Title
非冬眠動物における冬眠様低体温誘導に関する研究( 本文
(Fulltext) )
Author(s)
島岡, 弘樹
Report No.(Doctoral
Degree)
博士(獣医学) 甲第553号
Issue Date
2020-03-13
Type
博士論文
Version
ETD
URL
http://hdl.handle.net/20.500.12099/79354
※この資料の著作権は、各資料の著者・学協会・出版社等に帰属します。非冬眠動物における
冬眠様低体温誘導に関する研究
2 0 1 9 年
岐阜大学大学院連合獣医学研究科
(岐阜大学)
島
岡 弘 樹
非冬眠動物における冬眠様低体温誘導に関する研究
2
目次
略語一覧・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・ 3 諸言・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・ 4 実験材料及び実験方法・・・・・・・・・・・・・・・・・・・・・・・・・・・ 6 第1 章・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・11 中枢性アデノシンA1 受容体の活性化によるラットの冬眠様低体温誘導 結果 考察 第2 章・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・22 イソフルランによるラットの冬眠様低体温誘導 結果 考察 第3 章・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・45 イソフルランによるマウスの冬眠様低体温誘導 結果 考察 総合考察・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・65 結論・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・71 謝辞・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・72 文献・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・733
略語一覧
ALT: alanine aminotransferase AST: aspartate aminotransferase BUN: blood urea nitrogen CHA: N6-cyclohexyladenosine
CPT: 8-cyclopentyl-1,3-dimethylxanthine LDH: lactate dehydrogenase
4
諸言
哺乳動物の体温は一定の範囲内で維持されており,環境温度が変化しても,体温が大 きく変動することはない。そのため,何らかの要因で体温が低下することで,不整脈(3) や免疫機能の低下(60)などの有害な事象が生じる。一方で,低体温状態には医学的な メリットがあることも明らかとなっており,臨床分野において,低体温療法として人工 的に低体温に誘導することが行われている。例えば,心筋梗塞(72),脊髄損傷(33), 脳外傷(13)による傷害の軽減や予後を改善することが報告されている。 この低体温療法で適用される体温は,一般的に32℃から 34℃の間である(21)。しか しながら,低体温療法における最適な体温については議論の余地がある。Busto らは, ラットにおいて30℃の体温が 36℃の体温に比べて虚血による脳傷害を軽減すると報告 している(8)。一方,35-36℃の極めて軽度の低体温と 33℃の低体温による効果が同等 であるという報告もなされている(46,64)。様々な議論があるものの,低体温療法の 治療効果には低温による代謝の抑制が重要であることを考慮すると,代謝の抑制が温度 に依存する(17)ことから,より低い体温を用いた低体温療法はより治療効果が高くな ると考えられる。実際に臓器保存の分野では,低体温療法の一般的な温度である 33℃ 前後の体温よりも著しく低い4℃での臓器保存が行われている(26)。そのため,低体温 療法でもより低い体温に誘導することでより良い治療成績を得られることが期待され る。 しかし,ヒトを含む非冬眠動物は体温が 20℃以下になると,心室細動や完全房室ブ ロックなどの不整脈が生じ,最終的に死亡する(19,34,37)ため,極度の低体温状態 に誘導することは容易ではない。対照的に,冬眠動物は低体温に対して高い耐性を持つ ことが知られている。シリアンハムスターやジリスなどの冬眠動物は,冬眠期間中に環 境温度に近しい程度まで体温が低下しても死亡することはない(10,25)。また,冬眠 時には通常時と比べて著しく心拍数が低下するものの,正常な洞調律は維持されている5 (30,56,58)。この冬眠の仕組みを応用することで,非冬眠動物を冬眠の様な極度の 低体温状態に安全に誘導することが可能となれば,より良い低体温療法の開発につなが ると考えられる。 本研究は,低体温の持つ医学的に有用な特性を医療に応用することを目的に,非冬眠 動物を冬眠様の極度の低体温状態に誘導する方法の検討を行った。第1 章では,冬眠の 体温制御に着目した低体温誘導法を試みた。冬眠時の体温低下には中枢のアデノシン A1 受容体の活性化が重要であると報告されている(76)。そこで,中枢のアデノシン A1 受容体を薬理学的に活性化することで,非冬眠動物であるラットを冬眠様の極度の低体 温に誘導することが可能かについて検討した。この方法により 15℃まで体温を下げる ことが可能となったが,事前の薬剤投与のための外科手術が必要となることは,臨床応 用では不都合となり得る場合がある。そこで第2 章では,事前の処置を必要としない吸 入麻酔を用いて,ラットの冬眠様低体温誘導が可能かについて検討した。第3 章では, 同じ非冬眠動物であるマウスにおいても冬眠様低体温誘導が可能であるかについて検 討した。本研究の結果から,非冬眠動物においても適切な手法を用いることにより,冬 眠様の極度の低体温状態に誘導し,低体温状態を6 時間まで維持することが可能である ことが明らかとなった。これにより冬眠様の極度の低体温の持つ医療効果について研究 することが可能となった。
6
実験材料及び実験方法
実験動物 8-10 週齢の雄の Sprague-Dawley ラットと 10-12 週齢の雄の ddY マウスを使用した。 動物は日本SLC(静岡,日本)より購入した。明暗周期 12 時間,室温 22℃の環境下で 飼育した。水道水と飼料(MF;オリエンタル酵母工業,東京,日本)を自由に摂取で きるようにした。本研究における動物実験は全て,岐阜大学動物実験委員会の承認を 受けたものであり,岐阜大学動物実験取扱規程に従って実施した。 ラットの側脳室へのカニューレの設置 ラットにペントバルビタール(ソムノペンチル®;共立製薬,東京,日本)を腹腔内 投与(50 mg/kg)することにより麻酔し,脳固定装置(SR-5R-HT;ナリシゲ製作所,東 京,日本)にbregma と lambda が水平になるように固定した。切皮して頭蓋骨を露出し, bregma の位置を測定した。9 mm のガイドカニューレ(AG-9;エイコム,京都,日本) をbregma から AP: -4.0 mm L: 5.0 mm DV: 7.0 mm の位置に設置して固定した。術後 1 週 間の回復期間をおいてから実験に使用した。 アデノシンA1 受容体アゴニストの側脳室投与によるラットの低体温誘導 アデノシンA1 受容体アゴニストである N6-cyclohexyladenosin(CHA)は生理食塩水 に溶解し,濃度が4 nmol/μL になるように調製した。CHA の投与は 9 mm のインジェク ションカニューレ(AMI-9;エイコム,京都,日本)をポリエチレンチューブで接続し た10 μL のマイクロシリンジ(701 SNR;ハミルトン,ネバダ,アメリカ)を用いて行った。背部の皮下に設置したトランスポンダー(IPTT-300;BioMedic Data Systems,デ ラウェア,アメリカ)により皮下温度を計測した。22℃の環境下でラットの側脳室に CHA(20 nmol/head)を投与後,4℃の冷蔵庫にラットを移動させた。ラットの体温が低
