドンATG はEcoRIリンカーからBmcecB21では51-53, 他の3つでは54-56 の場所にあった。 成熟タンパク質 の アミノ酸 配列 (Morishima
etal.,
1990)の上流に , シグナルペプチドを含むと考えられるアミノ酸残基が 26個あった。 また, 成熟タンパク質ではC末端アミノ酸はイソロイシ ンであるが, c DNAの 配列ではさら にグリシ ンーリジンの2アミノ 酸が付加していた。 翻訳終止コドンTGAはBmcecB21では 240- 243 の位置に, BmcecB3, -B19, -B24では243 -246 の位置にあった。
3'非翻訳領域にはAT 2 - 4 A配列が数カ所 に みられた( Fi g.4)。 一般
的なポリA付加シグナルAATAA AAおよびポリAテイルは 4つのク
ローンの いずれにおいても認められなかったが, ポリA シグナルと類
似する配列 はいくつ かみられ, BmcecB3ではATAA Aが 394-398に,
BmcecB 3 GTACAGCGCCGCTTGTGTCTTAACGTCCGCGTTTGAATTTACATAAACTTACAATGAATT 60 BmcecB19 -ーーーーーーーーーーーーーーーーーーーーーーーーーーーー-C-ーーーーーーーーーーーーーーーーーーーーーーーーーーー- 60 BmcecB21 --GC--ーーーーーーーーーーーーーー甲『ーーーーー-G--ーーーーーーーーーーーーーーーーーーーーーーーーC- 57 BmcecB24 -ーーーーーーーーーーーーーーーーーーーーーーーーーーーーーC---ーーーーーーーーーーーーーーーーーーーーーーーーー- 60 Me七AsnPhe 3
BmcecB 3 TCGCAAAGATCCTATCCTTCGTCTTCGCTCTGGTGCTGGCTTTGAGCATGACCAGCGCTG 120 BmcecB19 ー由ーーーーーーーーーーーーーーーーーーーーーーーーーーーーーーーーーーーーーーーーーーーーーーーーーーーーーーーー--120 BmcecB21 -ーーーーーーーーーーーーーーーーーーーーーーーー由ーーーーーーーーーーーーーーーーーーーーーーーーーーーーーーーーーー117 BmcecB24 -ーーーーーーーーーーーーーーーーーーーーーーーーーーー四ーーーーーーーーーーーーーーーーーーーーーーーーーーーーーーー120 AlaLys工leLeuSerPheValPheAlaLeuValLeuAlaLeuSerMe七ThrSerAla 23
BmcecB 3 CTCCCGAGCCCAGGTGGAAGATCTTCAAGAAAATTGAAAAAATGGGCAGGAACATTCGTG180 BmcecB19 -ーーーーーーーーーーーーーーーーーーーーーーーーーーーーーーーーーーーーー『ーーーーーーーーーーーーーーーーーーーー-180 BmcecB21 ---ーーーーーーーーーーーーーーーーーーーーーーーーーーーーーーーーーーーーーーーーーーーーーーーーーーーーーーーー-177 BmcecB24 -ーーーーーーーーーーーーーーーーーーーーーーーーーーーーーーーーーー-GーーーーーーーーーーーーーーーーーーC----180
山叫pr
r
q叩山PheLysLys山luLysBmcecB 3 ACGGCATCGTCAAAGCGGGCCCGGCGATCGAGGTCCTTGGTTCGGCTAAAGCTATAGGAA240 BmcecB19 ----G-ーーーーーーーーーーーーーーーーーーーーーーーーーーーーーーーーーーーーーーーーーーーーーーーーーC-ーーー240 BmcecB21 --ーーーーーーーーーーーーーーーーーーーーーーーーーーーーーーーーーーーーーーーーーーーーーーーーーーーーーーーーーー237 BmcecB24 -ーーーーーーーーーーーーーーーーー四ーーーーーーーーーーーーーーーーーーC-ーーーーーーーーーーーーーC-ーーーーー-240
λspGly工levalLysAlaGlyProAla工leGluValLeuGlyserAlaLysAla工leGly 63
BmcecB 3 AATGATCTTTAAAA CGACTGCCATAAATACTAGTATTATTTλATCTACTCGTTAATTTTλ300 BmcecB19 -ーーーーーーーーーーーーーーーーーーーーーーーーーーー-T-ーーーーーーーーーーーーーーーーーーーーーーーーーーーーー300 BmcecB21 -ーーーー-A-ーーーーーーー-TA-A-ーーーーーーーーーーーーーーーーーーーA-ーーーーーーーーーーーーーーーーーー29 7 BmcecB24 ---ーーーーーーーーーーーー-TAC--ーーーーーーーーーーーーーーーーーーーーー---ーーーーーーーーーーーーー300
