雑誌名 星薬科大学紀要
号 60
ページ 1‑9
発行年 2018‑12‑10
URL http://id.nii.ac.jp/1240/00000811/
1.
悪性新生物 (がん) は、 1980年代初頭より約30年 間にわたり死亡原因の第1位だった脳血管疾患を逆転 し1位となっている。 現在、 がんによる年間の死亡者 数 は37万人を超え、 総死亡者数の約30% を占める疾 患にまでに増加している (平成27年度厚生労働省死因 順位別にみた死亡数・死亡率の年次 推 移: https://ww w.mhlw.go.jp/toukei/saikin/hw/jinkou/geppo/nengai15 /dl/gaikyou27.pdf)。 がんの罹患数も人口の超高齢化に 伴い増加し続けており、 がん患者の化学療法は、 がん専 門病院ならびに大規模な病院だけでなく、 一般病院や診 療所においても日常的に広く行われるようになってきて いる。 最近では、 免疫チェックポイント阻害薬や分子標 的薬など新しい作用機序をもつ抗がん剤や新規抗がん剤 を組み込んだレジメンが次々と開発され、 がん医療の急 速な高度化・細分化が進んでいるが、 がんの治療は多く のレジメンが存在するため従来の細胞障害作用を持つ抗 がん剤も現役で活躍している。 そのため、 医師、 薬剤師 を含む多職種の専門性を活かしたチーム医療が重要とな り、 がん患者に最善の医療を提供することが重要である と考えられている。
近年では、 適切な抗がん剤選択により生存効果の延長 が見込まれるようになっているが、 副作用の強い抗がん 剤治療では、 副作用による患者のquality of life (QOL) の著しい低下により、 治療を中断するケースが多い。 最 近の報告より、 抗がん剤副作用経験者のうち副作用対策 への薬剤が処方された患者は半数程度に留まり、 その効 果も限定的であるため、 がん患者の副作用軽減に対する アンメットメディカルニーズは非常に高いことが明らか となっている (ミクスOn line: https://www.mixonlin e.jp/Article/tabid/55/artid/44046/Default.aspx)。
現在、 我々の研究グループは抗がん剤の副作用の中で も、 倦怠感/疲労感、 下痢、 味覚障害、 食欲不振などの
発症機序を解析している1-10)。 がん化学療法中には、 体 がだるい、 ひどく疲れるといった疲労感/倦怠感が頻発 し、 抗がん剤治療を受けた患者の7割以上が体験する と言われている11)。 この疲労感/倦怠感は、 QOLの著 しい低下を招く大きな要因にもかかわらず、 十分な対応 が行われていない。 この抗がん剤による疲労感/倦怠感 は、 中枢および末梢組織の機能低下など、 多くの原因で 起こると考えられるが、 詳細な機序は不明である。 そこ で、 我々の研究グループは、 この倦怠感/疲労感の発症 メカニズム解明の一環として、 シスプラチンが骨格筋組 織に影響を及ぼすことにより引き起こされるのではない かと仮説を立て、 抗がん剤であるシスプラチン投与時の 骨格筋組織への影響を検討している。 シスプラチンは白 金ベースの抗がん薬であり、 多くのがん腫に用いられて いるがん化学療法のkey drugである。 一方、 がん患者 において骨格筋量が予後と相関すること、 ならびに進行 性の骨格筋萎縮が全身状態の増悪を反映することが報告 されており、 新たな予後予測因子となる可能性も示唆さ
れている12-14)。 したがって、 がん患者における骨格筋量
の減少は予後に直結することが考えられる。 本稿では、
シスプラチンによる筋萎縮作用のメカニズム解析および 増悪、 軽減因子の検討結果の一部を紹介する。
2. 1
前述したように倦怠感/疲労感の発症メカニズム解明 の一環として、 シスプラチンが骨格筋組織に影響を及ぼ すことにより引き起こされるのではないかと仮説を立て、
シスプラチン投与マウスの骨格筋が萎縮作用を引き起こ すか否かを解析した。
2-1.