7
下し,不動化した場合は,肛門から 5 cm の位置にサーミスタ(RET-3;Physitemp
Instruments,ニュージャージー,アメリカ)を挿入し,直腸温度を測定した。また,心 電図を計測するために四肢に記録用電極(OA213-067;ユニークメディカル,東京,日 本)を取り付けた。波形をアンプリファイア(DAM50;World Precision Instruments,フ ィラデルフィア,アメリカ)により増幅し,パワーラボ(AD Instruments,CO,名古屋, 日本)に記録した。 ペントバルビタール麻酔によるラットの低体温誘導 ペントバルビタールを腹腔内投与(50 mg/kg)することによりラットを麻酔した。肛 門から5 cm の位置にサーミスタを挿入し直腸温度を測定した。四肢に電極を取り付け て心電図の計測を行った。麻酔下でサーミスタと電極の設置を行った後,ラットを4℃ の冷蔵庫に移動させた。 イソフルラン麻酔と冷却によるラットの低体温誘導 気化器(HN-1070;夏目製作所,東京,日本)により気化したイソフルラン(ファイ ザー®;ファイザー,ニューヨーク,アメリカ)を濃度 2%,流量 1.0 L/分でマスクを用 いてラットに吸入させ,麻酔を行った。肛門から5 cm の位置にサーミスタを挿入し直 腸温度を測定した。また,四肢に記録用電極を取り付けて心電図の計測を行った。測定 機器の設置後,ラットを4℃の冷蔵庫に移動させた。低体温誘導中に特定の直腸温度に 到達した時点でイソフルランの濃度を調節した(第 2 章の結果参照)。体温を回復させ る場合には,室温22℃の環境に移動させ,白熱灯を用いてラットを保温した。別の実験 では,イソフルランによる低体温誘導中にアデノシンA1 受容体のアンタゴニストであ る8-cyclopentyl-1,3-dimethylxanthine(CPT)の側脳室内投与を行った。CPT は 100 mM の水酸化ナトリウム水溶液に溶かした後,リン酸緩衝生理食塩水で 12 nmol/μL となる ように希釈した。CPT の投与は,ラットを 4℃環境に移す 5 分前の時点と,4℃環境に
8 移動後に直腸温度が30℃,25℃,20℃,15℃にそれぞれ到達した時点で継続的に投与し た。1 回の投与量は 5 μL とした。 また,室温環境下でイソフルランにより麻酔をしたラットを,10℃に冷却したステン レスプレートに静置することによる低体温誘導も行った。15℃の低体温状態を維持する 場合にはプレートの温度を 14℃にした。体温を回復させる場合には,37℃に保温した プレートに移動させた。 イソフルラン麻酔と冷却によるマウスの低体温誘導 気化したイソフルランを濃度 2%,流量 1.0 L/分でマスクを用いてマウスに吸入させ て麻酔を行った。肛門から 2.5 cm の位置にサーミスタを挿入し直腸温度を測定した。 四肢に記録用電極を取り付けて心電図を記録した。測定機器の設置後,10°C に冷却し たステンレスプレートに静置した。直腸温度に応じてイソフルランの濃度を調節した (第3 章の結果参照)。15℃の低体温を維持する場合は,プレートの温度を 14℃にした。 体温を回復させる場合には,37℃に保温したプレートに移動させた。 血液生化学値の測定と解析 生化学自動分析装置(DRI-CHEM 3500V;富士フイルム,東京,日本)を用いて, 血漿中のAST(aspartate aminotransferase),ALT(alanine aminotransferase),LDH (lactate dehydrogenase),BUN(blood urea nitrogen)の測定を行った。ラットにおいて
は低体温誘導前,直腸温度が15℃に到達した直後,3 時間または 6 時間の低体温状態 維持後,体温回復後,1 日後,1 週間後の時点において,同一の個体から経時的に左右 の鎖骨下静脈より採血を行った。マウスにおいては,通常体温,15℃到達直後,3 時 間または6 時間の低体温状態維持後,体温回復後,1 週間後の時点において,それぞ れ別個の個体から後大静脈より全採血を行った。血液は採血後に直ちに富士ヘパリン チューブ(富士フイルム,東京,日本)に移し,4℃,1,200 × g,10 分の条件で遠心
9 分離をし,血漿を回収した。血漿は測定に供するまで-30℃で保存した。低体温誘導 による体温変化と採血時点の模式図を図1 として示した。 測定した血液生化学値を用いて,各採血時点での血液生化学値の平均値を算出し た。体温回復による影響,低体温維持による影響,低体温維持と体温回復による影響 を明らかにするために,下の式を用いて計算を行った。 (体温回復による影響)=15℃到達直後に体温を回復させた時点(図 1③)の血液 生化学値の平均値-15℃到達時点(図 1②)の血液生化学値の平均値 (低体温維持による影響)=低体温状態を3 時間または 6 時間維持した時点(図 1 ④)の血液生化学値の平均値-15℃到達時点(図 1②)の血液生化学値の平均値 (低体温維持と体温回復による影響)=低体温状態を3 時間または 6 時間維持した 後に体温を回復させた時点(図1⑤)の血液生化学値の平均値-15℃到達時点(図 1 ②)の血液生化学値の平均値 統計処理 数値は平均値±標準偏差で表した。二群間の比較は Student の t 検定,または対応の あるt 検定を使用した。多群間の比較には一元配置分散分析または二元配置分散分析を 行った後に,Tukey-Kramer 法により群間の有意差を事後検定した。統計学的有意差の基 準としてp<0.05 を用いた。
10 図1 低体温誘導による体温変化と採血時点の模式図 低体温誘導中の体温変化と採血時点を模式図として示した。縦軸は直腸温度、横軸は低 体温誘導開始からの経過時間を示す。○と数字は採血時点を示している。①低体温誘導 前の時点(対照群),②15℃到達時点(低体温維持時間 0h),③15℃到達直後に体温を回 復させた時点,④低体温維持後(低体温維持時間 3h または 6h),⑤低体温維持後(3h または 6h)に体温を回復させた時点,⑥低体温誘導から 1 日後の時点,⑦低体温誘導 から1 週間後の時点
40
直腸温度
(ºC
)
0
経過時間
15
2
1
4
3
6
7
0
5
11
第
1 章
中枢性アデノシン
A1 受容体の活性化によるラットの冬眠様低体温誘導
