⑱ オニヤブソテツ−ニオウヤブマオウ群落(表
7
) ニオウヤブマオウは土壌の堆積した海岸の攪乱地に 生えるイラクサ科の植物で,高さは1.5m 前後になっ て群落をつくる。本群落はニオウヤブマオウが厚い葉 を広げびっしりと生え優占するが,ハチジョウススキ, リュウキュウチク,カタバミ等の攪乱種が常在する。 平島はノヤギが多く植生にダメージを与えてるが,ニ オウヤブマオウはシカやノヤギなどの大型草食動物は 採食を避けるため群落をつくり発達する。全島の周囲 で土壌がたまったところに点々と群落を発達させてい る。 崖地群落 ⑲ ホソバワダン−ボタンボウフウ群落(表7
) 強風時に海水飛沫が達する風が強いところでは,岩 隙地に植被率が低い群落が形成される。群落の高さは 0.5m 前後,ボタンボウフウが優占し,ホソバワダン, オニヤブソテツ,ハチジョウススキなどが常在する。 群落の面積は小規模で断崖地や断崖地の前面等に多 い。トカラ列島では,ボタンボウフウは多年生の高さ が1m に達するコダチボタンボウフウが分布すること が知られるが(志内 ・ 堀田,2015),平島においても コダチボタンボウフウ混在している。両者は幼苗では 識別できず,調査区内のものはボタンボウフウとした。 また,ホソバワダン,ボタンボウフウともノヤギの好 餌植物でノヤギの口の届くところは被食される。この ためノヤギも上れない急崖地に群落は形成される。 ⑳ ハチジョウカグマ群落(表7
) 潮風の影響が,少ない急斜面や断崖地には大型シダ 植物のハチジョウカグマが優占する群落が形成され る。本群落には低木も混じるのが一般的で,先駆種の アカメガシワ等も含まれるが,ハチジョウシダ,ホシ ダ,ツワブキによって識別されるが,ハチジョウスス キ,ハマボッスが常在する。なお,本群落はトカラ地 域では普遍的な群落であるが,平島ではリュウキュウ チクに被圧され分布は少ない。 ㉑ オイランアザミ群落(表7
) オイランアザミは1m に達する悪石島を南限とする キク科植物で平島は南限地帯に当たる。また,平島で はゴボウと呼ばれ主に根茎が食されるが種子島等では とげを取り除いた葉柄や葉身が食用となる。 本群落はやや肥沃な礫地や岩隙地等にオイランアザ ミが草本層に優占する群落で,イソテンツキ,ハチ ジョウススキ,グンバイヒルガオが常在し,植被率が 40 ~ 80%と空隙が多い。花期の夏季には群落の高さ は1m 前後,植被率も100%近くになるものと予想さ れる。 ノヤギにとってはこのとげのため食べにくい不嗜好 植物の1つで,ロゼット状になってとげを外に向かわ せることで被食を免れ群落を形成できる。 ㉒ ハマゴウ群落(表7
) ハマゴウは主に砂丘地に矮性の低木群落をつくるが, 礫地や岩隙地でも群落をつくる。九州以北では主にチ ガヤ,ハマゴウを区分種にしてチガヤ−ハマゴウ群集 として,種子島以南ではクロイワザサ,ハマゴウを区 分種とするクロイワザサ−ハマゴウ群集に群落単位区 分されており,平島ではクロイワザサ−ハマゴウ群集 に帰属するものと思われるが,かつて平島で確認され ていた(平田,1995)クロイワザサを調査区内で確認 できずハマゴウ群落とした。海岸浸食が著しく,かつ て砂丘地の砂が大幅に減少し陸側が堆積した礫や岩角 地になっているところもあり,海岸植生の先端がハマ ヒルガオを区分種にもつハマゴウ群落となっている。 本群落はグンバイヒルガオを区分種にもつグンバイヒ ルガオ下位単位と含まない典型下位単位に下位区分さ れる。 グンバイヒルガオ下位単位(表7
)はグンバイヒル ガオを含み時にハマゴウを欠いてグンバイヒルガオが びっしりと繁り優占する。砂丘地先端に形成されるハ マアズキ−グンバイヒルガオ群集が,クロイワザサ− ハマゴウ群集と生育場所を分けずに合体したような群 落である。南の浜港のコンクリート護岸や東の浜の礫 地海岸等で調査した。 典型下位単位(表7
)はグンバイヒルガオを含まず, ハチジョウススキを構成種に持ち,時にハチジョウス スキが優占する。 ハマゴウはシカの不嗜好植物の1つで,シカの多い 屋久島や阿久根大島の海岸にも大規模な群落を形成し ているがノヤギに対しても不嗜好植物となっている。 ㉓ ハチジョウススキ群落(表7
) ハチジョウススキが優占する群落は,人為的攪乱の ある道路辺等には普遍的な群落であるが,海岸で起表
8
湿地耕作地群落組成表 ㉗セリーシチトウイ群落 ㉘オランダガラシ群落 ㉙ヒデリコーコナギ群落 ㉚ヤナギタデ群落 ㉛クサネム群落 ㉜ハイニシキソウーホナガイヌビユ群落 ㉝ハスノハカズラ群落 ㉞ナピアグラス群落 群落番号 ㉗ ㉘ ㉙ ㉚ ㉛ ㉜ ㉝ ㉞ 調査区番号 22 23 61 10 63 71 69 62 60 91 67 35 調査月日 (2016年) 3月16日 3月16日 11月9日 3月16日 11月9日 11月10日 11月9日 11月9日 11月9日 11月12日 11月9日 3月18日 標高(m) 5 5 95 130 95 95 95 95 95 115 95 130 方位 E E − − − − − − − − − − 傾斜 (゜) 10 10 0 0 0 0 0 0 0 0 0 0 調査面積(m × m) 1×2 1×5 1×5 2×2 1×5 2×3 2×5 2×3 2×2 10×10 1×15 5×5 備考 低木層(S)の高さ(m) 3 低木層(S)の植被率(%) 80 草本層(H)の高さ(m) 1.2 0.4 0.3 0.8 1 1.5 1 0.8 0.5 0.3 1 2 草本層(H)の植被率(%) 70 90 100 90 80 90 100 95 95 100 10 100 出現種数 8 7 7 4 16 10 11 9 16 18 17 3 和名 階層 22 23 61 10 63 71 69 62 60 91 67 35 セリ−シチトウイ群落区分種 Oenanthe javanica セリ H 2・3 5・4 ・ ・ + ・ 1・1 ・ ・ ・ ・ ・ Pteris fauriei ハチジョウシダ H 1・2 1・2 ・ ・ ・ ・ ・ ・ ・ ・ ・ ・ Hydrocotyle sibthorpioides チドメグサ H +・2 2・3 ・ ・ ・ ・ ・ ・ ・ ・ ・ ・ Cyperus malaccensis ssp. monophyllus シチトウイ H 4・4 + ・ ・ ・ ・ ・ ・ ・ ・ ・ ・ オランダガラシ群落区分種 Nasturtium officinale オランダガラシ H ・ ・ 5・5 5・4 + + ・ ・ ・ ・ ・ ・ ヒデリコ−コナギ群落区分種Colocasia esculenta var. aquatilis ミズイモ H ・ ・ ・ 1・1 4・4 3・3 5・4 ・ ・ ・ ・ ・
Echinochloa crus-galli イヌビエ H ・ ・ ・ ・ + 3・3 1・1 ・ ・ ・ ・ ・ Alternanthera sessilis ツルノゲイトウ H ・ ・ ・ ・ + ・ 1・1 ・ ・ ・ ・ ・ Fimbristylis miliacea ヒデリコ H ・ ・ ・ ・ + 2・2 ・ ・ ・ ・ ・ ・ Monochoria vaginalis コナギ H ・ ・ ・ ・ ・ 3・3 1・2 ・ ・ ・ ・ ・ ヤナギタデ群落区分種 Ranunculus sieboldii シマキツネノボタン H ・ ・ ・ ・ ・ ・ ・ + ・ ・ ・ ・ Polygonum hydropiper ヤナギタデ H ・ ・ 1・2 ・ ・ ・ 2・3 5・4 ・ ・ ・ ・ クサネム群落区分種 Aeschynomene indica クサネム H ・ ・ 1・1 ・ ・ ・ ・ 2・3 5・5 ・ ・ ・ Polygonum japonicum シロバナサクラタデ H ・ ・ ・ ・ ・ ・ ・ ・ 1・2 ・ ・ ・ ハイニシキソウーホナガイヌビユ群落区分種 Euphorbia chamaesyce ハイニシキソウ H ・ ・ ・ ・ ・ ・ ・ ・ ・ + ・ ・
Artemisia indica var. orientalis ニシヨモギ H ・ ・ ・ ・ ・ ・ ・ ・ ・ 1・1 ・ ・
Ipomoea Batatas Lam. var. edulis サツマイモ H ・ ・ ・ ・ ・ ・ ・ ・ ・ 5・5 ・ ・
Amaranthus glacilis ホナガイヌビユ H ・ ・ ・ ・ ・ ・ ・ ・ ・ 2・3 ・ ・
Digitaria ciliaris メヒシバ H ・ ・ ・ ・ ・ ・ ・ ・ ・ 1・2 ・ ・
ニオウヤブマオ群落区分種
Musa paradisiaca var. sapientum バナナ S ・ ・ ・ ・ ・ ・ ・ ・ ・ ・ 5・4 ・
Stephania japonica ハスノハカズラ H ・ ・ ・ ・ ・ ・ ・ + ・ ・ 2・2 ・ Boehmeria gigantea ニオウヤブマオ H ・ ・ ・ ・ ・ ・ ・ ・ ・ ・ 2・2 ・ ナピアグラス群落区分種 Pennisefum purpurreum ナピアグラス H ・ ・ ・ ・ ・ ・ ・ ・ ・ ・ ・ 5・5 その他の種 Centella asiatica ツボクサ H + ・ ・ ・ ・ ・ ・ +・2 + +・2 +・2 ・ Commelina diffusa シマツユクサ H ・ ・ ・ ・ + 1・1 1・2 1・1 1・2 ・ ・ ・ Ludwigia octovalvis キダチキンバイ H ・ ・ 1・1 ・ 1・1 ・ 3・3 1・1 1・2 ・ ・ ・ Eclipta prostrata タカサブロウ H ・ ・ 1・1 ・ + ・ ・ ・ 1・2 + ・ ・ Polygonum longisetum イヌタデ H ・ ・ ・ ・ ・ 1・1 1・2 2・3 1・2 ・ ・ ・ Panicum repens ハイキビ H ・ ・ ・ ・ +・2 ・ ・ ・ 2・3 + ・ ・ Paspalum urvillei タチスズメノヒエ H ・ ・ ・ ・ ・ 2・2 ・ 2・2 ・ + ・ ・ Thelypteris acuminata ホシダ H + 1・2 ・ ・ ・ ・ ・ ・ ・ ・ ・ + Pleioblastus linearis リュウキュウチク H ・ ・ ・ ・ ・ ・ ・ ・ ・ + 1・2 ・
Miscanthus sinensis var.