我々は、 シスプラチンをマウスに1日1回、 4日間投 与し、 大腿四頭筋ならびに後肢筋の重量ならびに筋線維 断面の直径を測定したところ、 特にシスプラチン投与に
酒 井 寛 泰
星薬科大学 生体分子薬理学研究室
Analysis of cisplatin-induced muscle atrophy
Hiroyasu SAKAI
Department of Biomolecular Pharmacology, Hoshi University
同等にまで体重を減少させたdietary restrictions (DR;
食事制限) 群よりもさらに減少していた。 この結果は、
シスプラチンによる摂食量/栄養摂取低下により引き起 こされる筋萎縮だけではなく、 シスプラチン自体が骨格 筋に対し、 筋萎縮を及ぼしている可能性が考えられる。
2-2.
種々の筋萎縮状態において筋特異的E3ユビキチンリ ガーゼであるMuscle RING Finger-1 (MuRF1) およ び muscle atrophy F-Box (MAFbx; also known as Atrogin-1) の発現が上昇することが知られている。 こ れらは筋萎縮原因遺伝子として周知されており、 筋萎縮 の特異的なマーカーとして同定されている15-18)。 そこで、
定量的 RT-PCR 法にて大腿四頭筋中のこれらの筋萎縮
原因遺伝子の発現を解析したところ、 MuRF1および
Atrogin-1 遺伝子の発現は著明に増加し、 シスプラチン 投与と同じ体重減少を示す食事制限群よりもさらに亢進 していた (Fig.3 A and B)。 Casitas B-cell lymphoma proto-oncogene b (Cbl-b) お よ び lysine 63-linked autoubiquitination of TNF receptor associated factor 6 (TRAF6) も骨格筋に発現するE3 ユビキチンリガー ゼ として知られているが19, 20)、 これらの発現もシスプ ラチン投与時に発現増加するものの、 食事制限群と比較 して変化がなかった。 MuRF1および Atrogin-1の転写 因 子 と し てForkhead box O1 (FOXO1) お よ び FOXO3が同定されている21, 22)。 そこで、 これらの遺伝 子発現を解析したところ FOXO1およびFOXO3はシ スプラチンにより発現増加したが、 FOXO3については 食事制限群よりもさらに発現が亢進していた (Fig. 3E and F)。
Fig. 1 ! "#
$%
A
!B
"#C
$%&'(#D $%&'(#E)
*p<0.05, **p<0.01 ***p<0.001 vs. VehicleVehi)
##p<0.01 ###p<0.001 vs. DR)*+,-1./0
Fig. 2 &'()$%
(123456745849:;457<=>?@
AABCDD D$
%&'((123 HE EFGHB-D6
$%&'((123=BCDD8IJ@K
! < L A )*p<0.05 ***p<0.001 vs. Vehicle)
##p<0.01 vs. DR)*+,-(1)./0
2-3.