冬眠動物であるシリアンハムスターとジリスにおいて,中枢のアデノシンA1 受容体 が冬眠の導入と維持に重要な役割を果たす(36,76)。これと一致するように,シリア ンハムスターにおいて中枢のアデノシン A1 受容体を薬理学的に活性化し,4℃の環境 下に置くことにより,冬眠と同様の極度の低体温状態に誘導することが可能であると報 告されている(59)。また,非冬眠動物であるラットにおいても,15℃の環境下で側脳 室にアデノシンA1 受容体のアゴニストである CHA を投与することにより,25℃の低 体温状態に誘導することが可能であると報告されている(78)。このため,中枢のアデ ノシンA1 受容体の活性化は,冬眠動物と非冬眠動物に関わらず,体温低下の機能を持 つことが考えられる。 また,シリアンハムスターは,ペントバルビタール麻酔と冷却を組み合わせることに より強制的に低体温状態に誘導することも可能である(59)。しかし,この方法では薬 理学的に中枢のアデノシンA1 受容体を活性化した場合や,自然の冬眠時には見られな い房室ブロックが生じており,また,非冬眠動物の低体温時に見られる異常心電図であ るJ 波(27,53)も生じている。このことから,中枢のアデノシン A1 受容体の活性化 には,体温の低下をもたらす作用に加えて,低体温時に正常な心拍動を維持する機能を 持つ可能性が示唆される。非冬眠動物が低体温になった場合,20℃以下では心停止が生 じる(34)ため,冬眠動物のような極度の低体温状態に誘導することはできないと考え られている。しかし,非冬眠動物であるラットにおいても,中枢のアデノシンA1 受容 体に低体温時の心拍動の保護機能があるならば,CHA の側脳室内投与による低体温誘 導を行うことで,Tupone らが報告した 25℃(78)よりも低い,20℃以下の極度の低体 温状態にラットを誘導できる可能性がある。12
本章では,ラットの側脳室内にアデノシンA1 受容体のアゴニストである CHA を投
与し,4℃の環境下に移動させることにより,冬眠の様な極度の低体温状態に誘導する ことが可能かについて検討した。
13
結果
CHA の側脳室内投与と冷却によるラットの冬眠様低体温誘導 室温22℃の環境下で CHA を側脳室内に投与した後,直ちに室温 4℃の冷蔵庫にラッ トを移動させた場合,ラットは震えを起こし,体温の低下は起きなかった。CHA の投 与後,室温22℃の環境下に静置した場合,1 時間以内にラットの活動性の低下が観察さ れた。この状態ではラットの皮下温度が平均1℃程度低下した。活動性の低下と皮下温 度の減少を確認後,ラットを室温22℃の環境から,4℃の冷蔵庫に移動させた場合,ラ ットは震えを起こすことはなかった。ラットの体温は時間経過に伴い低下し,20℃以下 の体温でも生存した(図2)。しかし,4℃環境に置き続け,直腸温度が 14℃以下まで低 下すると心停止を起こした。そのため,心停止がみられなかった最低体温である直腸温 度15℃の状態をラットの冬眠様低体温とした。 ラットの体温が 25℃付近になると不動化が起きたため,サーミスタと電極を取り付 けて直腸温度と心電図の記録を行った。25℃の心拍数は 128 ± 50 回/分であった。心拍 数は,直腸温度の低下に伴って減少した。直腸温度が15℃の時点での心拍数は 47 ± 9 回 /分であった。図 3a と 3b に示すように,どの体温においても,正常な洞調律は基本的に 維持されていた。しかし,時折心拍のスキップが観察された(図3c)。 CHA による低体温状態からの回復 CHA による誘導した低体温からラットを回復させることが可能かを明らかにするた めに,ラットの体温が15℃に到達した直後に,ラットを 22℃の環境下に置き,白熱灯 で温めることにより体温を回復させた。心拍数は体温が上昇するにつれて増加した。体 温の回復の途中において,ラットは正常な洞調律を基本的に維持していたが,心拍のス キップが時折観察された(図3d)。体温回復の開始直後は,ラットは不動化したままだ ったが,体温が30℃付近になると自発的な行動と震えが生じた。14 ペントバルビタールによるラットの低体温誘導 ラットにペントバルビタール麻酔と冷却を施すことにより,中枢のアデノシンA1 受 容体の活性化がない状態で低体温に誘導した。ペントバルビタールの腹腔内投与後に, 4℃の冷蔵庫に移動させたところ,経時的に体温が低下した(図 4a)。体温が低下するに つれて心拍数は減少した。(図4b)。しかし,ラットの体温が 22.5℃を下回ると,心拍数 が急激に低下した(図4b)。25℃までは洞調律に異常は生じなかったが,心拍数が急激 に減少する直前に,6 匹中,3 匹のラットで完全房室ブロックが,1 匹のラットで心室 細動が観察された(図4c)。異常心電図の有無に関わらず,全てのラットが心停止を起 こした。 ペントバルビタールがCHA による低体温誘導に与える効果 CHA によるラットの低体温誘導で,正常な洞調律が維持された理由は,麻酔状態で は活性化されない何らかの機構が作動しているのではないかと仮説を立てた。この仮説 を検証するため,CHA により低体温に誘導中であるラットに,ペントバルビタールを 腹腔内投与した。6 匹中 2 匹のラットで心室細動が,3 匹のラットで房室ブロックが生 じた(図5)。1 匹のラットでは異常心電図が生じなかったが,全ての個体が最終的に心 停止を起こした。
15 図2 CHA による低体温誘導時のラットにおける皮下温度と直腸温度の変化 CHA(20 nmol/head)を側脳室内に投与したラットの皮下温度と直腸温度の典型例を示 す。ラットの活動性の低下と皮下温度の減少を確認後,環境温度(Ta)が 22℃の室内か ら4℃の冷蔵庫に移した。●と□はそれぞれ皮下温度と直腸温度を示す。直腸温度はラ ットが低体温により不動化した時点で,肛門からサーミスタを挿入することにより測定 した。直腸温度は時間経過と共に低下し,最終的に15℃に到達した。同様の結果が 9 匹 の別々のラットより得られた。15℃到達までの平均時間は 312 ± 88 分であった。
40
35
30
25
20
15
10
5
0
300
270
30
60
120
180
240
0
90
150
210
Ta:4ºC Ta:22ºCCHA 側脳室内投与
皮下温度
直腸温度
温度
(ºC
)
経過時間 (分)
16 図3 CHA による低体温誘導時のラットにおける心電図波形 CHA による低体温誘導中のラットの心電図波形の典型例を示す。P 波と QRS 群を矢頭 と矢印でそれぞれ示した。ラットは直腸温度が25℃(a),15℃(b)の時点で正常な洞 調律を維持していた。体温の低下中に時折,心拍のスキップがみられた(c)。また,体 温の回復時にも心拍のスキップが時折みられた(d)。同様の結果が 9 匹のラットより得 られた。
a
直腸温度25ºC 1 秒 1 m V 1 秒 1 m V 1 秒 1 m Vb
直腸温度15ºCc
体温低下中の心拍のスキップ SHI-391 心電図 (V) -10 -5 0 5 10 4:50 4:50.5 4:51 4:51.5 4:52 4:52.5 4:53 4:53.5 4:54 4:54.5 4:55 4:55.5 4:56 4:56.5 0:04:49.875 1 秒 1 m Vd
体温回復中の心拍のスキップ SHI-393 心電図 (V) -10 -5 0 5 10 2:15:29.8 2:15:30 2:15:30.2 2:15:30.4 2:15:30.6 2:15:30.8 2:15:31 2:15:31.2 2:15:31.4 2:15:31.6 2:15:31.8 2:15:32 2:15:32.2 2:15:32.4 2:15:32.6 2:15:32.8 2:15:33 2:15:33.2 2:15:33.4 2:15:33.6 2:15:33.8 2:15:34 2:15:34.2 2:15:34.4 2:15:34.6 2:15:34.8 2:15:35 2:15:35.2 2:15:35.4 2:15:35.6 2:15:29.71317 図4 ペントバルビタールによる低体温誘導時の直腸温度と心拍数と心電図 ペントバルビタールによる低体温誘導を行った,各ラットの直腸温度(a)と心拍数(b) の変化をそれぞれ示す。ペントバルビタール(50 mg/kg)を腹腔内投与したラットの直 腸温度は時間経過により低下した(a)。心拍数も直腸温度が 25℃付近に低下するまでは 経時的に低下した。直腸温度が22.5℃を下回ると,急激に減少した(b)。直腸温度 25℃ 付近までは異常な心電図は生じなかった(c)。対照的に,直腸温度が 22.5℃を下回ると, 6 匹中 1 匹のラットで心室細動が,3 匹のラットで完全房室ブロックが観察された(d)。 P 波と QRS 群を矢頭と矢印でそれぞれ示した。
