腫瘍特性にリンクしたリン酸化蛋白質の解析
田 中 正 光
秋田大学大学院医学研究科 分子生化学
(平成
21
年11
月26
日掲載決定)Tyrosine phosphorylation mediated signaling in solid tumors Masamitsu Tanaka
Department of Molecular Medicine and Biochemistry, Akita University Graduate School of Medicine, Akita 010
-8543, Japan
は じ め に
癌治療のうえで最も深刻な問題点である転移,浸潤 をいかに阻止するかという課題に対し,シグナル伝達 の立場から,治療の標的となる分子を選択するアプ ローチがよく行われている.蛋白質相互作用による 信号伝達の立場から,癌の転移,浸潤過程において 特異的に活性化される信号経路を同定し,その経路を 遮断するうえで有効な標的蛋白質のツールをデザイン する方向性である.癌細胞の正常細胞とは異なった信 号経路を探索する目安のひとつとして,蛋白質のチロ シンリン酸化レベルの異常が知られている.チロシン リン酸化は蛋白質のリン酸化全体の数
%
であるが,それが重要であることは,レトロウイルスの腫瘍原性 の元となる癌遺伝子産物や増殖因子受容体の多くがチ ロシンキナーゼである事,細胞の増殖,分化,運動性 や接着などに広く関与していることによる.現在知ら れている多くの分子標的療法は,チロシンキナーゼを 標的対象にしているが,その多くは対象の分子特異性 が高いと言えず,また酵素活性を持つキナーゼ蛋白質 は広範囲の下流分子に影響を及ぼし,正常細胞にも必 要な経路もすべて抑制してしまう副作用が問題とな る.そこで,キナーゼによりリン酸化を受ける基質蛋
白質を治療標的分子にすることは,より個別の信号経 路だけを遮断し,副作用の少ない治療設計が期待でき る.本稿では,難治性の癌であるスキルス胃癌に関与 するチロシンリン酸化蛋白質の中から,最近同定され た
Ossa
とEph/ephrin
ファミリーを中心に解説する.1. スキルス胃癌における
チロシンリン酸化蛋白質の同定スキルス胃癌は最も予後の悪い胃癌のタイプであ り,浸潤性が激しく,胃壁を破った癌細胞が腹腔にこ ぼれ落ちて,腹膜播種を起こしやすい特徴を有する.
そこでスキルス型癌が腹膜播種してゆく過程で,強く リン酸化されている蛋白質を探索した.ヒトスキルス 癌由来の胃癌細胞を用い,ヌードマウスでの腹膜播種 組織でリン酸化の亢進した蛋白質を精製し,質量分析 による同定を行った.リン酸化蛋白質は,リン酸化チ ロシンと特異的に結合する
SH2
ドメインによるア フィニティー精製と,抗リン酸化チロシン抗体による 二段階の精製を用いた.その結果,細胞の基質接着班 の形成や細胞移動にとって重要なことが知られているp130Cas
やFAK
などとともに,CDCP1,Ossa/C9orf10 などの蛋白質が同定された(図1).このうち CDCP1
は1
型の細胞膜貫通蛋白質で,Srcの主要な基質蛋白 質である.その機能として,CDCP1はSrc
とPKC δ
の足場蛋白質として働き,SrcによるPKC δ
のリン酸 化を媒介することで,細胞の足場非依存性増殖を促進 している1)
.CDCP1はスキルス胃癌において,周辺の 正常胃粘膜に比較し高い発現がみられ,スキルス胃癌 細胞が腹膜播種の過程において腹腔内で浮遊した状態Corresponcence : Masamitsu Tanaka, M.D.
