ウサギ角膜上皮再構築能を有する幹細胞および 前駆細胞の同定とその性状に関する研究
Identification and cellular properties of stem/progenitor cells that are capable to reconstruct corneal epithelium
in the rabbit model
亀石 統子
Sumako KAMEISHI
生命理工学専攻
分子生理学研究2016年 2月
ウサギ角膜上皮再構築能を有する幹細胞および 前駆細胞の同定とその性状に関する研究
Identification and cellular properties of stem/progenitor cells that are capable to reconstruct corneal epithelium
in the rabbit model
2016年 2月
亀石 統子
Sumako KAMEISHI
早稲田大学大学院 先進理工学研究科 生命理工学専攻 分子生理学研究
目次
第1章 緒言
1-1.上皮組織の分類と構造 ... 1
1-2.眼組織の構造、疾患 ... 2
1-3.幹細胞研究 ... 4
1-3.上皮組織再生医療の発展... 9
1-4.本研究のねらい ... 10
第2章 角膜上皮幹細胞疲弊症モデルの作製とその評価 2-1.序章 ... 12
2-2.実験材料および方法 ... 13
2-2-1.動物 ... 13
2-2-2.角膜上皮欠損モデル(Corneal epithelial deficiency model)動物の作製方法 ... 14
2-3.実験結果、考察 ... 15
2-4.総括 ... 16
第3章 角膜上皮欠損モデルにおける上皮細胞と基底膜の再形成 3-1.序章 ... 17
3-2.実験材料および方法 ... 18
3-2-1.動物 ... 18
3-2-2.組織学的解析 ... 18
3-3.実験結果、考察 ... 21
3-3-1.角膜欠損モデル眼表面の解析 ... 21
3-3-2.上皮細胞の表現型と基底膜構成分子の関係 ... 25
3-3-3.48週角膜欠損モデルにおける輪部様組織の再構築 ... 30
3-3-4.組織修復過程における上皮間葉移行... 32
3-4.総括 ... 34
3-5.今後の展開 ... 37
第4章 角膜上皮幹細胞表現型同定のための輪部上皮SP 細胞研究への着手 4-1.序章 ... 38
4-2.実験材料および方法 ... 40
4-2-1.動物 ... 40
4-2-2.輪部上皮細胞の単離 ... 41
4-2-3.Hoechst 33342 染色による輪部上皮SP 細胞の単離 ... 41
4-3.実験結果、考察 ... 42
4-3-1.Hoechst 33342 染色濃度条件決定 ... 42
4-3-2.次世代シークエンサ解析に向けた輪部上皮SP 細胞とNSP 細胞の分取とRNA 抽 出、逆転写、増幅の実施 ... 45
4-3-3.輪部上皮SP 細胞とNSP 細胞におけるAbcg2 の発現量比較 ... 46
4-4.総括 ... 47
第5章 ウサギ輪部上皮SP 細胞における包括的遺伝子発現解析と表現型の解明 5-1.序章 ... 49
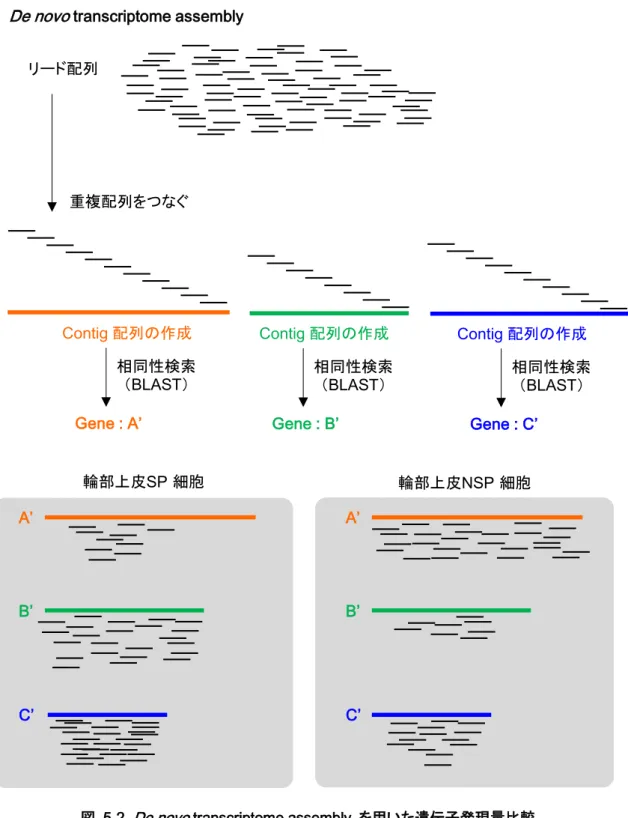
5-2.材料と方法 ... 52
5-2-1.Ion Proton を用いたシークエンス解析 ... 52
5-2-2.新規参照配列の構築 (de novo transcriptome assembly) ... 53
5-2-3.輪部上皮 SP 細胞、NSP 細胞の遺伝子発現プロファイルの作成と遺伝子発現量比 較、Gene ontology (GO) 解析 ... 53
5-2-4.定量的遺伝子発現解析 ... 54
5-2-5.シングルセルqRT-PCR解析 ... 54
5-2-6.輪部上皮SP 細胞を用いた低酸素培養 ... 54
5-3.実験結果、考察 ... 55
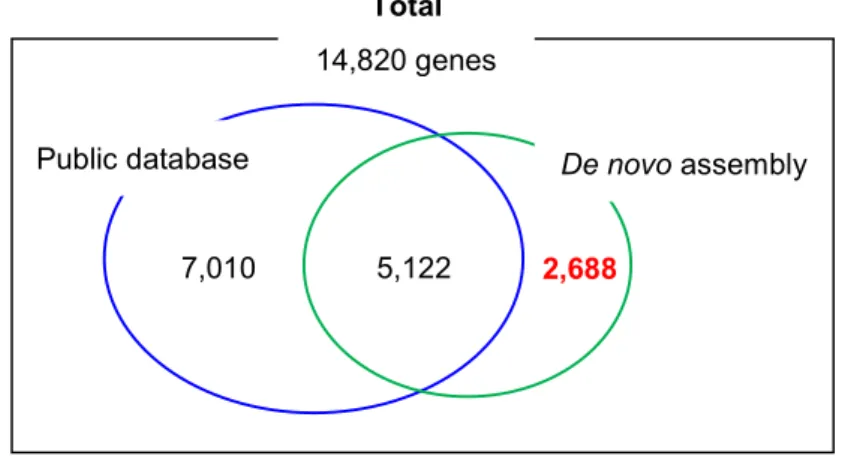
5-3-1.Public database (NCBI) と de novo transcriptome assembly を用いた遺伝子発 現プロファイルの作成 ... 55
5-3-3.DAVID Bioinformatics Resources 6.7 を用いたGene ontology 解析による輪部上 皮SP 細胞の表現型解明 ... 57
5-3-3.血管内皮細胞マーカーにおける定量的遺伝子発現解析 ... 64
5-3-4.シングルセルqRT-PCR 解析 ... 67
5-4.総括 ... 74
第6章 多臓器間比較におけるVcam1発現細胞の解析 6-1.序章 ... 75
6-2.材料と方法 ... 76
6-3.実験結果、考察 ... 76
6-4.総括 ... 79
第7章 総括と今後の展望 7-1.本研究の総括 ... 81
7-2.結語、展望 ... 87
謝辞 ... 89
参考文献 ... 91
研究業績 ... 104
【略語一覧】
ABC Avidin-biotin complex
Abcb1 ATP-binding cassette subfamily B member 1 Abcg2 ATP binding cassette subfamily G member 2 Bmi1 BMI1 proto-oncogene, polycomb ring finger BrdU 5-bromo-20-deoxyuridine
Cdh1 E-cadherin, Cadherin 1
(E-
カドヘリン)Cdh5 Cadherin 5, VE-cadherin
(VE-
カドヘリン)Emcn Endomucin
EMT Epithelial-mesenchymal transition
(上皮間葉移行)ES cells Embryonic stem cells
(胚性幹細胞)Esam Endothelial cell-selective adhesion molecule
GO Gene ontology
HEPES 4-(2-HydroxyEthyl)-1-PiperazineEthaneSulfonic acid HIF1
αHypoxia-inducible factor 1-alpha
iPS cells Induced pluripotent stem cells
(人工多能性幹細胞)K12 Keratin 12
(ケラチン12
)K13 Keratin 13
(ケラチン13
)K14 Keratin 14
(ケラチン14
)K19 Keratin 19
(ケラチン19
)Krt 15 Keratin 15
(ケラチン15
)MEndoT Mesenchymal-endothelial transition MET Mesenchymal-Epithelial transition
Muc5 Mucin 5AC
(ムチン5AC
)NGS Next generation sequencer
(次世代シークエンサ)NSP non-SP
PBS Phosphate buffered saline
(リン酸緩衝生理食塩水)Pcdh12 Protocadherin 12, VE-cadherin-2
Selp Selectin P
SP Side population
Stab1 Stabilin 1
Tek TEK tyrosine kinase, endothelial, TIE2 TGF-
βTransforming growth factor-β
TPS-A Triptostatin-A
Vcam1 Vascular cell adhesion molecule 1
【図表一覧】
図
1-1.