condensatus ハチジョウススキ H ・ 1・2 ・ ・ ・ ・ ・ ・ ・ ・ + ・
Paspalum oribiculare スズメノコビエ H ・ ・ ・ ・ + ・ ・ ・ 1・2 ・ ・ ・
Viola yedoensis var.
pseudo-japonica リュウキュウコスミレ H ・ ・ ・ ・ ・ ・ ・ ・ + ・ 1・1 ・ Oplismenus compositus エダウチチヂミザサ H ・ ・ ・ ・ ・ ・ ・ ・ 1・1 ・ + ・ Oxalis corymbosa ムラサキカタバミ H ・ ・ ・ ・ ・ ・ ・ ・ ・ 1・2 + ・ Lemna aequinoctialis ナンゴクアオウキクサ H ・ ・ ・ 1・2 ・ 2・3 ・ ・ ・ ・ ・ ・ Rumex japonica ギシギシ H ・ ・ ・ 1・2 ・ ・ ・ ・ ・ + ・ ・ Oxalis corniculata カタバミ H ・ ・ ・ ・ ・ ・ ・ ・ ・ +・2 ・ ・
出現1回の種 Also in 22:Phyla nodiflora イワダレソウ H + ,Scirpus ternatanus オオアブラガヤ H 1・2,in 23:Hydrocotyle maritima ノチドメ H +・2,in 35:Polygonum capita
tum ツルソバ H + ,in 60:Cynodon dactylon ギョウギシバ H + ,Cyperus polystachyos イガガヤツリ H + ,Diodia teres オオフタバムグラ H + ,Rhynchospora rubra イ
ガクサ H + ,Splanum nigrum イヌホウズキ H 1・1,in 61:Cardamine flexuosa タネツケバナ H + ,Lindernia antipoda スズメノトウガラシ H + ,in 63:Scirpus mucronatus var.
robustus カンガレイ H 2・2,Acorus calamus var. angustatus ショウブ H 2・2,Juncus effusus var. decipiens イ H + ,Cyperus difformis タマガヤツリ H + ,Sacciolepis
indica ハイヌメリ H + ,in 67:Youngia japonica オニタビラコ H + ,Miscanthus sinensis ススキ H 1・1, Alpinia intermedia アオノクマタケラン H + ,Glochidion obovatum
カンコノキ H + ,Acalypha autralis エノキグサ H + ,Rubus croceacanthus var maximowiczii リュウキュウバライチゴ H + ,Sambucus chinensis ソクズ H + ,Rubus sie
boldii ホウロクイチゴ H + ,:in 69:Gynostemma pentaphyllum アマチャヅル H + ,Scirpus juncoides イヌホタルイ H +・2,in 71:Arthraxon hispidus コブナグサ H 2・3,in
91:Bothriospermum tenellum ハナイバナ H + ,Cerastium glomeratum オランダミミナグサ H + ,Cyperus iria コゴメガヤツリ H + ,Commelina benghalensis マルバツユク
表
9
路 傍 ・ 路 上 ・ 耕 作 地 植 物 群 落 ㉟ ア オ イ ゴ ケ 群 落 ㊱ ツ ボ ク サ -チ ガ ヤ 群 落 ㊱ -1 -a 典 型 下 位 単 位 ㊱ -1 -b 典 型 下 位 単 位 シ バ ハ ギ フ ァ シ ス ㊱ -1 -c 典 型 下 位 単 位 オ オ バ コ フ ァ シ ス ㊱ -2 -a ハ チ ジ ョ ウ ス ス キ 下 位 単 位 ギ ョ ウ ギ シ バ フ ァ シ ス ㊱ -2 -b ハ チ ジ ョ ウ ス ス キ 下 位 単 位 シ バ フ ァ シ ス ㊱ -2 -c ハ チ ジ ョ ウ ス ス キ 下 位 単 位 ㊲ ハ イ キ ビ 群 落 ㊳ ア オ チ カ ラ シ バ 群 落 ㉟ ㊱ ㊲ ㊳ ㊱ -1 ㊱ -2 ㊱ -1 -a ㊱ -1 -b ㊱ -1 -c ㊱ -2 -a ㊱ -2 -b ㊱ -2 -b 調 査 区 番 号 2 52 66 64 39 65 28 13 72 29 1 50 11 92 38 40 86 51 48 36 47 調 査 月 日 ( 20 16 年 ) 3 月 15 日 11 月 8 日 11 月 9 日 11 月 9 日 3 月 19 日 11 月 9 日 3 月 16 日 3 月 16 日 11 月 10 日 3 月 16 日 3 月 15 日 11 月 8 日 3 月 16 日 11 月 12 日 3 月 19 日 3 月 19 日 11 月 11 日 11 月 8 日 11 月 7 日 3 月 18 日 11 月 7 日 標 高 ( m ) 13 0 5 95 95 14 0 95 21 0 5 12 5 12 5 13 0 75 12 0 12 0 14 0 14 0 14 5 80 80 13 0 80 方 位 − W N W − − − − W − − W − − − − − − − − − − 傾 斜 ( ゜) 0 30 20 0 0 0 0 5 0 0 30 0 0 0 0 0 0 0 0 0 0 調 査 面 積 ( m × m ) 0 . 2 × 5 2 × 3 2 × 5 5 × 10 5 × 5 5 × 10 1 × 5 3 × 8 10 × 10 5 × 5 1 × 15 3 × 5 5 × 15 10 × 10 5 × 5 5 × 5 10 × 10 10 × 10 5 × 5 5 × 5 5 × 5 備 考 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 草 本 層 ( H ) の 高 さ ( m ) 0 . 2 0 . 1 0 . 05 1 . 2 0 . 1 1 0 . 2 0 . 1 0 . 5 1 0 . 5 0 . 8 0 . 8 0 . 8 0 . 3 0 . 5 1 . 5 0 . 8 1 1 . 2 1 草 本 層 ( H ) の 植 被 率 ( % ) 40 30 70 10 0 80 80 80 95 95 90 70 95 10 0 95 80 90 10 0 10 0 10 0 10 0 10 0 出 現 種 数 6 7 12 18 8 32 19 12 27 17 32 19 17 14 15 18 10 10 5 5 7 和 名 階 層 2 52 66 64 39 65 28 13 72 29 1 50 11 92 38 40 86 51 48 36 47 ア オ イ ゴ ケ 群 落 区 分 種 D ic ho nd ra re pe ns ア オ イ ゴ ケ H 3 ・ 4 3 ・ 4 4 ・ 4 ・ ・ ・ ・ ・ ・ ・ ・ ・ ・ ・ ・ ・ ・ ・ ・ ・ ・ ツ ボ ク サ ー チ ガ ヤ 群 落 区 分 種 D ic ho nd ra re pe ns チ ガ ヤ H ・ ・ ・ 3 ・ 3 1 ・ 1 ・ 1 ・ 2 ・ 2 ・ 3 1 ・ 1 1 ・ 1 3 ・ 3 4 ・ 4 4 ・ 4 4 ・ 4 2 ・ 2 ・ 1 ・ 2 ・ 1 ・ 2 ・ C en te lla a si at ic a ツ ボ ク サ H ・ ・ ・ 1 ・ 2 + 2 ・ 2 1 ・ 2 ・ 1 ・ 2 ・ 1 ・ 2 ・ 1 ・ 2 ・ + 1 ・ 2 ・ ・ ・ ・ ・ Vi ol a ye do en si s va r. ps eu do -ja po ni ca リ ュ ウ キ ュ ウ コ ス ミ レ H ・ ・ ・ ・ ・ + ・ ・ + ・ ・ ・ ・ ・ ・ ・ ・ ・ ・ ・ ・ ハ チ ジ ョ ウ ス ス キ 下 位 単 位 区 分 種 M isc an th us