東京農業大学
博士論文
中国産食用茸ツブイボタケ (Thelephora vialis) 含有
vialinin A の RBL-2H3 細胞における TNF-α 産生阻害
メカニズムの解明
平成 26 年 3 月 20 日
食品栄養学専攻
吉岡 泰淳
東京農業大学
博士論文
中国産食用茸ツブイボタケ (Thelephora vialis) 含有
vialinin A の RBL-2H3 細胞における TNF-α 産生阻害
メカニズムの解明
指導教授 阿部尚樹
平成 26 年 3 月 20 日
食品栄養学専攻
吉岡 泰淳
目次
目次
目次
目次
Abstract
序論
... 1使用機器
... 11使用試薬
... 13第一章 Vialinin A の TNF-α 発現阻害活性の様式の検討
第一節 Vialinin A 及び DMT の TNF-α mRNA 発現への影響 ... 17
第二節 Vialinin A 及び DMT の TNF-α 産生量と放出量 ... 19
第三節 考察 ... 21
第四節 実験の部 ... 23
第二章 Vialinin A 標的分子の探索
第一節 探索プローブの調製 ... 32
第二節 探索プローブの TNF-α 放出阻害活性試験 ... 34
第三節 DMT の RBL-2H3 細胞内での局在性の検討 ... 40
第四節 Biotin-DMT を用いた vialinin A 標的分子の探索 ... 43
第五節 考察 ... 48
第六節 実験の部 ... 50
第三章 Vialinin A 及び vialinin A 標的分子 USP5、TNF-α 産生との関
連性
第一節 Vialinin A の脱ユビキチン化酵素活性阻害 ... 64
第二節 USP5 ノックダウン RBL-2H3 細胞における TNF-α 放出及び
産生、転写への影響 ... 68
第三節 Vialinin A の IκB 分解阻害を介した NF-κB シグナルへの影
響 ... 78
第四節 考察 ... 80
第五節 実験の部 ... 82
総括 ... 92
英文要約 ... 95
引用・参考文献 ... 102
謝辞 ... 107
1
序論
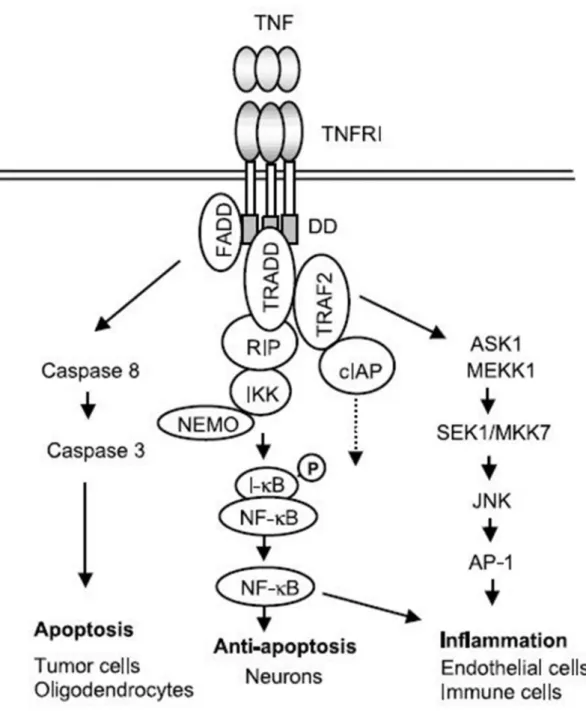
好塩基球細胞及びマスト細胞は分泌された炎症性メディエーターによるアレ ルギー反応に重要な役割を果たしている[1, 2]。高親和性 IgE 受容体 (FcεRI) に より活性化をされると、マスト細胞は最終的にアレルギー反応や炎症反応を惹 起させるヒスタミンやサイトカイン、ケモカインのようなメディエーターを放 出する[3, 4]。マスト細胞はその刺激による転写因子の活性化により産生される 炎症性サイトカインである腫瘍壊死因子 (TNF) -α 及びインターロイキン (IL) -4 などを分泌する[5, 6]。TNF-α は、細胞増殖及び分化、アポトーシスの調節を 行う炎症性サイトカインであり、他のサイトカインの産生や免疫応答を引き起 こす。そのため、無調節な TNF-α の産生は、関節リウマチやクローン病などの 炎症性疾患の発症と関連性があると言われている[7-9]。このように、活性化さ れた好塩基球細胞やマスト細胞における選択的、TNF-α を標的とした制御物質 は、関節リウマチ及びクローン病の治療薬候補として期待されている。 TNF は分子量 26 kDa の N 末端が細胞質内に存在する II 型膜結合タンパク質 として成熟化され、ゴルジ体を経由して形質膜表面に輸送された後、TNF 変換 酵素 (TACE) により Ala-Val 間が切断され 17 kDa の可溶型タンパクとなって細 胞外へと放出される[10, 11]。原形質膜上に発現する TNF には I 型 1 回膜貫通型 タンパク質である p55TNFRI と p75TNFRII の 2 種類の受容体が知られており、 TNF の多彩な作用の多くは TNFRI を介して起こされると考えられている。放出 された TNF は三量体を形成し、TNFRI に結合すると、細胞内領域に結合してい た抑制因子である silencer of death domains が活性化され、受容体の三量体化が引 き起こされる。これが引き金となり一連の細胞内シグナルが開始する[12]。まず、 TNFRI の細胞質内の death domain (DD) と呼ばれる領域に Fas-associated death domain が会合すると、caspase 8 が引き寄せられ、アポトーシスへ至る死のシグ2
ナルが起動する。さらに、DD への TNF receptor-associated death domain の会合を 介して TRAF2、RIP などのアダプター分子が結合する。TRAF2 は MAPKKK と しても機能し、JNK を活性化し c-Jun のリン酸化と転写因子 AP-1 の活性化を引 き起こす。一方、nuclear factor of κ chain gene in B cells (NF-κB) は通常その抑制 因子である IκB によりその核内移行シグナルがマスクされ細胞質に局在してい るが、RIP を介した IκB kinase (IKK) の活性化によりリン酸化された IκB はユビ キチン化を受けてプロテアソームにより分解され、NF-κB の活性化と核への移 行が引き起こされる。血管内皮細胞においてはこれらの転写因子の活性化を介 して ICAM-1 や VCAM-1 などの接着因子が発現し、血液中から好中球や好酸球 などの炎症細胞を局所に浸潤させ炎症反応を引き起こす[13, 14] (Figure 0-1)。
3
4
マスト細胞の FcεRI を介したシグナル伝達の研究は目覚ましい発展を遂げて きた[15, 16]。FcεRI は α 鎖 β 鎖と 2 本の γ 鎖の複合体からなり、α 鎖に結合した IgE に多価の抗原が結合すると IgE の架橋を介して FcεRI の凝集が生じる。する と、もともと FcεRI の β 鎖に結合していた Src ファミリーチロシンキナーゼであ る Lyn が近傍の β 鎖 γ 鎖の ITAM をリン酸化し、細胞質からさらに Lyn と Syk を呼び寄せる。Syk はホスホリパーゼ (PLC) γ、guanine nucleotide exchange factor である Vav1、SLP-76、LAT などのシグナル分子の活性化を引き起こすことが明 らかにされている。これらのアダプター分子は複合体を形成し Lipid raft と呼ば れるシグナル伝達の足場を形成し、マスト細胞の活性化に極めて重要な役割を 果たしている[17]。PLCγ は PIP2を加水分解して IP3と DG を産生し、細胞内 Ca2+ ストアからの Ca2+放出と PKC の活性化を引き起こす。Ca2+ストアの枯渇は形質 膜の CRAC チャネルを活性化し、細胞外から持続的な Ca2+流入を引き起こす。 この細胞質内 Ca2+濃度の上昇と PKC 活性化が脱顆粒とともに TNF 遊離におい ても必須であることが知られている[18, 19]。マスト細胞株であるラット好塩基 性白血病細胞 (RBL-2H3) は細胞表面に FcεRI を発現し抗原刺激により脱顆粒 を起こすことから、FcεRI を介した細胞内シグナルの研究に広く用いられてきた。 RBL-2H3 細胞を抗原または Ca2+イオノフォアのイオノマイシンで刺激すると、 脱顆粒とほぼ同じ濃度依存性をもって TNF を遊離する[20]。しかし、脱顆粒が 秒から分単位の速やかな反応であるのに対し、TNF 遊離は 1 時間後から認めら れはじめ、3-4 時間後に最大となるゆっくりした反応である。また、TNF mRNA 発現レベルは刺激後 30 分から 1 時間で最大に達し、TNF タンパクレベルは 2 時間後に最大となる。TNF mRNA は転写因子により合成が誘導されるが、3’末 端の非翻訳領域に AU-rich element を有し、転写後は速やかに分解を受ける。そ のため、細胞を刺激すると新たな転写が開始されるとともに mRNA の安定性が 高まり、その効果の総和として mRNA レベルは上昇すると考えられている。ま
5
た、この細胞株では顆粒内の TNF 貯蔵は認められず、もっぱら TNF の新たな 産生・放出の解析のためのモデルとして利用できると考えられている[21] (Figure 0-2)。
6 ユビキチン (Ub) 化は、プロテアソームでの分解をはじめとして、タンパク質 の運命や機能を様々な調節する翻訳後修飾である。しかしそれは不可逆的なも のではなく、拮抗する脱 Ub 化反応が存在する。ヒトゲノムには 100 種類以上も の脱 Ub 化酵素がコードされており、それぞれが様々な細胞内部位で固有の基質 タンパク質を脱 Ub 化することにより、Ub 化によるタンパク質の機能調節を負 に制御していることが解明されてきている。 Ub は、酵母からヒトまで高度に保存された 76 アミノ酸からなるタンパク質 であり、その C 末端カルボキシル基を介して細胞内タンパク質の Lys 残基にイ ソペプチド結合により付加される (モノ Ub 化)。Ub には 7 個の Lys 残基 (Lys6, 11, 27, 29, 33, 48, 63) が存在し、これらの Lys にさらに Ub が付加されることにより、 標的タンパク質上で Ub 鎖が伸長する (ポリ Ub 化)。細胞内には Ub のどの Lys を 介して形成したポリ Ub 化も存在し、各々が異なる機能を担うと考えられている [22]。プロテアソームでのタンパク質分解シグナルとして働くのは、Ub の Lys48 を介して連結したポリ Ub 化である[23]。Lys63 を介したポリ Ub 化は、NF-κB シグナリングや DNA 修復などのプロセスに関与し[24] 、リソソームへの膜タン パク質の選別輸送シグナルとしても働く[25, 26]。モノ Ub 化もまた、リソソーム への選別輸送シグナルとして働いている[27]。また最近、Lys11 連結型 Ub 化も プロテアソームでの分解に重要であること、N 末端の遊離のアミノ基を介して ペプチド結合 (イソペプチド結合ではない) でつながった直鎖型ポリ Ub 化が NF-κB のシグナル伝達経路で働いていることも報告され[28-30]。多様なタイプ の Ub 化が実に様々な細胞機能を担っていることが解明されてきている。脱 Ub 化酵素はプロテアーゼファミリーに属し,Ub とその標的タンパク質間、あるい は Ub 鎖中の Ub 間のイソペプチド結合を切断する加水分解酵素である。そして 近年、Ub 化による細胞機能制御における脱 Ub 化酵素の重要性が次々と解明さ れ、脚光を浴びている。
