内分泌甲状腺外会誌 32(2):68-73,2015
乳癌薬物療法の新展開 特集1
乳癌のアロマターゼ阻害剤耐性機序
東北大学大学院医学系研究科分子機能解析学分野*,横浜労災病院乳腺外科**
林 慎一* 木村万里子*
,**
Resistant mechanisms of aromatase inhibitors in breast cancer
Department of Molecular and Functional Dynamics, and Center for Regulatory Epigenomics and Diseases, Tohoku University Graduate School of Medicine*, Department of Breast Surgery, Yokohama Rosai Hospital**
Shin-ichi Hayashi* and Mariko Kimura*,**
エストロゲン受容体( ER)陽性乳癌ではホルモン療法が有効であるものの,およそ3分の1は再発す る。再発のメカニズムは数多く研究されてきたが,完全に解明されたわけではなく,ホルモン療法耐性,
特にアロマターゼ阻害剤( AI)耐性は代替的な細胞内 ERシグナルの獲得が関係すると考えられている。
筆者らは癌組織検体と癌細胞株で ERの転写活性をモニタリングすることでそのメカニズムを研究してき た。エストロゲン応答配列( ERE-) GFPアデノウィルス試験では AI無効例は多様な ER活性を示し,抗 エストロゲン剤への感受性も様々であったが,これは耐性には複数の機序が存在することを示唆してい る。また, ERE-GFP導入 ER陽性乳癌細胞株から AI耐性を模倣する6種類の異なるタイプの耐性株を樹 立し,リン酸化依存性やアンドロゲン代謝物依存性など,複数の代替的な ER活性化経路が AI耐性に関 わることを明らかにした。フルベストラントや mTOR阻害剤への反応も個々の耐性株で異なっていた。
これらの結果は ER陽性乳癌の分類をさらに細分化することが,ホルモン療法だけでなく新しい分子標的 薬などの治療選択に極めて重要であることを示唆している。
Key words: エストロゲン受容体(estrogen receptor),アロマターゼ阻害剤(aromatase inhibitor),ホ ルモン療法(hormonal therapy),アンドロゲン(androgen),mTOR(mTOR)
はじめに
エストロゲンは乳癌の進行に中心的な役割を果たしてお り,エストロゲンシグナルを阻害するホルモン療法はきわ めて有効でありかつ重要である。現在,ホルモン療法には 2つの治療戦略が存在する(図1)。一つはエストロゲン 受容体α(ERα)に競合的に結合し,作用を阻害する抗エ
別冊請求先:〒980-8575 宮城県仙台市青葉区星陵町2 -1 東北大学大学院医学系研究科分子機能解析学分野 林 慎一 E-mail address: [email protected]
ストロゲン剤治療であり,もう一つはエストロゲンの合成 を阻害するアロマターゼ阻害剤( AI)によるエストロゲ ン枯渇療法である。これらのホルモン療法は乳癌の予後を 著しく改善した。約 70%の乳癌は ERα発現腫瘍であり,
ERαを標的とするホルモン療法の対象となる。 ER陽性乳 癌におけるホルモン療法は重篤な有害事象を伴わない非常 に有効な治療法であり,進行再発乳癌だけでなく補助療法 の局面でも広く使用されている[1]。しかしながら,一定 数の再発は避けられず,ホルモン療法耐性,特にタモキシ フェン( TAM)耐性に対する臨床試験が数多く行われて いる[2]。一方, AIは様々な場面において TAMに勝るこ とが大規模臨床試験で証明されている[3]。そのため,
図1 . 乳癌の内分泌療法とその耐性機序 AIはより適切なホルモン療法として,閉経後患者の進行 再発治療や術後補助療法で抗エストロゲン剤より優先され ることとなった。卵巣機能の遮断は閉経前 ER陽性乳癌に 対する有効な治療戦略の一つであるが,エストロゲン枯渇 耐性の獲得は ER陽性乳癌でホルモン療法耐性が進展し,
腫瘍がより悪性化する上で重要なステップといえる。エス トロゲン枯渇耐性の機序は,いくつかの施設でエストロゲ ンを除去した培地で長期間培養された細胞を用いて研究さ れ て き た[ 4 ~ 6]。 こ れ ま で の 報 告 で は, 耐 性 株 は MAPKや PI3K/Aktシグナル経路とのクロストークや膜 ERの関与[7]によってエストロゲン過敏性を獲得するこ とが示唆されている。しかし,正確なメカニズムは十分理 解されているとはいえず,また耐性に関係する他のメカニ ズムが存在するかという点も含め,多くの疑問が残されて いる。筆者らはエストロゲン応答配列( ERE-) GFPアデ ノウィルス試験で再発検体を解析し, ER活性と抗エスト ロゲン剤への感受性は各症例によって異なることを見出し た[8]。この問題に取り組むため,われわれは ER陽性乳 癌細胞株MCF-7,T47Dを用いてAI治療を模倣するいくつ かの培養条件からコロニーを単離し,複数の細胞株を樹立 し た。MCF-7,T47Dに は 以 前 確 立 し た 定 常 的 に ERE- GFPレポーター遺伝子を組み込んだ細胞を親株として使用 した[9]。この細胞は ER活性を反映して GFPが発現する ため,生細胞において蛍光をみることで ER活性を評価す ることができる。このシステムを用いて AI耐性を模倣す る6種類のクローン株を樹立し,その性質を解明した。今 回われわれはこれらの複数の AI耐性メカニズムを概観し,