!2"Cox2#$
%&'
TNF-、 IL-6など23)による炎症が筋萎縮の引き金に な る こ と が 知 ら れ て い る た め 、 proinflammatoly cytokine である TNF-、 IL-6 および IL-1ならびに Cox2の遺伝子発現を検討したところ全ての群において 変化は認められなかった (Fig. 4)。 したがって、 シス プラチンによる筋萎縮作用には骨格筋炎症の関与は少な いことが示唆された。
2-4. insulin-like
growth factor-1"IGF-1#myostatin
"Mstn#$()*%&'
骨格筋量はタンパク質同化および異化経路のバランス によって維持されている。 IGF-1/PI3K/ Akt/ mammal- ian target of rapamycin (mTOR) pathwayはタンパ
ク質合成を介した同化作用を示し、 Mstn/Smads or Mstn/FOXOs/MuRF1 and Atrogin-1/ubiquitin- proteasome経 路 は 異 化 作 用 を 示 す 。 IGF-1お よ び Mstn シグナリングの活性化はそれぞれ筋肥大および筋 萎縮を引き起こすことが知られている24)。 そこで、 まず、
IGF-1の遺伝子発現がどの組織で引き起こされているか
を検討したところ、 肝臓での発現が極めて高かった (Fig. 5A)。 肝臓でのIGF-1遺伝子発現変化を観察した ところ、 食事制限およびシスプラチン投与においても発 現変化は引き起こされていなかったが、 大腿四頭筋にお ける遺伝子発現では溶媒群、 食事制限群と比較してシス プラチン投与群では有意に亢進していた (Fig. 5B and C)。
Mstnの遺伝子は舌、 食道、 皮膚および骨格筋 (大腿 四頭筋) に強い発現が認められた (Fig. 6A)。 それら の中でも骨格筋 (大腿四頭筋) において著明な遺伝子発 現が確認できたので、 シスプラチン投与および食事制限 マウスの大腿四頭筋でのMstn遺伝子の発現変化を検討 したところ、 溶媒群、 食事制限群と比較してシスプラチ ン投与群ではMstnの遺伝子発現は有意に亢進していた (Fig. 6B) 。 さ ら に 、 Mstnの 下 流 で 発 現 上 昇 す る cyclin-dependent kinase (Cdk) inhibitor p2125) の遺 伝子発現亢進も有意な亢進をしていた (Fig. 6C)。
Fig. 3 +,-./012$3456
MuRF1"A#7Atrogin-1"B#7Cbl-b"C#7 TRAF6"D#7FOXO1"E# FOXO3"F#$
()*%&'8
*p<0.05, **p<0.01 ***p<0.001 vs. Vehicle#p<0.05
###p<0.001 vs. DR 1
Fig. 4 +,-./012$3456
TNF-"A#7IL-6"B#7IL-1"C# Cox2"D#$()*%&'8
1
2-5. MuRF1
Atrogin-1Akt
FOXO3a Smad2
筋萎縮原因遺伝子であるMuRF1およびAtrogin-1の タンパク質発現量は遺伝子発現量と相関してシスプラチ ン投与マウスの大腿四頭筋において溶媒群および食事制 限群と比較して有意に発現増加していた (Fig. 7)。 Akt
およびFOXO3aのリン酸化は食事制限群と比較して、
シスプラチン投与マウスの大腿四頭筋において有意に減 少していた。 また、 Mstnの下流でリン酸化されること が知られているSmad2のリン酸化は食事制限群と比較 して、 シスプラチン投与マウスの大腿四頭筋において有 意に亢進していた (Fig. 7)。
以上のことより、 シスプラチンの投与は、 IGFを介 するタンパク質合成経路 (同化経路) が抑制され、
Mstnを介するタンパク質分解経路 (異化経路) が亢進 することにより筋萎縮が引き起こされることを明らかに した。
3. !"#$%
&' ()*+,-./30
多くの医学研究および動物を用いた基礎研究により運
動は筋肥大を引き起こす因子であることは周知の事実で ある。 そこで我々のグループでは、 シスプラチンにより 引き起こされる筋萎縮において運動負荷がどのような効 果を引き起こすかを検討した。
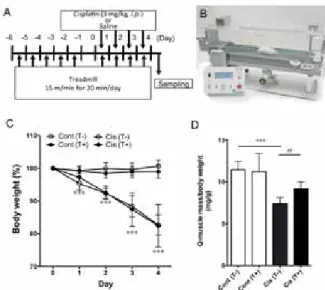
3-1. 1234 567
!"#$%&' ()*+,-89
シスプラチンの投与は有意に体重減少を引き起こすが、
トレッドミルを用いた運動の負荷 (Fig. 8A) はシスプ ラチンによる体重減少には変化を与えなかった (Fig. 8 C)。 しかしながら、 シスプラチンによる大腿四頭筋質 量低下はトレッドミルを用いた運動負荷により有意に抑 制された (Fig. 8D)。
3-2. 1234 :;<=67
!"#$%&' ()*+,-89 トレッドミルを用いた運動負荷を課したシスプラチン 投与群の大腿四頭筋の筋原線維を観察したところ、 トレッ ドミルを用いた運動負荷は、 シスプラチン誘発筋原線維 径の減少を有意に抑制した (Fig. 9)。
Fig. 5 >?@ABCDE1234
IGF-1 FGH
IGF-1 A
! "# IGF-1
$%B ! &'
()* IGF-1 $%C+**p<0.01 vs.