s in en sis v ar . c on de ns at us ハ チ ジ ョ ウ ス ス キ H ・ ・ ・ ・ ・ ・ ・ 2 ・ 2 2 ・ 2 1 ・ 2 ・ 2 ・ 3 1 ・ 2 2 ・ 2 + ・ 2 + 1 ・ 2 ・ ・ ・ ・ Vi ci a sa tiv a カ ラ ス ノ エ ン ド ウ H ・ ・ ・ ・ ・ ・ ・ ・ ・ ・ 1 ・ 2 ・ + ・ + + ・ 2 ・ ・ ・ ・ ・ P ae de ria s ca nd en s ヘ ク ソ カ ズ ラ H ・ ・ ・ ・ ・ ・ ・ ・ ・ + ・ + 1 ・ 2 ・ ・ + ・ ・ ・ ・ ・ シ バ ハ ギ フ ァ シ ス 区 分 種 D es m od iu m h et er oc ar po n シ バ ハ ギ H ・ ・ + ・ 5 ・ 4 4 ・ 4 ・ ・ ・ ・ ・ ・ 1 ・ 2 ・ ・ ・ ・ ・ ・ ・ ・ オ オ バ コ フ ァ シ ス 区 分 種 P la nt ag o as ia tic a オ オ バ コ H ・ ・ ・ ・ ・ ・ 4 ・ 4 ・ ・ ・ ・ ・ ・ ・ ・ ・ ・ ・ ・ ・ ・ ギ ョ ウ ギ シ バ フ ァ シ ス 区 分 種 C yn od on d ac ty lo n ギ ョ ウ ギ シ バ H ・ ・ ・ ・ 1 ・ 2 ・ ・ 5 ・ 4 2 ・ 2 ・ ・ ・ ・ 2 ・ 3 2 ・ 2 2 ・ 2 ・ 1 ・ 2 1 ・ 2 ・ + シ バ フ ァ シ ス 区 分 種 Zo ys ia ja po ni a シ バ H ・ ・ ・ ・ ・ ・ ・ ・ 4 ・ 4 ・ ・ ・ ・ 2 ・ 3 ・ ・ ・ ・ ・ ・ ・ ハ イ キ ビ 群 落 区 分 種 P an ic um re pe ns ハ イ キ ビ H ・ ・ ・ 2 ・ 3 ・ ・ ・ ・ 1 ・ 2 ・ ・ ・ 2 ・ 3 1 ・ 2 2 ・ 2 4 ・ 4 5 ・ 5 5 ・ 5 5 ・ 5 5 ・ 5 1 ・ 2 ア オ チ カ ラ シ バ 群 落 区 分 種 Pe nn ise tu m a lop ec ur oid es f. vi rid es ce ns ア オ チ カ ラ シ バ H ・ ・ ・ ・ ・ ・ ・ ・ ・ ・ ・ ・ ・ ・ ・ ・ ・ 1 ・ 2 ・ ・ 5 ・ 4 P en ni se tu m a lo pe cu ro id es チ カ ラ シ バ H ・ ・ ・ ・ ・ ・ ・ ・ ・ ・ ・ ・ ・ ・ ・ ・ ・ ・ ・ ・ 1 ・ 2 そ の 他 の 種 P le io bl as tu s lin ea ris リ ュ ウ キ ュ ウ チ ク H ・ 1 ・ 2 ・ + ・ 1 ・ 1 1 ・ 2 ・ ・ 1 ・ 1 2 ・ 2 2 ・ 3 1 ・ 2 ・ ・ 1 ・ 1 ・ 1 ・ 1 + ・ 1 ・ 2 O xa lis c or ni cu la ta カ タ バ ミ H ・ 1 ・ 2 ・ ・ ・ 1 ・ 2 + 1 ・ 2 + 1 ・ 2 1 ・ 2 + ・ + + ・ ・ ・ ・ ・ ・ P as pa lu m u rv ill ei タ チ ス ズ メ ノ ヒ エ H + ・ ・ 2 ・ 3 ・ ・ ・ ・ ・ 1 ・ 2 ・ ・ 2 ・ 3 1 ・ 2 ・ + ・ + + + + H yd ro co ty le m ar iti m a ノ チ ド メ H 2 ・ 3 ・ ・ ・ + ・ ・ ・ + ・ 1 ・ 2 ・ ・ ・ + + + ・ 2 ・ + + ・ Vi ol a ye do en si s ノ ジ ス ミ レ H ・ ・ ・ ・ ・ ・ 1 ・ 2 + ・ 2 ・ 1 ・ 2 1 ・ 2 ・ + ・ + + ・ ・ ・ ・ ・ E m ili a so nc hi fo lia ウ ス ベ ニ ニ ガ ナ H ・ ・ + ・ ・ + ・ ・ + + ・ + ・ ・ ・ + ・ ・ ・ ・ ・ Yo un gi a ja po ni ca オ ニ タ ビ ラ コ H ・ ・ ・ ・ ・ + ・ 1 ・ 2 ・ ・ 1 ・ 2 + ・ ・ ・ + ・ ・ ・ ・ ・ E cl ip ta p ro st ra ta タ カ サ ブ ロ ウ H ・ + 2 ・ 2 1 ・ 1 ・ 2 ・ 3 ・ ・ ・ ・ ・ + ・ ・ ・ ・ ・ ・ ・ ・ ・P as pa lu m o rib ic ul ar e ス ズ メ ノ コ ビ エ H ・ ・ ・ ・ ・ 1 ・ 2 ・ ・ 2 ・ 3 ・ ・ ・ ・ 2 ・ 2 ・ + + ・ ・ ・ ・ B oe hm er ia n iv ea v ar . n ip po ni ca カ ラ ム シ H ・ ・ 1 ・ 1 ・ ・ 1 ・ 1 ・ ・ ・ + 2 ・ 3 + ・ ・ ・ ・ ・ ・ ・ ・ ・ S po ro bo lu s fe rti lis ネ ズ ミ ノ オ H ・ ・ ・ ・ ・ 1 ・ 2 ・ ・ 1 ・ 2 ・ ・ 1 ・ 2 ・ 1 ・ 2 ・ ・ ・ + ・ ・ ・ P ol yg on um c hi ne ns e ツ ル ソ バ H ・ ・ ・ ・ ・ ・ 1 ・ 2 ・ ・ 3 ・ 4 1 ・ 2 ・ + ・ ・ ・ ・ ・ ・ ・ ・ Fi m br is ty lis d ic ho to m a f. fl o rib un da ク グ テ ン ツ キ H ・ ・ ・ 2 ・ 2 ・ 1 ・ 2 ・ ・ ・ ・ ・ + ・ 2 ・ 2 ・ 2 ・ ・ ・ ・ ・ ・ ・ A na ga lli s ar ve ns is f. co er ul ea ル リ ハ コ ベ H ・ ・ ・ ・ + ・ ・ ・ ・ ・ 1 ・ 2 ・ ・ ・ + ・ 2 + ・ 2 ・ ・ ・ ・ ・ M is ca nt hu s si ne ns is ス ス キ H ・ ・ ・ ・ ・ ・ ・ ・ 1 ・ 2 ・ ・ ・ ・ ・ 1 ・ 1 + ・ ・ ・ ・ ・ C om m el in a di ffu sa シ マ ツ ユ ク サ H ・ ・ ・ + ・ + ・ ・ ・ ・ ・ ・ ・ ・ ・ ・ + ・ ・ ・ ・ B ot hr io sp er m um te ne llu m . ハ ナ イ バ ナ H ・ ・ 2 ・ 2 ・ ・ ・ 1 ・ 2 ・ ・ ・ + ・ ・ ・ ・ ・ ・ ・ ・ ・ ・ W ah le nb er gi a m ar gi na ta ヒ ナ ギ キ ョ ウ H ・ ・ ・ ・ ・ 1 ・ 2 ・ ・ + ・ + ・ ・ ・ ・ ・ ・ ・ ・ ・ ・ G na ph al iu m ja po ni cu m チ チ コ グ サ H ・ ・ ・ ・ ・ ・ + ・ + ・ ・ ・ + ・ ・ ・ ・ ・ ・ ・ ・ Vi ol a m an ds hu ric a va r. tri an gu la ris ア ツ バ ス ミ レ H ・ ・ ・ ・ ・ ・ ・ ・ + ・ ・ ・ ・ ・ + + ・ ・ ・ ・ ・ E up ho rb ia c ha m ae sy ce ハ イ ニ シ キ ソ ウ H ・ + ・ ・ ・ 2 ・ 3 ・ ・ 1 ・ 2 ・ ・ ・ ・ ・ ・ ・ ・ ・ ・ ・ ・ C er as tiu m g lo m er at um オ ラ ン ダ ミ ミ ナ グ サ H + ・ ・ ・ ・ ・ + 1 ・ 2 ・ ・ ・ ・ ・ ・ ・ ・ ・ ・ ・ ・ ・ A rte m is ia in di ca v ar . o rie nt al is ニ シ ヨ モ ギ H + ・ ・ ・ ・ ・ 2 ・ 2 ・ ・ ・ 2 ・ 2 ・ ・ ・ ・ ・ ・ ・ ・ ・ ・ Ly si m ac hi a ja po ni ca コ ナ ス ビ H ・ ・ ・ ・ ・ 1 ・ 2 + ・ 2 ・ ・ ・ 1 ・ 1 ・ ・ ・ ・ ・ ・ ・ ・ ・ ・ S et ar ia g la uc a va r. pa lli de -fu sc a コ ツ ブ キ ン エ ノ コ ロ H ・ ・ ・ 3 ・ 3 ・ 1 ・ 1 ・ ・ 1 ・ 1 ・ ・ ・ ・ ・ ・ ・ ・ ・ ・ ・ ・ G al iu m s pu riu m f. s tri go su m ヤ エ ム グ ラ H ・ ・ ・ ・ ・ ・ ・ ・ ・ + 1 ・ 3 ・ ・ ・ + ・ ・ ・ ・ ・ ・ K um m er ow ia s tri at a ヤ ハ ズ ソ ウ H ・ ・ ・ ・ ・ ・ ・ ・ 1 ・ 2 ・ ・ ・ + 2 ・ 3 ・ ・ ・ ・ ・ ・ ・ S te lla ria a qu at ic a ウ シ ハ コ ベ H ・ ・ ・ ・ ・ ・ + + ・ 2 ・ 2 ・ ・ ・ ・ ・ ・ ・ ・ ・ ・ ・ Fi m br is ty lis d ic ho to m a f. an nu a テ ン ツ キ H ・ ・ ・ ・ ・ + ・ ・ + ・ ・ ・ ・ 2 ・ 2 ・ ・ ・ ・ ・ ・ ・ Th el yp te ris a cu m in at a ホ シ ダ H ・ ・ ・ ・ ・ ・ + ・ ・ ・ 1 ・ 1 ・ ・ ・ ・ ・ ・ ・ ・ ・ ・ Lu dw ig ia o ct ov al vi s H ・ ・ ・ ・ ・ + ・ ・ ・ ・ ・ ・ ・ ・ ・ ・ + ・ ・ ・ ・ C re pi di as tru m la nc eo la tu m ホ ソ バ ワ ダ ン H ・ ・ ・ ・ ・ 1 ・ 1 ・ ・ ・ ・ ・ ・ 1 ・ 1 ・ ・ ・ ・ ・ ・ ・ ・ E ch in oc hl oa c ru s-ga lli イ ヌ ビ エ H ・ ・ ・ ・ ・ ・ ・ ・ ・ ・ ・ ・ ・ ・ ・ ・ ・ + ・ ・ + H yd ro co ty le s ib th or pi oi de s チ ド メ グ サ H ・ ・ ・ ・ ・ ・ ・ ・ ・ ・ + ・ 2 ・ ・ ・ ・ ・ + ・ ・ ・ ・ O xa lis c or ym bo sa ム ラ サ キ カ タ バ ミ H ・ ・ ・ ・ ・ ・ 1 ・ 2 ・ ・ 2 ・ 2 ・ ・ ・ ・ ・ ・ ・ ・ ・ ・ ・ C on yz a su m at re ns is オ オ ア レ チ ノ ギ ク H ・ ・ ・ ・ ・ + ・ ・ ・ ・ ・ ・ + ・ ・ ・ ・ ・ ・ ・ ・ C yp er us ro tu nd us ハ マ ス ゲ H ・ 1 ・ 2 ・ ・ ・ + ・ 2 ・ ・ ・ ・ ・ ・ ・ ・ ・ ・ ・ ・ ・ ・ ・ C ar da m in e fl e xu os a タ ネ ツ ケ バ ナ H ・ ・ + ・ ・ + ・ ・ ・ ・ ・ ・ ・ ・ ・ ・ ・ ・ ・ ・ ・ A ca ly ph a au tra lis エ ノ キ グ サ H ・ ・ 1 ・ 2 + ・ ・ ・ ・ ・ ・ ・ ・ ・ ・ ・ ・ ・ ・ ・ ・ ・ C yp er us p ilo su s オ ニ ガ ヤ ツ リ H ・ ・ ・ + ・ ・ ・ ・ ・ ・ ・ ・ ・ ・ ・ ・ 1 ・ 2 ・ ・ ・ ・ O pl is m en us u nd ul at ifo liu s チ ヂ ミ ザ サ H ・ ・ ・ ・ ・ ・ 2 ・ 2 ・ ・ ・ 2 ・ 2 ・ ・ ・ ・ ・ ・ ・ ・ ・ ・ S id a rh om bi fo lia キ ン ゴ ジ カ H ・ ・ ・ ・ ・ ・ ・ ・ ・ ・ 1 ・ 1 + ・ 2 ・ ・ ・ ・ ・ ・ ・ ・ ・ A ju ga d ec um be ns キ ラ ン ソ ウ H ・ ・ ・ ・ ・ ・ ・ ・ + ・ 1 ・ 1 ・ ・ ・ ・ ・ ・ ・ ・ ・ ・ D uc he sn ea c hr ys an th a ヘ ビ イ チ ゴ H ・ ・ ・ ・ ・ ・ ・ ・ ・ 2 ・ 3 ・ ・ + ・ ・ ・ ・ ・ ・ ・ ・ C ra ss oc ep ha lu m c re pi di oi de s ベ ニ バ ナ ボ ロ ギ ク H ・ ・ ・ ・ ・ + ・ ・ ・ + ・ ・ ・ ・ ・ ・ ・ ・ ・ ・ ・ G er an iu m c ar ol in ia nu m ア メ リ カ フ ウ ロ H ・ ・ ・ ・ + ・ ・ ・ ・ ・ ・ ・ ・ ・ + ・ ・ ・ ・ ・ ・ M ed ic ag o hi sp id a ウ マ ゴ ヤ シ H ・ ・ ・ ・ + ・ 2 ・ ・ ・ ・ ・ ・ ・ ・ ・ ・ + ・ ・ ・ ・ ・ P hy sa lis a ng ul at a セ ン ナ リ ホ オ ズ キ H ・ ・ + ・ ・ ・ ・ ・ ・ ・ ・ + ・ ・ ・ ・ ・ ・ ・ ・ ・ P hy sa lis m in im a ヒ メ セ ン ナ リ ホ オ ズ キ H ・ ・ + ・ ・ + ・ ・ ・ ・ ・ ・ ・ ・ ・ ・ ・ ・ ・ ・ ・ M ur da nn ia n ud ifl or a ア レ チ イ ボ ク サ H ・ ・ ・ ・ ・ 2 ・ 3 ・ ・ + ・ ・ ・ ・ ・ ・ ・ ・ ・ ・ ・ ・ 出 現 1 回 の 種 A ls o in 1 :C yc as re vo lu ta ソ テ ツ H + , Fi cu s pu m ila オ オ イ タ ビ H 1 ・ 2 , Fa rfu gi um ja po ni cu m ツ ワ ブ キ H + , Vi ol a gr yp oc er as f. lu ci da リ ュ ウ キ ュ ウ タ チ ツ ボ ス ミ レ H 1 ・ 2 , C ar ex b re vi cu lm is ア オ ス ゲ H + , R ub us p al vi fo liu s ナ ワ シ ロ イ チ ゴ H 2 ・ 3 , S te lla ria m ed ia コ ハ コ ベ H 1 ・ 2 , Li liu m lo ng ifl or um テ ッ ポ ウ ユ リ H 1 ・ 2 , B ot hr io ch lo a pa rv ifl or a ヒ メ ア ブ ラ ス ス キ H + ・ 2 , S is yr in ch iu m a la nt ic um ニ ワ ゼ キ シ ョ ウ H + , E rig er on ca na de ns is ヒ メ ム カ シ ヨ モ ギ H + , in 2 :A gr op yr on ts uk us hi en se v ar . t ra ns ie ns カ モ ジ グ サ H + , in 11 :V io la m an ds hu ric a ス ミ レ H + , in 13 :V ite x ro tu nd ifo lia ハ マ ゴ ウ H 1 ・ 2 , Tr ig on ot is p ed un cu la ris キ ュ ウ リ グ サ H 1 ・ 1 , C or on op us d id ym us カ ラ ク サ ガ ラ シ H + , W ed el ia b ifl or a キ ダ チ ハ マ グ ル マ H + , C he no po di um v irg at um マ ル バ ア カ ザ H + , in 28 :R an un cu lu s si eb ol di i シ マ キ ツ ネ ノ ボ タ ン H + , C yn od on d ac ty lo n ギ ョ ウ ギ シ バ H 2 ・ 2 , Vi ci a hi rs ut a ス ズ メ ノ エ ン ド ウ H + , in 29 :R ub us c ro ce ac an th us v ar . m ax im ow ic zi i リ ュ ウ キ ュ ウ バ ラ イ チ ゴ H 3 ・ 3 , H er iti er a lit to ra lis サ キ シ マ フ ヨ ウ H + , in 36 :P en ni se tu m p ur pu re um ナ ピ ア グ ラ ス H + ・ 2 , in 38 :T rif ol iu m du bi um コ メ ツ ブ ツ メ ク サ H + , in 50 :B oe hm er ia g ig an te a ニ オ ウ ヤ ブ マ オ H 1 ・ 2 , S te ph an ia ja po ni ca ハ ス ノ ハ カ ズ ラ H + , Tu bo ca ps ic um a no m al um v ar .o bt us um マ ル バ ハ ダ カ ホ ウ ズ キ H + , O pl is m en us c om po si tu s エ ダ ウ チ チ ヂ ミ ザ サ H 2 ・ 3 , A be lm os ch us m os ch at us リ ュ ウ キ ュ ウ ト ロ ロ ア オ イ H 3 ・ 3 , E up ho rb ia s up in a コ ニ シ キ ソ ウ H + , in 51 :C hl or is g ay an a ア フ リ カ ヒ ゲ シ バ H 1 ・ 2 , Fi m br is ty lis d ip hy llo id es ク ロ テ ン ツ キ H + , in 52 :D ig ita ria h en ry i ヘ ン リ ー メ ヒ シ バ H 1 ・ 2 , in 64 :P ol yg on um lo ng is et um イ ヌ タ デ H + , A es ch yn om en e in di ca ク サ ネ ム H 2 ・ 2 , P ol yg on um h yd ro pi pe r ヤ ナ ギ タ デ H 1 ・ 1 , D io di a te re s オ オ フ タ バ ム グ ラ H + , Tr ic ho sa n th es ro st ra ta ケ カ ラ ス ウ リ H 1 ・ 2 , C hr ys an th em um o rn at um v ar . t ok ar en se ト カ ラ ノ ギ ク H + ・ 2 , P as pa lu m th un be rg ii ス ズ メ ノ ヒ エ H + , in 65 :L ys im ac hi a m au rit ia na ハ マ ボ ッ ス H 1 ・ 2 , R hy nc ho sp or a ru br a イ ガ ク サ H + ・ 2 , C yp er us b re vi fo liu s va r. le io le pi s ヒ メ ク グ H 1 ・ 2 , C yp er us ir ia コ ゴ メ ガ ヤ ツ リ H + , O xa lis s tri ct a オ ッ タ チ カ タ バ ミ H + ・ 2 , E le us in e in di ca オ ヒ シ バ H + , in 66 :G al iu m g ra ci le ns コ バ ノ ヨ ツ バ ム グ ラ H + , C om m e lin a be ng ha le ns is マ ル バ ツ ユ ク サ H 1 ・ 2 , in 72 :Z oy si a m at re lla v ar . p ac ifi ca コ ウ ラ イ シ バ H 2 ・ 3 , C ar ex b ru nn ea コ ゴ メ ス ゲ H + , O en ot he ra la ci ni at a コ マ ツ ヨ イ グ サ H + , D ig ita ria ra di co sa コ メ ヒ シ バ H 1 ・ 2 , La sp ed ez a cu ne at a メ ド ハ ギ H 1 ・ 2 , in 86 :S ci rp us m uc ro na tu s va r. ro bu st us カ ン ガ レ イ H 1 ・ 2 , S al ix p ie ro tii オ オ タ チ ヤ ナ ギ H 1 ・ 1 , in 92 :L as pe de za c un ea ta v ar . s er pe ns ハ イ メ ド ハ ギ H 1 ・ 2 , B id en s pi lo sa L . v ar . m in or シ ロ ノ セ ン ダ ン グ サ H +
こった土砂崩れ等の自然攪乱で自然植生が破壊された 立地にも群落は形成される。高さ1.2m のハチジョウ ススキが優占し,リュウキュウチク,グンバイヒルガ オ,ソナレムグラなどが常在する。ハマゴウ群落やホ ソバワダン−マルバニッケイ群集などの矮性低木林や 風衝低木林等が破壊されたところに成立していた。 ㉔ マルバハダカホオズキ群落(表
7
) 東の浜海岸では生帯として先端部は直径が30㎝ほど ある礫地海岸になっている。この岩隙地に厚く柔らか そうな濃い緑の葉をもつマルバハダカホオズキが優占 する群落が塊状になって点々と分布している。マルバ ハダカホオズキの他にはハマエノコロが随伴し,カタ バミやツボクサも常在するが被度は低い。ノヤギが多 く出没する海岸部で冬期に濃い緑の群落はよく目立つ。 草食獣が採食を避ける有毒植物からなる不嗜好植物群 落の1つと考えられる。 ㉕ コウボウシバ群落(表7
) 本群落はコウボウシバが優占する低茎の草本群落で 砂丘地に成立する。平島には現在砂丘地は海岸浸食の ため減少しつつあり,西海岸部では礫地海岸ばかりで 砂丘地は皆無に近い。わずかに残っていた砂丘の傾斜 地に成立しており,ごく小規模な6㎡ほどの群落であ る。ヘンリ−メヒシバやハマボッス,イソテンツキな どが随伴する。 ㉖ チガヤ−リュウキュウチク群落(表7
) 風衝の強い海岸部から山脚部に懸けてはチガヤが優 占する群落が成立する。平島では山脚部から山地部で はリュウキュウチク群落が広く占めるが,海岸部でテ ラス状になった突端部や海岸の小丘に本群落は成立す る。群落の高さは0.5m ~ 1m 前後でチガヤ,リュウ キュウチクの被度が高い。ヒナギキョウやツルモウリ ンカ,ハイニシキソウなど低茎の植物が随伴する。 湿生地群落・水田雑草群落 ㉗ セリ−シチトウイ群落(表8
) 東の浜海岸で地下水がしみ出す傾斜地や湿地ではセ リ,ハチジョウシダ,チドメグサ,シチトウイが区分 種となる群落が確認された。シチトウイは貧養な立地 のため高さは1.2m 前後と低い。水路に沿って小規模 な群落ではあるが,シチトウイあるいはセリが優占す る。シチトウイは平島では魚の干物をつるすときや大 根を干すときにひもとして利用した(用澤氏談)こと から古い時代に伝わってきた栽培植物と考えられる。 ㉘ オランダガラシ群落(表8
) 地下水が流入する集落近くの水田放棄地や水路にオ ランダガラシがびっしりと優占する群落が形成される。 オランダガラシのほかは周辺の湿性植物からなり,水 路に沿って5㎡程度の小群落である。オランダガラシ は現在野菜として利用されている。 ㉙ ヒデリコ−コナギ群落(表8
) 平島では現在イネは栽培されず水田の一部には里芋 の1種のミズイモが栽培されている。ここではコナギ, イヌビエ,ツルノゲイトウ,ヒデリコ,ミズイモを区 分種とするヒデリコ−コナギ群落が確認された。イネ に替わり植栽されたミズイモが優占する中に典型的な 水田雑草が小規模な群落をつくっていた。 ㉚ ヤナギタデ群落(表8
) ヤナギタデは,河川植生として流路に沿ってヤナ ギタデが優占する1年生の植物群落を形成する。また, 水田放棄地の中で降水時には冠水する立地に同様に群 落をつくる。平島でも水田放棄地で水路に沿ってヤナ ギタデが優占する群落を形成している。シマキツネノ ボタンを区分種とするほか,シマツユクサ,キダチキ ンバイ,タカサブロウ,イヌタデ,ハイキビ等が随伴 する。 ㉛ クサネム群落(表8
) 水田放棄地にはクサネム,シロバナサクラタデを区 分種としてクサネムが優占する群落が抽出された。ヤ ナギタデ群落と隣接するため随伴する種も類似するが, ヤナギタデ群落に比較してやや貧栄養な立地で冠水時 に水深の浅いところに成立していた。 畑地雑草群落 ㉜ ハイニシキソウ−ホナガイヌビユ群落(表8