c
心室細動 完全房室ブロック 400 300 200 100 0 20 36 38 直腸温度 (˚C) 心拍数 (b p m ) 34 32 30 28 26 24 22a
b
d
直腸温度25ºC 90 40 35 30 25 20 15 60 30 0 冷却時間(分) 直腸温度 (ºC ) 1 秒 1 m V 1秒 1 m V18 図5 CHA による低体温誘導時にペントバルビタールを投与したラットの心電図 CHA による低体温誘導中にペントバルビタールを腹腔内投与したラットの心電図を記 録した。P 波と QRS 群を矢頭と矢印でそれぞれ示す。異常な波形を示したラットの心 電図波形の典型例を示した。6 匹中 2 匹のラットが心室細動を示し,3 匹のラットが完 全房室ブロックを示した。 1 秒 1 m V
a 心室細動
b 完全房室ブロック
19
考察
ヒトを含む多くの哺乳類では体温が 20℃以下になると生存することができないとさ れている(19,34,37)。一方,冬眠動物であるジリスやシリアンハムスターは,冬眠 時に極度の低体温状態になるにも関わらず死亡することはない(10,25)。この冬眠の 制御には,中枢のアデノシンA1 受容体が重要であると言う報告がある(36,76)。実際 に,非冬眠時のシリアンハムスターにアデノシン A1 受容体のアゴニストである CHA を側脳室内に投与することにより,冬眠を誘発することが可能である(59)。第 1 章で はこの制御に着目し,非冬眠動物であるラットでも,中枢のアデノシンA1 受容体を薬 理学的に活性化することで,直腸温度 20℃以下の冬眠の様な低体温状態に誘導するこ とが可能かについて検討した。 室温環境下(22°C)で側脳室内への CHA 投与を行い,直ちにラットを 4℃の環境下 に移動させると,震えを起こし,体温が低下することはなかった。この結果はtupone ら の結果と矛盾する(78)。彼らは,環境温度 15℃に置いたラットの側脳室内に CHA を 投与すると体温低下が起きると報告している。この結果の違いの原因として,側脳室内 投与時の環境温度の違いが考えられる。今回の実験では,22℃環境下に置いたラットに CHA を投与した後に,4℃の環境下に移動させている。中枢のアデノシン A1 受容体の 活性化による低体温は,孤束核から生じる交感神経抑制経路が活性化することで,褐色 脂肪における熱産生が抑制されるためと考えられている(78)。低温環境が褐色脂肪の 活動を亢進することを考えると,22℃の環境から 4℃に移動させたことによる熱産生亢 進のシグナルが,アデノシンA1 受容体の活性化による熱産生抑制シグナルを上回った 可能性がある。実際に,CHA を投与後に室温で静置した場合は,ラットの活動性は低 下し,皮下温度も低下した。この状態を確認した後にラットを4℃の環境下に移動させ ることで,非冬眠動物であるラットに心停止を起こすことなく 15℃の極度の低体温状 態に誘導することに成功した。このことは,非冬眠動物においても冬眠の様な極度の低20 体温状態に誘導することが可能であることを示している。室温環境下で体温の低下を確 認した後に4℃の環境下に移した場合は,震えや体温の上昇が見られなかったことから, アデノシンA1 受容体の活性化を介した褐色脂肪の熱産生の抑制が十分に起きていると 考えられる。 シリアンハムスターを含む冬眠動物と異なり,非冬眠動物は 20℃以下の体温では心 拍動を維持できないと考えられている(19,37)。実際にペントバルビタール麻酔と冷 却による低体温誘導では,ラットに心停止が生じた。この結果は冬眠動物のものとは対 照的である。シリアンハムスターをペントバルビタール麻酔と冷却により低体温に誘導 した場合,心拍数は減少するものの心拍動は維持されることが報告されている(59)。 また,ラットの摘出した心臓は30℃から 16℃の温度で不整脈を起こし,16℃から 10℃ で心停止するが,冬眠動物であるジュウサンセンジリスの摘出した心臓はより低い温度 である10℃以下でも拍動する(7)。これらは,冬眠動物の心臓が非冬眠動物に比べて高 い低温耐性を持つことを明確に示している。しかしながら,CHA による低体温誘導で は 20℃以下の体温でも正常な洞調律が維持されたことから,ペントバルビタールによ る低体温誘導で生じたラットの心停止は,心臓の機能を維持することができる下限の温 度を下回ったためではないと考えられる。 CHA による低体温誘導中にペントバルビタールの腹腔内投与を行うと,異常心拍動 が生じた。CHA 単独による低体温誘導では生じないことから,CHA による心拍動の維 持機能は麻酔下の状態では機能しないことが示唆される。麻酔下の動物では全体的に自 律神経系が抑制されることから,この機能の少なくとも一部は自律神経系の調節が関与 していると考えられる。冬眠動物においては,冬眠中に自律神経系の調節が行われてお り,それによる心拍動の制御が行われているという報告がある(30,48,57,84)。自 律神経系の制御の破綻は不整脈や心筋傷害を引き起こすことが知られており,例えば, 急性の脳傷害の後には一過性の左心室の機能低下が生じる(63)。そのため,適切な自 律神経系の制御が,低体温状態の心拍動を維持するために重要であると考えられる。
21 CHA の側脳室内投与が迷走神経を介して徐脈を起こすこと(78)に加え,中枢のアデ ノシン A1 受容体が神経伝達物質の放出の抑制や細胞の過分極を起こすこと(82)を考慮 すると,この制御の一部には,迷走神経の興奮を抑制している神経の抑制が関与してい ると考えられる。 本章では,中枢のアデノシンA1 受容体を活性化することにより,非冬眠動物である ラットを冬眠様の極度の低体温状態に誘導することができることが明らかとなった。ま た,低体温状態において心拍動を維持するためには自律神経の適切な制御が重要である 可能性が示唆された。
22
第
2 章
イソフルランによるラットの冬眠様低体温誘導
前章では,冬眠に重要な役割を果たす中枢のアデノシン A1 受容体の機能に着目し, 非冬眠動物であるラットを,極度の低体温状態に誘導することができるかどうかについ て検討した。その結果から,ラットにおいて,中枢のアデノシンA1 受容体を薬理学的 に活性化し,冷却を行うことにより 15℃という極度の低体温状態に誘導する方法を確 立した。また,低体温誘導中に注射麻酔薬であるペントバルビタールを腹腔内投与する ことで,心停止が起こることから,低体温状態で心拍動を維持するためには自律神経系 の適切な制御が重要であると考えられた。 CHA を側脳室内に投与し冷却することにより,非冬眠動物であるラットを低体温状 態に誘導することが可能であったが,この方法では薬剤を側脳室内に投与するため,あ らかじめ脳に対する手術が必要となる。臨床分野では低体温療法は,脳梗塞などの虚血 性疾患の治療に応用されていることから,事前の脳手術が必要とすることが不都合とな りうる。そのため,可能であるのならば,より臨床応用がしやすい簡便な手法であるこ とが望ましい。 本章では,より臨床応用に適している,事前の外科手術を必要としない冬眠様低体温 誘導法の検討を行った。前章で示唆された低体温時の自律神経の制御の重要性に着目し, 麻酔の導入と覚醒が素早い吸入麻酔であるイソフルラン麻酔を用いることによる低体 温誘導法の確立について試みた。また,低体温療法では誘導の目標とする温度に加えて, 低体温を維持する時間についても重要である。そこで,ラットに重大な後遺症や傷害を もたらすことなく何時間まで低体温状態を維持し,回復させることが可能かについて検 討を行った。23