Department of Molecular Medicine and Biochemistry, Akita University Graduate School of Medicine, 1
-1
-1 Hondo, Akita 010
-8543, Japan
Tel : 81
-18
-884
-6077 Fax : 81
-18
-884
-6078
E
-mail : [email protected]
-u.ac.jp
※平成
21
年1
月9
日 新任教授就任講演での生存を助けていることなどが示唆された
2)
.2. Ossa/C9orf10
Ossa(Oxidative-
stress associated Src activator) は,
酸化ストレスに伴う
Src
の新規活性化因子として命名 した3)
.癌細胞は生体のなかで,虚血や血流再開,炎症,抗癌剤や光線療法,放射線療法など多くの場面におい て酸化ストレスに晒されているが,それに抵抗して生 き延びる機構を備えている
4,5)
.その防御機構のひとつ に,酸化ストレスに応じてSrc
ファミリーの活性化が 誘導され,抗アポトーシスシグナルが生じることが知 られているが,そのメカニズムは不明である.Srcは そのC
末のチロシン部位(Y530)がリン酸化を受け ると,自身のSH2
ドメインと結合し,また分子内でSH3
ドメインとプロリンリッチ配列との結合で折り たたまれるため不活性型となる.一方で,これらの分 子内結合が無い状態ではキナーゼドメインが開いてい るため,エフェクター蛋白質に対してキナーゼ活性を 有する活性型となっている.細胞内ではそれらの構造 が中間体を含めて平衡状態を保っていると考えられて いるが,Srcが他の蛋白質と結合し,自己の分子内結 合による折りたたみが防がれている状況では,安定的に
Src
は活性化型となる(図2).そこで Ossa
とSrc
の相互作用につき調べてみたところ,Ossaはそのチ ロシンリン酸化を介してSrc
のSH2
ドメインと結合 し,またN
末付近のプロリンリッチ配列によってSrc
のSH3
ドメインと結合能を有していた.このOssa
と の蛋白質間結合によってSrc
の自己折りたたみが防が れる結果,Src
キナーゼの活性化が誘導される(図2).
Ossa
を発現しているスキルス胃癌細胞に紫外線照射 や 過 酸 化 水 素 処 理 を す る とROS(Reactive oxygen
図
1. ヌードマウスに腹膜播種した胃癌組織(左図)から精製した,チロシンリン酸化蛋白質(右図).
同定したいくつかの蛋白質名を記した.
図
2. Src
の活性化機構.分子内結合で折りたたまれた不活化状態(左).Ossaとの結合などにより,
分子内結合が阻害され活性化型となった状態(右).
中央はその中間体を示す.
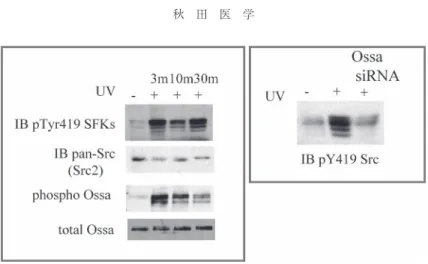
species :
活性酸素)が産生され,これに応答して数 分以内にSrc
ファミリーの活性化が誘導される.その 時RNAi
によってOssa
の発現を低下させた細胞ではROS
産生に応じたSrc
の活性化が阻害されていた(図3).つまり,酸化ストレス抵抗性としての Src
キナーゼの活性化には,Ossaが必要であると考えられた.
Ossa
はSrc
との相互作用により抗アポトーシスシ グナルを誘導する酸化ストレスに応答した,Ossaによる
Src
活性化の 意義を調べるため,そのシグナル伝達機構を解析した.まず
Ossa
は,胃癌組織でチロシンリン酸化の亢進し ている蛋白質として同定されたように,Src
ファミリー と結合してSrc
を活性化すると同時に,Srcの基質と してリン酸化を受ける(図3).同リン酸化チロシン
部位に対して,PI3キナーゼのp85
サブユニットが結 合することで,結局Ossa
はSrc
とPI3
キナーゼと複 合体を形成する足場蛋白質となっていた.その結果,Src
によるPI3
キナーゼの直接的な活性化を経て,PI3 キナーゼの標的蛋白質であるAkt
が活性化される.Akt
は抗アポトーシス作用を持つ主要な分子であるこ とが知られているが,実際Ossa
の発現をノックダウ ンさせた胃癌細胞は,紫外線照射や過酸化水素処理に よるアポトーシスが著名に亢進する.また逆にOssa