Vehicle+,-./1012
Fig. 6 >?@ABCDE1234
MstnFGH
Mstn A ! &'()* Mstn
$%B ! &
'()*p21 $%C+
***p<0.001 vs. Vehicle+###p<0.001 vs. DR+,-./1 012
3-3.
!"#$%&'()*
+,
トレッドミルを用いた運動負荷を課したシスプラチン 投与群の大腿四頭筋のMuRF1およびAtrogin-1の遺伝 子発現を解析したところ、 トレッドミルを用いた運動負 荷は、 シスプラチン誘発筋原線維径の減少を有意に抑制 した (Fig. 10)。
3-4. Mstn-.IGF-1
#/0&1234567 !"#
$%&'()*+,
シ ス プ ラ チ ン 投 与 群 の 大 腿 四 頭 筋 のAktお よ び
FOXO3aのリン酸化の減少はトレッドミルによる運動
負荷により有意に抑制された (data not shown; (3) を参照)。 また、 シスプラチン投与によるMstnの遺伝
子発現亢進ならびにIGF-1の遺伝子発現低下もトレッ ドミルによる運動負荷により有意に抑制された (Fig.
11 A and B)。 トレッドミルの運動負荷によりMstn の下流でリン酸化されるSmad2のリン酸化亢進は抑制 され、 IGF-1/PI3K/Akt/mTORの下流でリン酸化される
Fig. 7 8MuRF19
Atrogin-1 :;<=>?@ Akt9FOXO3a -. Smad2 AB?>?
MuRF1 Atrogin-1 !"#A-C$%
Akt !" &'("#D and E$%
FOXO3a !" &'("#F and G$%
Smad2 !" &'(" #H and I$)
*p<0.05, **p<0.01 ***p<0.001 vs. Vehicle (Vehi))
##p<0.01 ###p<0.001 vs. DR)*+,-#1$./0
Fig. 8 CDEF-. =GEF
!"#$%&'()*+, &12345678 9#A$%0:;<=>9#B$%?@A BC :;<=>9D0EF1234GH#C$%
?IBC :;<=>9D0 EF1234GH#D$
Cont#T or Tred-$:1234JKsaline L%Cont#T or Tred +$:1234M.salineL%Cis#T or Tred -$: 1234JKL%Cis#T or Tred +$: 1 234M.L)***p<0.001 vs. Cont#T-$)
##p<0.01 ###p<0.001 vs. Cis#T-$)*+,-#3$ ./0
Fig. 9 HIJEF
!"#$%&'()*+,
Cont#T -$%Cont#T +$%Cis#T -$ Cis#T +$ LNOP HE QRST#A-D$%:;<=
>9D0EF1234U%?NOPVBC DWXYKF#E$)Bar scale: 100 Zm)*p<0.05 ***p<0.001 vs. Cont#T-$)###p<0.001 vs. Cis#T-$)*
+,-(3)./0
p70S6 kinaseはシスプラチン投与により抑制され、 ト レッドミルの運動負荷により有意に回復した。
以上のことより、 トレッドミルによる運動負荷はシス プラチンによるIGF-1を介するタンパク質合成経路 (同化経路) の抑制、 Mstnを介するタンパク質分解経
26