7
これまでに見つかっている脱 Ub 化酵素のアミノ酸配列から、脱 Ub 化酵素は 構造的に Ub C-terminal hydrolase (UCH) ファミリー、Ub-specific protease (USP) ファミリー、ovarian tumor protease (OUT) ファミリー、Josephin ファミリー、お よび JAMM ファミリーの 5 つのサブファミリーに分類される[31, 32]。これらの うち、JAMM ファミリー以外はすべて Cys プロテアーゼであり、それらの活性 中心は、イソペプチド結合のカルボニル炭素を求核攻撃する Cys とそれに先立 ってその Cys を脱プロトン化する His、さらに多くの場合、その His を極性化す る Asp / Asn が触媒 triad を形成している。これらの Cys、His 残基周辺のアミノ 酸配列は各サブファミリーごとに保存されており、USP ファミリーの場合は特 に Cys ボックス、His ボックスと呼ばれている[31]。一方、JAMM ファミリー は Zn2+依存性のメタロプロテアーゼであり、活性中心の近傍に配位した Zn2+が、 イソペプチド結合のカルボニル炭素を求核攻撃する水分子を極性化する[31]。触 媒ドメインのアミノ酸配列の相同性から、出芽酵母ゲノムには 19 種類、ヒトゲ ノムには 100 種類以上の脱 Ub 化酵素がコードされていると推定されている。 脱 Ub 化酵素には、これまでに 3 つの主要な働きが知られている。1 つめは、 Ub の生合成に関わる機能である。Ub は前駆体 (複数の Ub がタンデムにつなが った直鎖型ポリ Ub 鎖、あるいはリボソームタンパク質との融合タンパク質) と して翻訳され[31] 、さらに、Ub 間やリボソームタンパク質との間のペプチド結 合が脱 Ub 化酵素によって切断され、遊離の Ub が生成される。多くの脱 Ub 化 酵素がこの切断活性を有しており、Ub 前駆体のプロセシングだけを専門に行う 脱 Ub 化酵素はないと考えられている[31]。2 つめは、細胞内の遊離の Ub 量の ホメオスタシスに関する機能である。脱 Ub 化酵素には、Ub 化タンパク質がプ ロテアソームやリソソームに送り込まれる前に Ub をタンパク質から外して回 収する Ub のリサイクル機能をもつものが存在する。プロテアソームの 19S 調節 サブユニットの lid (蓋部) には JAMM ファミリーの脱 Ub 化酵素 Rpn11/POH1 が、
8
base (基部) には USP ファミリーに属する Ubp6/USP14 が含まれ、いずれも分解 されるタンパク質から Ub を切り離す[33-35]。また、酵母の USP ファミリー酵 素 Doa4 は、Ub 化されて液胞に選別輸送されるタンパク質をエンドソームで脱 Ub 化して、Ub を回収する[36]。最近、細胞内に存在する遊離のポリ Ub 鎖が遊 離 Ub のリザーバーとなっており、Doa4 は大量の Ub が必要となるストレス応答 時にこの Ub 鎖を切断することにより、遊離 Ub 量の維持に働いていることも明 らかにされている[37]。さらに Ubp14/USP5 (イソペプチダーゼ T) にも、ポリ Ub 鎖を切断して遊離 Ub を供給する役割が報告されている[38]。そして 3 つめの 機能が Ub 化によるタンパク質の調節に拮抗する働きである。脱 Ub 化酵素の数 の多さから、その大部分は様々な細胞内部位でそれぞれに固有のタンパク質を 脱 Ub 化することにより、この機能を担っていると予想されてきた。そして近年、 次々とそれらの基質タンパク質が同定され、脱 Ub 化酵素の具体的な役割が明ら かになってきている[39]。
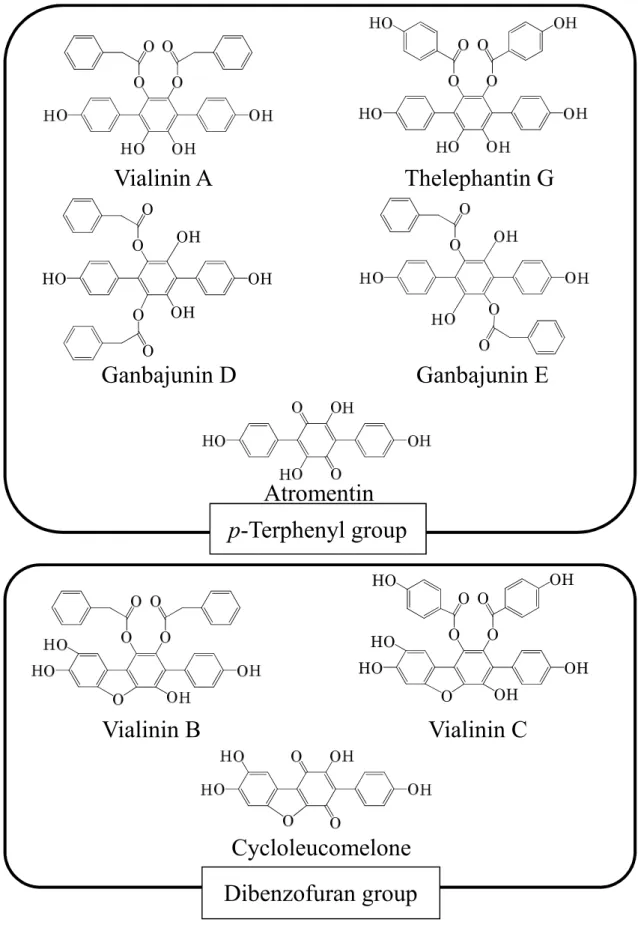
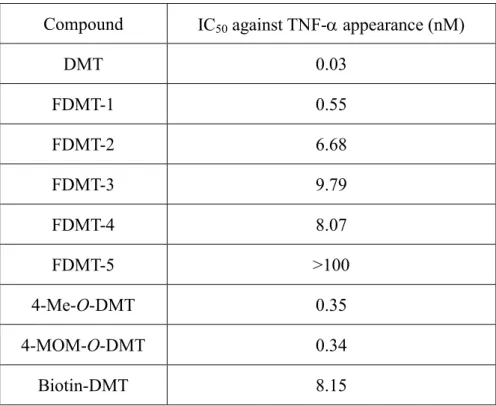
9 当研究室では、茸に含有される生理活性物質に着目し研究を行っている。中 国雲南省にある昆明食用茸研究所より有償にて購入した数種の中国産食用茸乾 燥子実体に含まれる物質を抗酸化性などのスクリーニング試験に供した。その 結果、中国産食用茸ツブイボタケ乾燥子実体より、強い抗酸化活性及び特異的 TNF-α 放出阻害活性を有する新規化合物 vialinin 類を見い出し、構造決定した[40, 41]。これまでにツブイボタケより複数の vialinin 類及びその類縁化合物を単離し ている (Figure 0-3)。これらは基本骨格の違いにより、p-terphenyl 骨格を有する ものと dibenzofuran 骨格を有するものとに分類できる。これら化合物の中で、 vialinin A (IC50: 0.09 nM) 及び vialinin B (IC50: 0.02 nM)、vialinin C (IC50: 894 nM)、
thelephantin G (IC50: 54.1 nM)のみが強力な TNF-α 放出阻害活性を示した。さらに
構造活性相関研究により、p-terphenyl 化合物の p-2’,3’,4,4’’-terphenyl- tetraol の構 造が活性発現に必須である pharmacophore であることが明らかとなり、vialinin A の側鎖部分である 2 つの phenyl acetate をより安定な methyl 基に変更した化合物 5’,6’-Dimethyl-1,1’:4’,1’’-terphenyl-2’,3’,4,4’’-tetraol (DMT, IC50: 0.02 nM) が vialinin A よりも強い TNF-α 放出阻害活性を有することが示された[42, 43]。 これらの結果より、活性アナログ DMT をリード化合物として標的分子探索ツ ールを調製し、vialinin A の RBL-2H3 細胞内標的分子の探索を行い、標的分子と vialinin A の関連性及び標的分子と TNF-α 産生の関連性について詳細に検討する ことで、vialinin A の RBL-2H3 細胞における TNF-α 放出阻害メカニズムを解明 することを目的とし研究を行った。
10
Vialinin A
Thelephantin G
Ganbajunin D
Ganbajunin E
Atromentin
p-terphenyl group
Vialinin B
Vialinin C
Cycloleucomelone
Dibenzofuran group
Figure 0-3 Bioactive compounds isolated from Thelephora vialis. OH OH O HO O O O OH OH OH O O O O O HO HO HO OH
p-Terphenyl group
Dibenzofuran group
11
使用機器
・Evaporator System
Vacuum controller: DPE-1210 (EYELA) Cool man: C-560 (SIBATA)
Rotary evaporator: N-1000 (EYELA) Digital water bath: SB-1000 (EYELA) ・DAD System
Pump: L-2130 HITACHI ELITE LaChrom Detector: L-2455 HITACHI ELITE LaChrom Column: Capcell pak C18UG120 (Shiseido)
Column: Capcell pak C18UG120 (15 mmF×250 mm) Column: Capcell pak C18UG120 (4.6 mmF×150 mm) ・HPLC System
Pump : PU-2080 (Jasco) Detector : UV-2070 (Jasco) Column oven : CO-2065 (Jasco) Integrator : choromatocorder (Sic) ・2-Dimention electrophoresis system
Isoelectric focusing electrophoresis system: IPG-IEF Type-PX (anatech) Power supply: Powerphorestar Pro 3900 (anatech)
SDS-PAGE system: Dual-270K (anatech)
・MALDI-TOFMS system: AXIMA Performance (shimadzu) ・Thermo mixer: M・BR-022UP (Taitec)
12
・Thermo mixer: Thermo mixer comfort (eppendorf) ・Centrifugal evaporator system
Centrifugal evaporator: CVE-2000 (EYELA) UNI TRAP: UT-1000 (EYELA)
Pump: DTC-22 (ULVAC)
・Spectrophotometer: U-0080D (HITACHI) ・Spectrophotometer: U-1500 (HITACHI) ・Spectrophotometer:
・Rotator: RT-50 (Taitec) ・Shaker: Shake-LR (Taitec)
・Centrifuge: himac CT15RE (HITACHI) ・Centrifuge: Avanti 30 (BECKMAN) ・Centrifuge: LC-131 (TOMY)
・Western blotting system: TE 77 PWR (GE Healthcare) ・Fluorescence microscope: OLYMPUS IX70
・MICRO PLATE READER: MTP-32 (CORONA ELECTRIC) ・CO2 INCUBATOR: MCO-96 (SANYO)
・Image analysis system: Image Quant 400 (GE Healthcare) ・Image analysis system: FluoroPhorestar 3000 (Anatech) ・SDS-PAGE system: PAGERUN AE-6531P/M (ATTO)