) サツマイモ畑等の富栄養な立地ではハイニシキソウ, ニシヨモギ,ホナガイヌビユ,メヒシバを区分種とし て本群落が抽出された。高さは30㎝前後で耕作や収穫 の前後で植被率や被度等は変化する。本群落はノヤギ 被害を防ぐため防護柵を設置している畑地での調査結果であり,他所では確認できなかった。 ㉝ ハスノハカズラ−ニオウヤブマオウ群落 バナナ植栽地ではバナナの下生えにニオウヤブマオ ウ,ハスノハカズラの群落が抽出された。バナナは道 路沿いの農地に植栽されたもので,ノヤギの侵入があ るため他の雑草は食され,ノヤギが採食しない不嗜好 植物で有毒植物のニオウヤブマオウ,ハスノハカズラ が繁茂したものと考えられ,同様の群落は近隣の臥蛇 島でも確認されている(寺田,1999)。道路周辺で数 カ所ハスノハカズラがお互いに絡みつき盛り上がるよ うに優占する群落が確認されている。 ㉞ ナピアグラス群落(表
8
) ナピアグラス(ネピアグラス)は,アフリカ原産の C4植物で草丈は5m を越すこともあるイネ科の多年草 である。超大型の飼料作物として昭和初期に日本に導 入され , 平島にも飼料として導入されたが,栄養価も 少ないため現在は栽培されることは少なく放置されて いる。本群落は水田放棄地に高さが2m あるナピアグ ラスが侵入し群落をつくったものであるが,ナピアグ ラス1種が優占し,随伴する種はまれである。 路上植物群落・路傍植物群落 ㉟ アオイゴケ群落(表9
) 道路周辺のコンクリートの隙間や未舗装の道路上, 廃土捨て場などに高さが3㎝に満たないアオイゴケが びっしりと生え優占する群落が点在する。1㎡前後の きわめて小規模な群落で,やや湿潤な立地を反映し, タカサブロウやタチスズメノヒエなどが常在する。 ㊱ ツボクサ−チガヤ群落(表9
) 定期的に道路整備等の刈り取りが行われる道路周辺 ではチガヤ,ツボクサ,リュウキュウコスミレを含む 群落が形成される。チガヤは被度が高く,優占する事 もある。本群落はハチジョウススキ,ヘクソカズラ, カラスノエンドウを区分種にしてハチジョウススキ下 位単位(表9
)と典型下位単位(表9
)に識別される。 典型下位単位は刈り取り等の頻度がハチジョウスス キ下位単位より高く,その中にはシバハギが優占する シバハギファシス(表9
),オオバコが優占するオオバ コファシス(表9
)がある。 シバハギファシスは高さが20㎝前後のシバハギが びっしりと優占する群落でヘリポートなど地表面を均 一化する整地を行った場所を定期的に管理し,時間が 経過している適潤地に成立する。オオバコファシスは 半陰の湿潤から適潤の立地でオオバコが優占する。未 舗装の道路上で,半陰から日陰で,土壌は粘土から泥 土上に成立する。幅1 ~ 2m 長さ2m 程度のいずれも 小規模な群落である。 ハチジョウススキ下位単位はシバが優占するシバ ファシス(表9
),ギョウギシバが優占するギョウギシ バファシス(表9
),いずれも優占しない群落に識別さ れる。シバファシスはかつて公園造成が行われてシバ が張られた陽地で,シバ張り後時間が経過し,26種の 植物が侵入し群落を形成している。ギョウギシバファ シスは,車両等の往来が多くない未舗装道路の縁で ギョウギシバがびっしり生える群落で,道路中心部近 くでは他植物の混入はなく,道路末端では路傍雑草が 混入する。 そのほかリュウキュウトロロアオイが群生する群落 やコウライシバの被度の高い群落などもハチジョウス スキ下位単位に含めた。 ㊲ ハイキビ群落(表9
) 湿潤な路傍でハイキビが優占する群落が確認され た。群落の高さは1m 前後でびっしりとハイキビが生 え,ギョウギシバ,リュウキュウチク,タチスズメノ ヒエが常在する。また,水田放棄地にも密生した群落 をつくる。 ㊳ アオチカラシバ群落(表9
) チカラシバやアオチカラシバは路上植物群落の構成 種として知られる。本群落は牧場辺の路傍に高さ1m 前後のアオチカラシバがびっしりと生える。ギョウギ シバ,タチスズメノヒエ,リュウキュウチク等が随伴 する。 (3
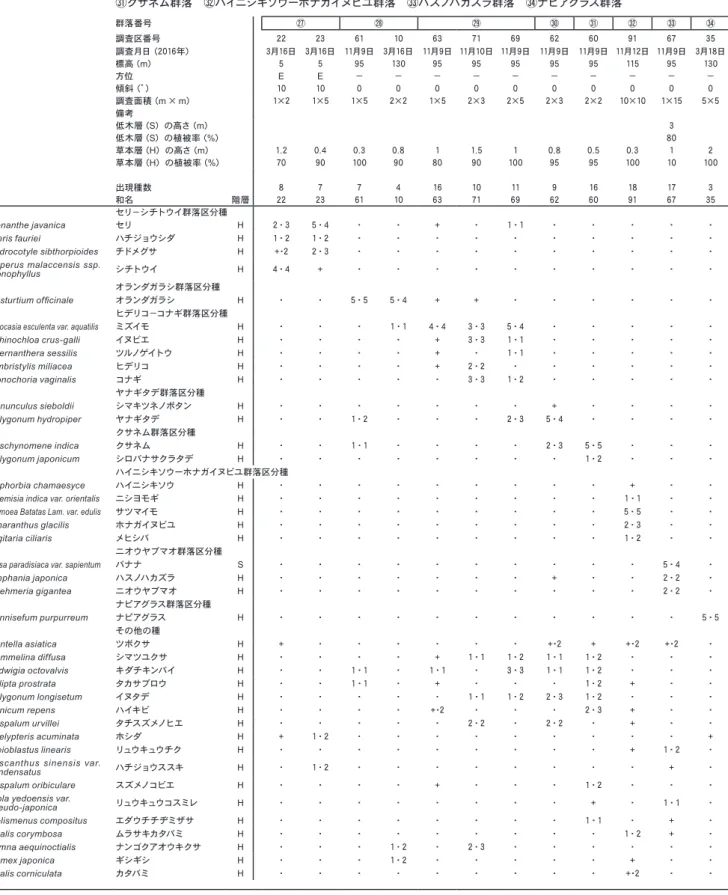
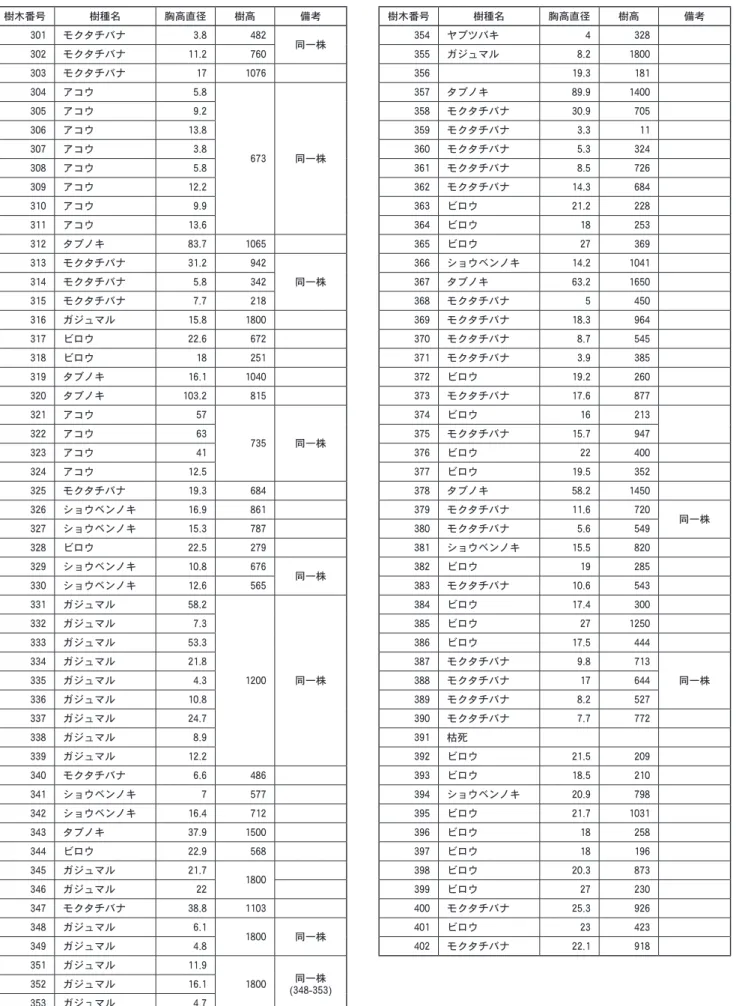
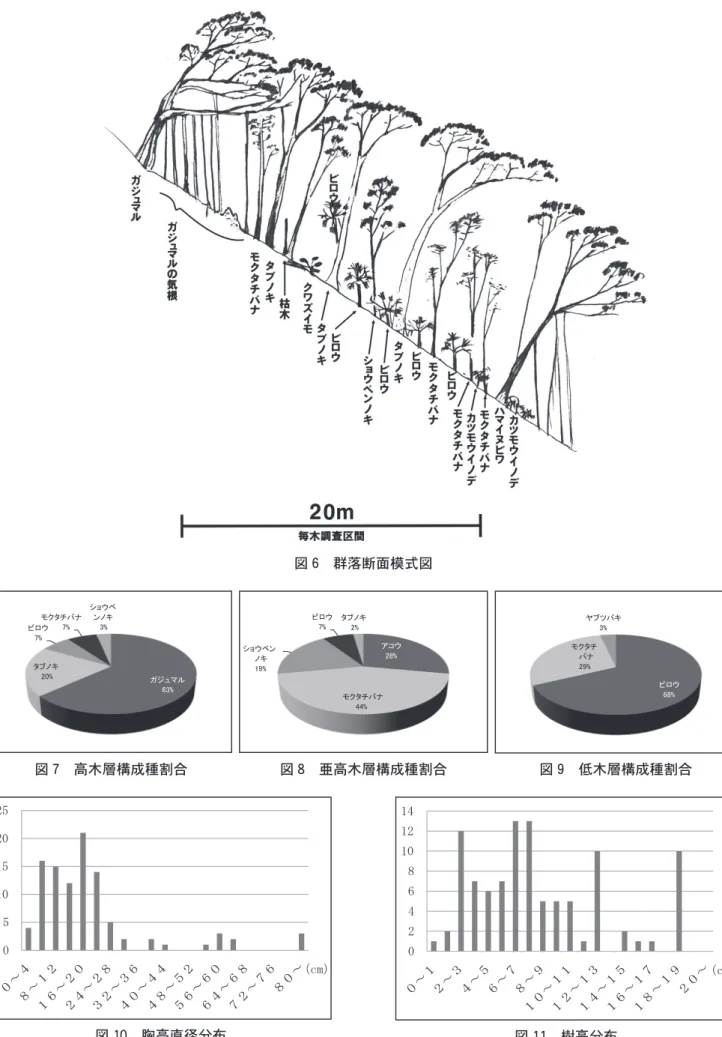
)毎木調査・樹冠投影図作成 今回群落区分されたムサシアブミ−タブノキ群集ヤ ブツバキ亜群集の典型的な林分にコドラードを設けて 毎木調査,樹冠投影図作成,群落断面模式図作成を実 施した。 20m 四方の方形枠を設定し,地上120㎝前後で,直 径が5㎝以上の個体について識別番号を付し,樹種, 樹高,胸高直径を記録する毎木調査表(表10
)とその表
10
毎木調査票 樹木番号 樹種名 胸高直径 樹高 備考 301 モクタチバナ 3.8 482 同一株 302 モクタチバナ 11.2 760 303 モクタチバナ 17 1076 304 アコウ 5.8 673 同一株 305 アコウ 9.2 306 アコウ 13.8 307 アコウ 3.8 308 アコウ 5.8 309 アコウ 12.2 310 アコウ 9.9 311 アコウ 13.6 312 タブノキ 83.7 1065 313 モクタチバナ 31.2 942 同一株 314 モクタチバナ 5.8 342 315 モクタチバナ 7.7 218 316 ガジュマル 15.8 1800 317 ビロウ 22.6 672 318 ビロウ 18 251 319 タブノキ 16.1 1040 320 タブノキ 103.2 815 321 アコウ 57 735 同一株 322 アコウ 63 323 アコウ 41 324 アコウ 12.5 325 モクタチバナ 19.3 684 326 ショウベンノキ 16.9 861 327 ショウベンノキ 15.3 787 328 ビロウ 22.5 279 329 ショウベンノキ 10.8 676 同一株 330 ショウベンノキ 12.6 565 331 ガジュマル 58.2 1200 同一株 332 ガジュマル 7.3 333 ガジュマル 53.3 334 ガジュマル 21.8 335 ガジュマル 4.3 336 ガジュマル 10.8 337 ガジュマル 24.7 338 ガジュマル 8.9 339 ガジュマル 12.2 340 モクタチバナ 6.6 486 341 ショウベンノキ 7 577 342 ショウベンノキ 16.4 712 343 タブノキ 37.9 1500 344 ビロウ 22.9 568 345 ガジュマル 21.7 1800 346 ガジュマル 22 347 モクタチバナ 38.8 1103 348 ガジュマル 6.1 1800 同一株 349 ガジュマル 4.8 351 ガジュマル 11.9 1800 同一株 (348-353) 352 ガジュマル 16.1 353 ガジュマル 4.7 樹木番号 樹種名 胸高直径 樹高 備考 354 ヤブツバキ 4 328 355 ガジュマル 8.2 1800 356 19.3 181 357 タブノキ 89.9 1400 358 モクタチバナ 30.9 705 359 モクタチバナ 3.3 11 360 モクタチバナ 5.3 324 361 モクタチバナ 8.5 726 362 モクタチバナ 14.3 684 363 ビロウ 21.2 228 364 ビロウ 18 253 365 ビロウ 27 369 366 ショウベンノキ 14.2 1041 367 タブノキ 63.2 1650 368 モクタチバナ 5 450 369 モクタチバナ 18.3 964 370 モクタチバナ 8.7 545 371 モクタチバナ 3.9 385 372 ビロウ 19.2 260 373 モクタチバナ 17.6 877 374 ビロウ 16 213 375 モクタチバナ 15.7 947 376 ビロウ 22 400 377 ビロウ 19.5 352 378 タブノキ 58.2 1450 379 モクタチバナ 11.6 720 同一株 380 モクタチバナ 5.6 549 381 ショウベンノキ 15.5 820 382 ビロウ 19 285 383 モクタチバナ 10.6 543 384 ビロウ 17.4 300 385 ビロウ 27 1250 386 ビロウ 17.5 444 387 モクタチバナ 9.8 713 同一株 388 モクタチバナ 17 644 389 モクタチバナ 8.2 527 390 モクタチバナ 7.7 772 391 枯死 392 ビロウ 21.5 209 393 ビロウ 18.5 210 394 ショウベンノキ 20.9 798 395 ビロウ 21.7 1031 396 ビロウ 18 258 397 ビロウ 18 196 398 ビロウ 20.3 873 399 ビロウ 27 230 400 モクタチバナ 25.3 926 401 ビロウ 23 423 402 モクタチバナ 22.1 918図