嘔気・嘔吐はがん化学療法中に引き起こされる副作用 の中でも著しいQOLの低下を引き起こす27-29)。 シスプ ラチンは催吐性リスク分類で高度リスク (high emetic risk) 薬に分類され、 投与量が50 mg/m2を超えると 90% 以上の患者に嘔気・嘔吐が発症する30)。 この嘔気・
嘔吐対策にNK1受容体アンタゴニスト、 5-HT3受容体 アンタゴニストおよびデキサメタゾンの三薬が制吐薬と して投薬される。 このデキサメタゾンはグルココルチコ イド (ステロイド) 性薬である。 以前よりデキサメタゾ ン等のステロイドは哺乳類筋管細胞16, 31) や骨格筋16) に お い て MuRF1 や Atrogin-1 の 発 現 亢 進 を 介 し て ubiquitin-proteasome系を介してタンパク質を分解す る (異化) 作用が知られている32)。 したがって、 シスプ ラチンとデキサメタゾンの共処置は筋萎縮を増強すると いうことが推測できるが、 この仮説を証明した基礎的な 研究はない。 そこで、 我々の研究グループはシスプラチ ンとデキサメタゾンを共処置した際の筋萎縮作用および 筋萎縮原因遺伝子の発現変化を検討した。
4-1.
!
デキサメタゾン (10 mg/kg, s.c.) の共投与はシスプ ラチンによる体重減少を増強させないにも関わらず、 大 腿四頭筋質量はシスプラチンまたはデキサメタゾンの単 独投与よりも共投与することによって有意に減少させた (Fig. 12A and B)。 さらに大腿四頭筋の筋原線維径の 測定でも単独投与よりも共投与において有意に短縮させ た (Fig. 12C- G)。
4-2. "
#$%& '()*+,MuRF1"
Atrogin-1-Mstn"IGF-1)*+./
0
デキサメタゾンは単独投与により筋萎縮原因遺伝子 (MuRF1およびAtrogin-1) の発現を有意に増加させた。
またシスプラチンまたはデキサメタゾンの単独投与より も共投与することによって有意に筋萎縮原因遺伝子の発 現を増加させた (Fig. 13A and B)。 これらの遺伝子 発現変化はタンパク質レベルでも相関した結果が得られ た (Fig. 13C and D)。 さらに、 Mstnの発現変化を検 討したところ、 デキサメタゾン単独でもMstnの遺伝子 発現を亢進したが、 シスプラチンを共処置することでさ らなる遺伝子発現増加が認められた (Fig. 13E)。 また、
Fig. 10
MuRF1; AAtrogin-1; B !"#
$%&'()*+,-./0
***p<0.001 vs. ContT-0###p<0.001 vs. CisT-0 12343567'
Fig. 11 MstnA58
IGF-1B9: !"#$%&'()
*+,-./0 Smad2
58 p70S6 kinase;<:9: !"#$
%&'()*+,-./0*p<0.05, **p<0.0158**
*p<0.001 vs. ContT-0#p<0.05 58###p<0.001 vs.
CisT-012343567'
デキサメタゾン投与によっても有意なIGF-1遺伝子発 現低下が認められたが、 シスプラチン共投与によって遺 伝子発現低下が増強されなかった (Fig. 13F)。
本研究結果によりシスプラチンおよび制吐目的で投薬 されるデキサメタゾンの共投与が骨格筋萎縮を悪化させ ることマウスを用いて証明された26)。
5.