AI耐性乳癌に対する次治療としてのフルベストラントや mTOR阻害剤の有効性にも触れる。
再発乳癌におけるAI治療後のER活性
AI無効化における ERの関与を解明するため, AI治療後 の再発腫瘍の ER活性を筆者らが開発した ERE-GFPアデノ ウィルス試験で分析した[8~10]。10例中5例では有意な ER活性を示し,そのほとんどはフルベストラントなどの 抗エストロゲン剤に対して程度の差はあれ,感受性をもっ ていた(未投稿データ)。この結果,再発例において AI耐 性の機序は単一ではなく,二次ホルモン療法の感受性も 様々であることが示された。これは臨床において観察され る治療反応性とよく一致する。
複数のAI耐性機序をモデル化する細胞の樹立
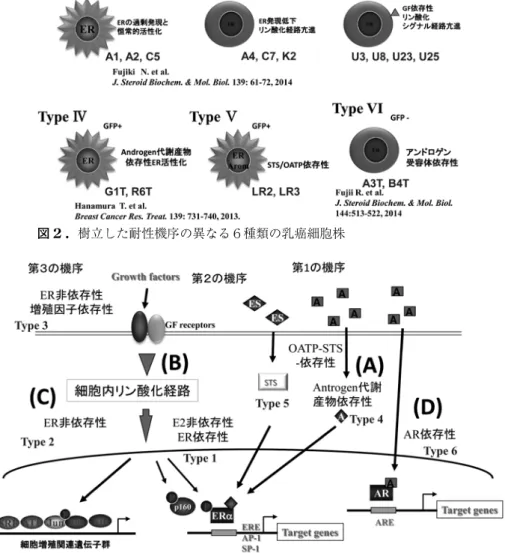
AI耐性機序の研究では,ERE-GFPを組み込んだMCF-7
( E10), T47D( TE8)から異なる耐性機序を示す6種類 の耐性株を樹立した(図2)。これらの耐性株の特徴を解 明することで, AI耐性の機序は3つのカテゴリーに分類 されることが明らかになった(図3)。まず第1にアロマ ターゼに依存しない経路によるエストロゲン生合成やエス トロゲン様作用をもったアンドロゲン代謝産物の供給によ って,AIの効果が回避されるもの(Type4,5細胞)である。
第2に細胞内リン酸化シグナル経路を経由した ERのエス トロゲン非依存的活性化に関係するもの( Type1細胞),
そして三番目は ER非依存的に増殖するメカニズムを獲得 するもの( Type2, 3, 6細胞)である。以下これらのメカ ニズムについて解説する。
ERに対するアロマターゼ非依存的リガンドの合成 第一の耐性メカニズム( Type4細胞)の例では, E10細 胞は3カ月間ステロイド除去かつテストステロン添加の条
図2 . 樹立した耐性機序の異なる6種類の乳癌細胞株
図3 . エストロゲン非依存性(AI耐性)のメカニズム
OATP:organic anion transporter peptides, STS:steroid sulfatas e
件で培養された( AI治療条件を模倣している)。生存細胞 のなかから GFP発現をモニターすることで ER活性を示す 安定的なバリアントを選択した。このバリアント細胞株で は ア ン ド ロ ゲ ン や 5α-androstane-3β,17 β-diol(3 β -diol)によってエストロゲン応答配列が誘導され,細胞 が増殖した[ 11]。この細胞株では HSD3B1の発現が上昇,
アンドロゲン受容体( AR)の発現が低下してアンドロゲ ン過剰条件へ適応し,3β-HSD type1によってジヒドロテ ストステロン( DHT)が 3β-diolに変換されていることが 判明した。異所性の HSD3B1発現や ARの抑制の結果,ア ンドロゲン過剰条件へと適応したものである。間質細胞と の共培養で局所的なアンドロゲンからのエストロゲン産生 を模倣したところ,この細胞株では親株 E10より AIに対す る感受性が低かった。この結果から,アンドロゲン代謝物 はエストロゲン様効果をもち,生存や増殖をサポートして いることが示唆された(図3A)。さらにわれわれは,原発 乳癌でもこのメカニズムが機能しているものがあり,補助 ホルモン療法への感受性に関与している可能性が認められ
た[ 12]。 ER陽性乳癌におけるアンドロゲン代謝物依存的 細胞増殖は AI耐性機序において,なんらかの役割を果た していることが示唆される。
細胞内リン酸化シグナル経路によるERの活性化 MCF-7-E10細胞を3カ月ステロイド枯渇培地で培養す ると,生存細胞内に ERE-GFPの発現がみられる細胞とみ られない細胞のコロニーが出現した。生細胞でGFPをみな がらこれらを単離し,別々にクローン化することでエスト ロゲン枯渇耐性( EDR)細胞株を樹立し,その結果,高 い ER活性をもつ3つのバリアントと ER活性をもたない3 つのバリアントが得られた[ 13]。 ERE-GFP陽性の3つの EDR細胞株では ERの過剰発現といくつかの ER標的遺伝子 の高い発現がみられた。さらに,細胞内リン酸化シグナル 経路の解析では,他の報告と同様,リン酸化ERα(Ser167)
とAkt(T308)の著しい変化が認められた[14~16]。包括 的リン酸化プロテオミクスでも同じく, Akt経路が ERα のリン酸化に寄与する可能性が示された。結果として, PI3K/
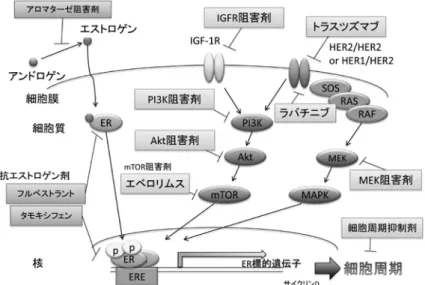