13
使用試薬
Acetone ... 国産化学株式会社 Acetonitrile ... 国産化学株式会社 Acrylamide ... 和光純薬工業株式会社 Agarose (DNA/RNA electrophoresis grade) ... フナコシ株式会社 2-Amino-2-hydroxymethyl-1,3-propanediol ... 和光純薬工業株式会社 Ammonium bicarbonate ... 和光純薬工業株式会社 Ammonium persulfate ... 和光純薬工業株式会社 Bradford reagent ... SIGMA Bromophenol blue (BPB) ... 和光純薬工業株式会社 Boric acid ... 和光純薬工業株式会社 Casein ... 和光純薬工業株式会社 Citric acid ... 和光純薬工業株式会社 α-Cyano-4-hydroxycinnamic acid ... SIGMA 3,3’-diaminobenzideine tetrahydrochloride (DAB) ... 和光純薬工業株式会社 Dimethyl Sulfoxide (DMSO) ... 和光純薬工業株式会社 1,1-Diphenyl-2-picrylhydrazyl (DPPH) ... 和光純薬工業株式会社 Dinitrophenyl-bovine serum albumin (DNP-BSA) ... LSA Dithiothreitol (DTT) ... 和光純薬工業株式会社 Dulbecco’s Modified Eagle Medium (DMEM) ... Nissui Ethanol ... 和光純薬工業株式会社 Ethidium Bromide Solution ... Bio-Rad Ethyl acetate ... 和光純薬工業株式会社
14
Ethylenediamine-N,N,N’,N’-tetraaacetic acid (EDTA) ... 株式会社同人科学研究所 Fetal Bovine Serum (FBS) ... Gibco Gelatin ... 和光純薬工業株式会社 anti-Glyceraldehyde 3-phosphate dehydrogenase (GAPDH)... Santa cruz Glycine ... 和光純薬工業株式会社 Glycerol ... 和光純薬工業株式会社 Goat anti-mouse IgG-HRP ... Santa Cruz Goat anti-rabbit IgG-HRP ... Santa Cruz Guanidine hydrochloride ... 和光純薬工業株式会社 4-(2-Hydroxyethyl)-1-piperazineethanesulfonic acid (HEPES) ... ... 株式会社同人科学研究所 Hybond ECL membrane ... GE Healthcare Hydrochloric acid ... 和光純薬工業株式会社 anti-IκB-α ... 和光純薬工業株式会社 Instantblue ... Expedeon K48-linked tetra-Ub ... Boston Biochem Kanamycin sulfate ... Meiji Seika LDH assay kit ... Promega Lipofectamine RNAiMAX ... Invitrogen Methanol ... 和光純薬工業株式会社 2-Mercaptoethanol ... 和光純薬工業株式会社 3MM CHR ... Whatman Monoclonal anti DNP- Immunoglobulin E (IgE) ... SIGMA p-Nitrophenyl-N-acethyl-β-D-glucosaminide ... 和光純薬工業株式会社
15
Opti-MEM ... Gibco Phenylmethysulfonyl fluoride (PdMSF) ... SIGMA Phosphate Buffered Saline 50 tablets (PBS) ... SIGMA Potassium chloride ... 関東化学株式会社 Potassium dihydrogen phosphate ... 和光純薬工業株式会社 ProteoMass Brdykinin Fragment 1-7 MALDI-MS Standard ... SIGMA ProteoMass P14R MALDI-MS Standard ... SIGMA
Primer ... Invitrogen Rainbow Molecular Weight Marker (High-Range) ... GE Healthcare Rainbow Molecular Weight Marker (Low-Range) ... GE Healthcare Rainbow Molecular Weight Marker (Full-Range) ... GE Healthcare Rat TNF-α Quantkine ELISA Kit ... R&D systems Rat IL-4 Quantkine ELISA Kit ... R&D systems RNase free water ... Qiagen RNeasy Plus Mini Kit ... Qiagen siRNA ... Invitrogen Streptavidin sepharose high performance... GE Healthcare Sodium dodecyl sulfate ... 和光純薬工業株式会社 Sodium hydrogen carbonate ... 和光純薬工業株式会社 SYBR Green PCR Master Mix ... Applied Biosystems Tacrolimus ... Fermetek TaqMan Reverse Transcription Reagents ... Invitrogen ... Tetramethylethylenediamine (TEMED) ... 和光純薬工業株式会社 TLC Silica gel 60 F254 ... Merck
16
Trypsine ... Promega TRITON X-100 ... SIGMA Ubiquitin ... Boston Biochem Ub-AMC ... Boston Biochem UCH-L1 ... Life Sensor UCH-L3 ... Life Sensor USP2 core ... Life Sensor USP4 ... Life Sensor anti-USP4 ... Santa Cruz anti-USP5 ... Abgent USP8 ... Boston Biochem anti-USP13 ... Proteintec
17
第一章 Vialinin A の TNF-α 発現阻害活性の様式の検討
第一節 Vialinin A 及び DMT の TNF-α mRNA 発現への影響
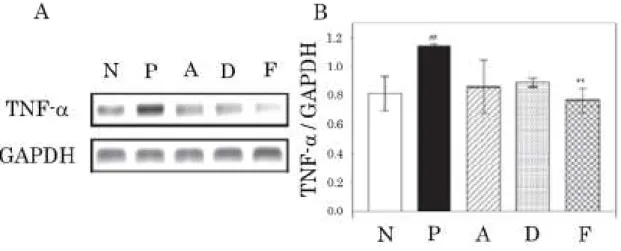
中国産食用茸ツブイボタケ (Thelephora vialis) 乾燥子実体より単離構造決定 した vialinin A とその活性アナログ DMT が RBL-2H3 細胞において強力に TNF-α 放出を阻害することから、その阻害メカニズム解明のため、vialinin A 及び DMT の TNF-α mRNA 発現に対する影響について検討した。ポジティブコントロール として用いたタクロリムス (F) は、細胞質内に存在する FK506 結合タンパク質 12 と複合体を形成し、カルシニューリン活性を阻害することにより、NF-AT 脱 リン酸化反応を阻害することで、TNF-α に代表される種々のサイトカインの発 現を阻害することが明らかとなっている。そのため、タクロリムスは抗体を介 した抗原刺激 (P) と比較し、TNF-α mRNA 発現の有意な減少を示した。 Vialinin A (A) 及び DMT (D) はタクロリムスと同様に、細胞外への TNF-α の放出は強力 に抑制するにも関わらず、TNF-α mRNA 発現に対して強力に抑制することはな かった。(Figure1-1)18
N: Non-stimulated, P: stimulated (control),
A: Vialinin A-treated, D: DMT-treated, F: Tacrolimus-treated
Figure 1-1 Inhibitory effects of vialinin A and DMT against TNF-α mRNA expression in RBL-2H3 cells.
19
第二節 Vialinin A 及び DMT の TNF-α 産生量と放出量
Vialinin A の TNF-α における阻害様式の検討を行うため、抗体を介する抗原刺 激 RBL-2H3 細胞において、様々な濃度の vialinin A 及び DMT の TNF-α 産生及 び放出に与える影響について検討した。ここでの放出量は細胞外に放出された TNF-α 量を、産生量は細胞外に放出された TNF-α 量と細胞内に残存する TNF-α 量の和で示した。対照として用いたタクロリムスは、報告されている TNF-α の 産生過程を抑制する作用メカニズムから予想されるように、産生量も放出量も 濃度依存的な抑制を示した。それに対し、Vialinin A と DMT は、放出量は濃度 依存的に抑制するが、産生量については低濃度 (0.001 nM) において強力に抑制 するものの、濃度依存性を持たないという特徴的な阻害様式を持つことが観察 された。この結果は、vialinin A 及び DMT による TNF-α 放出阻害活性機構は、 タクロリムスとは異なり、産生過程と放出過程の複数の阻害点を持つ、新規な 阻害様式であることが示唆された (Figure1-2)。20
Figure 1-2 Effects of vialinin A and DMT on TNF-α production and release in/from RBL-2H3 cells.
