低 親 和 性IgGレ
セ プ ター(FcγRII)を
介 す る
ヒ ト好 塩 基 球 の活 性 化機 序 に関 す る研 究
-フ
ロ ー サ イ ト メ ー タ ー に よ る カ ル シ ウ ム 動 態 の 解
析-岡 山大学 医 学部 第二 内科 学教 室(指 導:木 村 郁郎 教 授)
洲
脇
俊
充
(平成6年3月28日
受 稿)
Key words: basophils, FcƒÁRII, fluo-3, Ca2+ mobilization
緒 言 ヒ ト好 塩 基 球 は,全 白血 球 の0.5∼1.0%を 占 め る に す ぎ な い が,木 村 ら が 臨 床 ア レ ル ギ ー と の 関 係 を報 告 し1), Ishizakaら がIgEの 標 的 細 胞 で あ る こ と を明 らか に して 以 来2),肥 満 細 胞 と と も に 気 管 支 喘 息 をは じめ と す る 多 くの ア レ ル ギ ー 疾 患 に 関 与 す る こ と が 知 られ て い る.さ ら に,好 塩 基 球 は,肥 満 細 胞 と共 に,そ の 膜 表 面 上 でIgEのFc部 分 に 対 す る 高 親 和 性 レ セ プ タ ー(FcεRI)を 介 し てIgEと 結 合 し,多 価 ア レ ル ゲ ン が 膜 表 面IgEに 結 合 し てFcεRIが cross-linkingさ れ る と,細 胞 内 顆 粒 に 貯 蔵 さ れ て い た ヒ ス タ ミン等 の 化 学 伝 達 物 質 が 放 出 され あ る い は ロ イ コ ト リエ ン 等 の 脂 質 代 謝 産 物 の 新 た な 合 成,放 出 が 誘 導 さ れ る.ま た,最 近,イ ン タ ー ロ イ キ ンー3(IL-3)で プ ラ イ ム す る こ と に よ り,好 塩 基 球 がIL-4を 産 生 す る3)こ と も報 告 さ れ て お り, FcεRIを 介 して 種 々 の 免 疫 ・ア レ ル ギ ー 反 応 に 深 く関 与 して い る こ とが 明 らか に さ れ つ つ あ る. 一 方,ア トピー 型 の 小 児 喘 息 を代 表 とす るI 型 ア レ ル ギ ー 反 応 で はIgE反 応 系 が 基 本 で あ る が,非 ア トピ ー 型,特 に,木 村 の 提 唱 す る 中 高 年 発 症 型 難 治 性 喘 息4)では, IgEの 関 与 は 少 な く, む し ろIII型, IV型 ア レ ル ギ ー 反 応 に よ る細 胞 反 応 型 ア レ ル ギ ー5)の 関 与 が 想 定 され て い る.ま た, 近 年,木 村6), Itoら7)は 遅 発 型 気 道 反 応(LAR) がIgG1抗 体 に よ り惹 起 さ れ る こ と を 報 告 し て お り,好 塩 基 球 系 の 細 胞 反 応 は ア ト ピー 型 喘 息 症 例 の み で な く,非 ア ト ピー 型,特 に,中 高 年 発 症 型 難 治 性 喘 息 で も認 め られ,こ れ ら の 喘 息 病 態 で はIgEよ り も む し ろIgGの 関 与 が 想 定 さ れ る.ま た,好 塩 基 球 膜 表 面 に は, IgGのFc 部 分 に 対 す る 低 親 和 性 レ セ プ タ ー(FcγRII= CD32w)も 存 在 す る こ とが 明 らか と な っ て い る が8-11),そ の 免 疫 ・ア レル ギ ー 反 応 に お け る役 割 に つ い て は 未 だ 不 明 な 点 が 多 い. 以 上 よ り,好 塩 基 球 に お け るFcγRIIの 重 要 性 が 示 唆 され るが,こ れ ま で の と こ ろ 詳 細 な 検 討 は 少 な い.か か る観 点 か ら,教 室 のTanimoto ら12)は,フ ロー サ イ トメ ー ター を用 い,ネ ガ テ ィ ブ セ レ ク シ ョ ン法 に て 好 塩 基 球 を 精 製 す る方 法 を 確 立 し, Takahashiら8)は,こ の 方 法 を 応 用 して 好 塩 基 球 のFCγRIIを は じめ とす る表 面 マ ー カー に つ い て 検 討 し た.さ ら に今 回 著 者 は, FcγRIIのcross-linkingに よ る好 塩 基 球 の 活 性 化 機 構 を解 明 す る 目的 で, phycoerythrin(PE) 標 識 モ ノ ク ロ ー ナ ル 抗 体 との 二 重 蛍 光 染 色 に よ り末 梢 血 か ら 比 重 遠 沈 法 に よ っ て 部 分 精 製 した 好 塩 基 球 を用 い て, Ca2+指 示 薬 のfluo-3に よ る細 胞 内Ca2+濃 度 の 測 定 に 応 用 し た. 対 象 と 方 法 1.対 象 健 常 成 人6例 を 対 象 と し た.年 齢 は30歳 ∼38 歳(平 均32.0歳)で,性 別 は 男5例,女1例 で あ っ た. 847
2.方 法 1)好 塩 基 球 の 部 分 精 製(比 重 遠 沈 法) 末 梢 血 好 塩 基 球 の 部 分 精 製 は, Leonardら の 方 法13)に従 い 比 重 遠 沈 法 に よ り行 った.す な わ ち, EDTA加 末 梢 血 を 等 張Percoll(Pharmacia 社)の 不 連 続 勾 配(d=l.079及 びd=1.070) の 上 に 重 層 し,室 温, 700×g, 15分 間 遠 沈 し, 1.070と1.079の 間 に 形 成 さ れ る単 核 球 層 を採 取
し, basophil-enriched mononuclear cells (MNCs)を 得, Cat+, Mg2+を 含 ま な いHanks' balanced salt solution(HBSS)(S量gma社)+
10mM EDTA(Sigma社)+0.1% BSA(Sigma 社)(以 下HBSS-EDTA-BSA)に て 洗 浄 し た. 赤 血 球 が 混 在 した 場 合 に は 蒸 留 水 に て 溶 血 させ (hypotonic lysis),同 量 の1.8%食 塩 水 で 等 張 と し, HBSS-EDTA-BSAに て 洗 浄 した.こ の 時 点 に お け る 好 塩 基 球 の 純 度 は7-12%で,混 在 す る細 胞 は 殆 どが リ ン パ 球 で あ り他 の 顆 粒 球 の 混 在 は 認 め な か っ た.な お,好 塩 基 球 の 染 色 に は 好 酸 球 ・好 塩 基 球 同 時 算 定 液1)を 用 い た. 2)好 塩 基 球 以 外 の 細 胞 のPE蛍 光 染 色 と fluo-3の 封 入 1)で 作 成 したbasophil-enriched MNCsの う ち,好 塩 基 球 以 外 の 細 胞 を染 色 す る 目的 で, 各 細 胞 群 に 比 較 的 特 異 的 な下 記 のPE標 識 モ ノ ク ロ ー ナ ル 抗 体(リ ュ ウ シ リー ズ, Beckton-Dickinson社)を 加 え,ま ず4℃,暗 所 で30分 反 応 させ た. (1) CD2(Leu-5B): T細 胞 (2) CD14(Leu-M3):単 球 (3) CD16(Leu-11B): NK細 胞,好 中 球 (4) CD19(Leu-12): B細 胞 次 に, Ca2+指 示 薬 と してfluo-3(Dojindo社) を最 終 濃 度 が4μMと な る よ うに 加 え,室 温,暗 所 で さ ら に40分 反 応 させ, Ca2+, Mg2+を 含 む HBSSに て2回 洗 浄 後, 1×106cells/mlの 細 胞 浮 遊 液 と した.