上皮組織の構造とターンオーバー図
1-2.
眼組織の構造図
2-1.
角膜上皮欠損モデルの作製図
2-2.
角膜上皮欠損モデルの経時的観察図
3-1. 4
週角膜上皮欠損モデルにおける組織学的解析図
3-2. 24
週角膜上皮欠損モデルにおける組織学的解析図
3-3. 48
週角膜上皮欠損モデルにおける組織学的解析図
3-4.
ウサギ角膜、輪部、結膜組織におけるIV
型コラーゲンα鎖の分布図
3-5.
モデル眼表面上における基底膜構成成分IV
型コラーゲンα5
鎖の分布図
3-6.
基底膜構成成分IV
型コラーゲンα1, 2
鎖の分布とその変化図
3-7.
正常輪部組織と48
週角膜上皮欠損モデル境界部の比較図
3-8.
正常眼組織におけるビメンチンの発現部位図
3-9.
角膜上皮欠損モデル眼組織におけるビメンチンの発現部位図
3-10.
上皮細胞の分化・未分化とEMT
・MET
の関連図
4-1. SP
細胞、NSP
細胞におけるHoechst 33342
色素の挙動 図4-2. SP, NSP
細胞分取方法図
4-3. Hoechst 33342
染色濃度決定図
4-4. Hoechst 33342
濃度2.5 µg/ml
の染色結果図
4-5.
角膜上皮SP
細胞、NSP
細胞の形態図
4-6.
増幅産物の電気泳動図
4-7. SP
細胞とNSP
細胞におけるAbcg2
の遺伝子発現比較図
5-1. NCBI genome database (Oryctolagus cuniculus)
を用いた遺伝子発現量 比較図
5-2.
De novo transcriptome assembly を用いた遺伝子発現量比較図
5-3. NCBI genome database
とDe novo transcriptome assembly の統合に よる新規参照配列の構築図
5-4.
遺伝子発現プロファイルの性状確認図
5-5.
発現量に10
倍以上差がある遺伝子集団を用いたヒートマップ図
5-6
. 上皮細胞マーカーにおける定量的遺伝子発現解析図
5-7
. 血管内皮細胞マーカーにおける定量的遺伝子発現解析図
5-8
. 血管内皮細胞/
未分化細胞マーカーにおける定量的遺伝子発現解析図
5-9
. 血管内皮細胞マーカー、未分化細胞マーカー、間葉系細胞マーカーにおけるシングルセル解析
図
5-10. ATP-binding cassette sub-family B (Abcb1)
の遺伝子発現解析図
5-11.
低酸素誘導因子Hypoxia-inducible factor-1a (HIF-1a)
の定量的遺伝子発 現解析図
5-12.
輪部上皮SP
細胞の低酸素培養と免疫組織化学的解析図
6-1.
輪部組織と口腔粘膜組織におけるVcam1
陽性細胞の局在図
6-2.
口腔粘膜上皮組織におけるVcam1
陽性細胞の解析図
6-3.
小腸上皮組織におけるVcam1
陽性細胞の解析図
7-1.
上皮細胞-
間葉系細胞-
血管内皮細胞間相互作用(既往研究)図
7-2
. 上皮細胞-
間葉系細胞-
血管内皮細胞間相互作用(仮説)表
1-1.
上皮幹細胞マーカーの探索研究表
1-2.
上皮幹細胞の評価法と上皮幹細胞マーカーの探索研究表
3-1. 1
次抗体希釈濃度表
4-1
. 多臓器におけるSP
細胞の存在頻度表
4-2
.SMARTer
を用いたサンプルcDNA
の増幅表
5-1.
輪部上皮SP
細胞で発現量が10
倍以上低い遺伝子群におけるGO
解析表
5-2.
輪部上皮SP
細胞で発現量が10
倍以上高い遺伝子群におけるGO
解析表
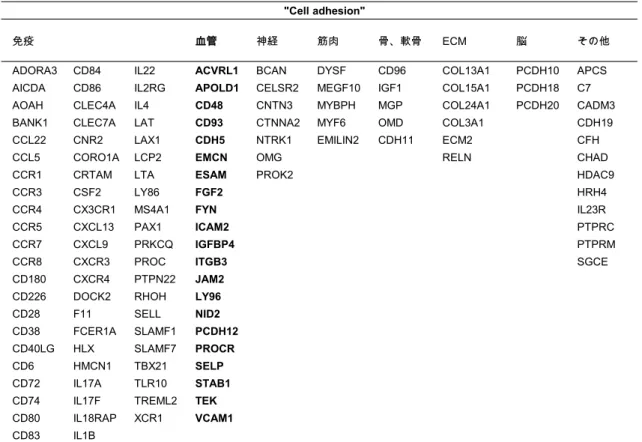
5-3.
“Cell adhesion
”に含まれる遺伝子➨㸯❶
⥴ゝ
㸬ୖ⓶⤌⧊ࡢศ㢮ᵓ㐀
ୖ⓶⤌⧊ࡣ༢ᒙୖ⓶ࠊከิୖ⓶ࠊ㔜ᒙୖ⓶ࡢ㸱ࡘศ㢮ࡉࢀࡿ㸦⾲ 㸧ࠋ༢ ᒙୖ⓶ࡣ୍ᒙࡢ⣽⬊ࡀᆒ୍୪ࡪ⤌⧊࡛࠶ࡾࠊከิୖ⓶ࡣᰕ≧ࡢ⣽⬊ࡀ୍ᒙ
୪ࡪࡀࠊ⫼ࡢ␗࡞ࡗࡓ⣽⬊ࡀΰᅾࡋࠊ᰾ࡢ⨨ࡶ␗࡞ࡿࠋ㔜ᒙᡥᖹୖ⓶ࡣ
ᩘᒙࡢ⣽⬊ࡀ㔜࡞ࡗࡓୖ⓶࡛࠶ࡿ㸦Ώࡽ 㸧ࠋ㔜ᒙᡥᖹୖ⓶ࡣ⓶ࡸゅ⭷ࠊ
ཱྀ⭍⢓⭷࡞య⾲Ꮡᅾࡍࡿୖ⓶⤌⧊ከࡃࡳࡽࢀࡿࠋ⣽⬊ྠኈࡀᐦ᥋⤖ྜ
ࡋࡓᵓ㐀㸦ᐦ᥋⤖ྜࠊࢱࢺࢪࣕࣥࢡࢩࣙࣥ㸧ࢆࡾࠊయእࡽ␗≀ࡢධࢆ
㜵ࡄࡓࡵࡢࣂࣜᶵ⬟ࢆᯝࡓࡍ࠸࠺㔜せ࡞ᙺࢆᢸࡗ࡚࠸ࡿࠋࡲࡓࠊ⣽⬊ࡀ
࠸ࡃࡘࡢᒙࢆ࡞ࡋࠊ✚ࡳ㔜࡞ࡿᵓ㐀㸦㔜ᒙ㸧ࢆࡿࡀࠊ௦ㅰࡼࡾᖖ⣽
⬊ࡀධࢀ᭰ࢃࡿ㸦ࢱ࣮࣮ࣥ࢜ࣂ࣮㸧ࠋ᭱⾲ᒙࡢ⤊ᮎศ⣽⬊ࡣࡀࢀⴠࡕࡿࡀࠊ ᇶᗏᒙᏑᅾࡍࡿୖ⓶ᖿ⣽⬊ࡀศࡋࠊ๓㥑⣽⬊ࡸศ⣽⬊ࢆୖᒙ౪⤥ࡍ
ࡿࠋࡇࡢࡼ࠺ࠊ⬇Ⓨ⏕ࡢࡳ࡞ࡽࡎᡂయࡢ⤌⧊ෆ࡛ࡶศࡋࠊᜏᖖᛶࡢ⥔ᣢ
⾲
⾲ ୖୖ⓶⤌⧊䛾ศ㢮㻌
䜘䛟䜟䛛䜛❧య⤌⧊Ꮫ㻌 䠄Ώ௰୕䚸ᐑ⃝㑻㻌䠅䜘䜚ᘬ⏝䞉ᨵኚ㻌
1
ࢆᢸ࠺ୖ⓶ᖿ⣽⬊ࡀᏑᅾࡍࡿ㸦ᅗ 㸧ࠋ
㸬║⤌⧊ࡢᵓ㐀ࠊᝈ
ゅ⭷⤌⧊ࡣ┤ᚄ⣙
PPࠊཌࡉ⣙ ȝP
ࡢ⤌⧊࡛࠶ࡾࠊ⾲ᒙࡽゅ⭷ୖ⓶ ᒙࠊ,ᆺࢥ࣮ࣛࢤࣥࢆ㇏ᐩྵࡴゅ⭷ᐇ㉁ᒙࠊゅ⭷ෆࡢỈศࢆㄪ⠇ࡋゅ⭷ࡢ㏱᫂ᛶࢆ⥔ᣢࡍࡿゅ⭷ෆ⓶ᒙࡢ㸱ᒙࡽᵓᡂࡉࢀ࡚࠸ࡿ ┿㘠ࡽ ࠋゅ⭷ୖ⓶
⤌⧊࡛ࡣ௦ㅰࡼࡾ⣽⬊ࡀ⨨ࡁࢃࡾࠊ⾲ᒙ⣽⬊ࡽ⬺ⴠࡍࡿࡀࠊᇶᗏ⣽⬊ࡀ ศࡍࡿࡇ࡛⣽⬊ࡀ⿵ሸࡉࢀࡿࠋゅ⭷ୖ⓶ᇶᗏ⣽⬊ࡣࠊゅ⭷⤌⧊⤖⭷⤌⧊
ࡢቃ⏺㒊࡛࠶ࡿ㍯㒊⤌⧊ࡢᇶᗏୖ⓶ᒙᏑᅾࡍࡿゅ⭷ୖ⓶ᖿ⣽⬊ࡽ౪⤥ࡉࢀ
ࡿࡇࡼࡗ࡚ࠊゅ⭷⾲㠃ࡢᜏᖖᛶࡀ⥔ᣢࡉࢀ࡚࠸ࡿ㸦ᅗ 㸧ࠋゅ⭷ୖ⓶⤌⧊
ࡀയᐖࢆཷࡅࡿ㍯㒊⤌⧊Ꮡᅾࡍࡿゅ⭷ୖ⓶ᖿ๓㥑⣽⬊ࡀศࡋࠊศୖ⓶
⣽⬊ࢆ౪⤥ࡍࡿࡇ࡛⤌⧊ࢆಟࡍࡿ &RWVDUHOLVHWDOࠋ
ᅗ
ᅗ ୖୖ⓶⤌⧊䛾ᵓ㐀䛸䝍䞊䞁䜸䞊䝞䞊
ᇶᗏ⭷
ᇶᗏୖ⓶⣽⬊
⾲ᒙ⣽⬊
௦ㅰ䛻䜘䜚䛜䜜ⴠ䛱䜛
┒䜣䛻ศ䛩䜛
2
基底細胞はヘミデスモソームを介して基底膜のラミニンなどを含むラミナルシ ダ
(Lamina lucida)
に接着し、さらにそれがIV
型コラーゲンを含むラミナデン サ(Lamina densa)
に結合している(
真鍋ら, 2003)
。輪部基底層に存在する角膜 上皮幹細胞は直下に存在する基底膜に接着し、増殖や分化が制御されていると 考えられている(Schlotzer-Schrehardt et al., 2007)
。しかし、アルカリ腐蝕、酸 腐食、薬剤毒性などの外傷や、スティーブン-
ジョンソン症候群や眼類天疱瘡な どの疾患により輪部組織が障害を受けると、角膜上皮幹細胞が失われ、角膜上 皮幹細胞疲弊症となる。スティーブン-
ジョンソン症候群とは発熱を伴う急性の 上皮疾患であり、全身の皮膚に特徴的は多形滲出性紅斑を生じる。眼症状とし ては結膜において炎症が起きることや角膜組織においても血管新生が生じ、眼 表面全体が乾燥したような状態となる(
真鍋ら, 2003)
。また眼類天疱症とは慢 性の瘢痕性水疱性疾患であり、主として結膜・口腔・咽頭に症状がみられる。角膜上皮欠損も頻発し、角膜周辺部において血管新生が生じ、最終的には高度 の眼球乾燥状態になり角膜表面は角化する
(
真鍋ら, 2003)