21
第三節 考察
中国産食用茸ツブイボタケ乾燥子実体より p-terphenyl 骨格を有する複数の化 合物を単離、構造決定した。その中でも vialinin A 及び thelephantin G が強力な TNF-α 放出阻害活性を示した。このことから、合成サンプルを含めた構造活性 相関研究により、p-terphenyl-2’,3’,4,4’’-tetraol の構造が TNF-α 放出阻害活性にお いて vialinin A の pharmacophore であることが明らかとなった。Vialinin A の全合 成研究において、その中間体として合成された活性アナログ DMT が vialinin A よりも強い TNF-α 放出阻害活性を示すことが明らかとなったことから、vialinin A と活性アナログ DMT の TNF-α 放出阻害活性パターンを比較することとした。 まず初めに、TNF-α mRNA 発現に与える影響について検討を行った。この実験 では、臨床で免疫抑制剤として使用されているタクロリムスを陽性対照として 用いた。タクロリムスは、細胞に透過した後、FK506BP12 と複合体を形成し、 カルシニューリンの活性化を阻害することで、種々のサイトカインの産生及び 脱顆粒、脂質メディエーターの産生を阻害する物質である。Vialinin A と DMT は、TNF-α mRNA 発現の阻害は観察されたものの、陽性対照のタクロリムスほ どは阻害しないことが示された。さらに、vialinin A と DMT が TNF-α の産生量 と放出量それぞれにに与える影響について検討を行った。放出量は、細胞外に 放出された TNF-α 量を ELISA にて定量し、産生量は、細胞外に放出された量と 細胞内に残存する量の和で示した。タクロリムスは TNF-α の産生段階を阻害す ることの報告と矛盾なく、産生量と放出量共に濃度依存的に阻害されることが 観察された。それに対し、vialinin A 及び DMT は、放出量は濃度依存的に阻害 されるが、産生量は低濃度で強力に阻害されるものの、濃度依存性をもたない 同様の阻害パターンを示すことが観察された。このことから、vialinin A 及び22
DMT が TNF-α の産生段階と放出段階の複数点において阻害を発現していること が示唆された。また、既存の阻害様式であるタクロリムスとは別の新規の阻害 様式を持つことが明らかとなった。
23
第四節 実験の部
4-1. RBL-2H3 細胞 化学発がんにより生じたラットがん化好塩基球をセルライン化し、抗原・抗 体反応及びカルシウムイオノフォアなどの刺激により脱顆粒を引き起こしやす い性質を有する細胞で肥満細胞株の一種である RBL-2H3 細胞を用いた。肥満細 胞の研究にはラットやマウスの細胞がよく用いられており、その存在部位によ り結合組織 connective tissue mast cell (CTMC) 型と粘膜肥満細胞 mucosal mast cell (MMC) の 2 型に分けられ、RBL-2H3 細胞は CTMC 型である。4-1-1. 10% FBS-DMEM 培養液の調製
Dulbecco’s Modified Eagle Medium (Nissui) 10 g 7.5% NaHCO3 (Wako) 20 mL
250 mg/mL Kanamycin (Meiji Seika) 0.4 mL Fatal Bovine Serum (Gibco) 100 mL Total (Distilled Water) 1000 mL
4-1-2. RBL-2H3 細胞の継代培養 細胞は 37°C、5% CO2存在下の CO2インキュベーター内で、10% FBS-DMEM 5 mL を含む T-25 cm2フラスコを用いて培養した。コンフルエントで 5×105 cells/mL の細胞濃度に調整し、細胞の継代は 3 日に 1 回行った。継代の際、培養上清を 取り除き、新鮮な培養液を加えた後、cell scraper で細胞をはがし、2×104 cells/mL になるように 10% FBS-DMEM 培養液で希釈して新しいフラスコに細胞を植え 継ぎ、CO2インキュベーター内で培養を行った。
24 4-1-3. RBL-2H3 細胞の融解 液体窒素中に保存しておいた細胞の入ったチューブを 37°C で融けきる直前ま で融解し、10% FBS-DMEM を 8 mL と細胞融解液 2 mL を 15 mL 遠沈管にて混 合し、1200 rpm, 4°C, 3 min で遠心分離を行った。上清除去後、新たに 10% FBS-DMEM を加え、T-25 cm2フラスコに移し、37°C にて CO2インキュベーター 内で培養を開始した。 4-1-4. 細胞の凍結 T-25 cm2フラスコ中の培養液を除き、新たに 10% FBS-DMEM を 5 mL ずつ加
え、細胞を cell scraper ではがし、15 mL 遠沈管に移した。1200 rpm, 4°C, 3 min で遠心分離を行い、上清を除去後、80%FBS-DMSO を培養液と等量 (2 mL) ずつ 混合し、あらかじめ冷やしておいた凍結用チューブに 2 mL ずつ分注し、-80°C で凍結した後、液体窒素中で凍結保存した。
25
4-2. リアルタイム PCR による TNF-α mRNA 発現の解析サンプルの調製
T-25 cm2フラスコで RBL-2H3 細胞を培養し、Monoclonal anti-DNP IgE 抗体を添 加し、16 時間感作させた。2% FBS-DMEM で 2 回洗浄後、100 nM vialinin A 及 び 100 nM DMT、100 nM タクロリムスを添加し、15 分間処理した。その後、 DNP-BSA を抗原として添加し、1 時間刺激した。細胞上清を分取し、細胞を TNF-α RNA 発現解析サンプルとした (scheme1-1)。分取した細胞上清中の TNF-α 量を ELISA 法を用いて定量することで、化合物の処理が適正に行われていることを 確認した。
RBL-2H3 cells (2.0×105 cells/flask) in T-25 cm2 flask. ←Monoclonal anti-DNP IgE
Incubated for 16 h at 37°C
Washed twice with 2% FBS-DMEM
←100 nM vialinin A or 100 nM DMT or 100 nM tacrolimus Incubated for 15 min at 37°C
←DNP-BSA
Incubated for 1 h at 37°C Cell sample
26
4-3. Vialinin A 及び DMT 処理 RBL-2H3 細胞からの RNA 抽出
RBL-2H3 細胞に Monoclonal anti-DNP IgE 抗体を 16 時間感作させ、その後に 100 nM vialinin A 及び 100 nM DMT、100 nM タクロリムスで処理し、15 分後に 抗原として DNP-BSA を加えた。抗原刺激 30 分後、細胞を RNA protect cell reagent (QIAGEN) を用いて固定し、RNeasy Plus kit (QIAGEN) を用い、添付のプロトコ ールに従って、RNA の抽出を行った(scheme1-2)。RNA protect cell reagent を 10000 rpm, 5 min の遠心分離で除去し、Buffer RLT を加え細胞を完全に破砕後、溶解物 を QIA shredder spin column にて 14000 rpm, 2 min 遠心分離し、均一化した。それ を gDNA Eliminator spin column にて 10000 rpm, 30 s 遠心分離した。その産物に 70%エタノールを加え混合し、RNeasy spin column にて 10000 rpm, 15 s 遠心分離 した。さらに、Buffer RW1 及び Buffer RPE で 10000 rpm, 15 s 遠心分離を行うこ とで精製し、RNase-free water で 10000 rpm, 1 min 遠心分離を行うことで RNA を 溶出した。精製した RNA は、U-0080 spectrophotometer を用いて、OD260 nm/OD280 nm比から純度を算出し、OD260 nm値から RNA 濃度を求めた。精製した RNA は
27 Cell sample
←RNA protect cell reagent
Resuspended and centrifuged for 5 min, 10,000 rpm ←Buffer RLT (lysis buffer)
Applied to QIAshredder spin column Centrifuged for 2 min, 14,000 rpm ←equal volume of 70%ethanol Applied to RNeasy spin column Centrifuged for 15 sec, 10,000 rpm ←Buffer RW1 (wash)
Centrifuged for 15 sec, 10,000 rpm ←Buffer RPE (wash)
Centrifuged for 15 sec, 10,000 rpm ←Buffer RPE (wash)
Centrifuged for 2 min, 10,000 rpm Centrifuged for 1 min, 10,000 rpm (dry) ←RNase free water
Incubated for 1 min Applied to collection tube
Remove tocentrifuge for 10,000 rpm, 1 min RNA extraction sample
28 4-4. Total RNA の逆転写
RBL-2H3 細胞より精製した RNA は RNase-free water で希釈し、逆転写に用い た。逆転写反応は、TaqMan Reverse Transcription Reagent (Invitrogen) を用い、添 付のプロトコールに従って、以下のように行った(scheme 1-3)。RT 反応液*1
18 µL に精製 total RNA 溶液 2 µL を加え、反応試料溶液とした。RT 反応は、15 min, 37°C で逆転写を行い、85°C, 5 s で reverse transcriptase を失活させた。 *1 RT 反応液 TaqMan RT Buffer 2.0 µL 5.5 mM MgCl2 4.4 µL 500 µM dNTP mix 5.0 µL 2.5 µM random hexamer 1.0 µL 0.4 Unit/µL RNase inhibitor 0.4 µL 1.25 Unit/µL reverse transcriptase 0.5 µL