結果
イソフルランの吸入と冷却によるラットの冬眠様低体温誘導 ラットをイソフルラン麻酔下(イソフルラン濃度 2%)で 4℃の冷蔵庫に移すと,ラ ットの直腸温度は時間経過と共に低下した。ラットの直腸温度が27.5℃まで低下した時 点でイソフルランの吸入を停止した場合,5 匹全てのラットが震えを行った。低温環境 下にも関わらず,これらのラットの直腸温度は回復した(図6)。直腸温度が 25℃の時 点でイソフルランの吸入を停止した場合,5 匹中 3 匹のラットが自発的な行動を示し, 体温を回復させた(図7)。1 匹のラットは,一時的に直腸温度を回復させたが,最終的 に直腸温度は低下した(図 7)。別の 1 匹のラットは,イソフルランの吸入の停止後も 自発的な行動を示すことなく,直腸温度は低下し続けた(図7)。一方,体温低下の過程 でイソフルランの吸入を停止することなく持続した場合は,直腸温度が 25℃を下回る と,3 例中全てのラットで心停止を起こした。 イソフルランの吸入を25℃で停止することにより,5 例中 2 例が低体温に誘導できた が,確実な方法ではなかった。また,イソフルランの吸入を続けた場合に心停止が生じ た理由として,低体温時の麻酔深度が深いことが原因であることが考えられた。そこで, より確実な低体温誘導法を確立するために,低体温誘導中に吸入しているイソフルラン 濃度を段階的に下げることで,確実な低体温誘導が可能か検討した。直腸温度が27.5℃ に低下するまではイソフルランの濃度を 2%で吸入を行い。27.5℃に到達した時点でイ ソフルランの濃度を1%に下げた。直腸温度が 22.5℃に低下した時点でイソフルランの 吸入を停止した。この方法では,イソフルランの吸入を停止した後も,ラットの直腸温 度と心拍数は低下し続け,目標の温度である 15℃に全てのラットが到達した(図 8a, b)。また低体温誘導中に異常な心電図は発生せず,直腸温度が 15℃に到達した時点で もP 波と QRS 群が連動する正常な洞調律を示していた(図 9)。心電図波形の解析では, 低体温誘導前のPR 間,QRS 群,QT 間の時間は,それぞれ 45.8 ± 4.2,18.8 ± 2.7,68.824 ± 9.1 ミリ秒であったが,15°C 到達時には 209.6 ± 15.8,56.7 ± 20.8,221.0 ± 85.2 ミリ秒 と有意に延長していた。なお,直腸温度が15℃に到達後も 4℃の低温環境下に置き続け ると,ラットの直腸温度は低下し続け,全てのラットが心停止を起こした。 この方法により誘導した低体温状態からラットを回復させることが可能かを検討す るために,直腸温度が15℃に到達したラットを室温 22℃の環境に移動させ,白熱灯に よりラットを暖めた。全てのラットで,直腸温度が 20℃以上に上昇してから自発的な 行動と震えが観察された。ラットは体温の回復途中に死亡することはなく,体温の回復 後にも異常な行動は見られなかった。また,心拍数は体温の回復とともに上昇し,体温 の回復途中や体温回復後に異常な心電図は生じなかった(図10)。 また,15℃の低体温状態の維持を行うために,冷却したステンレスプレートによる低 体温誘導法を試みた。濃度2%のイソフルラン麻酔下のラットを 10℃に冷却したステン レスプレートに静置した。冷蔵庫で確立した方法と同様に,直腸温度が27.5℃に到達す るまでは 2%の濃度で吸入を行い,直腸温度が 27.5℃から 22.5℃の間は 1%の濃度で麻 酔し,22.5℃以下の体温ではイソフルランの吸入を停止したところ,全てのラットが 15℃の低体温状態に誘導された。また,15℃到達後にステンレスプレートの温度を 14℃ に変更することで,15℃の極度の低体温状態を 6 時間まで維持することが可能であっ た。 イソフルラン麻酔と冷却によるラットの冬眠様低体温誘導中のCPT の経時的投与 第1 章において,中枢のアデノシン A1 受容体の活性化が低体温誘導に重要であるこ とを明らかとした。イソフルラン麻酔と冷却による低体温誘導においても,中枢のアデ ノシンA1 受容体の活性化が重要であるかを確認するために,イソフルランの低体温誘 導中にアデノシンA1 受容体のアンタゴニストである CPT を側脳室内に投与した。 イソフルランによる低体温誘導中にアデノシンA1 アンタゴニストを経時的に投与し たところ,5 例中全てのラットが,死亡することなく 15℃の低体温状態に到達した(図
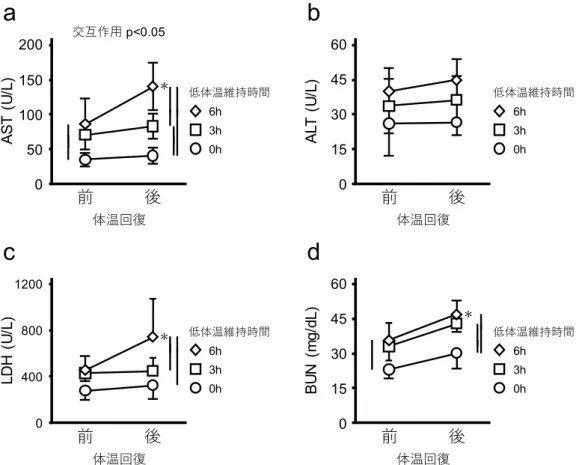
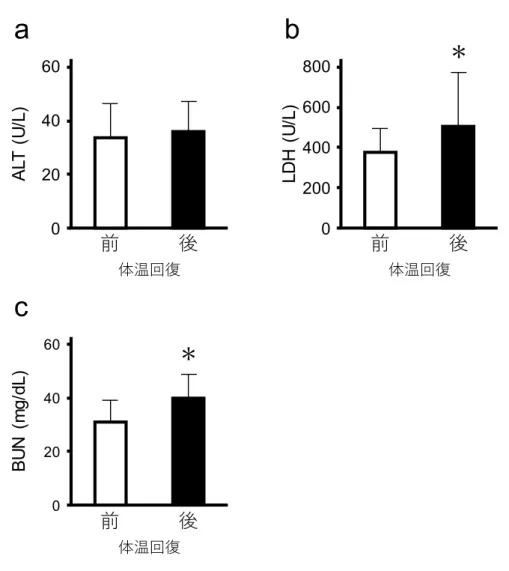
25 11)。15℃に到達するまでの時間に有意差はなかった(対照群 151.0±7.4 分 vs CPT 投与 群146.6±11.5 分 p>0.05)。低体温誘導中の心電図も異常は生じなかった(図 12)。15℃ 到達時のPR 間,QRS 群,QT 間の時間は,それぞれ 210.1 ± 16.8,44.2 ± 8.4,223.8 ± 72.1 ミリ秒であり,CPT を投与していない対照群と有意な差はなかった。 低体温の維持と回復がラットの血液生化学値に与える影響 低体温がラットに与える影響を評価するために,低体温誘導中の血液生化学値(AST, ALT,LDH,BUN)を測定した。体温が 15℃まで低下した時点では,血液生化学値に有 意な変化はなかった(図13)。しかし,低体温状態が 3 時間,6 時間と維持されること により,全ての項目で数値が有意に上昇した(図13)。LDH のみ,低体温を 6 時間維持 した後の値が3 時間維持した後の値に比べ有意に高かった(図 13c)。 全ての項目で,低体温の維持の有無に関わらず,低体温状態からの体温回復後の値は 体温回復前の値より有意に高かった。低体温の維持時間が,体温回復による血液生化学 値の上昇に影響を与えるかどうかを明らかにするため,体温回復前後の血液生化学値の データを二元配置分散分析により解析した。全ての項目で,低体温状態の維持時間と体 温の回復の間で有意な交互作用を示した(図14)。この結果は低体温状態の維持時間が, 体温回復による血液生化学値の上昇に影響を与えていること示唆しているが,どのよう に影響を与えているかは不明のままである。そのため,単純主効果について解析した。 全ての項目で,体温回復前の時点で,低体温維持時間による有意な差があった(図14)。 AST,LDH,BUN では,体温回復前の有意差が,体温回復後でも変化しなかった(図 14)。ALT では体温回復前の時点では,低体温を 3 時間維持した値と 6 時間維持した値 に有意な差はなかったが,体温回復後の時点では,6 時間維持した個体の値は 3 時間維 持した個体の値に比べて有意に高かった(図14b)。 低体温維持と体温回復が血液生化学値に相加的,もしくは相乗的に影響を与えている のかを明らかにするために,各時点の血液生化学値の平均値を用いて,体温回復の影響,
26 低体温維持による影響,低体温維持と体温回復による影響を計算した。3 時間維持した 場合と6 時間維持した場合の両方において,低体温の維持と回復による影響は,低体温 維持による影響と体温回復による影響の和より大きかった(図15)。 6 時間の低体温状態維持を経験した全てのラットは,1 週間後も生存していた。また, 1 週間後の血液生化学値は,低体温誘導前の値と有意差はなかった(図 16)。