Lys***
BmcecB 3 ATATGTAATTTAATTACAGCTT BmcecB19 ---ーーー----A-ーーーーーーーーー
BmcecB21 ---ーーーーーー由A---ーーーーーーーー ーーーーーー-C----ーーーー
nu nU 7' RJ FhJ 凋HE 3J 吋3 吋3 巾A- -
巾- -
巾・
- -
RA
-
、n
-
n』
-
巾4 -
m--
-
A - G 円」
-
EM -
円u -
-
TAAGACTTAATAATAA
BmcecB24 ー四ーーーーーーー-A-ーーーーー-AT-ACTCGTA---CG--TTCC-TTAT--T-ーーーーーーー-360
BmcecB 3 TAλATATAATTTGAAGTTAATGTCTGCCAAAGTTTTCTTAGTGATAλλCGTTATGTTGTA 410 BmcecB19 -ーーーーーーーーーーーーーーーーーーーーーーーーーーーーーーーーーーーーーーーーーーーーーーーーーーーーーーーーーC-410 BmcecB21 ---ーーーーーーーーーーーーーーーーーーーーーーーーーーーーー---C-ーーーーーーーー--G-ーーーーーーーーーー-407 BmcecB24 -ーーーーーーーーーーーーー-T---ーーーーーーーーーーーーーーーーーーーーーーー 402
BmcecB 3 AAAA TT BmcecB19 ー由ーT-AAAAA BmcecB21 ---T-
416 420 412
Fig. 3. Nucleotide and deduced amino acid sequences of four
cecropin B cDNAs from the silkworm, Bombyx mori. Dashed lines indicate the nucleotides which are identical to those of BmcecB3. The termination codon is indicated by asterisks. Arrow head shows the start position of mature peptides.
N Cコ
BmcecB 3
BmcecB19
BmcecB21
BmcecB24
5・-245 常肯肯肯肯 大台肯*肯* ****�同****
TCTTTAAAACGACTGCCATAAATACTAGTATTATTTAATCTACTCGTTAATTTTAATATGTAATTTAATTACAGCTTTAAGACTTAATAA
***合** 416-3'
TAAGACATTCAATTTTAAATATAATTTGAAGTTAATGTCTGCCAAAGTTTTCTTAGTGATAAACGTTATGTTGTAAAAATT
5'-245 ***セ 女******* 合犬、骨肯背骨 **** ****
TCTTTAAAACGACTGCCATAAATATTAGTATTATTTAATCTACTCGTTAATTTTAATATGTAATTAAATTACAGCTTTAAGACTTAATAA
合台育合*背 合脅*安 420-3'
TAAGACATTCAATTTTAAATATAATTTGAAGTTAATGTCTGCCAAAGTTTTCTTAGTGATAAACGTTATGTTGCAAAATTAAAAA
5'ー242 ******* *官、t合** **** ****
TATTTAAAACGTATACCATAAATACTAGTATTATTAAATCTACTCGTTAATTTTAATATGTAATTAAATTACAGCTTTAAGACTCAATAA
****** 412-3'
TAAGACGTTCAATTTTAAATATAATTTGAAGTTAATGTCTGCCAAAGTTTCCTTAGTGATAAGCGTTATGTTGTAAAATT
5'-245 ****合 安*台育台育 ***女 **** ***セ
TCTTTAAAACGTACGCCATAAATACTAGTATTATTTAATCTACTCGTTAATTTTAATATGTAATTAAATTACAATTACTCGTATAAGACT
台**肯 安*官官官官 安安*合* 402-3'
CGATTTCCATTATGATATTCAATTTTAAATATAATTTGAATTTAATGTCTGCCAAAGTTTTCTTAGT
Fig. 4. Nucleotide sequences of 3' non-coding regions of cecropin B cDNAs.