。これらの外傷や疾 患は輪部組織に存在する角膜上皮幹細胞が傷害を受け、無血管組織である角膜 表面は血管を有する結膜組織に被覆されることにより角膜の混濁や視力の低下図 1-2. 眼組織の構造 a) 眼組織の横断面 b) 前眼部の横断面を示している。
角膜
輪部 結膜
a b
3
が誘発される。このような角膜上皮幹細胞疲弊症に対して既存の治療法である 角膜移植が適用されているが、拒絶反応が生じ予後は不良である。またドナー 不足も深刻な問題となっている。そこでこれらの疾患に対して、自己由来の幹
/
前駆細胞を用いて根本的治療を行う再生医療に注目が集まっている。1-3 .幹細胞研究
幹細胞は自身と同じ細胞を産生する能力(自己複製能)と多様な細胞に分化 する能力(多分化能)を併せ持つ未分化な細胞であると定義される。幹細胞に は万能性を有する胚性幹細胞
(Embryonic stem cells : ES cells)
や多能性を有す る人工多能性幹細胞(Induced pluripotent stem cells : iPS cells)
と、成体内の複 数の組織に存在し多分化能を有する組織幹細胞(Somatic stem cells)
に大別さ れる。組織幹細胞は成体内のあらゆる組織に存在し、未分化性を保ちながら分 裂して、前駆細胞や分化細胞を産生することで組織の恒常性を維持すると考え られている(Knoblich, 2008)
。現在報告されている組織幹細胞は、主に造血幹細 胞(Nakauchi et al., 1999)
、骨髄由来間葉系幹細胞(Mabuchi et al., 2013)
、脂肪 由来間葉系幹細胞(Crisan et al., 2008)
、神経幹細胞(Suh et al., 2007)
、肝臓の 幹細胞(Wang et al., 2015)
、また上皮幹細胞においては毛包幹細胞(Blanpain et al., 2004; Snippert et al., 2010)
、小腸幹細胞(Barker et al., 2007; Holmberg et al.,
2006)
がある。特に本研究で用いた上皮組織においては、代謝により表層から剥がれ落ちる細胞を基底層に存在する上皮幹細胞が補い組織の恒常性が維持さ れている。上皮幹細胞はいまだ同定されていない組織幹細胞の一つであり、幹 細胞の維持機構やニッシェの解明も未解明のままであるが、これまでに様々な 取り組みがなされている。
4
幹細胞性を評価する方法としては、多組織の幹細胞と比較して基礎研究・臨 床応用ともに進められている造血幹細胞研究において新たな方法が確立され、
その方法を上皮幹細胞研究へと適用することで研究が遂行されてきた。
1967
年 には放射線照射マウスに骨髄細胞を移植し、脾臓に産生されるコロニーを計数 し、移植細胞の増殖能をin vivo
の系で評価する方法が確立されている(Till and McCulloch, 2012)
。さらに、1977
年にはチミジンのアナログである5-bromo-20-deoxyuridine (BrdU)
の投与により、in vivo
での細胞増殖の有無や 増殖速度の評価が可能となった(Schneider et al., 1977)
。BrdU
は細胞分裂の際 にチミジンの代用として新しく合成されるDNA
鎖に取り込まれる特徴を有す ることで、DNA
鎖を標識することができる。この手法を用いることで組織内で の幹細胞の局在部位の解明が可能になった。本手法を用いて1985
年には放射線 照射マウスに移植した細胞のうち、膵臓内でコロニーを形成する細胞と定義さ れたマウス造血幹/
前駆細胞(Colony forming unit in the spleen; CFU-S
)は、増 殖をしない細胞集団と増殖が極めて遅い細胞集団の少なくとも2
つの細胞集団 に分けられることが証明され(Pietrzyk et al., 1985)
、幹細胞の自己複製能を評 価する方法として考案された。これらの造血幹細胞研究をもとに、上皮幹細胞 の局在も研究されている(Bickenbach, 1981)
。Bickenbach
らは、マウスに対して
BrdU
投与後、1
時間では約90
% の上皮基底細胞の核がラベルされているのに対し、
10
日後の上皮基底細胞においては減少し、基底上層の細胞が標識され ることを明らかにした。さらに72
日後には大部分の細胞において標識が消失し、約
1%
の上皮基底細胞において強く発現することを明らかにした。この細胞は 分裂速度が遅いことから“ Slow cycling cells
”と名付けられ、幹細胞と類似した 特徴を持つことが報告されている(Braun and Watt, 2004)
。しかし、この方法の5
欠点として、
BrdU
投与時に分裂している細胞のみを標識できることである。前 述の通り幹細胞は分裂速度が遅い細胞であることが知られており、BrdU
投与時 にまさに分裂している幹細胞は極めて少ないことが予想される。したがって、実際には生体内に存在する全幹細胞を標識することは困難であると考えられる。
この
BrdU
を用いたin vivo
での評価法の課題をもとに、in vitro
での増殖活性 により短期間で幹細胞の自己複製能を評価する方法が考案された(Barrandon and Green, 1987)
。1987
年にBarrandon
らはヒト表皮細胞を培養し、6
日後 に継代(Secondary culture)
を行うと、異なる3
種類のコロニーが形成される ことを明らかにした。コロニーのサイズが一番大きいものはホロクローン(Holoclones)
と定義され、“ Holo = entire”
を意味し、コロニー形成細胞のすべ てが増殖速度の高い細胞により構成されているコロニーを意味している。逆に、ほとんど増殖しない細胞により構成されているコロニーはパラクローン
(Paraclones)
と定義された。“ Para = beyond ”
を意味し、完全に増殖の制限さ れた細胞であり小さなコロニーを形成する集団であることが知られている。ま た、その中間に属するコロニーはメロクローン(Meroclones; mero = partial)
で あり、増殖能の高い細胞と低い細胞の2
種類の細胞が混在しているコロニーで あることが定義されている(Barrandon and Green, 1987)
。増殖能が高い幹/
前駆 細胞様の性質を持つ細胞は大きなコロニーすなわちホロクローンを形成するこ とが示唆され、上皮細胞の自己複製能を評価する新たな方法が提唱された。こ のコロニー形成能を評価法として用いた検討が行われ、1995
年Jones
らはヒト 表皮組織においてインテグリンβ1
を発現する細胞は高い増殖能を有することを 明らかにしている(Jones et al., 1995)
。インテグリンスーパーファミリーは細 胞接着因子の一つであり、細胞外マトリックスや細胞表面抗原と結合すること6