3
樹冠投影図図
6
群落断面模式図 図10
胸高直径分布 図11
樹高分布 図7
高木層構成種割合 ガジュマル 63% タブノキ 20% ビロウ 7% モクタチバナ 7% ショウベ ンノキ 3% 図8
亜高木層構成種割合 アコウ 28% モクタチバナ 44% ショウベン ノキ 19% ビロウ 7% タブノキ 2% 図9
低木層構成種割合 ビロウ 68% モクタチ バナ 29% ヤブツバキ 3% (cm) 0 5 10 15 20 25 0 2 4 6 8 10 12 14 (cm)図
13
神山 図14
アコウにとりつかれた タブノキの巨木図
12
平島現存植生図個体の林冠が地表を覆っている形状についての樹冠投 影図(図
3
),方形枠中の上部から下部に向かって線を のばし,その線上にかかる樹木や草本植物についての 模式図(図6
)である。 踏査した林分中に対象樹木は101個体あったが,樹 木種としてはアコウ,ガジュマル,モクタチバナ, ショウベンノキ,タブノキ,ビロウ,ヤブツバキなど 7種のみである。 ムサシアブミ−タブノキ群集では一般に構成種数は 多いが,本林分中では少ない。本林分中ではガジュマ ル,アコウが多数の気根を出し,気根が地上に到達し て成長して屹立している。このため,株数はガジュマ ルは2株しかないが上部でつながり,高木層において ガジュマルの個体数は63%を占める(図7
)。亜高木層 においてはタブノキに付着したアコウが同様に気根を のばしているため1株でありながらも占める割合は高 い(図8
)(図9
)。 胸高直径(表10
,図10
)を見ると80㎝を越える大径 木が3本(83㎝,90㎝,102㎝)あり,いずれもタブノ キで1株はアコウに締め付けられている。胸高直径分 布では特徴的な点として56㎝で小ピーク,16㎝で最大 ピーク,8㎝で次のピークが見られる。 一般の森林であれば,最小単位の胸高直径付近が最 も多くなるのが普通であるが,本林分では小木の個体 数は少ない。 また,樹高分布(図11
)においては,18-19mに10, 12-13m に10,7-8m に13,6-7m に13のピークがある。 18-19m はガジュマル,12-13m はアコウの気根が成木 化したものによる。一般的に森林においては低木の個 体数が多いもので2m 未満の樹木は多数あるのが普通 である。この林分には3本しか確認されていない。 上記の特異的な傾向はノヤギの被食によるものと考 えられる。低木層のビロウの葉はかじられているが, 亜高木層,高木層の葉には及んでいない。また,調査 期間中1日放置したテープは夜間に引き倒されていた し,早朝の調査ではノヤギのにおい,また,林内には 多数のノヤギの糞が確認されている。 (4
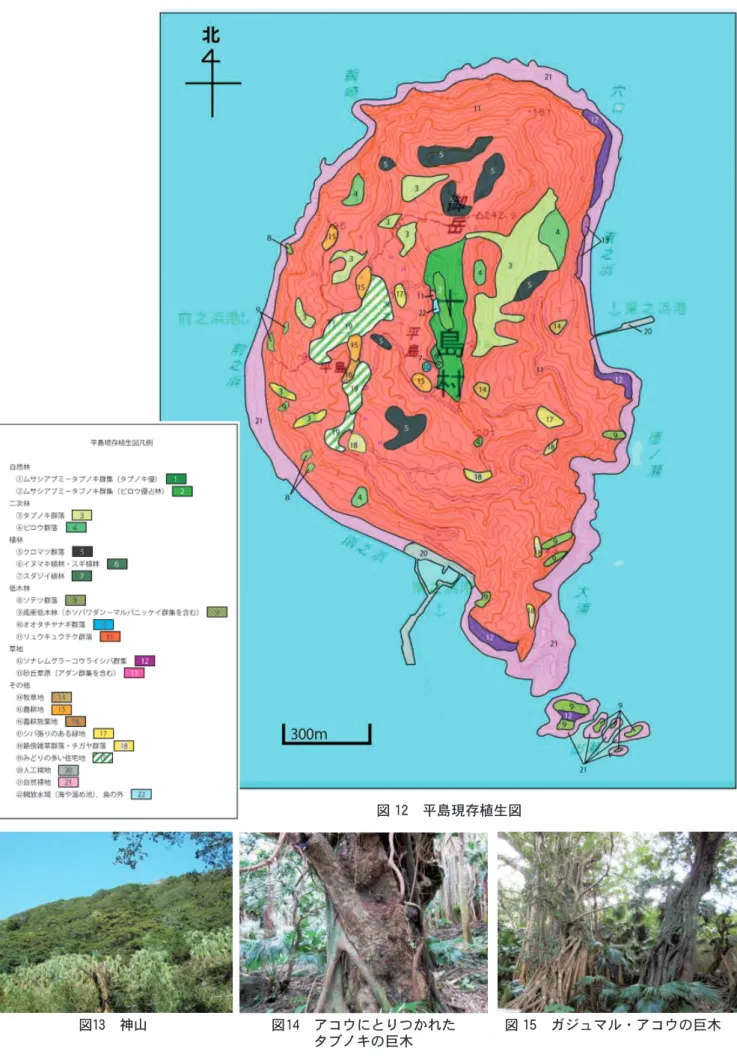
)現存植生図作成平島の現存植生を把握するため植物群落調査の知見 をもとに23の凡例で植生図を作成した(図
12
)。 植生分布の概要 自然林としてムサシアブミ−タブノキ群集が分布す る。本群集はタブノキが優占する林分とビロウが優占 する林分からなる。いずれも平島では神山と称される 集落の東部にある丘陵地で面積は4ha 前後である。タ ブノキが優占する林分は山脚部から山頂部にかけてで あり,ビロウが優占する林分はため池のある山脚部か ら中腹部にかけてである。 二次林の組成的には自然林と同じムサシアブミ−タ ブノキ群集と同じ群落のうちタブノキ群落およびは平 島の南北に延びる脊梁部のうち東部の中央部に分布す る。中腹部から標高200m 付近に広く分布する。ビロ ウ群落は東部中央部の山脚部や中腹部にやや広い群落 を占める。 植林としてクロマツ群落がかつては広く分布してい たが,マツクイムシ被害によって大半は枯死し残って いるものは島の脊梁部の南部と北部の山地部,水源地 付近である。なお,クロマツ植林の確認は空中写真と 現地調査を行ったが,大半のマツの活性がない。マツ クイムシによる枯死は今後も拡大し,数年後には植生 図には示されることがないほど小規模になることが予 想される。 イヌマキ植林,スギ植林,スダジイ植林とも狭小な 群落で中央部の神山に隣接する。また,イヌマキ植林 は東部の凹地部にも狭小な群落が見られる。スダジイ 植林は神山に隣接するが狭小である。 低木林としてソテツ群落,風衝低木林(ホソバワダ ン−マルバニッケイ群集を含む),オオタチヤナギ群 落があるがいずれも狭小である。島の80%以上はリュ ウキュウチク群落となり,海岸部から山頂部,耕作放 棄地などにも進出している。 草地にはソナレムグラーコウライシバ群集,アダン 群集,砂丘草原,牧草地,耕作地,シバ草地などがあ る。アダン群集,砂丘草原は東の浜に分布するが狭小 であるため凡例を統一した。ソナレムグラ−コウライ シバ群集は島の南部や東部,北東部の岩礁上に分布す る。ノヤギによる踏圧,掘り起こしによって破壊され て自然裸地に変化しているところとの判別は困難であ るが両者は接している。 耕作地は人口減とともに大きく減少しており集落の 近辺の耕作地は耕作放棄地その後リュウキュウチク林 に遷移している。学校や事業所も集落とともにみどり の多い住宅地とした。港湾施設は人口裸地とし,岩礁 海岸等の無植生部および自然災害,ノヤギによる土砂崩壊部は自然裸地とした。
4
考察 (1
)神山の森林の文化財的価値について ア 自然林として重要 温帯と亜熱帯気候の狭間にあるトカラ地域は日本で は同緯度に類がなく,この地域の自然林は重要である。 平島の神山は島の中央部にあり,南東側に集落を守る 衝立のように標高150-220m の範囲にある。神山の自 然林の広さは3-5ha 前後で植物社会学的には海岸型タ ブ林の1つでムサシアブミ−タブノキ群集の大隅半島 にあるものと類似する。その中にはタブノキの大径木 (胸高直径50−200㎝前後) ガジュマルやアコウが優 占するタブ林,ビロウが優占するビロウ林からなる。 タブノキの自然林は人が集中する低地部に多く全国的 に見ても少ない イ 神山として人を保護し,人に保護されてきた森 集落近くの丘陵はたきぎや農耕に使う肥料の供給源 として利用されることが多いが,平島では島の中央部 にあり,集落の後背の丘を神山と呼び神聖な場所とし て,樹木の伐採や落葉落枝の採集は行われてこなかっ た。 これは急峻な地形である丘の樹木伐採は土砂崩壊を 促すことを経験的に理解していたことや水田耕作や小 島での生活には安定的な水の供給は死活問題であり, 地域住民は森林の持つ貯水機能に気付いていたため神 山と名付け森を保護してきたものと推量される。神聖 な場所と位置づけることにより樹木の伐採や水質汚染 を免れ森が維持され,結果として村人の生命と財産, 健康を守ってきた。 また,南九州から沖縄にかけては,神聖な森が「モ イドン」(指宿市,錦江町,南大隅町など)「ガロウ 山」(種子島)「奥岳」(屋久島),「神山」(中之島,悪 石島,宝島,奄美大島など),「御嶽」(沖縄~八重 山)と呼ばれており,それぞれ地域の文化の基層とな り,重要な景観をつくっていた。それらは人の侵入に 制限を加え,地域本来の自然植生が保たれてきたが, 近代化とともに信仰が希薄になり,近年の開発事業に よって失われつつある中に,平島の神山は自然林とし て残っており,次世代に残すべき日本の自然を語る貴 重な文化財の1つと言える。 (2