図4 . ER陽性乳癌の治療標的
Akt経路によって ERがリン酸化されて活性化することが この細胞の増殖に重要であることが示唆された(図3B)。
ER非依存的増殖メカニズムの獲得
一方, GFP陰性細胞株の3つのバリアントは ER活性を 失っており,前述の3つのGFP陽性細胞と同様にクローン 化すると,c-Jun N-terminal kinase(JNK)の恒常的活性 化が示された。これは ERE-GFP陰性 EDR細胞でみられ,
JNK阻害剤によって抑制された。 IGF-1R特異的阻害剤は JNKのリン酸化を消失させたことにより,新しいシグナル 経路である IGF-IR/JNK経路が ER非依存的 MCF-7細胞の 増殖に重要であるようだ(図3C)。これらの結果により ER 陽性乳癌細胞は同時に2つ以上の耐性機序を獲得しうるこ とが示唆された。これは複数の耐性メカニズムが一つの腫 瘍内で同時に出現しうることを示しており,患者個人にお いても異なる耐性メカニズムが同時に存在しうることを意 味する。
さらに,われわれは AI耐性として別種の ER陽性乳癌細 胞株であるT-47Dから,前述の耐性株とは異なる安定的バ リアント細胞を樹立した[ 17]。本耐性細胞株では ARと PSAが過剰発現し,エストロゲンによっては増殖が刺激さ れず, ERの発現もみられなかった。これらの細胞ではア ンドロゲンによって著しく増殖が誘導された。加えて,マ イクロアレイの結果から,親株に比してバリアント細胞で はアンドロゲン誘導性遺伝子発現プロファイルが著しく高 発現していることが認められた(図3D)。そこで,21例の 原発巣と AI耐性後の転移巣のペア検体で免疫染色を行い,
免疫染色結果では PSAと Ki67が有意に上昇し, ERとプロ ゲステロン受容体( PgR)がいくつかの症例では転移巣に おいて原発巣より低下していることを観察した[ 17]。 ER 陽性乳癌でも症例によっては ER依存性から AR依存性へと
腫瘍細胞が変化することで AI耐性が出現することが示唆 された。 AR阻害剤は特定の患者群では有効である可能性 が示唆された。
AI耐性乳癌の治療戦略
臨床的に AI耐性を獲得した乳癌の多くは ER陽性を維持 しており,現在, AI耐性乳癌には抗エストロゲン剤であ るフルベストラントが多くの場合で使用されている。フル ベストラントはタモキシフェン同様,抗エストロゲン剤と して ERの機能を阻害すると同時に,タモキシフェンと異 なり, ERを分解する作用も有する[ 18]。フルベストラン トの第3相臨床試験である CONFIRM試験は閉経後 ER陽 性進行再発乳癌を対象に,フルベストラントを2種の濃度 群( 250mg対 500mg)で比較した試験であり,フルベスト ラント 500mgは忍容性の点で問題なく,全生存( OS)を 延長するという結果であった[19]。フルベストラントは前 述の AI耐性乳癌細胞株のなかで, ER陽性を保っている Type1,4,5細胞には強い増殖抑制効果を示したのに対し,
ER陰転化したType2,3,6細胞には無効であった。つまり,
ER依存性 AI耐性機序(アロマターゼ非依存的リガンド供 給やエストロゲン非依存的 ER活性化)(図1)はフルベス トラントに感受性をもっているが, ER非依存的 AI耐性機 序( ER非依存的増殖)では無効であることを示す結果で あった。
これまでの AI耐性獲得後の乳癌治療はホルモン剤の単 剤投与が治療の柱であったが,基礎研究の結果から AI耐 性乳癌に対する臨床上の治療戦略を考えると, ERが陽性 のままであれば, ERシグナル経路と他の増殖推進経路を 同時に標的とすることが重要であると考えられる(図4)。 なかでも ER陽性乳癌では PIK3CAの変異が 28~ 47%みら れており[20],PI3K/Akt/mTOR経路は ERシグナル経路
との間にクロストークが存在すること,また AI耐性乳癌 ではしばしば PI3K/Akt/mTORシグナル経路の亢進がみ られていることから,この経路は重要なターゲットである と考えられている。 mTOR阻害剤エベロリムスは,非ス テロイド型 AI耐性の進行再発乳癌において,ステロイド 型 AIエキセメスタンとの併用によりエキセメスタン単剤 投与と比較し無増悪生存期間を有意に延長する結果となっ た( BOLERO2試験)[ 21]。同薬剤は全生存では差がつか なかったものの[22],その臨床効果は病勢のコントロール に有益であると報告されている[ 23]。 AIとエベロリムス の併用は,ホルモン剤と分子標的治療の併用という新たな 標準治療の先駆けとして注目され,現在は同様の薬剤の臨 床試験が多く組まれている。なかでも, PI3Kを標的とす る阻害剤は有効性が期待される薬剤であり,特にサブユニ ット特異的 PI3K阻害剤は汎 PI3K阻害剤と比較し,薬剤効 果と有害事象の忍容性の点で優れた効果を発揮すると考え られている[ 24]。問題は, ER陽性乳癌では多くの症例で PI3CAの遺伝子変異がみられるにも関わらず,それが PI3K阻害剤効果のバイオマーカーにはなっていないこと である[25]。エベロリムスにおいても同様にバイオマーカ ーの特定は問題となっており,基礎研究において,著者ら の PI3K/Akt/mTORシグナル経路が亢進している全ての タイプの AI耐性細胞に対しても,エベロリムスは高い抗 腫瘍効果を示したが,臨床的にはより薬剤効果の期待でき る患者群を選別するマーカーによる治療適用が必要である と考えられる。