我々の研究グループはシスプラチンをマウスに投与す ると筋萎縮作用が引き起こされることを初めて証明した。
その機序には骨格筋のタンパク質異化作用を持つMstn の発現が骨格筋で亢進しオートクライン的に作用し PI3K/Akt経路を抑制することでFOXOs、 特にFOXO3 のリン酸化を抑制することでFOXO3の転写活性を亢進 させ筋萎縮原因遺伝子であるMuRF1およびAtrogin-1 の発現亢進を導く。 MuRF1およびAtrogin-1はE3ユ ビキチンリガーゼ であるためタンパク質の過剰なユビ キチン化を引き起こし、 26s proteasomeによりタンパ ク質分解が亢進され筋萎縮を誘発する。 さらに、 シスプ ラチンは骨格筋のタンパク質同化作用を持つIGF-1の 発現を低下させることによりタンパク質合成系が抑制さ れることもシスプラチンの筋萎縮作用の原因の1つと
考えられる (Fig. 14)1)。 また、 シスプラチンによる筋 萎縮作用は運動負荷によって抑制されること明らかにし た3)。 臨床的にシスプラチンを用いたがん化学療法前お よび療法中に運動をすることで筋萎縮を抑制できる可能 性が考えられるが、 がん患者の負担を考えると、 運動負 荷よりも良い対策を他に模索することが重要である。 さ らに、 我々の研究グループは、 シスプラチンの催吐作用 の対策に使用されるデキサメタゾンが筋萎縮を悪化させ る可能性も示唆した26)。 したがって、 デキサメタゾンは シスプラチン使用時の制吐療法に非常に有益であるが、
がん患者の予後に関与する筋萎縮に注意して使用する 必要である。 今後、 がん患者の予後およびQOLに直結 すると言っても過言ではないシスプラチンの筋萎縮作用 の簡便な診断方法や対応策などを探索するとともに、 が ん患者個人々に対してシスプラチンを適正使用できるよ う研究を通して貢献して行きたい。
Fig. 12 デキサメタゾンおよびシスプラチン共投与によるマウ ス体重変化 (A)、 大腿四頭筋質量 (B) および筋原線維径 の変化 (C-G)。 デキサメタゾン (Dex; 10 mg/kg, s.c.) は常法のシスプラチン (Cis; 3 mg/kg, i.p.) 投与の30 分 前 に 投 与 し た 。 Bar scale: 100 μm。 *p<0.05,
**p<0.01 および***p<0.001 vs. Control。 #p<0.05 およ び ###p<0.001, $$$p<0.001。 参考論文 (26) より引用
Fig. 13デキサメタゾンおよびシスプラチン共投与による大腿 四頭筋の MuRF1 (A)、 Atrogin-1 (B)、 Mstn (E) およ
び IGF-1 (F) 遺伝子発現変化。 デキサメタゾンおよびシ
スプラチン共投与による大腿四頭筋の MuRF1 および Atrogin-1タンパク紙レベルの変化 (C and D)。*p<0.05,
**p<0.01 および***p<0.001 vs. Control。 #p<0.05 およ び ##p<0.01 および ###p<0.001, $$p<0.01 および $$$
p<0.001。 参考論文 (26) より引用
援事業 (平成26年−平成30年, 事業番号: S1411019)、
JSPS 科研費 (若手研究 (B) 15K18880、 基盤 (C) 18K06706) の助成を受けたものです。 また、 本研究の 遂行および本稿を執筆するにあたり、 ご協力いただいた 甲斐友規氏、 木村三奈美氏、 田端祥子氏、 伊佐洋佑氏、
矢部早織氏、 猪俣茉耶氏、 月村友香氏、 浅見真穂氏、 木 寅聡子氏、 内藤宏秋氏、 茂木桃子氏、 亀井淳三教授をは じめとする生体分子薬理学研究室の皆様および湯本哲郎 教授、 里 史明講師をはじめとする薬剤師職能開発研究 部門の皆様に心より感謝申し上げます。
【利益相反】
開示すべき利益相反はない。
Fig. 14
1) Sakai H, Sagara A, Arakawa K, et al. Mechanisms of cisplatin-induced muscle atrophy. Toxicol Appl Pharmacol 2014; 278:190-9.
2) Sakai H, Sagara A, Matsumoto K, et al. Neutrophil recruitment is critical for 5-fluorouracil-induced diarrhea and the decrease in aquaporins in the colon. Pharmacol Res 2014; 87:71-9.