29 4-5. リアルタイム PCR 法による測定
リアルタイム PCR は検出方法に SYBR Green PCR Master Mix を用いたインタ ー カ レ ー タ ー 法 を 採 用 し 、 Power SYBR Green PCR Master Mix (Applied Biosystems) を用い、添付のプロトコールに従って、以下のように行った(scheme 1-3)。PCR 反応液*218 µL に逆転写により得られた cDNA 2 µL を加えて、反応試 料溶液とした。PCR 反応は、94°C, 10 s 前処理した後、denature、annealing、extension を 1 サイクルとして増幅反応を行い、その後 72°C, 5 min 保持した。PCR 後に融 解曲線分析を行い、Tm 値を確認した。使用したプライマーはそれぞれ、TNF-α forward primer: 5’-ACT CCC AGA AAA GCA AGC AA-3’, TNF-α reverse primer: 5’-CGA GCA GGA ATG AGA AGA GG-3’, glyceraldehyde 3-phosphate dehydrogenase (GAPDH) forward primer: 5’-GGC ACA GTC AAG GCT AGA ATG G-3’ GAPDH reverse primer: 5’-GGA GTC AGT TCC ACA GGCTG-3’である。温度 及び時間 (denature, annealing, extension)、サイクル数は、TNF-α: 94°C for 25 s, 55°C for 40 s, 72°C for 35 s (39 cycles); GAPDH: 94°C° for 10 s, 62°C for 20 s, 72°C for 35 s (39 cycles) である。蛍光強度より定量を行い、さらに PCR 産物をエチジ ウムブロマイドを含む 2%アガロースゲル電気泳動により分画し、UV 照射下で、 バンドを IQuant Capture 400 system (GE Healthcare) にて定量を行い、確認した。 GAPDH はサンプルの標準化に用いて、相対値として比較を行った。
30 *2 PCR 反応液
Power SYBR Green PCR Master Mix 10 µL 20 µM forward primer 0.5 µL 20µM reverse primer 0.5 µL RNase-free water 7 µL
Total 18 µL
RNA extraction sample
←RT reaction buffer
Incubated for 15 min at 37°C Incubated for 5 sec at 85°C cDNA sample
←PCR reaction buffer Incubated for PCR condition
Denature Annealing Extension
Incubated for 5 min, 75°C Analysed for melting curve PCR products
31 4-6. TNF-α 放出量及び産生量の測定 4-6-1. TNF-α 発現阻害活性試験サンプル調製 3.0×105 cells/mL になるように 10%FBS-DMEM で調製した RBL-2H3 細胞を 1 mL ずつ 24 穴プレートに分注した。さらに各 well に 30 ng/mL になるように調製 した Monoclonal anti DNP IgE 抗体を加え、37°C、16 時間感作した。2% FBS-DMEM に溶解した vialinin A 及び DMT、tacrolimus を加え、15 分間前培養 した。その後、抗原として DNP-BSA を 10 µg/mL となるように加え、37℃で CO2
インキュベーター内にて 3 時間培養した。その後、培養上清を回収し、それを 細胞培養上清サンプルとした。さらに、細胞を 0.2% tritonX-100-PBS で溶解し、 細胞内残存サンプルとした。
4-6-2. TNF-α 放出量及び産生量の定量
RBL-2H3 細胞に発現した TNF-α の定量には、rat TNF-α Quantikine ELISA kit (R&D systems) を用い、添付のプロトコールに従い、測定を行った。あらかじめ TNF-α 抗体が塗布してある 96 穴マイクロプレートに assay buffer 50 µL と測定す るサンプル 50 µL を入れ、室温で 2 時間静置した。Wash buffer で 5 回各 well を 洗浄し、HRP-conjugated anti-TNF-α を 100 µL 加え、室温で 2 時間静置した。そ の後、発色基質を 100µL 加え、30 分後に反応停止薬として stop solution 100 µL を加えた。反応停止後、OD450 nmを測定し、付属されているスタンダードを用い て定量した。TNF-α 放出量は、細胞培養上清サンプル中に含有する TNF-α を示 し、産生量は、細胞培養上清サンプル中に含まれる TNF-α 量と細胞内残存サン プルに含有される TNF-α 量の和で示した。
32
第二章 Vialinin A 標的分子の探索
第一節 探索プローブの調製
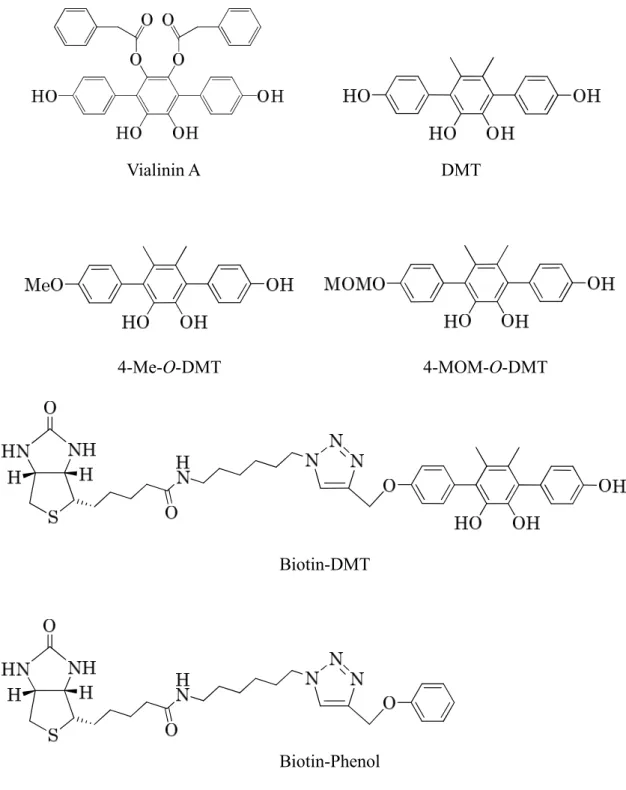
1-1. Vialinin A の全合成
Vialinin A は、sesamol を出発原料に、11 steps、全収率 28%で全合成を達成し た。鍵反応として、電子リッチなアリルトリフレートとフェニルボロン酸のダ ブル鈴木―宮浦カップリングと bis-MOM ether の酸化的脱保護を用いた (Figure 2-3)。 1-2. Vialinin A 活性アナログ DMT の調製 DMT は、Vialinin A のファーマコフォアを有する誘導体として合成され、その TNF-α の放出阻害活性は vialinin A よりも強力な IC50値 0.02 nM を与えた。この 合成は、vialinin A と同様、ダブル鈴木―宮浦カップリングを鍵反応とし、商業 的に利用可能な 3,4-dimethylphenol を出発原料としてわずか 5 steps で達成される ことから、vialinin A 標的分子探索ツールのリード化合物とした (Figure 2-3)。
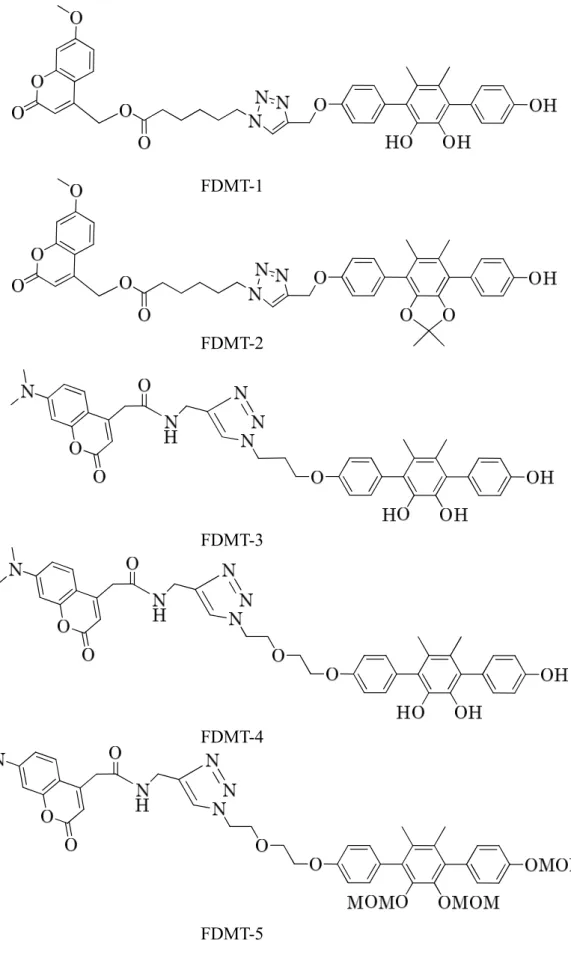
33 1-3. Biotin 標識化プローブの調製 Vialinin A 標的分子の探索研究のため、そのツールとして使用する Biotin 標識 化プローブ並びにクマリニル基を蛍光発色団とする蛍光標識化プローブの合成 を試みた。4 位での誘導体化による活性への影響を検討するため、vialinin A 活 性アナログ DMT の 4 位のヒドロキシル基をメトキシ基に置換した 4-Me-O-DMT とメトキシメチルに置換した 4-MOM-O-DMT を合成した。一方、DMT の 4 位の ヒドロキシル基部分にクリック反応を用いて 1,2,3-トリアゾール環を含むリン カーを介して Biotin 標識化プローブ Biotin-DMT を調製した。その対照として、 DMT 部分にフェニル基を置換した Biotin-Phenol を調製した (Figure 2-3)。 1-4. 蛍光標識化プローブの合成 蛍光標識化プローブ (FDMTs) もビオチン標識化プローブと同様、アルキル基 またはエチレングリコールを導入し、リンカーの形状の異なるプローブを複数 調製した。FDMT-1 と FDMT-2 はリンカーをエステル結合で、FDMT-3 及び-4、 -5 は、アミド結合で結合させた。FDMT-2 はカテコール部分をマスクすることで、 FDMT-1 の対照として調製した。FDMT-4 は、FDMT-3 に対し、より直線性の高 いリンカーを導入た。FDMT-5 は DMT のヒドロキシル基を MOM 基で保護する ことにより、FDMT-4 の対照とした (Figure 2-4)。
34
第二節 探索プローブの TNF-α 放出阻害活性試験
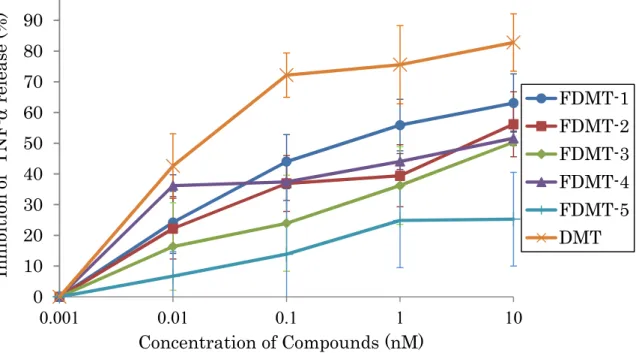
Vialinin A のファーマコフォアを有する DMT の TNF-α 放出阻害活性は、vialinin A よりも強い IC50値 0.02 nM を示した。さらに、vialinin A と DMT の TNF-α 産 生と放出の阻害パターンが同じ傾向を示したことから、同様な阻害メカニズム を有することが予想されることにより、DMT をリード化合物として探索プロー ブを調製することした。DMT の 4 位のヒドロキシル基をメトキシ基に置換した 4-Me-O-DMT とメトキシメチル基に置換した 4-MOM-O-DMT は TNF-α 放出阻害 活性の IC50値としてそれぞれ 0.35 nM 及び 0.34 nM を与えることから、活性への 影響は少ないものと判断し、4 位にリンカーを結合させることとした。蛍光標識 プローブ 5 種とビオチン標識化プローブについても検討を行った。エステル結 合で調製した FDMT-1 は IC50値 0.55 nM を、FDMT-2 は IC50値 6.68 nM を与えた。 アミド結合で調製した FDMT-3 は IC50値 9.79 nM、より直線性の高いリンカーを 導入した FDMT-4 は 8.07 nM、そして FDMT-5 は>100 nM を示した。Biotin-DMT は IC50値 8.15 nM を与えた (Figure 2-1 and 2-2, Table 2-1)。35
Figure 2-1 Inhibitory effects of DMT and FDMTs against TNF-α release from RBL-2H3 cells. 0 10 20 30 40 50 60 70 80 90 100 0.001 0.01 0.1 1 10 FDMT-1 FDMT-2 FDMT-3 FDMT-4 FDMT-5 DMT Concentration of Compounds (nM) In h ib it ion of T N F -α rel ea se (% )
36
Figure 2-2 Inhibitory effects of vialinin A, DMT, Biotin-DMT, 4-Me-O-DMT and 4-MOM-O-DMT against TNF-α release from RBL-2H3 cells.