で細胞の分化・増殖および器官形成の調節を行うと考えられている。インテグ リンはα鎖とβ鎖のヘテロダイマーを形成し、細胞種や細胞の状況により同じ 基質に対してもレセプターとして使用されるインテグリンの種類が異なり、周 囲の環境に応じて様々な情報を細胞内へと伝達する役目を担う重要な因子であ
る
(
小出et al., 2000)
。また、2000
年Kaur
らはヒト表皮組織においてインテグリン
α6
が上皮基底細胞に発現し、増殖能が高いことが示された(Kaur and Li, 2000)
。さらにヒト表皮細胞において、CD90
陽性細胞がin vitro
またin vivo
で のコロニー形成能が高いことが示されている(Nakamura et al., 2006)
。これら は特定の培養環境下での増殖能を基準に分化、未分化を評価したものであるが、幹細胞の定義である多分化能を評価することができない。したがって決定的な 上皮幹細胞マーカーを同定するためには不十分であり、さらなる検討が必要で あると考えられる。そこで、遺伝子改変マウスをもちいた上皮幹細胞研究が行 われた。⊿
Np63
欠損マウスでは、通常の重層扁平上皮の形成は困難であり、単 層の上皮が形成されることからp63
は上皮幹細胞の維持や分化、上皮組織の重 層化に重要であることが報告されている(Koster et al., 2005; Mills et al., 1999;
Yang et al., 1999)
(表1-2
)。角膜上皮幹細胞マーカーの研究も行われており、上皮細胞に発現する中間系フィラメントであるケラチン
14 (Keratin 14: K14)
やK19
の発現が、幹/
前駆細胞が局在する輪部上皮組織に発現することが報告さ れている(Barnard et al., 2001; Wang et al., 2003)
。また、2001
年Pellegrini
ら はヒト表皮組織と角膜上皮組織の上皮前駆細胞は⊿Np63
を発現し、これが増殖 に関与する因子であることを報告した(Pellegrini et al., 2001)
。さらに、間葉系 細胞マーカーであるビメンチン(Lauweryns et al., 1993)
や神経堤幹細胞マー カーであるp75 (Di Girolamo et al., 2008)
の発現もヒト輪部上皮組織に発現す7
ることが報告されている。このように、輪部上皮細胞には上皮細胞特異的なケ ラチンや間葉系細胞マーカー、神経堤細胞マーカーなど多様な表現型を示すこ とが報告されている。しかしそれらは組織学的解析を用いて輪部上皮基底細胞 に発現することのみ示され詳細が不明瞭であることから、角膜上皮幹細胞の表 現型解明においては未だ議論の余地があるといえる。上皮幹細胞マーカーの同 定のために上記の課題を克服するため、新たに考案されたのが細胞系譜解析
(Lineage-tracing)
を用いた解析である。特定の細胞が細胞分裂を繰り返した後、産生された細胞を
in vivo
内で追跡することができる。この手法を用いた検討に より、BMI1 proto-oncogene, polycomb ring finger (Bmi1)
遺伝子を発現する細胞 が上皮組織を構成細胞の起源であることが明らかになった。さらに上皮組織修 復にも寄与することが示され、Bmi1
は上皮幹細胞マーカーであることが強く示 された(Tanaka et al., 2013)
。Bmi1
は転写因子であり、セルソーターを用いた 細胞分離の際の標的分子として用いることは不可能であるが、Bmi1
陽性の上皮 細胞を解析することにより、上皮幹細胞の正体を解明できると期待される(表1-2
)。8
㸬ୖ⓶⤌⧊⏕་⒪ࡢⓎᒎ
ᇶ♏◊✲ㄢ㢟ࡀṧࡿ୍᪉࡛ࠊୖ⓶⏕་⒪ࡣࡢ⤌⧊ẚ㍑ࡋ࡚㎿㏿ᐇ
ࡉࢀ࡚࠸ࡿࠋࡑࡢ⌮⏤ࡢ୍ࡘࡣࠊᚰ➽⣽⬊ࠊ⫢⣽⬊➼ẚ㍑ࡋ࡚᪩ᮇࡼࡾ
LQ YLWUR
࡛ࡢ⣽⬊ᇵ㣴ἲࡀ☜❧ࡉࢀࡓࡇࡼࡾ⛣᳜∦ࡢస〇ࡀྍ⬟࡞ࡾࠊ⮫ᗋ ᛂ⏝ࡀ㐍ࡵࡽࢀ࡚࠸ࡿࠋᖺ*UHHQ
ࡽࡣ࣐࢘ࢫ⧄⥔ⱆ⣽⬊ 7ࢆࣇ࣮ࢲ࣮⣽⬊ࡋ࡚ඹᇵ㣴ࡍࡿࡇ࡛ࠊึࡵ࡚⛣᳜ྍ⬟࡞ᇵ㣴⾲⓶⣽⬊ࡢస〇
ᡂຌࡋࡓ *UHHQHWDOࠋࡇࢀࢆⓎ➃ࡋ࡚ࠊᖺ
3HOOHJULQL
ࡽࡣ㔜 ᗘ⇕യᝈ⪅ᑐࡋ࡚ᇵ㣴⾲⓶⣽⬊⛣᳜ࢆࡋࠊ⛣୍᳜ᖺᚋ࠾࠸࡚ࡶᴟࡵ࡚Ⰻ ዲ࡞ணᚋࢆᚓ࡚࠸ࡿ 3HOOHJULQLHWDOࠋࡲࡓᮏ◊✲ࡢᑐ㇟ࡋࡓゅ⭷⤌⧊࠾࠸࡚ࡣࠊᖺᮾிዪᏊ་⛉Ꮫඛ➃⏕་⛉Ꮫ◊✲ᡤ㜰Ꮫ║⛉ᩍ ᐊࡢඹྠ◊✲ࡼࡾࠊᇵ㣴ୖ⓶⣽⬊ࢩ࣮ࢺࢆ⏝࠸ࡓゅ⭷ୖ⓶⏕ࡀ⪃ࡉࢀࡓࠋ
ࣝ࢝ࣜ⭉⼃ࠊ㓟⭉㣗ࠊ⸆ẘᛶ࡞ࡢ㔜⠜࡞እയࠊࡲࡓࢫࢸ࣮ࣈࣥࢪࣙࣥ
ࢯࣥೃ⩌ࡸ║㢮ኳ⑁⒔࡞ࡢᝈࡼࡾᖿ⣽⬊ࡀᏑᅾࡍࡿ㍯㒊⤌⧊㞀ᐖࢆ
ཷࡅࡓゅ⭷ୖ⓶ᖿ⣽⬊⑂ᘢᝈ⪅ᑐࡋ࡚ࠊ⮬ᕫ⏤᮶ᇵ㣴⣽⬊ࢆ⛣᳜ࡍࡿࡇ
࡛ゅ⭷ୖ⓶ࡀᘓࡉࢀ࡚࠸ࡿ 1LVKLGDHWDOࠋ∦║ᛶᝈࡢᝈ⪅ᑐࡋ࡚
⾲
⾲ ୖୖ⓶ᖿ⣽⬊䛾ホ౯ἲ䛸ୖ⓶ᖿ⣽⬊䝬䞊䜹䞊䛾᥈⣴◊✲㻌
9
は、健常眼の角膜上皮幹
/
前駆細胞が存在する輪部上皮組織から細胞を採取・単 離した後、培養角膜上皮細胞シートを作製し移植する。しかし、スティーブン-
ジョンソン症候群や眼類天疱瘡などの疾患は両眼性疾患であることが多く、自 身の輪部上皮組織から角膜上皮幹/
前駆細胞を得ることは不可能である。そこで、大阪大学医学部眼科学教室との共同研究により、角膜上皮細胞の代用として口 腔粘膜上皮細胞を細胞ソースに用いて角膜上皮組織を再建する治療法が考案さ れた。患者の口腔粘膜上皮細胞を採取し、温度応答性培養皿を用いて培養する。
温度応答性培養皿を用いることによりタンパク質分解酵素を使用することなく、
温度を
20
℃に下げるという非侵襲的な方法で細胞を回収できるため、基底膜成 分を保持したシート状の細胞を患者に移植できる。培養上皮細胞シート移植後 の角膜表面は移植1
年後でも角膜の透明性が維持されており、視力の回復が認 められている(Nishida et al., 2004)
。角膜上皮組織はターンオーバーを繰り返し、2
週間で細胞が入れ替わることが知られているため、患者の体内には移植された 幹細胞が生着し、上皮組織の恒常性を維持すると推測される。しかし幹細胞マ ーカーが同定されておらず、モデル動物を用いた幹細胞の生体内挙動やニッシ ェの構築過程が未解明のままとなっている。そこで本研究では角膜組織に着目 し、角膜上皮幹/
前駆細胞や分化角膜上皮細胞、基底膜構成分子、結合組織内の 線維芽細胞、血管内皮細胞などの側面から多角的に検討することで角膜上皮幹 細胞研究を遂行した。1-4 .本研究のねらい
本研究では角膜上皮再生医療への貢献を目指して研究を遂行し、
in vitro
の検 討に終始することなく、生体内での観察結果をもとに仮説を立て、実証できる10
よう取り組んだ。第
2
章、第3
章ではin vivo
実験により組織修復過程における 分化上皮細胞や幹/
前駆細胞の挙動や幹細胞ニッシェであると考えられている基 底膜の再編成という側面から解析した。また第4
章、第5
章、第6
章ではそこ で得られた仮設をもとに上皮幹細胞の正体の解明を目指した。11