を過剰発現させた細胞では,これらの酸化ストレスに よるアポトーシスに対して有意な抵抗性がみられた.Ossa
はmRNA
結合蛋白質でIGFII
の産生を促進 するOssaのシグナル伝達をさらに検索する過程で,
Ossa
はIMP
-1(IGFII mRNA binding protein
-1) と 複
合体を形成することが分かったが,その結合はRNA
を介したものであった.解析の結果,OssaはそのC
末においてmRNA
との結合能を有しており,その一 つとしてIGFII mRNA
と結合し,そのmRNA
の安定 性を制御する.実際Ossa
の過剰発現によって,細胞 外に分泌されるIGFII
蛋白質の量も増加していた.IMP
-1
などIGFII
の産生に関わる他の蛋白質に対する 影響や,他にどのようなmRNA
とOssa
が結合するか などが,今後注目される.Ossaは,Src,PI3キナーゼとの相互作用を介した 信号伝達による経路と,C末の
RNA
結合によるIGFII
の産生促進の二つの独立した機序によって癌細胞の生 存を助ける抗アポトーシス作用をもつ蛋白質であるこ とが明らかになった(図4).ヒトの手術症例で Ossa
の発現分布を検索すると,スキルス胃癌の約7
割で腫 瘍部位にOssa
の発現が観察され,またOssa
の発現 を恒常的に抑制したスキルス胃癌細胞のヌードマウス における腫瘍形成を調べてみると,皮下移植腫瘍のサ イズや腹腔内接種による腹膜播種が有意に低下してお り,治療の分子標的になる可能性が示唆された(図5).
たとえば
Ossa
の機能により,抗癌剤による酸化スト レスに対して癌細胞が治療抵抗性を獲得している事な どが考えられ,今後の解析対象である.図
3. 紫外線照射による Src
の活性化にはOssa
が必要である.UV刺激によりSrc
が活性化(Tyr419のリン酸化で判定)され,それにより
Ossa
自身のリン酸化が生じる(左図).Ossaをノックダウンした細胞で はSrc
の活性化が誘導されない(右図).Eph/ephrin
ファミリーの癌における機能Ephファミリーは受容体型チロシンキナーゼのなか では最大の分子数をもち,そのリガンドである
ephrin
ファミリーとともに近年腫瘍における機能がクローズ アップされている.我々は以前ヒト胃癌組織で高発現 している受容体としてEphB2
をクローニングした事 から,Eph/ephrinファミリーの解析に携わっている.Eph/ephrin
の特徴はephrin
も細胞膜に固定されてい るため,Eph発現細胞とephrin
発現細胞の細胞間接 触により,双方向性のシグナルが誘導されることである
6,7,8)
.その結果生じる現象として細胞間の反発運動があげられる(図
6).Eph/ephrin
は神経細胞で発現 量が高いが,たとえばephrin
を発現している神経軸 索は対応するEph
を発現している脳の領域からは反 発され,軸索が誤った領域に投射されることを防いで いる.また,小腸などの絨毛では基底側と表層側の領 域で,それぞれEphB
受容体とephrin
-B
が発現して いるため,その境界領域では反発運動により両者の領 域の細胞が入り混じることなく,その絨毛内での位置 が規定される.他に,同ファミリーは血管の動静脈の 分化や,骨での骨芽細胞・破骨細胞の分化の制御,膵 臓ラ氏島でのインシュリンの分泌制御など多彩な機能 が知られている9,10)
.一方で,自他の報告からEph/
ephrin
は多くの腫瘍細胞で発現がみられ,その腫瘍における機能解析が最近進んできている.その中で我々 は,上皮細胞における機能や細胞内シグナル経路があ まり分かっていなかった
ephrin
-B1
の下流シグナルが癌の進展を促進していることを報告してきた.
Ephrin
-B1
のシグナル伝達は癌の進展を促進する Bタイプのephrin
の中でephrin
-B1
やephrin
-B2
は,多くの種類の癌において特に浸潤性の高い癌組織で高 発現していることが報告されている.実際,ephrin-
B1
を過剰発現させた膵臓癌細胞は,ヌードマウスの図
5. 恒常的に Ossa
をノックダウンした胃癌細胞(miOssa)は,コントロール(miLacZ)に比較し,ヌードマウス皮下腫瘍の縮小(左図)や,腸間膜播種の抑制(右図)が観察される.