こ の 段 階 に お け る ト リパ ン ブ ル ー 染 色 に よ る生 細 胞 率 は90%以 上 で あ っ た. 3)細 胞 内Ca2+濃 度 の 測 定 細 胞 内Ca2+濃 度 の 測 定 は, FACStarフ ロー サ イ トメ ー タ ー(Beckton-Dickinson社)を 用 い,二 重 蛍 光 染 色 を行 っ た ,蛍 光 色 素 の 励 起 は ア ル ゴ ン イ オ ン レー ザ ー を 使 用 し,波 長488nm, 400mWで 測 定 し た. 515nmの ロ ン グパ ス フ イ ル タ ー で ア ル ゴ ン イ オ ン レー ザ ー 光 を カ ッ トし, 560nmの ダ イ ク ロ イ ッ ク ブ イ ル ター を使 用 して そ れ ぞ れ の 蛍 光 を分 離 後, 525nmバ ン ドパ ス フ イ ル ター でfluo-3の 蛍 光 を 測 定 した. FSC, SSC,
FL-1はlinearscale, FL-2はlog scaleで 表
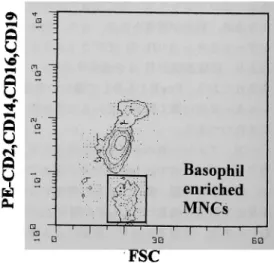
示 し, FSC閾 値 を調 節 して デ ブ リス を カ ッ ト し た.好 塩 基基球 分 画 は, Fig. 1の 如 くFSCとFL -2画 面 上 でPE非 染 色 細 胞 と して 同 定 可 能 で あ っ た.従 っ て,か か るFig. 1に 示 した 好 塩 基 球 分 画 に つ い て, FSC, FL-1画 面 上 で 各 刺 激 に 対 す る細 胞 内Ca2+濃 度 を経 時 的 に 解 析 した. 細 胞 内Ca2+濃 度 の 測 定 は,カ ル シ ウ ム イ オ ノ フ ォ アA23187(10μM, Sigma社)添 加 時 の 平 均 蛍 光 強 度 を最 大 値Fmaxと し14), 2mM MnCl2を カ ル シ ウ ム イ オ ノ フ ォ ア処 理 した 細 胞 に 添 加 し た 際 の 平 均 蛍 光 強 度(FMnCl2)よ り,最 小 値Fminを 算 出15)し て,各 実 験 ご と に 較 正 を行 い, fluo-3の 平 均 蛍 光 強 度 か ら,次 に 示 す 計 算 式 に て 算 出 し た16). [Ca2+]i(nM)=Kd×[F-Fmin]/[Fmax-F] Kd: Fluo-3の 解 離 定 数(400nM) F:検 体 のfluo-3平 均 蛍 光 強 度 Fmax:カ ル シ ウ ム イ オ ノフ ォ アA23187(10 μM)刺 激 後 の 蛍 光 強 度 Fm;n: 1.25×FMnCl2-0.25×Fmax
Fig. 1 Basophil-enrichedmononuclear cellsの サ イ ト グ ラ ム
4) Heat aggregated IgG(agg. IgG)の 作 成
多価IgGと して のagg. IgGは, chromatogra phically purified human IgG(Zymed社)に 63℃, 60分 間 熱 処 理 を 加 え た 後, 10000×g, 10 分 間 遠 沈 し, large aggregatesを 除去 して 作 成 し た. 5)ヒ ス タ ミ ン遊 離 好 塩 基 球 活 性 化 の 一 指 標 と し て,ヒ ス タ ミ ン 遊 離 値 を 測 定 し た.す な わ ち, basophil enriched MNCsを 好 塩 基 球 が1×103cells/mlと な る よ う に 調 節 してCa2+, Mg2+を 含 むHBSSに 浮 遊 させ, IgGレ セ プ ター を介 す る刺 激, anti-lgE(ε) (Kirkegaard & Perry Laboratories社),及 び カ ル シ ウ ム イ オ ノ フ ォ アA23187(1μg/ml) を加 え て37℃, 45分 間 反 応 させ た の ち, 4℃, 400×gで10分 間 遠 沈 し,上 清 中 の ヒ ス タ ミ ン を RIA法 で 測 定 し た.刺 激 物 質 の 代 わ りにHBSS を加 え 同 様 に 反 応 させ て 得 ら れ た もの を 自 然 遊 離 量 と し て 求 め,全 ヒ ス タ ミ ン量 は 細 胞 浮 遊 液 を100℃, 10分 間 熱 処 理 した もの を 遠 沈 後,上 清 を 測 定 に 供 した. RIA法 に よ る 測 定 は す べ て duplicateで 行 い,ヒ ス タ ミン遊 離 率 は,下 記 の 式 に よ り算 出 して 求 め た.な お,自 然 遊 離 量 は 全 ヒ ス タ ミ ン 量 の10%以 下 で あ っ た. ヒ ス タ ミン 遊 離 率(×100%) =
刺 激 に よ る遊 離 量-自 然遊 離 量/
全 ヒ ス タ ミ ン 量-自 然 遊 離 量 6) IL-3の 増 強 効 果 好 塩 基 球 に 対 す るIL-3の 増 強 効 果 を み る た め, basophil-enriched MNCsにrecombinant human IL-3(rhlL-3, Genzyme社)を100U/mlの 濃 度 で加 え て30分 か ら1時 間 反 応 させ た 後, Ca2+動 態 及 び ヒス タ ミン遊 離 に つ い て検 討 した. 結 果 1.好 塩 基 球 分 画 の 構 成 細 胞 ま ず, PE非 染 色 細 胞 と し て 示 した 細 胞 分 画 が 果 た して 好 塩 基 球 と して 妥 当 か ど う か に つ い て 検 討 した. 1) Fig. 1に 示 すPE非 染 色 細 胞 分 画 を FACStarに て ソ ー テ ィ ン グ し, May-Giemsa染 色 を 行 っ た.好 塩 基 球 の 純 度 は78-85%,ト リ パ ン ブ ル ー 染 色 に よ る 生 細 胞 率 は90%以 上 で, 混 在 す る細 胞 は 殆 どが リン パ 球 で あ り,他 の 顆 粒 球 の 混 在 は 認 め られ なか っ た. 2) Basophil enriched MNCsに 対 し て, PE 染 色 と同 時 にFITC標 識anti-IgE(ε)抗 体
(Kirkegaard & Perry Laboratories社)で 染 色 す る と, Fig. 2の サ イ トグ ラ ム に 示 す 如 く,
PE非 染 色 細 胞 群 は 表 面 に 多数 のIgEを 結 合 し
て お り,好 塩 基 球 と し て 妥 当 で あ る こ とが 判 明 し た.