第2章
角膜上皮幹細胞疲弊症モデルの 作製とその評価
2-1 .序章
ウサギ眼表面は
3
~5
層の重層扁平上皮で構成されている(Thoft and Friend, 1977; Wei et al., 1996; Wei et al., 1993)
。基底層に存在する未分化細胞は増殖を 通じて分化しながら表層へと移動し、最終的には脱落する。この代謝、すなわ ちターンオーバーは約2
週間かけて行われる。角膜と結膜の境界部に存在する 輪部組織には角膜上皮幹細胞が局在しており、分裂し、分化細胞を角膜中央部 へと供給することにより、角膜上皮の恒常性が維持される(Buck, 1985; Thoft
and Friend, 1983)
。重篤な外傷や、スティーブン-
ジョンソン症候群や眼類天疱瘡のような疾患により、この角膜上皮幹細胞が傷害を受けると、結膜組織が角 膜表面を被覆し、血管新生や角膜の混濁が生じて視力が低下する
(Ti et al., 2002; Tsubota et al., 1999)
。このような病理学的所見は角膜上皮幹細胞疲弊症と 呼ばれる。本疾患のモデル動物として研究されてきたのが、ウサギ角膜上皮幹 細胞疲弊症モデル(Rabbit limbal epithelial stem cell deficiency model; LSCD model)
である。研究対象とする動物種を選定する際、以下の事項を考慮する必要があった。
ヒトの角膜組織は、アメリカアイバンクより購入可能ではあるが、とても希少 であり入手が困難であることや
in vivo
での検討が不可能であることが大きな欠 点となっている。マウスを対象にする場合、生物資源が豊富であり、遺伝子改12
変マウスを用いた解析や特異的抗体を用いて細胞を単離、培養することができ、
とても有用であると考えられる
(Mine et al., 2005; Vauclair et al., 2007; Yoshida
et al., 2006b)
。しかしこれは注目する因子が特定されているときのみ使用可能であり、外科的処置を施す検討や多くの細胞を必要とする場合、眼組織が極めて 小さいため実施が困難となる。本研究は
in vivo
での上皮細胞とそれを取り巻く 微小環境であるニッシェ構築過程の解析や、特定の細胞を単離した後、遺伝子 発現プロファイルを作成して上皮幹/
前駆細胞マーカーを探索するといった、特 定の遺伝子やタンパクを決定するための研究である。したがってヒト角膜組織 や遺伝子改変マウスを用いることは不可能であると考えた。そこで本研究では ウサギ角膜上皮欠損モデルを作製し、解析した。分化細胞が局在する角膜上皮 と幹/
前駆細胞が存在する輪部上皮を外科的に切除したのち、さらにヘプタノー ル処理を施すことで化学的に細胞を除去して作製した(Ti et al., 2002)
。このモ デル眼表面では、角膜の混濁や結膜組織の浸潤、血管新生が観察され、ヒト角 膜上皮幹細胞疲弊症と類似した症状が現れることが示されている(Ti et al.,
2002)
。本研究では、ウサギ角膜上皮幹細胞疲弊症モデル(角膜上皮欠損モデル)を作製し、修復の過程を組織学的に解析した。まず本章では、眼表面に上皮組 織を欠損させ、そのモデルの評価を行った。
2-2 .実験材料および方法 2-2-1
.動物成体のオスの白色家兎(
New Zealand White Rabbit
、株式会社日本医科学動物 資材研究所、体重:約3 kg
)を飼育室に搬入後1
週間以上飼育し、実験に用い13
た。〔実験動物倫理委員会 審議済整理番号:
10-42
、11-122
、12-25
、13-99
、14-87
、15-43
〕2-2-2
.角膜上皮欠損モデル(Corneal epithelial deficiency model
)動物の 作製方法白色家兎(
New Zealand White Rabbit
、株式会社日本医科学動物資材研究所、体 重:約3 kg
)に対し、ケタラール0.4 ml
、ドミトール0.6 ml
の混合液を投与し 全身麻酔を行なった。麻酔下にあり、痛みを感じ得ない状態を保ち、眼脱臼し、眼瞼が閉じぬよう固定後、生理食塩水で眼球をよく洗浄した。その後、ゴルフ メスにより強膜に切開線を入れ、さらに角膜中央部に向かって上皮下の結合組 織を切開し、角膜実質に到達させた。そこから眼科用剪刀を用いて輪部上皮組 織および角膜上皮組織を
1
枚のシート状の組織片として切除し、眼球に外科的 損傷を与えた(図2-1
)。その後、ヘプタノールを含ませたガーゼで角膜表面を覆い、
5
分間静置するこ とで化学的損傷を与えた。反応終了後、ガーゼを取り除き、残ったヘプタノー ルを完全に除去するために生理食塩水で丁寧に洗浄した。最後に眼瞼をもとの 位置に戻してリンデロンとタリビット軟膏を塗布し、処置を終了した。また、図 2-1. 角膜上皮欠損モデルの作製
眼科用剪刀を用いて角膜および輪部上皮組織とその直下に存在する基底膜を切除し た様子を示している。
14
麻酔から覚醒しそうな場合は、適宜ケタラールとドミトールを
2
:3
の割合で混 合したものを約0.5 ml
再投与した。処置後1
週間は炎症を抑えるためにリンデ ロンとタリビットを1
日2
回投与した。また、経過観察として、処置後1, 2, 3,
4, 24, 48
週目に経過観察および蛍光色素(フルオレセイン)による染色を行った。処置後
4
週目、24
週目、48
週目にソムノペンチル(共立製薬株式会社)を用いて犠牲させ、組織学的解析に供した。
2-3 .実験結果、考察
角膜および輪部上皮組織を切除した後の経過観察の結果を図
2-2
に示した。角膜上皮組織は表層細胞、翼細胞、基底細胞から成り、最外層の表層細胞の頂 端部には密着結合が発達している。これは隣接細胞間を密につなぐことで水溶
1 週目 4 週目 24 週目 48 週目
正常組織
図 2-2. 角膜上皮欠損モデルの経時的観察
正常組織と角膜上皮欠損モデル作製後1週目、4週目、24週目、48週目の観察結 果を経時的に示している。上段が蛍光染色前、下段が蛍光染色後の様子である。上 皮細胞が存在する部位では、細胞-細胞間に密着結合(Tight junction)が発達し、蛍 光色素の沈着を防ぐが、上皮細胞が存在せず、角膜実質(結合組織)が眼表面上に 曝されている場合、色素の沈着が起き、緑色を呈する。Kameishi et al., 2015 より引 用・改変。
15
性分子に対する障壁となるため、外来物質に対する防御機能を有する。したが って蛍光色素を点眼すると、角膜実質が露出している部位は蛍光色素が浸透し 沈着するが、上皮組織が存在する場所は密着結合により色素の浸透が妨げられ て沈着しない
(Satake et al., 2008)
。本研究では処置から1
週間後には角膜実質 への蛍光色素の沈着が眼表面全体に観察された(図2-2
)。これに対し、4