図
4. Ossa
はSrc
とPI3
キナーゼの足場蛋白質として働き,PI3キナーゼ
/ Akt
の活性化を誘導し,細胞は酸化ストレス抵抗性を獲得する.その一方で
Ossa
はIGFII mRNA
と結合し,IGFIIの分泌を促進 することでも細胞の生存を促す.腹腔内移植した際の腹膜播種が亢進した
11)
.そのメカ ニズムとして,ephrin-B1
はタイトジャンクションを構成する
claudin
と相互作用する事で,細胞間接着を負 に 制 御 す る
12)
. ま たephrin
-B1
は そ のC
末 でdi-
shevelled
と結合し,そのシグナル伝達の結果RhoA
の活性化を誘導することで癌細胞の移動に寄与す る13)
.さらにephrin
-B1
のC
末を介したシグナル伝達 は,Arf1の活性化を経て細胞外基質分解酵素である マトリックスメタロプロテアーゼ(MMP)の分泌を 亢進することで,癌細胞の浸潤性を増すと考えられ た11)
(図7).この時 ephrin
-B1
を発現している癌細胞 に対して,EphB受容体による細胞外刺激が加わると,MMP
の分泌がより促進される.では実際の癌組織で も,このようなEphB
受容体を発現している細胞との 接触がみられるのか? 例えば腹膜播種の過程では癌
細胞が腹膜に生着するが,その表面を覆っている中皮 細胞では実際,EphB2やEphB4
の高い発現が認めら れる.このような場面ではEphB/ephrin
-B1
の相互作 用が癌の微小環境を規定して,ephrin-B1
を発現して いる癌細胞の局所浸潤を高めているのではないかと考 えている(図8).
癌の治療標的分子としての
Ephrin
-B1
Ephrin-
B1
を発現している癌細胞の進展を抑制する図
6. Eph
受容体とephrin
との相互作用による反発作用.細胞自体の移動や,神経軸索の投射,腸管 の上皮細胞の位置の制御などに関っている.
図
7. Ephrin
-B1
の腫瘍細胞における機能.1Ephrin-B1
を導入した膵臓癌細胞は,ヌードマウスにおける腹膜播種が促進する.2 RhoAの活性化などを介して,細胞移動を制御している.3 メタロプロテアーゼの細 胞外分泌過程を促進する.4 細胞間接着を負に制御する.
ため,
ephrin
-B1
の信号伝達を阻害する試みをおこなっ ている.まず,Ephrin-B1
のC
末を介したシグナルを 阻害するために,同部位のアミノ酸配列にHIV
の細 胞膜透過シグナルを付加した合成ペプチドを用い た14)
.このephrin
-B1 C
末ペプチド(EFNB1-C)を培
養細胞の培地中に添加すると,上記dishevelled
との結合や
RhoA・Arf1
の活性化が阻害され,またMMP
の分泌促進作用も抑制され,ephrin-
B1
のC
末に依存 するシグナル伝達を効率よく阻害できた.そこでin vivo
における効果を調べるため,ephrin-B1
を発現し ているスキルス胃癌細胞をヌードマウス腹腔内に接種 し,その後連日EFNB1
-C
ペプチドを同じく腹腔内に 投与して腹膜播種に対する影響を判定した.その結果,EFNB1
-C
ペプチド処理群では対照群に比較して,スキルス胃癌細胞の腸間膜や大網への播種,また直腸近 傍への播種が,腫瘍結節のサイズ,数いずれにおいて も 低 下 し て い た. 一 方 で,ephrin-
B1
は 対 応 す るEphB
受容体による刺激によって,細胞内領域のチロ シンリン酸化が生じる.このephrin
-B1
のチロシンリ ン酸化を介したシグナル伝達を遮断するephrin
-B1
変異体の過剰発現においても,やはりスキルス胃癌細胞 の腹膜播種の抑制を認めている
15)
.これらの事から,ephrin
-B1
のC
末を介したシグナルと,チロシンリン 酸化を介したシグナルの両方を遮断する事でephrin
-B1
を発現する癌細胞の進展をより効率よく阻害でき るのではないかと示唆された.お わ り に
以上,我々の報告してきた癌組織におけるチロシン リン酸化蛋白質の解析の中から
Ossa,ephrin
-B1
を中 心に解説した.今後の取り組みのひとつとして,この ようなリン酸化蛋白質を治療標的とした検索,基礎解 析を播種型の腫瘍組織を中心に広げてゆこうとしてい る.治療応用にあたっては,これらの実験で用いたephrin
-B1
のC
末ペプチドなどの正常細胞に対する影 響の検索が,今後の課題としてあげられる.図
8. 腹膜播種の形成過程(左図).Ephrin
-B1
を発現している癌細胞が,対応するEphB
受容体を発現している腸間膜の中皮細胞などと接触すると,ephrin-
B1
のシグナル伝達により癌細胞の浸潤が亢進するモデ ル図(右).文 献
1) Uekita, T., Jia, L., Narisawa
-Saito, M., Yokota, J., Ki- yono, T. and Sakai, R.