2. 好 塩 基 球 細 胞 内Ca2+動 態 の 解 析
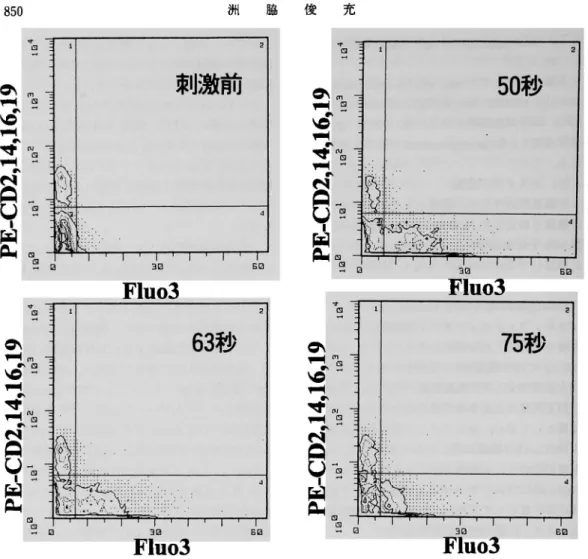
Basophil-enriched MNCsを,ま ずagg. IgG
l00μg/mlで 刺 激 し た際 の 細 胞 内Ca2+動 態 を 検 討 し た.そ の 結 果, Fig. 3の サ イ トグ ラ ム に 示 す 如 く刺 激 前 の 状 態 に 比 し,刺 激 後 約50秒 を ピ ー ク にPE非 染 色 細 胞 す な わ ち好 塩 基 球 のfluo -3蛍 光 強 度 が 右 に 偏 位 して お り,好 塩 基 球 内 Ca2+濃 度 が 増 加 し て い る こ と が 判 明 した. Fig. 4は 同 じデ ー タ を ゲ ー トを か け たPE非 染 色 細 胞 集 団 に つ い て ヒ ス トグ ラ ム 表 示 し た も の で あ る が,刺 激 前 の 状 態 に 比 しagg. IgG刺 激 に よ り明 らか にfluo-3蛍 光 強 度 が 増 加 して い た. Fig. 5は, fluo-3蛍 光 強 度 か ら 算 出 し た 細 胞 内Ca2+ 濃 度 を 経 時 的 に グ ラ フ化 した もの で あ るが,刺 激30秒 後 頃 か ら細 胞 内Ca2+濃 度 の 上 昇 が 認 め ら れ,約50秒 後 に ピ ー クに 達 した 後,約100秒 後
Fig. 2 PE標 識-anti-CD2, CD14, CDl6, CD19と FITC標 識-anti-IgEに よ る 二 重 蛍 光 染 色
Fig. 3 Agg. IgG(100μg/ml)刺 激 後 のfluo-3蛍 光 強 度 の サ イ ト グ ラ ム
Fig. 4 Agg. IgG(l00μg/ml)刺 激 後 のfluo-3蛍 光 強 度 の ヒ ス トグ ラ ム
Fig. 5 Agg. IgG(100μg/ml)刺 激 に よ る 細 胞 内 Ca2+濃 度 の 経 時 的 変 化
に は 刺 激 前 の 状 態 に戻 っ た. agg. IgGの250㎎/ ml, 500μg/mlの 刺 激 で は,濃 度 の 上 昇 と共 にCa2+ 濃 度 の ピ ー ク は や や 増 加 す る傾 向 が 認 め られ た
方 法 でanti-IgE(ε)抗 体0.1μg/ml刺 激 後 の 細
胞 内Ca2+濃 度 変 化 に つ い て 検 討 した と こ ろ,
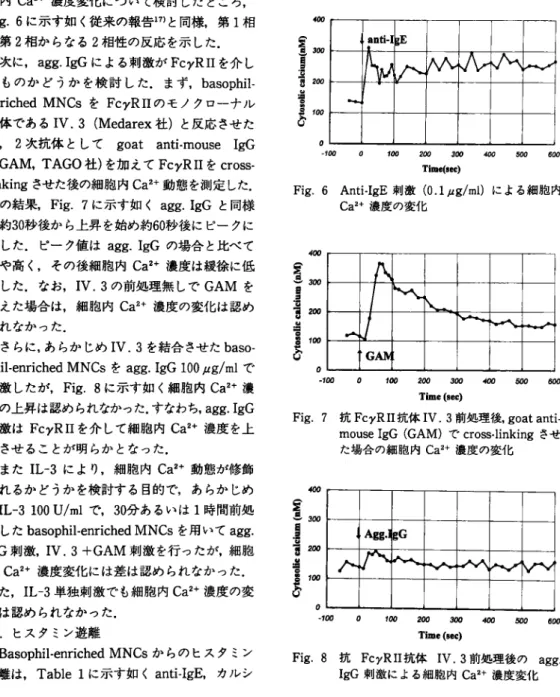
Fig. 6に 示 す 如 く従 来 の 報 告17)と 同様,第1相
と第2相 か ら な る2相 性 の 反 応 を示 し た.
次 に, agg. IgGに よ る 刺 激 がFcγRIIを 介 し
た も の か ど う か を 検 討 し た.ま ず, basophil
-enriched MNCsをFcγRIIの モ ノ ク ロ ー ナ ル
抗 体 で あ るIV. 3(Medarex社)と 反 応 させ た
後, 2次 抗 体 と し てgoat anti-mouse IgG
(GAM, TAGO社)を 加 え てFcγRIIをcross
linkingさ せ た 後 の 細 胞 内Ca2+動 態 を測 定 した.
そ の 結 果, Fig. 7に 示 す 如 くagg. IgGと 同様 に 約30秒 後 か ら上 昇 を始 め 約60秒 後 に ピ ー ク に 達 した.ピ ー ク値 はagg. IgGの 場 合 と比 べ て や や 高 く,そ の 後 細 胞 内Ca2+濃 度 は 緩 徐 に 低 下 し た.な お, IV. 3の 前 処 理 無 しでGAMを 加 え た 場 合 は,細 胞 内Ca2+濃 度 の 変 化 は 認 め られ な か っ た. さ らに,あ らか じめIV. 3を 結 合 させ たbaso phil-enriched MNCsをagg. IgG 100μg/mlで 刺 激 し た が, Fig. 8に 示 す 如 く細 胞 内Ca2+濃 度 の 上 昇 は認 め られ なか った.す な わ ち, agg. IgG 刺 激 はFcγRIIを 介 し て 細 胞 内Ca2+濃 度 を上 昇 させ る こ と が 明 らか と な っ た. ま たIL-3に よ り,細 胞 内Ca2+動 態 が 修 飾 さ れ る か ど う か を検 討 す る 目 的 で,あ らか じめ rhlL-3100U/mlで, 30分 あ るい は1時 間 前 処 理 し たbasophil-enriched MNCsを 用 い てagg.