AT2_4A elements and possible poly
(
A+)
signals are indicated by asterisks and underlines, respectively.Bm∞cB19 ではAATTAAA が 41 2-418 に, ATAAA が 39 6 -40 0 に位 置していた(Fig. 4)。
コンビューター解析
得られたcDNAクローンのオープンリーデイングフレームから読 み取られるセクロビンBタンパク質のアミノ酸配列およびその基とな る塩基配列に基づいて, コンビューター解析により構造上の特徴を推 定した。 Fig.5に疎水性アミノ酸残基の分布を示す。 シグナルペプチ
ドおよび成熟ペプチドのC末側の約半分は疎水'性に富んで、おり, 成熟 ペプチドのN末側約半分は親水性で、あることが明らかとなった。 塩基 配列のAT含量はFig. 6 に示すように3'非翻訳領域で著しく高く,
BmcecB3, -19, -21, -24でそれ ぞれ 78 .8, 78.9, 76.0, 79 .1 %で
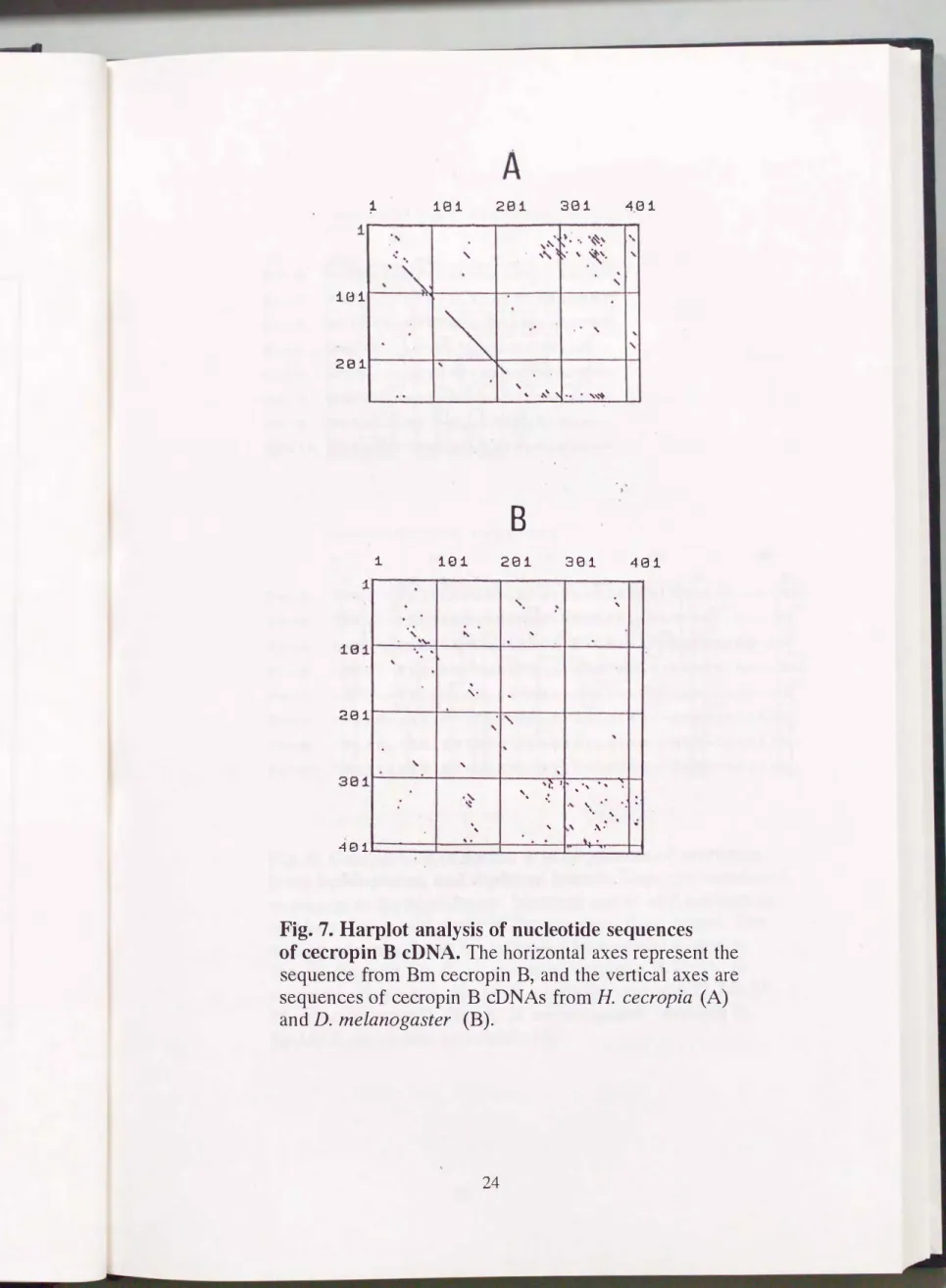
あった。 ハープロット解析によると(Fig. 7) , 塩基配列においてカ イコセクロビンBは同じ鱗趨目であるセクロピアサンのセクロビンB と相向性が高い一方, 双麹目であるキイロショウジョウバエのセクロ ビンBとは相向性が低し これらのホモロジーの値はそれぞれ71.3,
47.2%であった。 しかし, アミノ酸配列レベルでセクロビン類を比較 すると, キイロショウジョウバエをも含めた数種昆虫において高度に 保存されている領域が存在する(Fig.8 )。 特に黒三角で図中に表示 した残基は種聞を越えて保存されており, これはシグナルペプチド領
域で6箇所, 成熟ペプチド領域で7箇所数えられた。 相向性の高い領
域はここにあげた鱗題目のタンパク質において大きし 双麹目では小
.寸N 凶 ωω ωg 凶 -向。ωωロωロ
c ,
.