)ノヤギによる植生被害について 平島ではノヤギが多数生息し,牧場内や道路,海岸 部,リュウキュウチク林内,神山内でも出くわすこと が多いが,個体数については行政も把握していない。 フェリーが停泊する南之浜港に向かうときに 南端に ある離れ瀬の出瀬と平島の南部および東側の岩礁地帯 の植生でノヤギの植生破壊状況が確認できる。風衝の 強い出瀬が風衝低木林やコウライシバ群落に覆われて いるのに対して平島の岩礁地帯は植生が剝がされた自 然裸地が広い面積確認できる。また,南之浜港に向か う南部の傾斜地が土砂崩壊を起こし復旧工事が行われ ている。 ノヤギやシカなどの大型草食獣が住む地域ではその 密度によって植生変化が起こり,密度の低いところで は好餌植物が減少し,増加ととともに,不嗜好植物の 群落が形成され,その後不嗜好植物も被食され,さら に密度が高くなると,ついには土壌浸食が深刻化し土 砂崩壊に繋がる。 今回の調査でも海岸沿地に普通に見られるホソバワ ダン,ボタンボウフウ,ツワブキ,トカラノギクがあ まり見られないが,集落内に設置してあるアンテナの 柵内には,ノヤギの食害を受けないため,ホソバワダ ンの大株が花を付けていた。また,海岸部にはニオウ ヤブマオウ群落,ハマゴウ群落,マルバハダカホオズ キ群落,ハスノハカズラ群落等の不嗜好植物群落が形 成されている。 また,神山や植林地内では低木層,草本層の植生は 貧弱になっており林床植物はカツモウイノデ,クワズ イモ,ムサシアブミ等の不嗜好植物が群落を築き,そ の他の種はほとんど見あたらない。また,不嗜好植物 を含む多くの植物に食痕が残る。斜面上で不嗜好植物 が育っていないところでは裸地状態になり降雨によっ て土壌浸食も起こり,将来的には規模の大きな土砂崩 れや森林崩壊も懸念される(寺田,1999)。森林生態 系にとって危機的な状況になっているが,ノヤギの食 料の大半はリュウキュウチクと考えられる。リュウ キュウチクは5月から6月にかけて筍を出すが,その後 数量は減少するものの筍は出し続ける。森林破壊が起 こったところにも侵入しリュウキュウチク群落は増加 していく。 ノヤギは文化財的価値のある小型のトカラヤギはほ とんどおらず大型の個体(ザーネン種)ばかりを確認 している。森林生態系の保全のため以上に人々の生活 を守るためにもノヤギ対策が必要である。(
3
)海岸浸食による植生破壊について 平島は亜熱帯地域にありながら離水サンゴが少ない ため,イソマツ群落やイソフサギ群落は確認されない。 砂丘地植生はコウボウシバ群落,ハマゴウ群落,アダ ン群落の他は確認されなかった。亜熱帯の海岸であれ ば,他にハマニガナ群落やコオニシバ群集,ハマアズ キ−グンバイヒルガオ群集,ツキイゲ群落等は普通に 見られるが,これまで確認もされていない。 フロラを見ると,これまで確認されていたイワタイ ゲキ,ハマアズキ,シママンネングサ,クサトベラが 確認されていない。これらは現在進行中の海岸浸食の 影響が考慮される。 口之島以南の琉球列島の砂丘地植生は一般的には以 下のような順で帯状分布を示す。 ①無植生帯②ハマアズキ−グンバイヒルガオ群集③ク ロイワザサ−ハマゴウ群集④モンパノキ−クサトベラ 群集⑤アダン群集 平島ではかつて,アダン群集まで砂があり,砂丘植 生が発達していたが,現在はアダン群集も根のところ は岩が裸出している。また,アダン群集より海側に形 成されるクサトベラ群落(モンパノキ−クサトベラ群 集)は壊滅し,クロイワザサ−ハマゴウ群集(ハマゴ ウ群落)からは砂が激減し,表層に生えるクロイワザ サは消滅し,ハマゴウは礫地に取り残され,クサトベ ラは生育地を見いだせない。ハマアズキーグンバイヒ ルガオ群集で長い地上茎を持つグンバイヒルガオは礫 地や岩上地に生育地をシフトするが,ハマアズキは砂 丘地のみの生活域のため生育が限定される。クロイワ ザサ−ハマゴウ群集のクロイワザサも同様である。砂 の流出がありアダン群集より海側の群落構成種に大き な影響があったものと思われる。 また,前之浜,南の浜,東の浜等地名に浜が着いた ところはかつて砂丘地があったところであるが,現在, 砂丘地は東の浜にわずかに残り,ほかは砂が渚からは 消え礫浜に変貌している。このため砂丘地植物,砂丘 地植生は今後とも減少していくものと思われる。一方 今回増加した種としてマルバアカザ,コダチボタンボ ウフウ,ヘンリーメヒシバ,ハナカモノハシ,コオニ シバ,コウシュンシバ,シオカゼテンツキなどがあり, これまで調査機会が十分確保されていなかったものと 思われる。 今後も継続的な調査によって平島の自然の変化を植 生の変動の面からも記録していくことが重要である。 (4
)社会構造の変化と植生の変化 過疎化,高齢化,産業構造の変化等様々な社会構造 の変化によって地域の植生は変わってくる。 平島では昭和20年代まで人口が多く生活維持のため 焼き畑耕作を行い,その結果リュウキュウチク群落が 多くまた,水田も広く,水田雑草等も多様であった。 人口流出,余剰米等もあって水田耕作,焼き畑等は行 われなくなり,焼き畑は牧場,マツ植林地に変化し, そのマツ林はマツクイムシの侵入によってリュウキュ ウチク林に変化し,現在は8割以上がリュウキュウチ ク林となっている。植物相も帰化植物が増え,水田雑 草をはじめ湿地性の植物は減少している。 謝辞 今回の調査では多数の方々にお世話いただいた。 現地調査では平島自治会会長の日高利成氏,民宿経 営の用澤満男氏,日高峰夫氏には島内での侵入困難地 等の案内をしていただいた。また,地域の自然の変遷 や植物方言に関する貴重な情報もいただいた。十島村 立平島小・中学校教頭の中村幸一郎氏,教諭の宮田直 彦氏には急峻な斜面での毎木調査で測定記録等をお手 伝いいただいた。屋久島環境文化村研修センターの山 田島崇文氏には毎木調査のデータ処理を,鹿児島大学 教育学部准教授川西基博氏には樹冠投影図作成の労を 執っていただいた。元横浜国立大学教授大野啓一氏に は群落単位の所属に関して丁寧な指導をいただいた。 十島村および十島村教育委員会には現地滞在の諸事象 に便宜を図っていただいた。記して厚く感謝を申し上 げる。 参考・引用文献 Braun-Blanquet, J.(1964)Pflanzensoziologie.3.Aufl.865pp. 初島住彦(1986)改訂 鹿児島県植物目録,290pp.鹿 児島植物同好会,鹿児島. 初島住彦 (1991) 北琉球の植物,218pp. 朝日印刷 . 鹿児 島 . 初島住彦 (2004)九州植物目録,343pp.鹿児島大学学 研究総合博物館.鹿児島 . 日高和広・吉田昭穂(1971)宝島及び悪石島における 焼畑農業.鹿児島地理学会紀要,18:77-80. 平田浩(1995)植物 . 十島村誌:56-146. 宮脇昭 編著(1981) 日本植生誌 九州 , 473pp. 至文堂, 東京 .宮脇昭(1989)日本植生誌沖縄・小笠原,675pp. 至文堂, 東京. 宮脇昭・奥田重俊・藤原陸夫編(1994)日本植生便覧 (改定新版),871pp.至文堂,東京 . 森田康夫(2005)トカラ列島平島の植物採集記録.鹿児 島県立博物館研究報告,24:20-27. 成尾英仁 (1995)トカラ列島の地質.十島村誌 :4-48. 大野照好 (1991)トカラ列島の植生,トカラ列島学術 調査報告書 :30-56. 迫静男 (1991)トカラ列島の植物相,トカラ列島学術 調査報告書 :57-117. 志内利明・堀田満(2015) トカラ地域植物目録 . 368pp. 鹿児島大学総合研究博物館. 鈴木邦雄(1979)琉球列島の植生学的研究.横浜国大環 境科学研究センター紀要,5 :87-160. 鈴木時夫(1953)大隅半島中央部の森林植生の群落学 的研究 . 日本林學會誌 35(5):178-179. 立久井昭雄(1991)トカラ列島平島の植物.鹿児島県立 博物館研究報告,10: 11-20. 寺田仁志 (1997) トカラ列島・中之島の植生と現存植 生図.鹿児島県立博物館研究報告,16:1-48. 寺田仁志 (1999) 臥蛇島の現存植生と植物相(野生化 したシカ・ヤギが植生に及ぼす影響).南日本文 化,33:59-108. 寺田仁志 ・ 大屋哲(2007)鹿児島県喜界島の隆起珊瑚 礁上植物群落について,鹿児島県立博物館研究報 告,26:45-77.