(2007) CUB-domain con- taining protein 1 is a novel regulator of anoikis resis- tance in lung adenocarcinoma. Mol. Cell Biol., 27, 7649
-7660.
2) Uekita, T., Tanaka, M., Takigahira, M., Nakanishi, Y., Yanagihara, K. and Sakai R.
(2008) CUB-domain containing protein 1 regulates peritoneal dissemina- tion of gastric scirrhous carcinoma. Am. J. Pathol., 72, 1729
-1739.
3) Tanaka, M., Sasaki, K., Kamata, R.,Yanagihara, K., and Sakai, R.
(2009) A novel RNA-binding protein Ossa/C9orf10 regulates activity of Src kinases to protect cells from oxidative
-stress induced apoptosis. Mol Cell Biol, 29, 402
-413, 2009.
4) Ozben, T.
(2007) Oxidative stress and apo pto-sis : Impact on cancer therapy. J. Pharm. Sci., 96, 2181
-2194.
5) Brown, NS and Bicknell, R.
(2001) Hypoxia andoxidative stress in breast cancer. Oxidative stress:
its effects on the growth, metastatic potential and re- sponse to therapy of breast cancer. Breast Cancer Res., 3, 323
-327.
6) Henkemeyer, M., Orioli, D., Henderson, JT., Saxton, TM., Roder, J., Pawson, T. and Klein, R.
(1996)Nuk controls pathfinding of commissural axons in the mammalian central nervous system. Cell, 86, 35
-46 7) Kullander, K., Mather, NK., Diella, F., Dottori, M.,
Boyd, A.W. and Klein, R.
(2001) Kinase dependentand kinase independent functions of EphA4 recep- tors in major axon tract formation in vivo. Neuron, 29, 73
-84.
8) Batlle, E., Henderson, JT., Beghtel, H. et al.
(2002)Beta
-catenin and TCF mediate cell positioning in the intestinal epithelium by controlling the expression of EphB/ephrinB. Cell, 111, 251
-263.
9) Konstantinova, I., Nikolova, G., Ohara
-Imaizumi, M., Meda, P., Kucera, T., Zarbalis, K., Wurst, W., Na- gamatsu, S. and Lammert, E.
(2007) EphA-Eph- rin
-A
-mediated beta cell communication regulates insulin secretion from pancreatic islets. Cell, 129, 359
-370.
10) Zhao, C., Irie, N., Takada, Y., Shimoda, K., Miyamoto, T., Nishiwaki, T., Suda, T. and Matsuo, K.
(2006)Bidirectional ephrinB2
-EphB4 signaling controls bone homeostasis. Cell Metab., 4, 111
-1121.
11) Tanaka, M., Sasaki, K., Kamata, R. and Sakai R.
(2007) The C-
terminus of ephrin
-B1 regulates metalloproteinase secretion and invasion of cancer cells. J. Cell Sci., 120, 2179
-2189.
12) Tanaka, M., Kamata, R. and Sakai, R.(2005) Phos- phorylation of ephrin
-B1 via the interaction with claudin following cell
-cell contact formation.
EMBO J., 24, 3700
-3711.
13) Tanaka, M., Kamo, T., Ota, S. and Sugimura, H.
(2003) Association of Dishevelled with Eph-