� �
m 寸ヨ
困層圏・ ...
.苫巳4u ω 伺巳4o (l)
2 5
同 �
æ a 匂ωωロ唱ω聞こ。h判明ぷSOSac-M匂h国.mu.9同
同OロOZ窃O【ごHS∞ω召∞ω定。司自vS区OMM〈000.寸
22
0000 .寸
doH∞
ω』∞店℃。υ'ロOロ・何回。口。ヨ∞。仏ロS∞
ω召∞ωおυ一
℃店診OHH〈
do吉岡《回一』封切宅苫ω言。ω'Hd刊』。sgω何一。.唱・ω日
』Ea・2Z 6"e回 。ωの
23
。旧
% .LV
。
。
44 . 44 191 281
A
3B1 4811臼1
2臼1
B
1 181 281 381 481
11Jl
-、 、1 日 1
r 子 !
、\・
281 、
"
"、 、
-、\= ‘、、 ・" 、 、
、、 、
、、 、 入・
. ー. ・
401
Fig. 7. Harplot analysis of nucleotide sequences of cecropin B CDNA.The horizontal axes represent the sequence from Bm∞cropin B, and the vertical axes are sequences of ωcropin B cDNAs from H cecropia (A) and D. melanogaster (B).
24
�Slgnal peptldes
-20 -10
Bm-B Hc-B Hc-A
一 一 一 A A pu nH QM QM N Q FU Fu j L A 一S一F -a v
--刈lEUN 「tLIL --ullU [以lIH仇 ーートllト
v v F F V I F F JK」川 M川 nH
F .
N F M N M
A
D D A B
- 一 一 -
c s m m p H M D D S
�Mature peplldes
+1 +10 +初 +30 +40
v Bm-B
Hc-B Hc-A Hc-D Ms-D Dm-A Dm-B
Fig. 8. Comparison of amino acid sequences of cecropins from lepidopteran and dipteran insects. Gaps are introduced to maximize the homologies. Identical amino acid residues in one letter code with those of Bm cecropin B are boxed. The conserved amino acids are indicated by reversed triangles.
Bm-B: BmcecB, Hc-B: H cecropia cecropin B, Hc-A: H cecropia cecropin A, Hc-D: H cecropia∞cropin D, Ms-D:
M.sαta cecropin D, Dm-A: D. melanogaster∞cropin B,
Sp-IA: S. peregrina sarcotoxin IA.
さかった。 次に, カイコセクロビンB全アミノ酸配列をセクロピアサ ンのセクロビンA, B, C, タノくコスズメガのセクロビンD, ショウ ジョウノくエのセクロビンA, Bヲ センチニクノくエのザルコトキシンI Aの配列と比較した結果, Table 1に掲げるように, 塩基配列の場合 と同様, カイコセクロビンBは鱗麹目セクロビンBと相向性が高く,
双麹目のものとの相向性は低かった。 これはシグナルペプチド領域お よび成熟ペプチド領域双方にあてはまることである。 特に成熟ペプチ ド領域において双題目セクロビンとの差異が大であった。
考 察
カイコセクロビンBの4種のcDNAは , BmcecB3を基準にすると BmcecB19, - 21, - 24 の順に8, 15, 30箇所の塩基が異なっていた。
このようなcDNAの多様性はセクロビンB遺伝子の重複または個体 聞の多様性の反映であると推察される。 cDNAクローンにポリAテ イルやポリAシグナルが含まれなかった原因は, 3 I非翻訳領域が極め てATに富むためである可能性がある。 cDNAライブラリーを作製 する際に用いたオリゴ(d T)がポリAテイルの代わりにATに富む 部位にアニーリングしたので、あろう。 このATに富む領域においては,
A T 2-4Aエレメントが数カ所存在した。 このエレメントは, ヒトの
リンホカイン遺伝子GM-CSFおよびß-グロビン遺伝子などのよ
Table 1. Comparison of the homology in amino acid sequences deduced from nucleotide sequences of cecropin cDNAs between of Bombyx mori cecropin B and
cecropins from other insects. Designations for cecropins are as described in Fig. 8.
Signal peptide
(
1ρpidoptera) (%)
Hc・.B 76.9
Hc-A
61.5
Hc-D 57.7
Ms-D 53.9
(
Diptera)
Dm-A Dm-B
主主
Mature peptide
)6
ユ
5fb
%
0381
/t10δ〆oqu今コ
5 ユ5 oa 6
8
弓3 4・ 司、)
23.8 23.8 23.8
27