その他,細胞周期抑制剤やHDAC阻害剤,プロテアソー ム阻害剤など異なるコンセプトに基づいた薬剤も新規治療 ターゲットとして検討されている。なかでも, CDK4/6阻 害剤 Palbociclibは閉経後 ER陽性進行・再発乳癌における 一次治療として, AI剤レトロゾール単剤と比較し,併用 療法で有意に無増悪生存期間を延長した( PALOMA-1試 験)[ 26]。しかし,同試験で設定されたバイオマーカーに よる薬剤効果の期待できる群の選別は想定と逆の結果とな り,バイオマーカー研究の難しさを露呈することとなっ た[26]。さらに,IGF,MET,Src,MAPKなど他のシグ ナル経路[24,27]をターゲットにする阻害剤もまた,多く は抗エストロゲン剤との併用で臨床試験が施行され始めて きている。しかし,標的の増加がホルモン療法を進歩させ るとは単純にいえず,基礎研究において ER陽性乳癌の治 療標的候補となる薬剤は非常に多く報告され,各種分子標 的治療薬はホルモン療法耐性乳癌に対する有望な戦略の一 つと期待されてはいるが,バイオマーカーによる治療対象 の選別や,より効果的な薬剤の組み合わせの決定など,臨 床でより効果的に使用するには多くの課題が残されている。
本邦でも既に, mTOR阻害剤エベロリムスは AI剤エキ セメスタンとの併用で適応可能となったが,今後, AI耐
性乳癌治療においては内分泌治療薬と分子標的薬の併用が 主流となると思われる。しかし,どのような順序で,どの ように各種内分泌治療薬と新規分子標的薬を組み合わせた ら良いのか,大規模な臨床試験だけでなく, in vitro, in vivo両面からの細胞内での作用機序を考慮した基礎研究も 重要であろう。新たなコンパニオンバイオマーカーを見出 し,いかに費用対効果を挙げ,患者の利益に繋げていくか,
さらなる基礎と臨床の連携した研究が必要とされる。
【文 献】
1. Strasser-Weippl K, Goss PE : Advances in adjuvant hor- monal therapy for postmenopausal women. J Clin Oncol 23 : 1751-1759, 2005
2. Riggins RB, Schrecengost RS, Guerrero MS, et al. : Path- ways to tamoxifen resistance. Cancer Lett 256 : 1-24, 2007 3. Baum M, Buzdar A, Cuzick J, et al. : Anastrozole alone or
in combination with tamoxifen versus tamoxifen alone for adjuvant treatment of postmenopausal women with early- stage breast cancer : results of the ATAC (Arimidex, Tamoxifen Alone or in Combination) trial efficacy and safety update analyses. Cancer 98 : 1802-1810, 2003 4. Yue W, Wang JP, Conaway MR, et al. : Adaptive hyper-
sensitivity following long-term estrogen deprivation : in- volvement of multiple signal pathways. J Steroid Biochem Mol Biol 86 : 265-274, 2003
5. Sabnis GJ, Jelovac D, Long B, et al. : The role of growth factor receptor pathways in human breast cancer cells adapted to long-term estrogen deprivation. Cancer Res 65 : 3903-3910, 2005
6. Martin LA, Farmer I, Johnston SR, et al. : Elevated ERK1/
ERK2/estrogen receptor cross-talk enhances estrogen- mediated signaling during long-term estrogen deprivation.
Endocr Relat Cancer 12(Suppl1) : S75-84, 2005