3) Sakai H, Kimura M, Isa Y, et al. Effect of acute treadmill exercise on cisplatin-induced muscle atrophy in the mouse. Pflugers Arch 2017; 469:1495-505.
4) Sakai H, Sato K, Sato F, et al. Curcumin inhibits epigen and amphiregulin upregulated by 2,4,6- trinitrochlorobenzene associated with attenuation of skin swelling. Inflamm Res 2017; 66:663-78.
5) Sakai H, Yabe S, Sato K, et al. ELR(+) chemokine-mediated neutrophil recruitment is involved in 2,4,6- trinitrochlorobenzene-induced contact hypersensitivity. Clin Exp Pharmacol Physiol 2018; 45:27-33.
6) Sakai H, Tabata S, Kimura M, et al. Active Ingredients of Hange-shashin-to, Baicalelin and 6-Gingerol, Inhibit 5- Fluorouracil-Induced Upregulation of CXCL1 in the Colon to Attenuate Diarrhea Development. Biol Pharm Bull 2017; 40:2134-9.
7) Sakai H, Kai Y, Oguchi A, et al. Curcumin Inhibits 5-Fluorouracil-induced Up-regulation of CXCL1 and CXCL2 of the Colon Associated with Attenuation of Diarrhoea Development. Basic Clin Pharmacol Toxicol 2016; 119:540-7.
8) Sakai H, Kai Y, Takase K, et al. Role of peptide YY in 5-fluorouracil-induced reduction of dietary intake. Clin Exp Pharmacol Physiol 2016; 43:753-9.
9) Matsumoto K, Nakajima T, Sakai H, et al. Increased expression of 5-HT3 and NK 1 receptors in 5-fluorouracil- induced mucositis in mouse jejunum. Dig Dis Sci 2013; 58:3440-51.
10) Sakai H, Sagara A, Matsumoto K, et al. 5-Fluorouracil induces diarrhea with changes in the expression of inflamma- tory cytokines and aquaporins in mouse intestines. PLoS One 2013; 8:e54788.
11) op A, Manfredi AM, Bonura S. Fatigue in cancer patients receiving chemotherapy: an analysis of published studies.
Ann Oncol 2004; 15:712-20.
12) Delmonico MJ, Harris TB, Lee JS, et al. Alternative definitions of sarcopenia, lower extremity performance, and functional impairment with aging in older men and women. J Am Geriatr Soc 2007; 55:769-74.
13) Blauwhoff-Buskermolen S, Versteeg KS, de van der Schueren MA, et al. Loss of Muscle Mass During Chemotherapy Is Predictive for Poor Survival of Patients With Metastatic Colorectal Cancer. J Clin Oncol 2016; 34:1339-44.
14) Cho KM, Park H, Oh DY, et al. Skeletal muscle depletion predicts survival of patients with advanced biliary tract cancer undergoing palliative chemotherapy. Oncotarget 2017; 8:79441-52.
15) Lecker SH, Solomon V, Mitch WE, Goldberg AL. Muscle protein breakdown and the critical role of the ubiquitin- proteasome pathway in normal and disease states. J Nutr 1999; 129:227S-37S.
16) Bodine SC, Latres E, Baumhueter S, et al. Identification of ubiquitin ligases required for skeletal muscle atrophy.
Science 2001; 294:1704-8.
17) Gomes MD, Lecker SH, Jagoe RT, Navon A, Goldberg AL. Atrogin-1, a muscle-specific F-box protein highly expressed during muscle atrophy. Proc Natl Acad Sci U S A 2001; 98:14440-5.
18) Foletta VC, White LJ, Larsen AE, Leger B, Russell AP. The role and regulation of MAFbx/atrogin-1 and MuRF1 in skeletal muscle atrophy. Pflugers Arch 2011; 461:325-35.
19) Nakao R, Hirasaka K, Goto J, et al. Ubiquitin ligase Cbl-b is a negative regulator for insulin-like growth factor 1 signaling during muscle atrophy caused by unloading. Mol Cell Biol 2009; 29:4798-811.