27 図6 直腸温度 27.5℃の時点でイソフルランを停止した場合のラットの直腸温度の変化 このグラフはイソフルランと冷却による低体温誘導時に,ラットの直腸温度が27.5℃に 到達した時点でイソフルランの吸入を停止した場合の,各ラットの直腸温度の変化を示 している(n=5)。□などの各マーカーはそれぞれの個体を示す。横軸はラットを 4℃環 境下に移動させてからの時間を示している。全てのラットが震えや自発的な行動を示し, 低温環境下にも関わらず体温が回復した。白いバーはイソフルランを2%の濃度で吸入 している時間を示している。
150
90
60
0
30
120
180
210
30
20
10
直腸温度
(ºC
)
冷却時間
(分)
40
0
2% イソフルラン濃度28 図7 直腸温度 25℃の時点でイソフルランを停止した場合のラットの直腸温度の変化 このグラフはイソフルランと冷却による低体温誘導時に,ラットの直腸温度が 25℃に 到達した時点でイソフルランの吸入を停止した場合の各ラットの直腸温度の変化を示 している(n=5)。□などの各マーカーはそれぞれの個体を示す。横軸はラットを 4°C 環境下に移動させてからの時間を示している。5 匹中 3 匹のラットが震えを行い,直腸 温度を自発的に回復させた。2 匹のラットは吸入停止直後に,震えと自発的な行動を行 ったが,直腸温度が 22℃を下回ると不動化した。これらのラットの直腸温度は最終的 に15℃まで到達した。白いバーはイソフルランを 2%の濃度で吸入している時間を示し ている。
300
180
120
0
60
240
360
30
20
10
直腸温度
(ºC
)
冷却時間
(分)
40
0
2% イソフルラン濃度29 図8 イソフルラン濃度を段階的に軽減した場合のラットの直腸温度と心拍数の変化 (a)上のグラフはイソフルラン濃度を段階的に制御し,直腸温度が 22.5℃に到達した 時点でイソフルランの吸入を停止した場合の,各ラット(n=6)の直腸温度の変化を示 している 。直腸温度に従い,イソフルランの濃度を 2%から 0%に段階的に減らした。 横軸はラットを4℃環境下に移動させてからの時間を示している。ラットの直腸温度は イソフルランの吸入停止後も時間経過とともに低下した。(b)グラフは,低体温誘導 中のラットの直腸温度と心拍数の相関図を示している。横軸はラットの直腸温度を示し ている。ラットの直腸温度が低下するにつれて,ラットの心拍数は低下した。 34 30 26 22 18 300 200 38 14 100 400
直腸温度(ºC)
0 2% 1% イソフルラン濃度b
心拍数
(b
p
m
)
150 90 60 0 30 120 180 30 20 40 10 0a
直腸温度 (ºC ) 冷却時間(分) 2% 1% イソフルラン濃度30 図9 イソフルランによる低体温誘導中のラットの心電図 イソフルランによる低体温誘導中のラットの心電図波形の典型例を示した。低体温誘導 前(a),直腸温度 25℃(b),15℃(c)の心電図を示した。P 波と QRS 群を矢頭と矢印 でそれぞれ示した。直腸温度15℃の状態でも,正常な洞調律は維持されていた。同様の 結果が6 例中全てのラットで再現性良く観察された。
a
低体温誘導前
b
直腸温度25ºC
c
直腸温度
15ºC
1 秒
5
V
SHI-505 心電図 (V ) -15 -10 -5 0 5 10 15 15:58:30.1 15:58:30.2 15:58:30.3 15:58:30.4 15:58:30.5 15:58:30.6 15:58:30.7 15:58:30.8 15:58:30.9 15:58:31 15:58:31.1 15:58:31.2 15:58:31.3 15:58:31.4 15:58:31.5 15:58:31.6 15:58:31.7 15:58:31.8 15:58:31.9 15:58:32 15:58:32.1 15:58:32.2 15:58:32.3 15:58:32.4 15:58:32.5 15:58:32.6 15:58:32.7 15:58:32.8 15:58:32.9 15:58:33 15:58:33.1 15:58:33.2 15:58:33.3 15:58:33.4 15:58:33.5 15:58:33.6 15:58:33.7 15:58:33.8 15:58:33.9 15:58:34 2018/01/11 15:58:30.000 SHI-505 心電図 (V ) -15 -10 -5 0 5 10 15 16:52:30.1 16:52:30.2 16:52:30.3 16:52:30.4 16:52:30.5 16:52:30.6 16:52:30.7 16:52:30.8 16:52:30.9 16:52:31 16:52:31.1 16:52:31.2 16:52:31.3 16:52:31.4 16:52:31.5 16:52:31.6 16:52:31.7 16:52:31.8 16:52:31.9 16:52:32 16:52:32.1 16:52:32.2 16:52:32.3 16:52:32.4 16:52:32.5 16:52:32.6 16:52:32.7 16:52:32.8 16:52:32.9 16:52:33 16:52:33.1 16:52:33.2 16:52:33.3 16:52:33.4 16:52:33.5 16:52:33.6 16:52:33.7 16:52:33.8 16:52:33.9 16:52:34 2018/01/11 16:52:30.000 SHI-505 心電図 (V ) -15 -10 -5 0 5 10 15 18:35:05.3 18:35:05.4 18:35:05.5 18:35:05.6 18:35:05.7 18:35:05.8 18:35:05.9 18:35:06 18:35:06.1 18:35:06.2 18:35:06.3 18:35:06.4 18:35:06.5 18:35:06.6 18:35:06.7 18:35:06.8 18:35:06.9 18:35:07 18:35:07.1 18:35:07.2 18:35:07.3 18:35:07.4 18:35:07.5 18:35:07.6 18:35:07.7 18:35:07.8 18:35:07.9 18:35:08 18:35:08.1 18:35:08.2 18:35:08.3 18:35:08.4 18:35:08.5 18:35:08.6 18:35:08.7 18:35:08.8 18:35:08.9 18:35:09 18:35:09.1 18:35:09.2 2018/01/11 18:35:05.20031 図10 体温回復時のラットの心電図 イソフルランによる低体温誘導後に体温を回復させたラットの心電図波形の典型例を 示した。15℃到達直後(a),直腸温度 20℃(b),体温回復後(c)の心電図を示した。 体温回復後の心電図は2%のイソフルラン麻酔下で計測した。P 波と QRS 群を矢頭と矢 印でそれぞれ示した。 体温の回復途中,体温回復後のいずれの時点でも正常な洞調律 が維持されていた。同様の結果が6 例中全てのラットで再現性よく観察された。 