0% 10% 20% 30% 40% 50% 60% 70% 80% 90% 100% 0.001 0.01 0.1 1 10 In h ib it io n of T N F -α R el w a se Concentration of Compounds (nM) Vialinin A BDMT 4-Me-O-DMT 4-MOM-O-DMT DMT
37
Table 2-1 IC50 values of DMT derivatives against TNF-α release from RBL-2H3 cells.
Compound IC50 against TNF-α appearance (nM)
DMT 0.03 FDMT-1 0.55 FDMT-2 6.68 FDMT-3 9.79 FDMT-4 8.07 FDMT-5 >100 4-Me-O-DMT 0.35 4-MOM-O-DMT 0.34 Biotin-DMT 8.15
38
Vialinin A DMT
4-Me-O-DMT 4-MOM-O-DMT
Biotin-DMT
Biotin-Phenol
39 FDMT-1 FDMT-2 FDMT-3 FDMT-4 FDMT-5
40
第三節 DMT の RBL-2H3 細胞内での局在性の検討
DMT の RBL-2H3 細胞内での挙動を検討するため、蛍光標識化プローブ FDMTs を用いて検討を行った。エステル結合によりクマリニル基を結合している FDMT-1 並びに-2 は、薄層クロマトグラフィーによる検討から細胞培養液中で加 水分解され、蛍光発色団が失われることが確認された。これに対し、アミド結 合により蛍光発色団を結合している FDMT-3 及び-4、-5 は培養液中で分解され ることなく安定であることが確認された。蛍光顕微鏡により観察を行ったとこ ろ、リンカー部分にエチレングリコールを導入した FDMT-4 及び-5 は細胞内へ の十分な取り込みが確認されたのに対し、アルキル鎖のリンカーを有する FDMT-3 は取り込み量が極端に少ないことが示された (Figure 2-5)。以上の結果 より、細胞内への取り込みがリンカー部分の構造により影響することが示唆さ れた。さらに、細胞内分子との結合について検討を行った。細胞を FDMT-3 及 び-4、-5 で処理した後、抽出したタンパク質を電気泳動に供したもの (A) と、 細胞をライセートとした後に FDMT-3 及び-4、-5 で処理後、電気泳動に供した もの (B) とを比較した。SDS-PAGE に供したゲルを 360 nm で励起させて蛍光を 発するバンドを検出したところ、(A) では FDMT-4 で処理したレーンにのみ複数 のバンドが検出されたのに対し、(B) では、FDMT-3 及び-4 で処理したものに同 様な蛍光を示す複数のバンドが検出された。FDMT-5 で処理したものには、蛍光 を示すバンドは検出されなかった (Figure 2-6)。41 F igur e 2-5 L oc al iz at ion of F D M T s i n R B L -2H 3 c el ls .
42 F igur e 2-6 F ra ct iona ti on of F D M T s-b indi ng i nt ra ce ll ul ar m ol ec ul es i n R B L -2H 3 c el ls w it h S D S -P A G E .
43
第四節 Biotin-DMT を用いた vialinin A 標的分子の探索
Vialinin A 及び DMT の TNF-α 阻害メカニズムは、臨床で免疫抑制剤として使 用されているタクロリムスとは異なり、TNF-α の産生段階と放出段階をそれぞ れ阻害していることが示唆された。そこで、活性アナログ DMT をリード化合物 とするビオチン標識化プローブ biotin-DMT を調製し、RBL-2H3 細胞における vialinin A 標的分子の探索を行った。対照としてフェニル基にビオチン標識した プローブ biotin-phenol を用いた。抗体を介して抗原刺激した RBL-2H3 細胞の細 胞質画分にそれぞれのプローブを処理し、ストレプトアビジンカラムに供する ことで vialinin A 標的分子の精製を試みた結果、Biotin-DMT を処理したサンプル の 約 100 kDa 付 近 に バ ン ド が 検 出 さ れ た (Figure 2-7) 。 こ の 分 子 を MALDI-TOFMS に供し、peptide mass fingerprint (PMF) 法にて解析したところ、 脱ユビキチン化酵素の 1 つである ubiquitin-specific peptidase 5 (USP5) と一致し た (Figure 2-7, 2-8, 2-9)。対照として用いた biotin-phenol を処理したサンプルから は USP5 が検出出来なかったこと、さらに DMT との競合阻害実験にて標的分子 USP5 の検出量が減少したことなどから、USP5 は biotin-DMT の DMT 部分に結 合することが示された (Figure 2-10)。44
Figure 2-7 Isolation of a vialinin A-binding molecule from RBL-2H3 cells using a ligand-affinity probe.
45
Figure 2-8 Peptide fragments peaks of a target molecule of vialinin A in MALDI-TOFMS.
Figure 2-9 Protein score of peptide fragments of a vialinin A-targeting molecule by PMF analysis.
46
Table 2-1 Peptide mass of the vialinin A-binding protein for PMF analysis.
Start-End Mr (expt.) Mr (calc.) Delta Miss Sequence 122-133 1413.8527 1413.8231 0.0296 0 K.IVILPDYLEIAR.D 147-162 1575.8027 1575.8104 -0.0077 0 R.VTSAVEALLSADSASR.K 163-174 1443.7227 1443.7106 0.0121 1 R.KQEVQAWDGEVR.Q 206-221 1873.9427 1873.9356 0.0071 0 R.ENLWLNLTDGSILCGR.R 222-238 1977.9027 1977.8830 0.0197 1 R.RYFDGSGGNNHAVEHYR.E 223-238 1821.7427 1821.7819 -0.0392 0 R.YFDGSGGNNHAVEHYR.E 292-303 1479.7327 1479.6697 0.0630 0 K.TMTELEIDMNQR.I 361-379 2107.0227 2107.0222 0.0005 0 K.IFQNAPTDPTQDFSTQVAK.L 380-406 2824.4327 2824.3879 0.0448 0 K.LGHGLLSGEYSKPALESGDG EQVPEQK.E 511-524 1609.8327 1609.8675 -0.0348 1 R.QAEEEKVALPELVR.A 562-574 1539.8427 1539.8337 0.0090 0 R.FASFPDYLVIQIK.K 576-585 1208.7027 1208.6230 0.0797 0 K.FTFGLDWVPK.K 576-586 1336.7027 1336.7180 -0.0152 1 K.FTFGLDWVPKK.L 586-603 2114.0327 2114.0929 -0.0602 1 K. KLDVSIEMPEELDISQLR.G 829-836 1064.5427 1064.5291 0.0136 0 R.WVIYNDQK.V 837-855 2347.1127 2347.1671 -0.0543 1 K.VCASEKPPKDLGYIYFYQR.V
47
Figure 2-10 Competition assay assessing recombinant hUSP5 binding of vialinin A, DMT or Ub to a biotin-DMT probe.