週後 には角膜中央部にのみ蛍光色素が沈着していたことから、結膜側から角膜中央 部にかけて上皮組織が浸潤したことがわかった(図2-2
)。さらに、結膜側から 角膜中央部に向けて血管新生も観察され、結膜組織が角膜中央部へと浸潤した と考えられる。また約6
週後には上皮組織が角膜表面を完全に被覆し、24
週、48
週後も上皮組織は維持されていた。これらのモデル組織は正常組織と比較し て角膜が混濁し、血管新生も観察され、適切にモデルを作出できたと考えられ る。以後の上皮修復過程の解析に、本モデルを使用した。2-4 .総括
本研究で作出したモデルの眼表面において、結膜側から上皮細胞が浸潤し、
眼表面を被覆したこと、また角膜の混濁や血管新生も観察された。これは角膜 上皮幹細胞疲弊症患者の病態に類似しており、今後の解析に有用なモデルを作 出できたことが分かった。さらに、本研究では、ヒト眼組織と大きさや細胞の 性質が類似するウサギ眼組織を用いたが、これはヒトやマウス眼組織での研究 では実施不可能である傷害モデルの作製や
in vivo
での経時的な観察、組織学的 解析が可能となる点で適切な動物モデルであると考えられる。16
第3章
角膜上皮欠損モデルにおける 上皮細胞と基底膜の再形成
3-1 .序章
組織修復過程において、角膜上皮幹細胞は分裂して分化細胞を産生し、上皮 組織を再構築する。角膜上皮幹細胞は周囲の細胞や細胞外マトリクスなどの微 小環境(ニッシェ)との相互作用により制御されている
(Ihanamaki et al., 2004;
Li et al., 2007)
。また、体内で分泌された増殖因子や形態形成因子は特定の細胞外マトリクスへ結合し、隣接する細胞の増殖や分化に影響を及ぼすことが報告 されている
(Saharinen et al., 1999)
。さらに、基底膜の分子組成は多様性を示 し、組織により異なることが知られている。眼組織では、分化細胞が存在する 角膜組織と未分化な幹/
前駆細胞が存在する輪部組織において、基底膜構成成分 が異なることが報告されている(Schlotzer-Schrehardt et al., 2007)
。以上より、輪部組織特異的に存在する基底膜構成成分により、角膜上皮幹
/
前駆細胞の増殖 や分が制御されていると考えられる。本研究では、第2
章で確立したウサギ角 膜上皮欠損モデルを用いて、組織の修復過程を上皮細胞と基底膜構成成分に着 目して解析を行うこととした。特に、基底膜主要構成成分であるIV
型コラーゲ ンα鎖の分布を組織学的に解析することで、上皮細胞の表現型獲得と基底膜構 成成分の関連の解明を目指した。17
3-2 .実験材料および方法 3-2-1
.動物成体のオスの白色家兎(
New Zealand White Rabbit
、株式会社日本医科学動 物資材研究所、体重:約3 kg
)を飼育室に搬入後1
週間以上飼育し、実験に用 いた。〔実験動物倫理委員会 審議済整理番号:10-42
、11-122
、12-25
、13-99
、14-87
、15-43
〕3-2-2
.組織学的解析3-2-2-1.
凍結切片の作製方法クリオスタットを用いて
5 μm
に薄切し、MAS
コート付きスライドガラス(Micro
Slide Galass,
松浪硝子工業株式会社)に転写した。切片を室温で1
時間風乾させた。
3-2-2-3.
ヘマトキシリン・エオジン(HE)
染色3.2.2
の凍結切片の作製方法に従って作製した組織切片において、リン酸緩衝生理食塩水
(Ca, Mg
不含)(Phosphate buffered saline; PBS(-),
ナカライテスク)
で5
分間洗浄しコンパウンドを洗い流した。PBS(-)
をよく切り、ヘマトキシリ ン液(
マイヤー・ヘマトキシリン,
武蔵化学株式会社)
で2
分染色した。その後、水道水で洗浄、色出しを行った。次にエオジン液で
3
分間染色した後、水道水 で洗浄した。染色が完了した後、70
%、80
%、90
%、100
%エタノール(
和光純 薬株式会社)
の順に包埋切片をエタノールにつけて脱水、キシレン(
和光純薬株 式会社)
で透徹し封入した。18
3-2-2-4.
免疫組織化学的解析a)
酵素抗体法を用いた免疫組織化学的解析3.2.2
の凍結切片の作製方法に従って作製した組織切片においてPBS(-)
で5
分間洗浄しコンパウンドを洗い流した。内因性ペルオキシターゼを失活させるため ペルオシキダーゼ失活液
(Peroxidase Blocking Regent, Dako)
を滴下し、室温 で5
分間反応させた。PBS(-)
で5
分間洗浄した後、非特異吸着を防ぐために5
% ロバ血清含有PBS(-)
を滴下し、室温で1
時間ブロッキングを行った。次に1
% ロバ血清含有PBS(-)
で所定の濃度に希釈した1
次抗体を滴下し、4
℃で一晩反 応させた(表3-1
)。その後、PBS(-)
で10
分、計3
回洗浄し、1
% ロバ血清含 有PBS
で所定の濃度に希釈した2
次抗体を滴下し、室温で1
時間反応させた。このとき、発現の少ないタンパクについてはアビジン
-
ビオチン複合体(avidin-biotin complex : ABC)
を用いることにより検出の感度を高めた。ABC
法 を用いる場合、1
% ロバ血清含有PBS(-)
で所定の濃度に希釈したビオチン標識 された2
次抗体を滴下し、室温で1
時間反応させた後、PBS(-)
で10
分、計3
回洗浄した。続いてHRP
標識されたアビジンを室温で1
時間反応させた後、PBS(-)
で10
分、計3
回洗浄した。発色の際は発色基質(DAB, DAKO
)を添加 し、発色具合を見ながら1
~10
分反応させた。その後、核染色のためにヘマト キシリン液を用いて染色を行い、水道水で色出しを行った。洗浄・色出しを終 え、70
%、80
%、90
%、100
%エタノールの順にサンプルをエタノールにつけ て脱水を行い、最後にキシレンで透徹し封入を行った。また、抗ビメンチン抗体の染色においては切片を風乾・洗浄後、ホルムアル デヒド
(10%
ホルマリン液,
和光純薬株式会社)
に浸し、室温で10
分固定を行 った。またラット抗ヒトIV
型コラーゲンα鎖抗体を用いた染色においては切片19
を風乾させたのち、-
20
℃のアセトン(
和光純薬株式会社)
を用いて10
分固定 後、塩化カリウム-
塩酸(KCl-HCl, pH 1.5)
溶液で酸性処理を行うことで前処理 を行い、免疫染色に用いた。b)
蛍光色素を用いた二重染色3.2.2
の凍結切片の作製方法に従って作製した組織切片において、PBS(-)
で5
分間洗浄しコンパウンドを洗い流した。その後、
5
% ロバ血清含有PBS(-)
を滴 下し、室温で1
時間ブロッキングを行った。次に、1
% ロバ血清含有PBS(-)
で 所定の濃度に希釈した1
次抗体を滴下し、4
℃で一晩反応させた(表3-1
)。そ の後、PBS(-)
で10
分、計3
回洗浄し、1
% ロバ血清含有PBS
で所定の濃度に 希釈した2