SHI-639 Ch an ne l 7 (V ) -4 -2 0 2 4 2:16:05.22:16:05.32:16:05.42:16:05.52:16:05.62:16:05.72:16:05.82:16:05.92:16:062:16:06.12:16:06.22:16:06.32:16:06.42:16:06.52:16:06.62:16:06.72:16:06.82:16:06.92:16:072:16:07.12:16:07.22:16:07.32:16:07.42:16:07.52:16:07.62:16:07.72:16:07.82:16:07.92:16:082:16:08.12:16:08.22:16:08.32:16:08.42:16:08.52:16:08.62:16:08.72:16:08.82:16:08.92:16:092:16:09.1 2:16:05.140 SHI-639 Ch an ne l 7 (V ) -4 -2 0 2 4 3:40:48.13:40:48.23:40:48.33:40:48.43:40:48.53:40:48.63:40:48.73:40:48.83:40:48.93:40:493:40:49.13:40:49.23:40:49.33:40:49.43:40:49.53:40:49.63:40:49.73:40:49.83:40:49.93:40:503:40:50.13:40:50.23:40:50.33:40:50.43:40:50.53:40:50.63:40:50.73:40:50.83:40:50.93:40:513:40:51.13:40:51.23:40:51.33:40:51.43:40:51.53:40:51.63:40:51.73:40:51.83:40:51.93:40:523:40:52.1 3:40:48.100 SHI-639 Ch an ne l 7 (V ) -4 -2 0 2 4 2:28:54.92:28:552:28:55.12:28:55.22:28:55.32:28:55.42:28:55.52:28:55.62:28:55.72:28:55.82:28:55.92:28:562:28:56.12:28:56.22:28:56.32:28:56.42:28:56.52:28:56.62:28:56.72:28:56.82:28:56.92:28:572:28:57.12:28:57.22:28:57.32:28:57.42:28:57.52:28:57.62:28:57.72:28:57.82:28:57.92:28:582:28:58.12:28:58.22:28:58.32:28:58.42:28:58.52:28:58.62:28:58.72:28:58.82:28:58.9 2:28:54.900
a
直腸温度15℃b
直腸温度20ºCc
直腸温度36℃(イソフルラン2%麻酔下) 1 秒 5 V32 図11 イソフルランによる低体温誘導中に CPT を経時的に投与した場合のラットの体 温と心拍数の変化 これらのグラフはイソフルランによる低体温誘導中に CPT を経時的に投与した場合の ラットの体温(a)と心拍数(b)の変化の典型例を示す。矢印は CPT(60 nmol/5μL)を 投与した時点を示す。横軸は環境温度(Ta)が ۦ22℃の室内から 4℃の冷蔵庫に移動して からの時間を示す。イソフルラン麻酔下でCPT を側脳室内投与した 5 分後に 4℃の環 境に移した。その後,直腸温度が30℃,25℃,20℃,15℃に到達した時点でも CPT を 投与した。ラットの直腸温度と心拍数は時間経過とともに低下し,最終的にラットの直 腸温度は15℃に到達した。同様の結果が 5 匹のラットから再現性良く得られた。 Ta 22ºC:4ºC Ta 22ºC:4ºC 300 200 100 400 0
b
心拍数 (b p m ) 30 20 40 10 0a
直腸温度 (ºC ) 150 90 60 120 30 0 180 冷却時間(分) 150 90 60 0 30 120 180 冷却時間(分) 2% 1% イソフルラン濃度 2% 1% イソフルラン濃度33 図12 イソフルランによる低体温誘導中に CPT を経時的に投与した場合のラットの心 電図 イソフルランによる低体温誘導中に CPT を経時的に投与した場合のラットの各時点に おける心電図を示した。(a)CPT を投与する直前。(b)CPT を投与してから 5 分後(4℃ 環境に移す直前)。(c)直腸温度が 25℃に到達した時点。(d)直腸温度が 15℃に到達し た時点。P 波と QRS 群を矢頭と矢印でそれぞれ示した。CPT の投与は低体温誘導中の 心電図に変化を起こさなかった。同様の結果が5 匹のラットから再現性良く得られた。 SHI-525 心電図 (V ) -15 -10 -5 0 5 10 15 13:58:03.1 13:58:03.2 13:58:03.3 13:58:03.4 13:58:03.5 13:58:03.6 13:58:03.7 13:58:03.8 13:58:03.9 13:58:04 13:58:04.1 13:58:04.2 13:58:04.3 13:58:04.4 13:58:04.5 13:58:04.6 13:58:04.7 13:58:04.8 13:58:04.9 13:58:05 13:58:05.1 13:58:05.2 13:58:05.3 13:58:05.4 13:58:05.5 13:58:05.6 13:58:05.7 13:58:05.8 13:58:05.9 13:58:06 13:58:06.1 13:58:06.2 13:58:06.3 13:58:06.4 13:58:06.5 13:58:06.6 13:58:06.7 13:58:06.8 13:58:06.9 13:58:07 2018/02/06 13:58:03.000
a CPT投与直前
b CPT投与から5分後
c 直腸温度25ºC
d 直腸温度15ºC
SHI-524 心電図 (V ) -15 -10 -5 0 5 10 15 14:34:38 14:34:38.1 14:34:38.2 14:34:38.3 14:34:38.4 14:34:38.5 14:34:38.6 14:34:38.7 14:34:38.8 14:34:38.9 14:34:39 14:34:39.1 14:34:39.2 14:34:39.3 14:34:39.4 14:34:39.5 14:34:39.6 14:34:39.7 14:34:39.8 14:34:39.9 14:34:40 14:34:40.1 14:34:40.2 14:34:40.3 14:34:40.4 14:34:40.5 14:34:40.6 14:34:40.7 14:34:40.8 14:34:40.9 14:34:41 14:34:41.1 14:34:41.2 14:34:41.3 14:34:41.4 14:34:41.5 14:34:41.6 14:34:41.7 14:34:41.8 14:34:41.9 2018/02/05 14:34:38.000 SHI-525 心電図 (V ) -15 -10 -5 0 5 10 15 11:38:15.1 11:38:15.2 11:38:15.3 11:38:15.4 11:38:15.5 11:38:15.6 11:38:15.7 11:38:15.8 11:38:15.9 11:38:16 11:38:16.1 11:38:16.2 11:38:16.3 11:38:16.4 11:38:16.5 11:38:16.6 11:38:16.7 11:38:16.8 11:38:16.9 11:38:17 11:38:17.1 11:38:17.2 11:38:17.3 11:38:17.4 11:38:17.5 11:38:17.6 11:38:17.7 11:38:17.8 11:38:17.9 11:38:18 11:38:18.1 11:38:18.2 11:38:18.3 11:38:18.4 11:38:18.5 11:38:18.6 11:38:18.7 11:38:18.8 11:38:18.9 11:38:19 2018/02/06 11:38:15.000 SHI-525 心電図 (V ) -15 -10 -5 0 5 10 