IgG刺 激, IV. 3+GAM刺 激 を 行 っ た が,細 胞
内Ca2+濃 度 変 化 に は 差 は 認 め ら れ な か っ た. ま た, IL-3単 独 刺 激 で も細 胞 内Ca2+濃 度 の 変 化 は 認 め ら れ な か っ た. 3.ヒ ス タ ミン遊 離 Basophil-enriched MNCsか ら の ヒ ス タ ミン 遊 離 は, Table 1に 示 す 如 くanti-IgE,カ ル シ Fig. 6 Anti-lgE刺 激(0.1μg/ml)に よ る 細 胞 内 Ca2+濃 度 の 変 化
Fig. 7 抗FcγRII抗 体IV. 3前 処 理 後, goat anti-mouse IgG(GAM)でcross-linkingさ せ た 場 合 の 細 胞 内Ca2+濃 度 の 変 化
Fig. 8 抗FcγRII抗 体IV, 3前 処 理 後 のagg . IgG刺 激 に よ る 細 胞 内Caz+濃 度 変 化
Table 1 FcγRIIのcross-linkingに よ る ヒ ス タ ミ ン 遊 離 率(%HR)
ウ ム イ オ ノフ ォ ア刺 激 で は 高 率 で あ っ た が, agg. IgG(100, 250, 500μg/ml)あ るい はIV. 3+GAM
に よ る2種 の 刺 激 で は, IL-3の30分 間 の 前 処 理 の 有 無 に 拘 ら ず,有 意 な ヒ ス タ ミン 遊 離 は 起 こ ら な か っ た. 考 察 好 塩 基 球 は,そ の 膜 表 面 上 に 高 親 和 性IgEレ セ プ ター で あ るFcεRIと 共 に,低 親 和 性IgG レセプ ター のFcγRIIを 有 す るこ とがTakahashi ら の 研 究 に よ り明 ら か とな って い る8-1l)が,そ の 免 疫 ・ア レ ル ギ ー に お け る役 割 に つ い て は 未 だ 十 分 に は 解 明 さ れ て い な い.そ こ で 今 回,ヒ ト 末 梢 血 よ り比 重 遠 沈 法 に て 得 ら れ たbasophil enriched MNCsを 材 料 に,フ ロー サ イ トメー タ ー を 用 い て好 塩 基 球 の 細 胞 内Ca2+濃 度 を測 定 す る方 法 を考 案 し,そ の 上 昇 を指 標 と してFcεR Iの み な らず, FcγRIIを 介 して も好 塩 基 球 が 活 性 化 さ れ る か 否 か を 検 討 し た.そ の 結 果,ヒ ト 好 塩 基 球 はFcεRIだ け で な く, FcγRIIのcross-linkingに よ っ て も活 性 化 さ れ,細 胞 内Ca2+濃 度 の 上 昇 を 引 き起 こ す こ とが 示 され た.し か し, FcγRII刺 激 で は ヒス タ ミンの 遊 離 は 認 め られ ず, そ れ 以 外 の メ デ イ エ ー ター が 関 与 して い る こ と が 示 唆 さ れ た. Ca2+は 細 胞 外 か らの 情 報 を 細 胞 内 に 伝 達 す る セ カ ン ドメ ッ セ ン ジ ャ ー で あ り,細 胞 内Ca2+濃 度 の 上 昇 は 多様 な細 胞 内 情 報 伝 達 過 程 の ご く早 期 に 認 め られ る普 遍 的 な 現 象 で,細 胞 活 性 化 の 指 標 と さ れ て い る.好 塩 基 球 に お い て も, IgEレ セ プ タ ー のcross-linkingに よ り,ま ず 細 胞 内 Ca2+濃 度 の 上 昇 が 起 こ る18).引 き 続 い て セ リ ン プ ロ テ ア ー ゼ,メ チ ル トラ ン ス フ ェ ラ ー ゼ,ア デ ニ レ ー トシ ク ラー ゼ,ボ ス ホ リパ ー ゼC(PLC) な どの 膜 表 面 酵 素 の 活 性 化 が 誘 導 され,特 にPLC に よ りホス フ ァ チ ジル イ ノ シ トー ルが 細 胞 内Ca2+ 反 応 の 基 礎 とな る19, 20)イノ シ トー ル1, 4, 5-三 リ ン 酸 と1, 2-ジ ア シ ル グ リセ ロ ー ル に加 水 分 解 さ れ る. こ れ ま で ヒ ト好 塩 基 球 の 細 胞 内Ca2+濃 度 の 測 定 に は,デ ジ タ ル ビ デ オ 顕 微 鏡 に て 解 析 す る 顕 微 蛍 光 法18)が主 に 用 い ら れ て き た.し か し,こ の 方 法 は,ま ず 好 塩 基 球 を80%以 上 の 高 純 度 に 精 製 す る 必 要 が あ っ た.今 回 確 立 し た フ ロー サ イ トメー ター に よ る 方 法 で は,好 塩 基 球 を 高 純 度 に 精 製 す る必 要 が な く,末 梢 血 か ら比 重 遠 沈 法 を 用 い て10%程 度 に 精 製 す る だ け で,好 塩 基 球 の 細 胞 内Ca2+濃 度 を測 定 す る こ とが 可 能 と な っ た.ま た,好 塩 基 球 をIgE陽 性 細 胞 と して 同 定 す る方 法21)と異 な り,好 塩 基 球 を非 標 識 細 胞 と して 同 定 す る た め,測 定 前 の 段 階 で 好 塩 基 球 を活 性 化 した り脱 感 作 す る こ と な く,目 的 の 刺 激 で 活 性 化 さ れ る か ど う か を検 討 で き る 利 点 が あ る.一 方,多 数 の 細 胞 に お け るCa2+濃 度 の 平 均 を 測 定 す る た め,最 初 のCa2+濃 度 上 昇 は 同 期 して い る の で 大 き な ピー ク と して 認 め ら れ る が,そ れ 以 後 の 反 復 す る ピ ー クは 必 ず し も同 期 し て い な い の で,単 一 細 胞 レベ ル で 観 察 さ れ る 細 胞 内Ca2+濃 度 の オ シ レー シ ョ ン22)の解 析 が で き な い とい う欠 点 が あ る.ま た1つ の 検 体 に つ い て 一 度 解 析 を 開 始 す る と,メ デ イ エ ー タ ー の 測 定 な どの た め に 同 じ検 体 か ら同 時 に は 上 清 を採 取 で き な い こ と,あ る い は 混 在 す る細 胞 (主 に,小 リ ンパ 球,単 球)の 影 響 が 少 な か ら ず あ る こ と な ど が 問題 点 と考 え ら れ る.し か し, 比 較 的 少 量 の 検 体 を 用 い て 簡 便 に 種 々 の 刺 激 に 対 す る好 塩 基 球 の 細 胞 内Ca2+濃 度 変 化 の パ タ ー ン を解 析 で き る点 で ,有 用 と考 え られ る. ま た, Ca2+指 示 薬 と し て 用 い たfluo-3は, 第3世 代 蛍 光 色 素 と して 開 発 さ れ, Ca2+結 合 に よ る 蛍 光 強 度 の 増 加 が, quin-2(第l世 代), indo -1, fura-2(第2世 代)に 比 べ て 強 く,ま た励 起 波 長480∼500nM,蛍 光 波 長530nMで ア ル ゴ ン レー ザ ー フ ロ ー サ イ ト メー ター を用 い て 細 胞 内Ca2+濃 度 の 上 昇 を 決 定 で き る と い う利 点 が あ る23).カ ル ボ キ シ ル 基 を脂 溶 性 の ア セ トキ シ メ チ ル エ ス テ ル体 に し たfluo-3AMは,疎 水 性 で 細 胞 膜 を通 過 し て 細 胞 質 へ 入 っ た 後,非 特 異 的 な エ ス テ ル 水 解 酵 素 に よ っ て 加 水 分 解 され るた め,容 易 に 細 胞 内 へ 封 入 す る こ と が で き る.以 上 よ り, fluo-3とPE標 識 モ ノ ク ロ ー ナ ル 抗 体 を 用 い た 二 重 蛍 光 染 色 に よ り,ア ル ゴ ン レ ー ザ ー フ ロー サ イ トメ ー ター を 用 い て細 胞 内Ca2+濃 度 を 測 定 す る こ とが 可 能 と な っ た. こ の 方 法 を用 い て, anti-lgE(ε)に よ る細 胞 内Ca2+動 態 を検 討 し た と こ ろ, Fig. 6に 示 す
如 く,こ れ まで の 報 告 同 様24,25)第1相 と第2相 か らな る2相 性 のパ ター ン を示 し た.一 方, agg. IgG 刺 激 で は, C5a24)や 血 小 板 活 性 化 因 子(PAF)26) 類 似 の 一 相 性 パ タ ー ン を示 し た.ま た, IV. 3+
GAMに よ る 刺 激 で は, agg. IgG刺 激 と比 べ て
ピー ク に 達 し た 後 のCa2+濃 度 の低 下 は 緩 徐 で,
10分 以 上 の 経 過 で 刺 激 前 の レベ ル に 戻 っ た.こ れ は, FcγRIIが 低 親 和 性 の レ セ プ ター で あ り,
agg. IgG刺 激 が 免 疫 複 合 体 類 似 で,よ り生 理 的
にFcγRIIをcross-linkingさ せ た の に 対 し,
IV. 3+GAMに よ る刺 激 で は, FcγRIIを 非 生
理 的 にcross-linkingさ せ た 結 果 で あ る と考 え ら れ る.し か し,何 れ の 刺 激 もPAFと 同 様26)に ヒ ス タ ミン遊 離 は 認 め られ な か っ た.こ の こ と は,細 胞 内Ca2+濃 度 の 一 過 性 上 昇 だ け で は 脱 顆 粒 を起 こ す に は 十 分 で な い17)ことを示 唆 してお り,ヒ ス タ ミン と共 に ロ イ コ ト リエ ン を も遊 離 す るFcεRIのcross-linkingと,ヒ ス タ ミン 遊 離 の 見 ら れ な いFcγRIlのcross-1inkingと で はCa2+を 含 め た そ の 後 の 細 胞 内 情 報 伝 達 過 程 に相 違 が あ る と考 え られ る.