次抗体を滴下し、室温で1
時間反応させた。抗体反応終了後、PBS(-)
で10
分、計3
回洗浄し、1
% ロバ血清含有PBS(-)
で所定の濃度に希釈した2 次抗体を滴下し、遮光しながら室温で1
時間反応させた。抗体反応終了後、PBS(-)
で10
分、計3
回洗浄した。同様に2
種類目の抗体においても上記の手順で染色 を行った。最後に、1
% ロバ血清含有PBS(-)
で所定の濃度に希釈したDAPI
を 滴下し、室温で10
分反応させ、核染色を行い、封入剤(Prolong Gold, Thermo Fisher Scientific
)を用いて封入した。20
㸬ᐇ㦂⤖ᯝࠊ⪃ᐹ
㸬ゅ⭷Ḟᦆࣔࢹࣝ║⾲㠃ࡢゎᯒ
ṇᖖゅ⭷ୖ⓶ࡢ┤ୗᏑᅾࡍࡿゅ⭷ᐇ㉁ࡣ↓⾑⟶⤌⧊࡛࠶ࡾࠊࢥ࣮ࣛࢤࣥࡀ つ๎ⓗ㓄ྥࡍࡿ࣓ࣛࣛᵓ㐀ࢆࡿࠋࡇࢀᑐࡋࠊ㍯㒊࣭⤖⭷⤖ྜ⤌⧊࠾࠸
࡚ࡣ⾑⟶ࡀ㇏ᐩ࡞⤌⧊࡛࠶ࡿࡇࡀ▱ࡽࢀ࡚࠸ࡿ 0DUVKDOO*(ࠋᮏ◊✲
࡛సฟࡋࡓゅ⭷ୖ⓶Ḟᦆࣔࢹࣝࡢゅ⭷ᐇ㉁࠾࠸࡚ࡣࠊ᪂⏕⾑⟶ࡀᏑᅾࡋࠊࢥ
࣮ࣛࢤࣥ⧄⥔ࡶࣛࣥࢲ࣒㓄ྥࡋ࡚࠸ࡿࡇࡀࢃࡗࡓ㸦ᅗ
D㸧ࠋࡇࢀࡽࡣ
ࢫࢸ࣮ࣈࣥࢪࣙࣥࢯࣥೃ⩌ᝈ⪅ࡢ║⤌⧊ぢࡽࢀࡿゅ⭷⾲㠃ࡢ⾑⟶᪂⏕㢮ఝࡋ࡚࠸ࡓ (VSDQDHWDOࠋḟච⤌⧊Ꮫⓗゎᯒ౪ࡋࡓ⤖ᯝࠊ
㐌ࣔࢹࣝ║⾲㠃࠾࠸࡚⤖⭷ୖ⓶⣽⬊࣐࣮࣮࡛࢝࠶ࡿࢣࣛࢳࣥ㸦.HUDWLQ.㸧ࡸ⤖⭷ᮼ⣽⬊࣐࣮࣮࡛࢝࠶ࡿ࣒ࢳࣥ $&
㸦0XFLQ$&0XF㸧ࡀⓎ⌧ࡋ࡚࠸ࡓ㸦ᅗ EF㸧ࠋࡲࡓࠊゅ⭷ୖ⓶⣽⬊࣐࣮࣮࡛࢝࠶ࡿࢣࣛࢳࣥ
㸦.㸧
ࡢⓎ⌧ࡣほᐹࡉࢀ࡞ࡗࡓ㸦ᅗ G㸧ࠋࡋࡓࡀࡗ࡚ࠊゅ⭷ࠊ㍯㒊ୖ⓶ษ㝖ᚋ⤖⭷⤌⧊ࡀゅ⭷୰ኸ㒊ᾐ₶ࡋࡓࡇࡀࢃࡗࡓ㸦ᾐ₶⤖⭷ୖ⓶㸧ࠋ
.DPHLVKLHWDO ㆤ䞷ዘ㟈⮘ᇭ㻌
嫷
嫷 㶰㶰㔦⇢゛摗䉒ㄵ㻌
21
次に
24
週モデル眼表面において組織学的解析に供したところ、角膜中央部に肥 厚した上皮層が観察された(図b. 3-2, black bar
)。この部位を免疫組織化学的解 析に供したところ、K13
は角膜周辺部で発現するのに対し、肥厚した上皮細胞 ではK13
の発現は観察されず(図3-2, b black bar
)、Muc5
を発現する結膜杯細 胞の局在も観察されなかった(図3-2, c, black bar
)。驚いたことに、モデル作 製時にK12
陽性細胞を除去したにも関わらず、この肥厚した上皮細胞は角膜上 皮細胞マーカーであるK12
を発現し、角膜上皮様細胞であることがわかった(図3-2, d, black bar
)。さらにこの細胞はK13
陽性の浸潤結膜上皮細胞に囲まれ、角膜上皮幹
/
前駆細胞が存在する輪部組織とは隔離されており、この角膜上皮様 細胞はおよそ2.5 mm
にわたって存在することがわかった。図 3-1. 4週角膜上皮欠損モデルにおける組織学的解析
角膜上皮欠損モデル作製後、4週の眼組織を採取し、解析を行った。(a) ヘマトキシリ ン・エオジン染色、また(b) 結膜上皮細胞マーカーであるケラチン13 (Keratin 13:
K13)、(c) 結膜上皮組織内に存在する杯細胞マーカーであるムチン5 (Mucin 5 :
Muc 5)、(d) 角膜上皮細胞マーカーであるK12 の免疫染色の結果を示している。ス
ケールバーは25 μm。Kameishi et al., 2015 引用・改変。
HE K13
K12 Muc5
a b
c d
22
48
週角膜欠損モデル眼表面上においても、K13
陽性の浸潤結膜上皮が存在した が(図3-3, b
左)、K13
を発現しない細胞も存在した(図3-3, b
右)。このK13
陰性細胞はK12
を発現することがわかり(図3-3, d
)、Muc5
陽性細胞も含まな いことが分かった(図3-3, c
)。したがって48
週角膜欠損モデル眼表面上にも 角膜上皮様細胞が存在することがわかり、およそ7.5 mm
にわたってK12
陽性 細胞が存在することがわかった。以上より、角膜上皮欠損モデル作出時に角膜 上皮また輪部上皮を切除したにもかかわらず、長期経過観察により角膜上皮様 細胞が再び出現することが分かった。図 3-2. 24週角膜上皮欠損モデルにおける組織学的解析
角膜上皮欠損モデル作製後、24週の眼組織を採取し、解析を行った。(a) ヘマ トキシリン・エオジン染色、また(b) 結膜上皮細胞マーカーであるケラチン13 (Keratin 13: K13)、(c) 結膜上皮組織内に存在する杯細胞マーカーであるムチン5 (Mucin 5 : Muc 5)、(d) 角膜上皮細胞マーカーであるK12 の免疫染色の結果を示 している。矢頭は境界部を、アスタリスクは血管を示している。また黒いバーは 肥厚した上皮を示している。スケールバーは25 μm。Kameishi et al., 2015 引 用・改変。
d b
c
*
K12 HE K13
Muc5 a
23
結膜上皮細胞は角膜上皮様細胞に分化転換することが報告されている
(Dua, 1998; Kinoshita et al., 1983)
。これは結膜上皮組織中に存在するムチンを発現す る結膜杯細胞の数が減少することにより示されている。また、これまでの結膜 上皮細胞の分化転換における既往研究は、角膜上皮細胞に発現する主要なケラ チンである64kD
のK3
を認識するモノクローナル抗体(AE5
)を用いた研究で あり、ウサギ角膜上皮欠損モデル眼表面の解析に使用されている。しかし、AE5
はK3
を発現しないはずの結膜上皮細胞とも交差反応性を持つことが報告され(Barbaro et al., 2010)
、結膜上皮細胞の分化転換においては議論の余地があるとされている。しかし近年、角膜上皮細胞の主要なケラチンタンパクであり、
K3
と ヘテロダイマーを形成するK12
に対するポリクローナル抗体が作出され、角膜 上皮特異的に認識し、角膜上皮と結膜上皮を識別できるようになっている(Boehlke et al., 2004; Kawasaki et al., 2006)
。本研究では角膜上皮細胞を特異的 に認識する抗K12
抗体を用い、正常眼組織において結膜上皮細胞に反応せず、図 3-3. 48週角膜上皮欠損モデルにおける組織学的解析
角膜上皮欠損モデル作製後、48週の眼組織を採取し、解析を行った。(a) ヘマトキ シリン・エオジン染色、また(b) 結膜上皮細胞マーカーであるケラチン13 (Keratin 13:
K13)、(c) 結膜上皮組織内に存在する杯細胞マーカーであるムチン5 (Mucin 5 :
Muc 5)、(d) 角膜上皮細胞マーカーであるK12 の免疫染色の結果を示している。矢
頭は境界部を、アスタリスクは血管を示している。スケールバーは25 μm。Kameishi et al., 2015 引用・改変。
a
d b
c
*
K12 K13 HE
Muc5
24
角膜上皮細胞のみ検出することを確認したのち、モデル眼表面の解析を進める ことで、角膜上皮様細胞の存在を明らかにした。
3-3-2