48
第五節 考察
中国産食用茸ツブイボタケに含有されている vialinin A 及びその活性アナログ である DMT が強力な TNF-α 放出阻害活性を有することが明らかとなった。ま たその阻害メカニズムが、臨床で免疫抑制剤として使用されているタクロリム スとは異なり、TNF-α の産生段階及び放出段階のそれぞれを阻害していること が観察され、新規経路による阻害メカニズムの存在が示唆された。そこで、DMT をリード化合物とするクマリニル基を蛍光発色団とする FDMTs を用いて RBL-2H3 細胞内での局在性及びビオチンを標識化した biotin-DMT を用いて標的 分子の探索を検討した。それぞれのプローブデザインとして DMT の 4 位のヒド ロキシ基にリンカーを介して蛍光発色団及び biotin を結合させることとした。4 位 の ヒ ド ロ キ シ 基 を 修 飾 す る こ と に よ る 活 性 へ の 影 響 を 検 討 す る た め 、 4-Me-O-DMT 及び 4-MOM-O-DMT を調製し、TNF-α 放出阻害活性を測定した。 DMT の IC50値 0.02 nM に対し、4-Me-O-DMT 及び 4-MOM-O-DMT の IC50値は それぞれ、0.35 nM 及び 0.34 nM を示したことから、活性は保持されたものと判 断した。この結果より、4 位のヒドロキシ基を介したプローブを調製することと した。FDMT-1 及び FDMT-2、FDMT-3、FDMT-4、FDMT-5、Biotin-DMT の IC50 値はそれぞれ、0.55 nM 及び 6.68 nM、9.79 nM、8.07 nM、>100 nM、8.15 nM を 与えた。FDMT-1 は、最も強い活性を示したが、蛍光プローブを用いた細胞への 取り込み実験から、FDMT-1 及び-2 は細胞内への取り込みが確認されなかったこ とより、TLC による検討を行ったところ、培養液中での蛍光プローブの蛍光発 色団と DMT 部分とをつなぐエステル結合に乖離が認められた。従って、FDMT-1 及び-2 は完全なかたちで細胞に取り込まれないことが示唆された。FDMT-3 は、 細胞内に少量しか取り込まれないのに対し、FDMT-4 及び-5 は十分量が取り込ま49 れることが観察された。この結果を受け、細胞膜透過の有無による影響を検討 するため、細胞破砕前後の蛍光プローブ処理による細胞内分子への結合実験を 行ったところ、リンカーの構造によらず、DMT の構造が細胞内分子への結合に 必須であることが観察された。これらの結果より、プローブのリンカー構造の 違いが細胞内への取り込みに影響ないことが示されたことから、biotin-DMT を 用いた RBL-2H3 細胞内標的分子の探索は細胞のライセートを用いることとした。 Biotin-DMT を用いた標的分子の探索研究により、vialinin A の標的分子の 1 つを 脱 Ub 化酵素の 1 種である Ubiquitin-Specific Peptidase 5 (USP5) と同定した。対 照実験に用いた biotin-phenol に USP5 が結合しないこと及び、vialinin A 及び DMT、 Ub との競合試験の結果から、USP5 が vialinin A の標的分子であることが強く示 唆された。
50
第六節 実験の部
6-1. TNF-α 放出阻害活性試験 6-1-1. TNF-α 放出阻害活性試験サンプル調製 3.0×105 cells/mL になるように 10%FBS-DMEM で調製した RBL-2H3 細胞を 1 mL ずつ 24 穴プレートに分注した。さらに各 well に 30 ng/mL になるように調製 した Monoclonal anti-DNP IgE 抗体 を加 え 、37°C、16 時間 感 作した 。2% FBS-DMEM に溶解した各サンプルを加え、15 分間前培養した。その後、抗原と して DNP-BSA を 10 µg/mL となるように加え、37℃で CO2インキュベーター内にて 3 時間培養した。その後、培養上清を回収し、それを細胞培養上清サンプ ルとした。
6-1-2. TNF-α 放出量及び産生量の定量
RBL-2H3 細胞に発現した TNF-α の定量には、rat TNF-α Quantikine ELISA kit (R&D systems) を用い、添付のプロトコールに従い、測定を行った。あらかじめ TNF-α 抗体が塗布してある 96 穴マイクロプレートに assay buffer 50 µL と測定す るサンプル 50 µL を入れ、室温で 2 時間静置した。Wash buffer で 5 回各 well を 洗浄し、HRP-conjugated anti-TNF-α を 100 µL 加え、室温で 2 時間静置した。そ の後、発色基質を 100µL 加え、30 分後に反応停止薬として stop solution 100 µL
を加えた。反応停止後、OD450 nmを測定し、付属されているスタンダードを用い
51 6-2. FDMTs の RBL-2H3 細胞内の局在性の検討 6-2-1. FDMTs を用いた RBL-2H3 細胞への取り込み実験 T-25 cm2フラスコに RBL-2H3 細胞を 2.0×106 cells/mL になるように培養し、 そこに 10% FBS-DMEM で希釈した 100 µM FDMTs をそれぞれ添加し、15 分間 処理した。処理後、PBS で 2 回洗浄し、10% FBS-DMEM を PBS で置換し、蛍光 顕微鏡 OLIMPUS IX70 で観察した。FDMTs の蛍光発色団であるクマリニル基は 励起波長 360 nm、蛍光波長 420 nm を持つため、ブルーライトバンドパスフィル ターを介することにより、クマリニル基の蛍光を観察した。 PBS
PBS tablets (sigma) 1 tablets Sterilized water 200 mL Total 200 mL 6-2-2. 培養液添加時の FDMTs の状態 RBL-2H3 細胞内に蛍光発色が確認できなかった FDMT-1 及び FDMT-2、蛍光 発色の弱かった FDMT-3 について、培養液中のそれぞれの FDMTs の状態につい て検討した。FDMT-1 及び FDMT-2、FDMT-3 を処理した培養液を TLC 板に塗布 し展開した。展開された TLC 板に UV: 254 nm を照射し、現れるスポットを観察 した。
52 6-3. FDMTs による RBL-2H3 細胞内分子への結合実験 6-3-1. サンプル調製 RBL-2H3 細胞への FDMTs 結合実験を、intact な細胞に FDMTs を処理した実験 と細胞破砕し細胞内分子を抽出したものに FDMTs を処理した実験の 2 種類の方 法で検討した。 A). 破砕処理前処理 T-25 cm2フラスコに 2.0×105 cells/mL になるように調製した RBL-2H3 細胞に 30 ng/mL になるように moclonal anti-DNP IgE 抗体を加え、37°C、16 時間感作し た。次に 100 µM FDMTs をそれぞれ加え 15 分間処理し、細胞内結合分子と結合 させた。そこに抗原として DNP-BSA を加え、3 時間刺激した。そして、PBS で 2 回洗浄後、0.2% tritonX-100-PBS を添加し、細胞を破砕した。そこに等量の sample buffer を加え、サンプルとした。 B). 破砕処理後処理 T-25 cm2フラスコに 2.0×105 cells/mL になるように調製した RBL-2H3 細胞に 30 ng/mL の monoclonal anti DNP IgE 抗体を加え、37℃、16 時間感作した。そこ に抗原として DNP-BSA を加え、3 時間刺激した。そして、PBS で 2 回洗浄後、 0.2% tritonX-100-PBS を添加し、細胞を破砕した。そこに、100 µM FDMTs を添 加し、15 分間処理し、細胞内結合分子に FDMTs をそれぞれ結合させた。そこに 等量の sample buffer を加え、サンプルとした。
53 0.2% TritonX-100-PBS PBS tablets 1 tablet Sterilized water 200 mL TritonX-100 400 µL Total 200 mL Sample buffer 0.5 M Tris-HCl (pH6.8) 2.5 mL 10% SDS 4.0 mL 2-Mercaptoethanol 1.0 mL Glycerol 2.0 mL 0.05% BPB 0.2 mL Total 10 mL
54 6-3-2. ポリアクリルアミド電気泳動
Sodium dodecyl sulfate- poly acrylamide gel electrophoresis (SDS-PAGE) は、 Stacking gel (濃縮ゲル) 及び running gel (分離ゲル) の 2 層ゲルを用いて行った。 分離ゲルはアクリルアミド濃度を 12%で調製した。濃縮ゲルの各ウェルにサン プル 20 µL、rainbow marker 5 µL ずつ分注し、PAGERUN (ATTO) を用いて 120 分、20 mA にて電気泳動を行った。泳動後、ゲルを fluorophorestar 3000 (anatech) を用いて、UV で励起し、FDMTs の染色を観察した。 Stacking gel 0.5 M Tris-HCl (pH6.8) 1.5 mL 2.9% acrylamide-1.0% Bis 0.6 mL 10% SDS 0.12 mL TEMED 7 µL 10% APS 20 µL Total 5 mL
55 Running gel 0.75 M Tris-HCl (pH8.8) 10 mL 2.9% acrylamide-0.5% Bis 8.0 mL 10% SDS 0.4 mL TEMED 20 µL 10% APS 150 µL Total 20 mL Running buffer Tris 3.0 g Glycine 14.4 g 10% SDS 5.0 mL Total 500 mL
56 6-4. Vialinin A 標的分子の探索研究 6-4-1. RBL-2H3 細胞内分子の抽出 RBL-2H3 細胞の細胞質画分は、Dignam の改良法にて分画を行った。RBL-2H3 細胞を 200×g、5 分の条件で遠心分離することでペレットの状態にし培養液を 取り除いた。そこに 2 倍量の buffer A を添加し懸濁し、20 分間氷上に静置した。 そして、0.05%になるように NP-40 を添加し、ボルテックスミキサーで 10 秒間 撹拌することで細胞膜を破砕した。抽出物を 20,000×g、4°C、5 分の条件で遠心 分離を行い、可溶化画分を分取した。その後、NP-40 の濃度が 0.01%になるよう に buffer B で希釈し、buffer C で 4°C、3 時間、透析を行った。透析後のサンプ ルを 10 倍に希釈し、20,000×g、4°C、30 分の条件で遠心分離を行い、その可溶 化画分を細胞質画分とした。タンパク質濃度を、Bradford reagent (sigma) を用い て測定した (Scheme 2-1)。
57 RBL-2H3 cells
←buffer A (on ice, 20 min) ←0.05% NP-40 (vortex, 10 sec) (Centrifugation, 4°C, 20,000 x g, 5 min) Collected supernatants ←buffer B (5-fold) (Dialyze, 4°C, 3 h) ←buffer B (10-fold) (Centrifugation, 4°C, 20,000 x g, 30 min) Collected supernatants Cytoplasmic extracts
58 Buffer A 10 mM HEPES (pH7.9) 10 mM KCl 0.1 mM EDTA 1 mM DTT 0.5 mM PMSF Buffer B 20 mM HEPES (pH7.9) 100 mM KCl 0.2 mM EDTA 10%(v/v) glycerol 1 mM DTT 0.2 mM PMSF Buffer C 20 mM HEPES (pH7.9) 100 mM KCl 0.2 mM EDTA 10%(v/v) Glycerol 0.1% Triton X-100 1 mM DTT 0.2 mM PMSF
59
6-4-2. Biotin 標識化プローブを用いた RBL-2H3 細胞内標的分子の単離
RBL-2H3 細胞内標的分子の探索には、有機合成した 2 種の biotin プローブ (biotin-DMT、biotin-phenol) を用いた。2 mg/mL RBL-2H3 細胞細胞質画分にそれ ぞれ 100 µM biotin-DMT 及び 100 µM biotin-phenol、vehicle として使用した DMSO を添加し、4°C で 1 時間撹拌した。非結合分子を取り除いた後、Streptavidin sepharose resin (GE Healthcare) を添加し、4°C で 15 分間反応させた。resin を取 り除き、SDS-PAGE sample buffer にて結合分子を溶出させた。溶出液を SDS-PAGE に供し、Instant Blue 染色にて結合分子を染色した。 6-4-3. MALDI-TOFMS サンプル作成 Biotin-DMT を 用 い た ア フ ィ ニ テ ィ ー ク ロ マ ト よ り 溶 出 さ れ た 分 子 を SDS-PAGE に供し、染色されたバンドを切り出した。切り出したゲルは、既存 のプロトコールに従い、脱色、還元、アルキル化、脱水、膨潤、乾燥を行った。 その後、トリプシン消化を行い、アセトニトリルでペプチドを抽出後、濃縮し た (Scheme 2-2)。