20) Paul PK, Bhatnagar S, Mishra V, et al. The E3 ubiquitin ligase TRAF6 intercedes in starvation-induced skeletal muscle atrophy through multiple mechanisms. Mol Cell Biol 2012; 32:1248-59.
21) Sandri M, Sandri C, Gilbert A, et al. Foxo transcription factors induce the atrophy-related ubiquitin ligase atrogin-1 and cause skeletal muscle atrophy. Cell 2004; 117:399-412.
22) Stitt TN, Drujan D, Clarke BA, et al. The IGF-1/PI3K/Akt pathway prevents expression of muscle atrophy-induced ubiquitin ligases by inhibiting FOXO transcription factors. Mol Cell 2004; 14:395-403.
23) Zhou J, Liu B, Liang C, Li Y, Song YH. Cytokine Signaling in Skeletal Muscle Wasting. Trends Endocrinol Metab 2016; 27:335-47.
24) Banerjee A, Guttridge DC. Mechanisms for maintaining muscle. Curr Opin Support Palliat Care 2012; 6:451-6.
25) Thomas M, Langley B, Berry C, et al. Myostatin, a negative regulator of muscle growth, functions by inhibiting myoblast proliferation. J Biol Chem 2000; 275:40235-43.
26) Sakai H, Kimura M, Tsukimura Y, et al. Dexamethasone exacerbates cisplatin-induced muscle atrophy. Clin Exp Pharmacol Physiol 2018 in press.
27) Hesketh PJ. Comparative review of 5-HT3 receptor antagonists in the treatment of acute chemotherapy-induced nau- sea and vomiting. Cancer Invest 2000; 18:163-73.
28) Coates A, Abraham S, Kaye SB, et al. On the receiving end--patient perception of the side-effects of cancer chemo- therapy. Eur J Cancer Clin Oncol 1983; 19:203-8.
29) Goedhals L, Heron JF, Kleisbauer JP, Pagani O, Sessa C. Control of delayed nausea and vomiting with granisetron plus dexamethasone or dexamethasone alone in patients receiving highly emetogenic chemotherapy: a double-blind, placebo-controlled, comparative study. Ann Oncol 1998; 9:661-6.
30) Schnell FM. Chemotherapy-induced nausea and vomiting: the importance of acute antiemetic control. Oncologist 2003; 8:187-98.
31) Wang L, Luo GJ, Wang JJ, Hasselgren PO. Dexamethasone stimulates proteasome- and calcium-dependent proteoly- sis in cultured L6 myotubes. Shock 1998; 10:298-306.
32) Hong DH, Forsberg NE. Effects of dexamethasone on protein degradation and protease gene expression in rat L8 myotube cultures. Mol Cell Endocrinol 1995; 108:199-209.
Analysis of cisplatin-induced muscle atrophy Hiroyasu SAKAI
Department of Biomolecular Pharmacology, Hoshi University
Cisplatin, a platinum-based anti-cancer drug, is one of the most effective broad-spectrum anti-cancer agents used against various cancers. It has been recently suggested that low skeletal muscle mass is predictive of mortality in patients with cancer. Although several molecules produced by the actual tumor itself contribute to skeletal muscle impairment, we suggested that the administration of cisplatin could increase levels of MuRF1 and Atrogin-1, possibly leading to muscle atrophy in the mouse. On the other hand, exercise is an important factor that induces muscle pro- tein synthesis and muscle hypertrophy. In addition, we recently indicated treadmill exercise could attenuate cisplatin-induced muscle atrophy. Furthermore, we show that the combination treatment of cisplatin and dexamethasone, one of the antiemetic drugs, exacerbated muscle atrophy in mice. Therefore, this treatment regimen might exacerbate muscle atrophy in cancer patients. In this review, I will describe above-mentioned pathophysiological mechanisms of cisplatin-induced muscle atrophy.