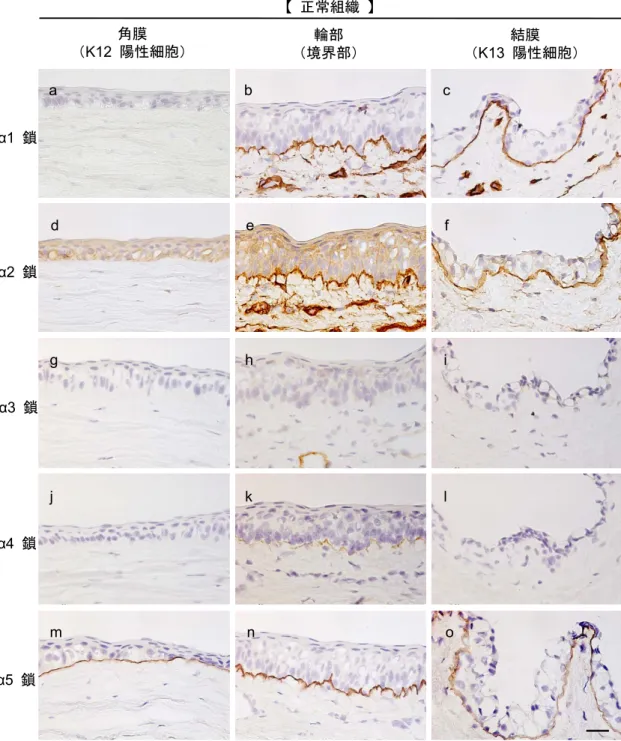
.上皮細胞の表現型と基底膜構成分子の関係IV
型コラーゲンは主要な基底膜構成分子であり、3
つのα
鎖が集合して三重ら せん構造をとる(Ninomiya et al., 1995; Thorner et al., 1996)
。α1-α1-α2(IV)
は 生体内で広く分布が確認されており、ヒト正常角膜組織には存在せず、輪部・結膜組織の基底膜に存在することが知られている。また、
α3-α4-α5(IV)
は角膜 組織特異的に局在し、α5-α5-α6(IV)
は角膜・輪部・結膜組織のすべてに分布す ることが分かっている(Guerriero et al., 2007; Schlotzer-Schrehardt et al.,
2007)
。このモノクローナル抗体はIV
型コラーゲンのC
末端領域を認識することが明らかになっており、
α1-5
鎖に対する抗体においてはウサギに交差性があ ることが示されている(Ninomiya et al., 1995)
。本研究ではIV
型コラーゲンα1
鎖、α2
鎖、α3
鎖、α4
鎖、α5
鎖に対するモノクローナル抗体(重井医学研究所 免疫部門 佐渡義一先生よりご供与)を用いてウサギ正常眼組織における基底膜 構成成分の解明を目指した。ウサギ眼組織において、IV
型コラーゲンα1
鎖とα2
鎖は結膜・輪部組織基底膜に存在し、角膜組織には発現しないことがわかり、さらに血管にも発現が見られた(図
3-4, a-f
)。IV
型コラーゲンα3
鎖は基底膜 に発現がみられなかったが、血管の一部に発現することが分かった(図3-4, g-i
)。IV
型コラーゲンα4
鎖は輪部組織基底膜のみに弱く発現することがわかった(図3-4, j-l
)。またIV
型コラーゲンα5
鎖は角膜・輪部・結膜組織すべての基底膜に存在することがわかった(図
3-4, m-o
)。25
図 3-4. ウサギ角膜、輪部、結膜組織におけるIV型コラーゲンα鎖の分布 正常眼組織におけるIV型コラーゲンα鎖の分布を免疫組織化学的解析を用いて検討 している。 IV型コラーゲン (a–c) a1、(d–f) a2、(g–i) a3、(j–l) a4、(m–o) a5 鎖の染 色結果を示している。スケールバーは25 μm。Kameishi et al., 2015 引用・改変。
【 正常組織 】 角膜
(K12 陽性細胞)
輪部
(境界部)
結膜
(K13 陽性細胞)
α5 鎖
a b c
d e f
m n o
α1 鎖
α2 鎖
α3 鎖
α4 鎖
g h i
j k l
26
これをもとに角膜上皮欠損モデル眼表面において解析を行った。
24
週、48
週角 膜上皮欠損モデルにおいて、抗IV
型コラーゲンα5
鎖抗体を用いて免疫組織化 学的解析を行った。その結果、IV
型コラーゲンα5
鎖の発現はモデル眼表面の一 部に発現が観察され(図3-5
)、これは角膜・輪部・結膜の眼表面全体に発現す る正常組織とは異なることがわかった((図3-4, m-o
)。連続切片を用いてK12
とK13
の免疫染色を行った結果、驚くべきことに、モデル眼表面に再出現したK12
陽性の角膜上皮様細胞下にのみIV
型コラーゲンα5
鎖が発現し、K13
陽性 の浸潤結膜上皮細胞下には発現しないことが分かった。図 3-5. モデル眼表面上における基底膜構成成分IV型コラーゲンα5鎖の分布 IV型コラーゲンα5鎖の免疫組織化学的解析の結果を示している。(a-c) 24週、(d-f) 48週角膜上皮欠損モデル組織であり、(a, d) 角膜上皮様細胞、(b, e) 境界部、(c, f) 浸潤結膜上皮の染色結果である。矢頭はIV型コラーゲンα5鎖の発現部位を示して いる。スケールバーは25 μm。Kameishi et al., 2015 引用・改変。
24 週 モデル
48 週 モデル
a c
【 角膜上皮欠損モデル 】
角膜上皮様細胞
(K12 陽性細胞)
浸潤結膜上皮
(K13 陽性細胞)
境界部
b
d e f
27
Endo
らは羊膜上で角膜上皮と結膜上皮を培養した場合、角膜上皮培養下でのみIV
型コラーゲンα5
鎖の発現が検出され、結膜上皮培養下では発現しないことを 報告している(Endo et al., 2004)
。以上より、本研究で観察されたin vivo
での 結果と既往研究によるin vitro
の検討により、角膜上皮細胞は組織修復や培養環 境下など分裂が盛んな状況において、早期よりIV
型コラーゲンα5
鎖を産生し 自身の周囲の環境を整え、恒常性を維持することが示唆される。これに対し、結膜上皮においては正常組織には存在が確認されたが、組織修復後や培養環境 下では発現がみられないことから、
IV
型コラーゲンα5
鎖の産生に直接関与する かを判断できず、直下に存在する繊維芽細胞が産生する可能性がある。このように、上皮細胞の表現型と基底膜の構成には密接な関係があると示唆 されたため、
IV
型コラーゲンα1, 2
鎖についても検討を行った。正常組織では 輪部上皮と結膜上皮細胞下基底膜に観察され、角膜上皮細胞下には発現しない 分子であるIV
型コラーゲンα1, 2
鎖は(図3-6, a-c
)、24
週角膜欠損モデルに おいては角膜上皮様細胞下基底膜を含む眼表面全体に存在することが分かった(図
3-6, d-f
)。これに対し、48
週角膜欠損モデルにおいては角膜上皮様細胞下基底膜には存在せず、境界部と浸潤結膜下に存在することが分かり、これは正 常組織における
IV
型コラーゲンα1, 2
鎖の分布と類似することが分かった(図3-6, g-i
)。したがって、上皮組織修復後、長期観察を行うと基底膜の構成が変動し、正常組織と類似した上皮細胞の表現型と基底膜構成成分の関係へと近づく ことが分かった。また、
IV
型コラーゲンα1, 2
鎖は正常組織とモデル組織の両 者において結合組織中のすべての血管に発現することが観察された(図3-6,
ア スタリスク)。28
図 3-6. 基底膜構成成分IV型コラーゲンα1, 2鎖の分布とその変化 正常眼組織 (a-c) と24週角膜上皮欠損モデル (d-f)、48週角膜上皮欠損モデル (g-i) において、角膜上皮細胞マーカーであるケラチン12(Keratin12: K12)と基底膜 の構成成分であるIV型コラーゲンα1, 2鎖の免疫組織化学的解析を行った。矢頭は IV型コラーゲンα1, 2鎖の発現部位を、アスタリスクは血管を示している。スケールバ ーは25 μm。Kameishi et al., 2015 引用・改変。
24 週 モデル
48 週 モデル
f
g h
d
【 角 膜 上 皮 欠 損 モ デ ル 】 角膜上皮様細胞
(K12 陽性細胞)
境界部 浸潤結膜上皮
(K13 陽性細胞)
e
i
*
*
*
*
*
*
*
* 正常組織
a b c
【 正 常 組 織 】
角膜
(K12 陽性細胞)
輪部 結膜
(K13 陽性細胞)
*
*
*