ヒ トに お い て, IgG reaginic antibody27)が 存 在 す るか ど うか は 異論 の あ る とこ ろで あ り, Ishiza-kaら は, agg. IgGと125Iで ラ ベ ル したanti-IgG
を一 緒 に 反 応 させ る こ とに よ り,ヒ ト好 塩 基 球 表 面 にIgGが 結 合 し て い る こ とを最 初 に 証 明 し た28).ま た, Takahashiら は フ ロー サ イ トメ ー タ ー と モ ノ ク ロー ナ ル 抗 体 を 用 い て,好 塩 基 球 表 面 にIgG,とIgG4が 結 合 し て い る こ と を 報 告 して い る8). IgG4抗 体 に 関 し て は,ヒ ス タ ミ ン が 遊 離 され る とす る報 告29-32)と,こ れ を否 定 す る報 告33-35),あ る い は 減 感 作 療 法 に お い て 遮 断 抗 体 と し て 働 く とい う報 告36,37)もあ り,そ の役 割 に つ い て は 未 だ 一 定 の 見 解 が 得 ら れ て い な い の が 現 状 で あ る.最 近, Lichtensteinら は,ア トピー 患 者 に お け るanti-IgGモ ノ ク ロー ナ ル 抗 体 に よ る ヒス タ ミン 遊 離 は,好 塩 基 球 上 のFcγR IIに 結 合 したIgG抗 体 に よ る もの では な く, FcεR
Iに 結 合 し たIgEに 対 す るIgG自 己 抗 体 を介
し て 間 接 的 にFcεRIをcross-linkingし た結 果 で あ る と して お り39),好 塩 基 球 のFcγRIIの cross-linkingに よ る ヒ ス タ ミン遊 離 が 認 め られ な か っ た 今 回 の 結 果 と一 致 す る.ま た, IgGlに 関 して,木 村6), Itoら7)はLARがIgG1抗 体 に よ り惹 起 され る と報 告 して い るが,好 塩 基 球 のIgGレ セ プ ター の状 態 を免 疫 走 査 電 顕 法 に よ り検 討 す る と,喘 息 患 者 の 免 疫 グ ロ ブ リ ン 密 度 は 重 症 例 やLARを 呈 す る症 例 に 高 密 度 で あ り40),血 清 中41)や気 管 支 肺 胞 洗 浄 液 中42)のIgG サ ブ ク ラ ス で検 討 す る と抗 原 特 異 的IgG1が 優 位 な症 例 で はLARを 呈 しや す い こ と が 知 られ て い る.以 上 よ り,好 塩 基 球 のFcγRIIを 介 す る活 性 化 に は, IgG,抗 体 が よ り強 く関 与 し て い る と考 え られ る. IL-3は,特 異 的 レ セ プ タ ー を 介 し て43,44,)好塩 基 球 の 増 殖,分 化,遊 走 因 子 と して 機 能 し,さ らに メデ ィ エ ー ター 遊 離 に も関 与 して い る45-47). ま た ヒ ト好 塩 基 球 は, C5a単 独 刺 激 で は ヒ ス タ ミ ン は 遊 離 す るが,ロ イ コ ト リエ ンC4(LTC4) は 産 生 し な い もの の, IL-3で10分 間 前 処 理 す る こ と に よ り, C5aで もLTC,を 産 生 す る よ うに な る と報 告 さ れ て い る45).か か る機 序 を 細 胞 内 Ca2+動 態 の 面 か ら解 析 す る と, C5a単 独 で は2 分 以 内 の 一 過 性 のCa2+濃 度 の 上 昇 しか 来 さ な い が, IL-3で18時 間 前 処 理 す る とfMLPに 類 似 の 持 続 的 なCa2+濃 度 の 上 昇 を示 す よ う に な り,そ れ がLTC、 産 生 に 必 須 の 要 因 と考 え ら れ て い る25).し か し, IL-3の8分 間 の 前 処 理 で は C5aに よ るLTC4産 生 を 引 き起 こす が, Ca2+濃 度 変 化 に は影 響 しな い とさ れ て お り48),今回 の 実 験 で も好 塩 基 球 に お い て はIL-3そ れ 自 身 がCa2+ 濃 度 を変 化 させ ず,か っFcγRIIのcross-linking に よ るCa2+濃 度 変 化 に は,少 な く と も1時 間 以 内 のIL-3前 処 理 で は 影 響 が み ら れ な か っ た. この こ とは, IL-3の 短 時 間 の プ ライム 効 果 はCa2+ 濃 度 変 化 の 相 違 だ け で は 説 明 で き な い こ と を示 し て お り,ヒ ス タ ミ ン遊 離 の 初 期 相 に お い て は, よ り局 所 的 なCa2+反 応 と かCa2+濃 度 の 上 昇 と平 行 して 起 こ る他 の 活 性 化 機 序 が よ り重 要 で あ る と考 え られ る. ア レ ル ギ ー の 遅 発 型 反 応(LPR)局 所 に は, 好 酸 球 と共 に 好 塩 基 球 が 存 在 す る こ とが,皮 膚49,50),鼻51-53),下気 道54,55)にお い て 認 め られ て い る.こ の こ とは,他 の 炎 症 細 胞 か ら好 塩 基 球 遊 走 因 子 が 出 て い る こ と を 予 想 させ る.実 際, 抗 原 に よ る遅 発 型 皮 膚 反 応 の 皮 膚 生 検 組 織 に は
IL-3, IL-4, IL-5, GM-CSFのmRNAが56), 喘 息 患 者 の 気 管 支 粘 膜 に はIL-5のmRNA57)が 発 現 して い る こ とがin situ hybridizationに よ っ て 認 め られ て い る.ま た, MasseyはGM-CSF 活 性 が ア レ ル ギ ー 患 者 の 皮 膚LPR局 所 に 存 在 す る こ と を バ イ オ ア ッ セ イ とELISAで 認 め て い る58).従 っ て, LPR局 所 で は,抗 原 に 反 応 し てT細 胞 か ら産 生,放 出 さ れ るIL-3, IL-5, GM-CSFが 好 塩 基 球 に 対 す る遊 走 活 性 を持 っ て い る59,60)ことか ら,組 織 へ の 好 塩 基 球 の 集 積 に 重 要 な 役 割 を 果 た して い る と考 え ら れ る61).一 方, IL-5は 好 塩 基球 に 対 し て は,ヒ ス タ ミン よ り も LTC4の 産 生 を増 強 す る作 用 が認 め られ てお り62), LPR局 所 で の 好 塩 基基球 か らのLTC、 産 生 の 可 能 性 も考 え ら れ る.ま た, FcγRIIは,ト リプ シ ン の 作 用 に よ り高 親 和 性 のFcγRIIと して 働 き 得 る ス タ ン バ イ レ セ プ タ ー で あ り63,64),ヒトの 単 球 に お い て は,プ ロ テ ア ー ゼ 処 理 に よ り高 親 和 性 と な っ たFcγRIIのcross-linkingに よ っ て 腫 瘍 壊 死 因 子(TNF)が 産 生 され る65).か か る 事 実 か ら,プ ロ テ ア ー ゼ の 豊 富 な 炎 症 の 場 に 遊 走 して き た好 塩 基 球 に お い て も, FcγRIIを 介 す る刺 激 が よ り増 強 さ れ て い る 可 能 性 が 示 唆 さ れ る. 肥 満 細 胞 は, anti-IgE刺 激 に よ り脱 顆 粒 を 起 こす の み な らず,各 種 サ イ トカ イ ン の 遊 離 を 誘 導 す る66,67)が, Paulら は,マ ウ ス の 脾 臓,骨 髄 のnon-B, non-T細 胞 を用 い た 一 連 の 実 験 か ら, non-B, non-T細 胞 がFcεRI,
FcγRIIのcross-linkingに よ りIL-4を 産 生 す る こ と を 報 告 して
い る68).更 に,こ のIL-4産 生non-B, non-T
細 胞 はFcεRI陽 性 で,形 態 学 的 に ほ とん ど が 好 塩 基 球 ま た は 好 塩 基 性 骨 髄 球 で あ り,肥 満 細 胞 よ り もむ し ろ好 塩 基球 が 主 なIL-4産 生 細 胞 で あ る と し て い る69).一 方, Piccinniら は,ヒ ト骨 髄 のnon-B, non-T細 胞 を用 い て 同 様 の 実 験 を行 い, IL-3の 存 在 下 でFcεR, FcγRの