7. Santen RJ, Song RX, Masamura S, et al. : Adaptation to estradiol deprivation causes up-regulation of growth fac- tor pathways and hyper sensitivity to estradiol in breast cancer cells. Adv Exp Med Biol 630 : 19-34, 2008
8. Gohno T, Seino Y, Hanamura T, et al. : Individual tran- scriptional activity of estrogen receptors in primary breast cancer and its clinical significance. Cancer Med 1 : 328-337, 2012
9. Yamaguchi Y, Takei H, Suemasu K, et al. : Tumor-stromal interaction through the estrogen-signaling pathway in hu- man breast cancer. Cancer Res 65 : 4653-4662, 2005 10. Matsumoto M, Yamaguchi Y, Seino Y, et al. : Estrogen
signaling ability in human endometrial cancer through the cancer-stromal interaction. Endocr Relat Cancer 15 : 451- 463, 2008
11. Hanamura T, Niwa T, Nishikawa S, et al. : Androgen me- tabolite-dependent growth of hormone receptor-positive
breast cancer as a possible aromatase inhibitor-resistance mechanism. Breast Cancer Res Treat 139 : 731-740, 2013 12. Hanamura T, Niwa T, Gohno T, et al. : Possible role of the
aromatase-independent steroid metabolism pathways in hormone responsive primary breast cancers. Breast can- cer Res Treat 143 : 69-80, 2014
13. Fujiki N, Konnno H, Kaneko Y, et al. : Estrogen response element-GFP (ERE-GFP) introduced MCF-7 cells demon- strated the coexistence of multiple estrogen-deprivation resistant mechanisms. J Steroid Biochem Mol Biol 139 : 61- 72, 2014
14. Cambell RA, Bhat-Nakshatri P, Patel NM, et al. : Phospha- tidylinositol 3-kinase/AKT-mediated activation of estro- gen receptor alpha : a new model for anti-estrogen resis- tance. J Biol Chem 276 : 9817-9824, 2001
15. Castano E, Vorojeikina DP, Notides AC : Phosphorylation of serine-167 on the human oestrogen receptor is impor- tant for oestrogen response element binding and tran- scriptional activation. Biochem J 326 : 149-157, 1997 16. Yamashita H, Nishio M, Kobayashi S, et al. : Phosphoryla-
tion of estrogen receptor alpha serine 167 is predictive of response to endocrine therapy and increases postrelapse survival in metastatic breast cacer. Breast Cancer Res 7 : R753-764, 2005
17. Fujii R, Hanamura T, Suzuki T, et al. : Increased androgen receptor activity and cell proliferation in aromatase inhib- itor-resistant breast carcinoma. J Steroid Biochem Mol Biol 144 : 513-522, 2014