60 Clipped gel
←50% methanol (65°C, 15 min, 4 times; decoloration)
←6 M guanidine hydrochloride, 10 mM DTT, 100 mM ammonium bicarbonate (15 min; reduction)
←6 M guanidine hydrochloride, 500 mM acrylamide, 100 mM ammonium bicarbonate (15 min; alkylation) ←50% methanol 10% acetate (10 min, 3times; wash) ←50% acetonitrile (10 min; anhydration)
←100% acetonitrile (10 min; anhydration)
←50 mM ammonium bicarbonate (10 min; swelling) ←100% acetonitrile (10 min; anhydrration)
←50 mM ammonium bicarbonate (10 min; swelling)
←50% acetonitrile/50 mM ammonium bicarbonate (10 min; anhydration) ←100% acetonitrile (10 min; anhydration)
(centrifugal concentration)
←10 ng/µL trypsine (gel digestion)
←50 mM ammonium bicarbonate (37°, overnight; digestion) ←50% acetonitrile (sonication, 15 min; extraction)
←100% acetonitrile (sonication, 15 min; extraction) (centrifugal concentration)
Peptide sample for MALDI-TOFMS
61 6-4-4. PMF 解析による標的分子の同定
濃縮した標的分子ペプチドサンプル及び 2 種混合スタンダード※1をマトリッ
クス α-Cyano-4-hydroxycinnamic acid と混合し、MALDI-TOFMS に供した。標的 分子の peptide mass fingerprint (PMF) 解析は MALDI-TOFMS AXIMA Performance を用いて以下の設定で行った。
※1
2 種混合スタンダード
ProteoMass Brdykinin Fragment 1-7 MALDI-MS Standard (SIGMA) ProteoMass P14R MALDI-MS Standard (SIGMA)
MALDI-TOFMS AXIMA Performance Sample plate: 384-wells plates
Turning mode: Reflectron Mass range: 1.0-3,000 Max Laser Rap Rate: 20 Power: 50
Shots: 5 Profiles: 200
Ion Gate: Blank, 700
62 Peak Processing
Peak width: 20
Smoothing methods: off Threshold Offset: 0.1-1.0 mV
Mascot Search Server Search Engine: PMF Data Base: NCBI
Taxonomy: Rattus norvegicus Digest Enzyme: Trypsin Missed Cleavages: 1
Fixed modification: Carbamidomethyl Variable Modifications: Oxidation (M) Treat Masses as: Monoisotopic
Mass Type: MH+ Mass List: 1,000-3,000
63
6-6. Biotin-DMT に対する DMT 及び vialinin A、Ub の競合試験
競合試験には、有機合成した biotin-DMT 及び DMT、vialinin A と大腸菌を用 いて大量発現させた recombinant human USP5 (rhUSP5)、市販の Ub を使用した。 10 µM Biotin-DMT とそれぞれの濃度 (5 µM 及び 10 µM、20 µM、50 µM) に調整 した vialinin A 及び DMT を 1.6 µg/mL rhUSP5 に添加し、その混合物を 4°C で 1 時間撹拌した。また、biotin-DMT に対する Ub の競合試験においても同様の方法 で処理を行った。その後、ストレプトアビジンビーズを添加し、4°C、15 分間撹 拌した。非結合分子を洗浄した後、biotin-DMT に結合した rhUSP5 を溶出させ、 SDS-PAGE で展開し、InstantBlue 染色にて可視化させた。
64
第三章 Vialinin A 及び vialinin A 標的分子 USP5、TNF-α 産生との関
連性
第一節 Vialinin A の脱ユビキチン化酵素活性阻害
Vialinin A の活性アナログ DMT をリード化合物とした biotin 標識化プローブ Biotin-DMT を用いて、vialinin A の RBL-2H3 細胞内標的分子を USP5 と同定した。 USP5 はユビキチン特異的プロテアーゼであり、細胞質内のフリーのポリユビキ チン鎖をモノユビキチン化することが知られている。このことから、rhUSP5 の 酵素活性に与える vialinin 類の影響について検討を行った。まず初めに、vialinin A 及び DMT、biotin-DMT について、Ub テトラマーを基質として阻害試験を行 った。Ub テトラマーと rhUSP5 を反応させると、テトラマーがトリマー、ダイ マー、モノマーへと分解されていく。vialinin A 及び DMT は強力に rhUSP5 の酵 素活性を阻害し、biotin-DMT にも酵素阻害活性が観察された (Figure 3-1)。さら に、蛍光基質 Ub-AMC を用いて vialinin A の rhUSP5 の酵素活性に与える影響に ついても検討を行った。Vialinin A は濃度依存的に rhUSP5 の酵素活性を強力に 阻害し、IC50値: 5.9 µM を与えた (Figure 3-2)。また、市販されている脱ユビキチ
ン化酵素 (USP4 及び USP2 core、USP8、UCH-L1、UCH-L3) に対する vialinin A の影響について検討を行った。Vialinin A は USP4 (IC50値: 1.5 µM) と UCH-L1
(IC50値: 22.3 µM) に対する酵素阻害活性も有することが明らかとなった (Figure
3-3)。このことから、vialinin A は標的分子の USP5 以外の複数の脱ユビキチン化 酵素に対して酵素阻害活性を有することが示唆された。
65
Figure 3-1 Inhibitory effects of vialinin A, DMT and biotin-DMT against rhUSP5 enzymatic activity with K48-linked tetra-Ub substrate.
66
Figure 3-2 Inhibitory effects of vialinin A against rhUSP5 enzymatic activity with Ub-AMC.
67 F igur e 3-3 Inhi b it or y e ff ec ts of vi al ini n A a g ai ns t de u b iqui ti na ti ng e nz ym at ic a ct ivi ty .
68
第二節 USP5 ノックダウン RBL-2H3 細胞における TNF-α 放出及び
産生、転写への影響
Vialinin A の RBL-2H3 細胞内標的分子が USP5 であり、vialinin A は USP5 に酵 素活性阻害を有することから、vialinin A の有する強力な TNF-α 産生阻害活性は USP5 の酵素活性の阻害を介して発現すると予想された。そこで、USP5 ノック ダウン RBL-2H3 細胞を調製し、USP5 の発現減少が TNF-α 産生に与える影響に ついて検討を行った。Vialinin A 標的分子の USP5 及び vialinin A により強力な酵 素活性の阻害が認められた USP4 及び USP5 とドメイン構造の類似している USP13 についてノックダウン細胞を調製することとした。トランスフェクショ ン試薬 Lipofectamine RNAiMAX を用いて、設計したそれぞれの 2 重鎖 siRNA を 細胞内に導入し、ノックダウンさせた細胞を抗体感作、抗原処理を行い、細胞 上清サンプルと細胞サンプルを得た。細胞サンプルから、RNA を抽出し、逆転 写にて cDNA を合成後、リアルタイム PCR 法にてそれぞれの脱ユビキチン化酵 素と内部標準の GAPDH の発現量を測定した。USP5 siRNA1 及び USP5 siRNA2 を処理したサンプルでは、Non-target siRNA を処理した Cont.に比較し、有意に 発現量の減少が認められたが、USP5 siRNA3 を処理したサンプルでは発現の減 少が見られなかった。また、USP4 siRNA1 及び USP4 siRNA2、USP13 siRNA1、 USP13 siRNA2 を処理したサンプルで Cont. siRNA 処理と比較し、mRNA レベル において、有意な発現減少が見られた (Figure 3-4)。さらに、タンパク質レベル でも有意な発現減少が観察された (Figure 3-5)。この細胞を用いて、TNF-α の細 胞外への放出量を ELISA 法にて測定した。USP4 及び USP13 をノックダウンさ せた細胞からの TNF-α 放出量は、Cont.細胞と比較し変化がみられなかったのに 対し、USP5 をノックダウンさせた細胞からの TNF-α 放出は有意に減少した
69 (Figure 3-6)。さらに、USP5 ノックダウン細胞からの TNF-α の細胞外への放出量 が有意に減少したことから、TNF-α 産生量について検討することとした。ここ での産生量とは、細胞外に放出された TNF-α と細胞内に残存する TNF-α の和で 示した。USP5 ノックダウン細胞における TNF-α 産生量は、Cont.と比較し有意 な減少が観察された (Figure 3-7)。また、TNF-α mRNA レベルについても検討し たところ、有意な減少が観察された (Figure 3-8)。TNF-α と同じ炎症性サイトカ イン IL-4 及び脱顆粒の指標である β-hexosaminidase の放出量の減少は観察され なかった (Figure 3-9, Figure 3-10)。この RNA 干渉の実験においての細胞毒性は、 LDH の細胞外への放出を検討することで評価し、細胞毒性が認められないこと が観察された (Figure 3-11)。
70 F igur e 3-4 E x p re ss ion of U S P s m R N A i n U S P s si R N A knoc kdow n c el ls .
71 F igur e 3-5 E x p re ss ion of p rot ei n l eve ls of U S P s i n U S P s s iR N A knoc kdow n c el ls .
72 F igur e 3-6 T N F -α r el ea se f rom U S P s s iR N A -knoc kdow n c el ls .
73
74 F igur e 3-8 E x p re ss ion of T N F -α m R N A i n U S P 5 s iR N A -knoc kdow n c el ls .
75 F igur e 3-9 IL -4 r el ea se f rom U S P s s iR N A -knoc kdow n c el ls .
76 F igur e 3-10 E ff ec t on β -he xos am ini da se r el ea se f rom U S P s s iR N A -knoc kdow n c el ls .
77 F igur e 3-1 1 C el l vi ab il it y f or U S P s s iR N A -knoc kdow n c el ls .