cross-linkingに よ るIL-4, IL-5のmRNAの 発 現 を 報 告 して お り70),実 際, IL-3で 前 処 理 す る こ と に よ り,正 常 末 梢 血 好 塩 基 球 がIL-4を 産 生 す る事 も報 告 さ れ て い る3). 以 上 の 如 く,T細 胞 か ら遊 離 さ れ たIL-3, IL -5, GM-CSFに よ り, LPR局 所 に 遊 走 し て き た 好 塩 基 球 は,プ ロ テ ア ー ゼ, IL-3, IL-5な ど に よ っ て よ り活 性 化 さ れ, IgG,免 疫 複 合 体 で FcγRIIが 刺 激 さ れ る こ と に よ り,ヒ ス タ ミン 以 外 の メ デ イ エ ー タ ー,す な わ ちLTC4或 い はIL -4, IL-5な どの サ イ トカ イ ン を 産 生 す る可 能 性 が 示 唆 さ れ,オ ー トク ラ イ ン あ る い は パ ラ ク ラ イ ン の 機 序 に よ る ア レ ル ギ ー 反 応 の 増 幅,あ る い は 他 の 細 胞 群 との 複 雑 な ネ ー トワ ー クの 形 成 に 関 与 して い る こ と が 推 察 され た. 結 論 ヒ ト好 塩 基 球 の 活 性 化 を,細 胞 内Ca2+濃 度 の 上 昇 を指 標 と し て フ ロ ー サ イ トメ ー ター を用 い て 検 出 す る 方 法 を確 立 し, IgEの み な らずIgG に よ っ て も好 塩 基 球 が 活 性 化 さ れ る か 否 か に つ い て 検 討 し た . 1.ヒ ト好 塩 基 球 に お い て, FcγRIIのcross-linkingに よ り細 胞 内Ca2+濃 度 の 上 昇 が 認 め ら れ た が, IL-3で30分 か ら1時 間 前 処 理 した 細 胞 との 間 に 明 らか な 相 違 は 認 め ら れ な か っ た. 2. FcγRIIのcross-linkingに よ る ヒ ス タ ミ ン 遊 離 は 認 め ら れ な か っ た. 以 上,ヒ ト好 塩 基 球 で は, FcγRIIのcross-linkingに よ りヒ ス タ ミン遊 離 は 認 め られ な か っ た が,セ カ ン ド メ ッ セ ン ジ ャ ー と して の 細 胞 内 Ca2+濃 度 の 上 昇 が 認 め られ た こ と か ら, IgG系 の 刺 激 で は ヒ ス タ ミ ン以 外 の メ デ イ エ ー タ ー が 関 与 して い る こ とが 想 定 さ れ た. 稿 を終 え るに あ た り,終 始 御 指 導 御校 閲 を賜 った 恩 師 木 村 郁 郎教 授 に深 甚 の 謝 意 を表 す る と共 に,直 接 御 指 導 戴 い た 高橋 清 講 師 に 深 謝 致 し ます. 本 論 文 の 要 旨は 第43回 日本 ア レル ギー 学会 総 会 に て発 表 し た. 文 献 1) 木 村郁 郎,守 谷欣 明,西 崎 良知,谷 崎勝 朗:気 管 支 喘 息 に お け る好 塩 基 球 の臨 床 的 意義 .ア レル ギー(1968)
17, 558-561.
2) Ishizaka K, Tomioka H and Ishizaka T: Mechanism of passive sensitization;
I. Presence of IgE and
IgG molecules on human leukocytes.
J Immunol (1970) 105, 1459-1470.
3) Brunner T, Heusser CH and Dahinden CA: Human peripheral blood basophils primed by interleukin
3 (IL-3) produce IL-4 in response to immunoglobulin E receptor stimulation.
J Exp Med (1993) 177,
605-611.
4) 木 村 郁 郎:喘 息 の 病 型 と その 本 質 論-中 高 年 発 症 型 難 治 性 喘 息 の 独 立 性.日 胸 疾 会 誌(1983) 21, 181-182. 5) 木 村 郁 郎:気 管 支 肺 病 変 に お け る ア レ ル ギ ー と リ ン パ 球.ア レ ル ギ ア(1990) 19, 12-16.
6) 木 村 郁 郎:遅 発 ア レ ル ギ ー の 発 症 機 序-細 胞 反 応 を 中 心 に-;第3回 免 疫 薬 理 シ ン ポ ジ ウ ム 記 録 集,冨 岡 玖 夫 編,デ ー ・エ ム ・べ ー ・ジ ャ パ ン,東 京(1985) pp 23-40.
7) Ito K, Kudo K, Okudaira H, Yoshinoya S, Morita Y and Nakagawa
T: IgG1 antibodies to house
dust mite (Dermatophagoides farinae) and late asthmatic response. Int Arch Allergy Appl Immunol
(1986) 81, 69-74.
8) Takahashi K, Takata
M, Suwaki T, Kawata N, Tanimoto Y, Soda R and Kimura I: New flow
cytometric method for surface phenotyping basophils from peripheral blood. J Immunol Methods
(1993) 162, 17-21.
9) Stain C,
Stockinger
H, Schael M, Jager U, Gossinger H, Lechner K and Bettelheim P: Human
blood basophils display a unique phenotype including activation linked membrane structures.
Blood
(1987) 70, 1872-1879.
10) Anselmino LM,
Perussia B and Thomas LL: Human basophils selectively express the FcyRII
(CD32w) subtype of IgG receptor.
J Allergy Clin Immunol (1989) 84, 907-914.
11) Valent P, Majdic O,
Maurer D, Bodger M, Muhm M and Bettelheim P: Further characterization
of surface membrane structure exprss on human basophils and mast cells. Int Arch Allergy Appl
Immunol (1990) 91, 198-203.
12) Tanimoto Y, Takahashi
K, Takata
M, Kawata N and Kimura I: Purification
of human blood
basophils using negative selection by flow cytometry.
Clin Exp Allergy (1992) 22, 1015-1019.
13) Leonard EJ, Roberts RL and Skeel A: Purification of human blood basophils by single step isopycnic
banding on percoll. J Leukocyte Biology (1984) 35, 169-177.
14) Deber CM, Tom-Kun J, Mack E and Grinsoein S: Bromo-A 23187: A non fluorescent
calcium
ionophore for use with fluorescent probes. Anal Biochem (1985) 146, 349-352.