18. Miller TW : Endocrine resistance : what do we know? Am Soc Clin Oncol Educ Book, 2013
19. Di Leo A, Jerusalem G, Petruzelka L, et al. : Final overall survival : fulvestrant 500mg vs 250mg in the randomized CONFIRM trial. J Natl Cancer Inst 106 : djt337, 2014
20. Zardavas D, Phillips WA, Loi S : PIK3CA mutations in breast cancer : reconciling findings from preclinical and clinical data. Breast Cancer Res 16 : 201, 2014
21. Baselga J, Campone M, Piccart M, et al. : Everolimus in postmenopausal hormone-receptor-positive advanced breast cancer. N Engl J Med 366 : 520-529, 2012
22. Piccart M, Hortobagyi GN, Campone M, et al. : Everolimus plus exemestane for hormone-receptor-positive, human epidermal growth factor receptor-2-negative advanced breast caner : overall survival results from BOLERO-2.
Ann Oncol 25 : 2357-2362, 2014
23. Diaby V, Adunlin G, Ali AA, et al. : Using quality-adjusted progression-free survival as an outcome measure to as- sess the benefits of cancer drugs in randomized-controlled trials : case of the BOLERO-2 trial. Breast Cancer Res Treat 146 : 669-673, 2014
24. Zardavas D, Baselga J, Piccart M : Emerging targeted agents in metastatic breaset cancer. Nat Rev Clin Oncol 10 : 191-210, 2013
25. Musgrove EA, Sutherland RL : Biological determinants of endocrine resistance in breast cancer. Nat Rev Cancer 9 : 631-643, 2009
26. Cadoo KA, Gucalp A, Traina TA : Palbociclib : an evi- dence-based review of its potential in the treatment of breast cancer. Breast Cancer (Dove Med Press) 6 : 123- 133, 2014
27. Johnston SR, Schiavon G : Treatment algorithms for hor- mone receptor-positive advanced breast cancer : going forward in endocrine therapy---overcoming resistance and introducing new agents. Am Soc Clin Oncol Educ Book, 2013
![図 1 . 乳癌の内分泌療法とその耐性機序 AIはより適切なホルモン療法として,閉経後患者の進行 再発治療や術後補助療法で抗エストロゲン剤より優先され ることとなった。卵巣機能の遮断は閉経前 ER陽性乳癌に 対する有効な治療戦略の一つであるが,エストロゲン枯渇 耐性の獲得は ER陽性乳癌でホルモン療法耐性が進展し, 腫瘍がより悪性化する上で重要なステップといえる。エス トロゲン枯渇耐性の機序は,いくつかの施設でエストロゲ ンを除去した培地で長期間培養された細胞を用いて研究さ れ て き た[ 4 ~ 6]](https://thumb-ap.123doks.com/thumbv2/123deta/7622837.2548130/2.892.179.687.80.418/ホルモンエストロゲン対するエストロゲンホルモンエストロゲ.webp)