15) Hesketh TR,
Smith GA,
Moore JP,
Taylor MV and Metcalfe JC: Free cytoplasmic
calcium
concentration
and the mitogenic stimulation of lymphocytes.
J Biol Chem (1983) 258, 4876-4882.
16) Vandenberghe
PA and Ceuppens JL: Flow cytometric measurement of cytoplasmic free calcium in
human peripheral blood T lymphocytes with fluo-3, a new fluorescent calcium indicator.
J Immunol
Methods (1990) 127, 197-205.
17) MacGlashan DW Jr and Botana LM: Biphasic Ca2+ responses in human basophils. J Immunol (1993)
150, 980-991.
18) Warner JA and MacGlashan DW Jr: Signal transduction events in human basophils-a
compara
tive study of the role of protein kinase-C in basophils activated by anti-IgE antibody and formyl
methionyl-leucyl-phenylalanine.
J Immunol (1990) 145, 1897-1905.
19) Nishizuka Y: Turnover of inositol phosopholipids and signal transduction.
Science (1984) 225, 1365
-1370 .
Rev Biochem (1987) 56, 159-193.
21) Bochner BS, McKelvey AA, Schleimer RP, Hildreth JEK and MacGlashan DW Jr: Flow cytometric
methods for the analysis of human basophil surface antigens and viability.
J Immunol Methods
(1989) 125, 265-271.
22) MacGlashan DW Jr and Guo CB: Oscillation in free cytosolic calcium during IgE-mediated stimula
tion distinguish human basophils from human mast cells. J Immunol (1991) 147, 2259-2269.
23) Minta A, Kao JPY and Tsien RY: Fluorescent indicators for cytosolic calcium based on rhodamine
and fluorescein chromophores.
J Biol Chem (1989) 264, 8171-8178.
24) MacGlashan DW Jr and Warner JA: Stimulus-dependent
leukotriene release from human basophils:
a comparative
study of C5a and Fmet-leu-phe.
J Leukocyte Biology (1991) 49, 29-40.
25) Knol EF, Verhoeven AJ and Roos D: Stimulus secretion coupling in human bosophilic granulocytes.
Clin Exp Allergy (1993) 23, 471-480.
26) Columbo M, Casolaro V, Warner JA, MacGlashan DW Jr, Kagey-Sobotka
A and Lichtenstein LM:
The mechanism of mediator release from human basophils induced by platelet-activating
factor. J
Immunol (1990) 145, 3855-3861.
27) Parrish WE: Short-term anaphylactic
IgG antibodies in human sera. Lancet (1970) 2, 591-592.
28) Ishizaka T, Sterk AR and Ishizaka K: Demonstration
of Fcy receptors on human basophil granulo
cytes. J Immunol (1979) 123, 578-583.
29) Vijay HM and Perelmutter L: Inhibition of reagin-mediated
PCA reactions in monkeys and hista
mine release from human leukocytes by human IgG4 subclass. Int Arch Allergy Appl Immunol (1977)
53, 78-87.
30) Nakagawa
T and de Weck AL: Membrane receptors for the IgG4 subclass on human basophils and
mast cells. Clin Rev Allergy (1983) 1, 197-206.
31) Nakagawa
T, Stadler BM, Heiner DC, Skvaril F and de Weck AL: Flow-cytometric
analysis of
human basophil degranulation.
II. Degranulation
induced by anti-IgE,
anti-IgG, and the calcium
ionophore A 23187. Clin Allergy (1981) 11, 21-30.
32) Fagan DL, Slaughter DA, Capra DC and Sullivan TJ:
Monoclonal antibodies to immunoglobulin
G, induce histamine release from human basophils in vitro. J Allergy Clin Immunol (1982) 70, 399
-404
.
33) Jimeno L, Lombardero M, Carreira J and Prado JMD: Presence of IgG4 on the membrane of human
basophils.
Histamine release is induced by monoclonal antibodies directed against the Fab but not
the Fc region of the IgG, molecule.
Clin Exp Allergy (1992) 22, 1007-1014.
34) Van Toorenenbergen
AW and Aalberse RC: IgG, and passive sensitization of basophil leukocytes.
Int Arch Allergy Appl Immunol (1981) 65, 432-440
35) Van Toorenenbergen
AW and Aalberse RC: IgG4 and release of histamine from human peripheral
leukocytes.
Int Arch Allergy Appl Immunol (1982) 67, 117-122.
36) Devey ME and Panzani R: The IgG subclass of antibodies to castor began allergen in patients with
allergic asthma; detection of high incidence of antibodies of the IgG4 subclass.
Clin Allergy (1975)
5, 353-361.
37) Van der Gissen M, Homan WL, van Karnebeek
G, Aalberse RC and Dieges PH: Subclass typing
of IgG antibodies formed by grass-pollen allergic patients during immunotherapy.
Int Arch Allergy
Appl Immunol (1976) 50, 625-639.
disease. Clin Exp Allergy (1991) 21, 289-296.
39) Lichtenstein LM, Kagey-Sobotka
A, White JM and Hanilton RG: Anti-human IgG causes basophil
histamine release by acting on IgG-IgE complexes bound to IgE receptors.
J Immunol (1992) 148,
3929-3936.
40) 岡田 千春,高 橋 清,宗 田 良,松 岡 孝,難 波 一 弘,荒 木 洋行,木 村 郁 郎,岸 本 卓 巳:遅 発 型 気 道 反 応 に お け る好塩 基 球 表 面 免疫 グ ロ ブ リンの検 討.ア レル ギー(1988) 37, 5-11. 41) 小 栗栖 和 郎:遅 発 型 気 道反 応 にお け るIgGサ ブ クラ ス抗 体 に 関す る研 究.第1編 血 清 中抗 原特 異 的IgG サ ブ ク ラス抗 体 と遅 発 型気 道 反 応 の 関連 につ いて.岡 山医 誌(1992) 104, 797. 807. 42) 小 栗栖 和 郎:遅 発 型 気 道 反応 にお け るIgGサ ブ クラ ス抗体 に 関す る研 究.第2編 気管 支 肺 胞 洗 浄 液 中抗 原 特 異 的IgGサ ブ クラ ス抗 体 と遅 発型 気 道 反 応 の関 連 につ い て.岡 山医 誌(1992) 104, 809-820.43) Hirai K, Morita Y, Misaki K, Ohta S, Suzuki K, Motoyoshi K and Miyamoto T: Modulation of
human basophil histamine release by hematopoietic
growth factors.
J Immunol (1988) 141,
3958-3964.
44) Valent P, Besember J, Muhm M, Majdic O, Lechner K and Bettelheim P: Interleukin 3 activates
human blood basophils via high-affinity binding sites. Proc Natl Acad Sci USA (1989) 86,
5542-5546.
45) Kurimoto Y, de Weck AL and Dahinden CA: Interleukin 3-dependent mediator release in basophils
triggered by C5a. J Exp Med (1989) 170, 467-479.
46) Schleimer RP, Derse CP, Friedman B, Gillis S, Plaut M, Lichtenstein LM and MacGlashan DW
Jr: Regulation of human basophil mediator release by cytokines.
I. Interaction with antiinflam
matory steroids.
J Immunol (1989) 143, 1310-1317.
47) Valent P, Schmidt G, Besemer J, Mayer P, Zenke G, Liehl E, Hinterberger W, Lechner K, Maurer
D and Bettelheim P: Interleukin-3 is a differentiation
factor for human basophils. Blood (1989) 73,
1763-1769.
48) Krieger M, von Tscharner
V and Dahinden CA: Signal transduction
for interleukin-3-dependent
leukotriene
synthesis in normal human basophils:
opposing role of tyrosine kinase and protein
kinase C. Eur J Immunol (1992) 22, 2907-2913.
49) Charlesworth
EN, Hood AF, Soter NA, Kagey-Sobotka
A, Norman PS and Lichtenstein LM:
Cutaneous late-phase response to allergen: mediator release and inflammatory
cell infiltration.
J
Clin Invest (1989) 83, 1519-1526.
50) Charlesworth EN, Kagey-Sobotka
A, Schleimer RP, Norman PS and Lichtenstein LM: Predoni
sone inhibits the apperance
of inflammatory
mediators
and influx of eosinophils
and basophils
associated with the cutaneous late-phase response to allergen.
J Immunol (1991) 146, 671-676.
51) Naclerio RM, Proud D, Togias AG, Adkinson NF Jr, Meyers DA, Kagey-Sobotka
A, Plaut M,
Norman PS and Lichtenstein LM: Inflammatory
mediators in late antigen-induced rhinitis.
N Engl
J Med (1985) 313, 65-70.
52) Bascom R, Wahs M, Naclerio RM, Pipkorn U, Galli S and Lichtenstein LM: Basophil influx occurs
after nasal antigen challenge: effect of topical corticosteroid pretreatment.
J Allergy Clin Immunol
(1988) 81, 580-589.
53) Iliopoulos O,
Baroody FM, Naclerio RM, Bochner BS, Kagey-Sobotka
A and Lichtenstein LM
Histamine-containing
cells obtained from the nose hours after antigen challenge have functional and
phenotypic characteristics
of basophils.
J Immunol (1992) 148, 2223-2228.
朗: House Dustに よ る気 管支 喘息 患 者 の遅 発 型 気 道 反 応 の発 症 機 序 に 関 す る検 討-気 管 支 肺 胞 洗 浄 を 中 心 に.ア レル ギ-(1988) 37, 67-74.
55) Liu MC, Hubbard WC, Proud D, Stealy BA, Galli SJ, Kagey-Sobotka A, Bleecker ER and Lichtenstein LM: Immediate and late inflammatory responses to ragweed antigen challenge of the
peripheral airways in allergic asthmatics. Am Rev Respir Dis (1991) 144, 51-58.
56) Kay AB, Ying S, Varney V, Gaga M, Durham SR, Moqbel R, Wardlaw AJ and Hamid Q: Messenger RNA expression of the cytokine gene cluster, interleukin 3 (IL-3), IL-4, IL-5, and granulocyte/macrophage colony-stimulating factors, in allergen induced late-phase cutaneous reac
tions in atopic subjects. J Exp Med (1991) 173, 775-778.
57) Hamid Q, Azzawi M, Ying S, Moqbel R, Wardlaw AJ, Corrigan CJ, Bradley B, Durham SR, Collins JV, Jeffery PK, Quint DJ and Kay AB: Expression of mRNA for interleukin-5 in mucosal
bronchial biopsies from asthma. J Clin Invest (1991) 87, 1541-1546.
58) Massey W, Friedman B, Kato M, Cooper P, Kagey-Sobotka A, Lichtenstein LM and Schleimer RP: Appearance of granulocyte-macrophage colony-stimulating factor activity at allergen-challeng
ed cutaneous late-phase reaction sites. J Immunol (1993) 150, 1084-1092.
59) Tanimoto Y, Takahashi K and Kimura I: Effects of cytokines on human basophil chemotaxis. Clin Exp Allergy (1992) 22, 1020-1025.
60) Yamaguchi M, Hirai K, Shoki S, Takaishi T, Ohta K, Morita Y, Suzuki S and Ito K: Haematopoietic growth factors induce human basophil migration in vitro. Clin Exp Allergy (1992) 22, 379-384.
61) Shute J: Basophil migration and chemotaxis. Clin Exp Allergy (1992) 22, 321-323.
62) Bischoff SC, Brunner T, De-Weck AL and Dahinden CA: Interleukin 5 modifies histamine release and leukotriene generation by human basophils in response to diverse agonists. J Exp Med (1990)
172, 1577-1582.
63) Van de Winkel JGJ, van Ommen R, Huizinga TWJ, de Raad MAHVM, Tuijnman WB, Groenen PJYTA, Capel PJA, Koene RAP and Tax WJM: Proteolysis induces increased binding affinity of
the monocyte type II FcR for human IgG. J Immunol (1989) 143, 571-578.
64) Tax WJM and van de Winkel JGJ: Human FcƒÁ receptor II: A standby receptor activated by proteolysis? Immunol Today (1990) 11, 308-310.
65) Debets JMH, van de Winkel JGJ, Ceuppens JL, Dieteren IEM and Buurman WA: Cross-linking of both FcƒÁRI and FcƒÁRII induces secretion of tumor necrosis factor by human monocytes requiring high affinity Fc-FcƒÁR interactions: Functional activation of FcƒÁRII by treatment with proteases or neuraminidase. J Immunol (1990) 144, 1304-1310.
66) Plaut M, Pierce JH, Watson CJ, Hanley-Hyde J, Nordan RP and Paul WE: Mast cell lines produce lymphokines in response to cross-linkage of FcƒÃRI or to calcium ionophores. Nature (1989) 339, 64
-67 .
67) Wodnar-Filipowicz A, Heusser CH and Moroni C: Production of the hematopoietic growth factors GM-CSF and interleukin-3 by mast cells in response to IgE receptor mediated activation. Nature (1989) 339, 150-152.
68) Ben-Sasson SZ, Gros GL, Conrad DH, Fikelman FD and Paul WE: Cross-linking Fc receptors stimulate splenic non-B, non-T cells to secrete interleukin and other lymphokines. Proc Natl Acad
Sci USA (1990) 87, 1421-1425.
SA, Galli SJ and Plaut M: Mouse splenic and bone marrow cell populations that express high affinity FcƒÃ receptors and produce interleukin 4 are highly enriched in basophils. Proc Natl Acad Sci USA (1991) 88, 2835-2839.
70) Piccinni MP, Macchia D, Parronchi P, Giudizi MG, Bani D, Alterini R, Grossi A, Rlcchi M, Maggi E and Romagnani S: Human bone marrow non-B, non-T cells produce interleukin 4 in response to cross-linkage of FcƒÃ and Fcy receptors. Proc Natl Acad Sci USA (1991) 88, 8656-8660.
The mechanism of human basophil activation
through the low affinity IgG receptor (Fc ƒÁ RII):
Analysis of calcium mobilization using a new flow cytometric method
Toshimitsu SUWAKI
Second Department of Internal Medicine,
Okayama University Medical School,
Okayama 700, Japan
(Director: Prof. I. Kimura)
A new method for flow cytometric analysis of calcium mobilization in human peripheral blood basophils without prior purification was developed. The method is based on dual color analysis of centrifugation-enriched mononuclear cell populations using fluo-3 and phycoeryth rin (PE)-conjugated CD2, CD14, CD16, CD19 monoclonal antibodies (mAb) to stain contaminated cells. This technique allows the detection of fluo-3 fluorescence as a measure of an increase in the cytoplasmic free calcium concentration ( [Ca2+] 1) while simultaneously discriminating PE-mAb-unlabelled basophils.
To clarify whether the human peripheral blood basophil is activated through the low affinity IgG receptor, Fc ƒÁ RII, as well as the high affinity IgE receptor, Fe ƒÃ R I, calcium mobilization after Fc ƒÁ RII stimulation was analyzed by this method. After cross-linking of Fc ƒÁ RII, transient [Ca2+] 1 elevation was observed but there was no apparent difference with interleukin-3 (IL-3)-treated cells, and no significant histamine release was observed with or without short pre-incubation of IL-3. These findings suggest that the cross-linking of Fc ƒÁ RII, not only Fc ƒÃ R I, can activate human basophils which may result in mediator release