15 12:28:08.1 12:28:08.2 12:28:08.3 12:28:08.4 12:28:08.5 12:28:08.6 12:28:08.7 12:28:08.8 12:28:08.9 12:28:09 12:28:09.1 12:28:09.2 12:28:09.3 12:28:09.4 12:28:09.5 12:28:09.6 12:28:09.7 12:28:09.8 12:28:09.9 12:28:10 12:28:10.1 12:28:10.2 12:28:10.3 12:28:10.4 12:28:10.5 12:28:10.6 12:28:10.7 12:28:10.8 12:28:10.9 12:28:11 12:28:11.1 12:28:11.2 12:28:11.3 12:28:11.4 12:28:11.5 12:28:11.6 12:28:11.7 12:28:11.8 12:28:11.9 12:28:12 2018/02/06 12:28:08.000 1 秒 5 V34 図13 低体温の導入と維持がラットの血液生化学値に与える影響 これらのグラフは AST(a),ALT(b),LDH(c),BUN(d)の平均値±標準偏差を示 している。横軸は採血の各時点を示している。対照群は低体温誘導を行っていない通常 体温のラットの血液生化学値の平均値を示している(n=5)。0h は低体温誘導により直 腸温度が15℃に到達した直後のラットの血液生化学値の平均値を示す(n=6)。3h と 6h はそれぞれ15℃の低体温状態を 3 時間または 6 時間維持したラットの血液生化学値の 平均値を示す(3 時間: n=7,6 時間: n=6)。全ての項目の値は,低体温を維持する時間が 延びることで有意に上昇した。横棒は各群間に有意差があることを示す(p<0.05)。
a
A S T (U /L ) 対照群 300 200 100 0 0h 3h 6h 低体温維持時間b
A LT ( U /L ) 対照群 100 75 50 25 0 0h 3h 6hc
LD H ( U /L ) 対照群 800 400 0h 3h 1200 0 1600d
B U N ( m g/ dL ) 対照群 25 20 15 10 5 0h 3h 0 低体温維持時間 6h 低体温維持時間 6h 低体温維持時間35 図14 低体温の維持と回復がラットの血液生化学値に与える影響 これらのグラフは,体温回復前後の各群のAST(a),ALT(b),LDH(c),BUN(d) の平均値±標準偏差を示している。横軸は採血時点を示している。○は低体温誘導によ り直腸温度が 15℃に到達した直後に体温を回復させたラットの血液生化学値の平均値 を示している(n=6)。□と◇はそれぞれ,15℃の低体温状態を 3 時間,6 時間維持した 直後に体温を回復させたラットの平均値を示している(3 時間: n=7,6 時間: n=6)。二元 配置分散分析により低体温状態の維持時間と体温の回復には有意な交互作用があるこ とが確認された。縦棒は各郡間に有意差(p<0.05)があることを示している。*は各低 体温維持群において体温回復の直前と直後において有意差(p<0.05)があることを示す。
b
A LT ( U /L ) 160 0 40 80 前 後 120a
A S T (U /L ) 400 0 100 200 300 前 後c
LD H ( U /L ) 3000 0 1000 2000d
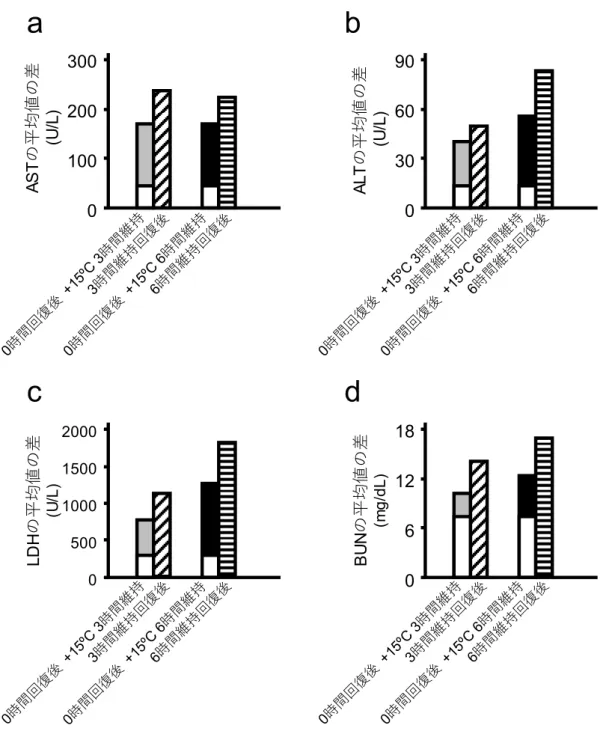
40 0 10 20 30 B U N ( m g/ dL ) 体温回復 交互作用p<0.05 体温回復 交互作用p<0.05 低体温維持時間 6h 3h 0h 低体温維持時間 6h 3h 0h 交互作用p<0.05 交互作用p<0.05 前 後 前 後 体温回復 体温回復 低体温維持時間 6h 3h 0h 低体温維持時間 6h 3h 0h * * * * * * * * * * * *36 図15 体温の回復による血液生化学値の上昇に低体温の維持が与える影響 これらのグラフは,AST(a),ALT(b),LDH(c),BUN(d)の各採血時点間での血液 生化学値の平均値の差を表わしている。“0 時間回復後”(白)は15℃到達直後に体温を 回復させた時点の血液生化学値の平均値から,15℃到達時点の血液生化学値の平均値を
a
b
300 0 100 A ST の平均値の差 (U /L ) 90 0 30 60 A LT の平均値の差 (U /L )c
d
1000 1500 LD H の平均値の差 (U /L ) 500 0 2000 18 0 6 12 200 BU N の平均値の差 (m g /d L )37 引くことにより算出した。“15℃3 時間維持”(グレー)は低体温状態を 3 時間維持した 時点の血液生化学値の平均値から,15℃到達時点の血液生化学値の平均値を引くことに より算出した。“15℃6 時間維持”(黒)は低体温状態を 6 時間維持した時点の血液生化 学値の平均値から,15℃到達時点の血液生化学値の平均値を引くことにより算出した。 “3 時間維持回復後”(斜線)は低体温状態を 3 時間維持した後に体温を回復させた時 点の血液生化学値の平均値から,15℃到達時点の血液生化学値の平均値を引くことによ り算出した。“6 時間維持回復後”(横線)は低体温状態を 6 時間維持した後に体温を回 復させた時点の血液生化学値の平均値から,15℃到達時点の血液生化学値の平均値を引 くことにより算出した。
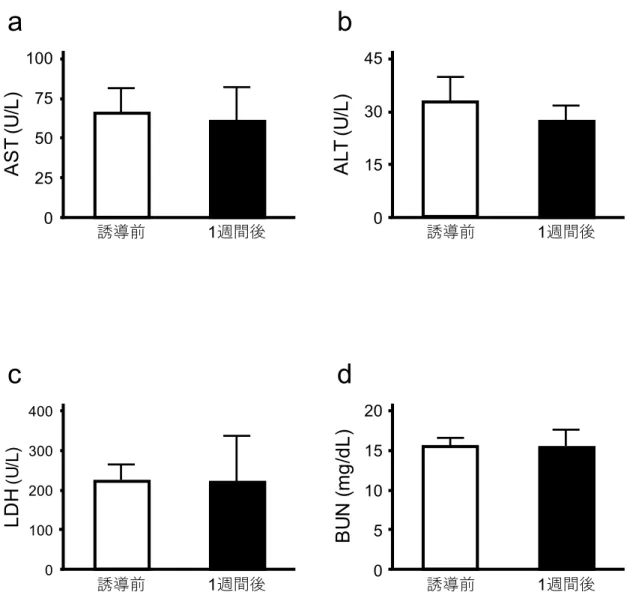
38 図16 低体温を 6 時間維持したラットの 1 週間後の血液生化学値 これらのグラフは低体温状態を 6 時間維持したラットの,低体温誘導前と 1 週間後の AST(a),ALT(b),LDH(c),BUN(d)の平均値±標準偏差を表わしている(n=6)。 全ての項目において有意な差はなかった。
a
100 0 25 75A
S
T
(U
/L
)
b
45 0 15 30A
LT
(U
/L
)
c
200 300L
D
H
(U /L ) 100 0 400d
20 0 5 15B
U
N
(
m
g
/d
L
)
50 10 誘導前 1週間後 1週間後 誘導前 誘導前 1週間後 1週間後 誘導前39
考察
第1 章の結果から,低体温時における自律神経の適切な制御が極度の低体温状態で心 拍動を維持するために重要であることが示唆された。しかしながら,第1 章で確立した 冬眠様低体温誘導法は事前の脳外科手術を必要とするため,脳の傷害に対する低体温療 法をする場合に不都合となり得る可能性がある。そこで,事前の外科手術を必要としな い低体温誘導方法として,吸入麻酔を用いた新たな低体温誘導法の確立を試みた。イソ フルランによる吸入麻酔と冷却を組み合わせることにより,ラットに心停止を起こすこ となく15°C の極度の低体温状態に誘導する方法を確立することができた。また,重度 な傷害を引き起こすことなく非冬眠動物の極度の低体温状態を 6 時間まで維持するこ とが可能であることが明らかとなった。低体温療法は,心筋梗塞(72),脊髄損傷(33), 脳外傷(13)の治療に用いられている。また,低体温療法の治療効果は,低体温に誘導 するまでの時間が短いほど効果が高いことが報告されている(74)。本章で確立したイ ソフルランの吸入と冷却を組み合わせる低体温誘導法は,脳への薬剤投与を必要とする ことなく素早く低体温に誘導できることから,極度の低体温を医療に応用する上で非常 に有益だろう。 イソフルランの吸入濃度を適切に調節することにより,ラットを冬眠様低体温に誘導 することに成功した。麻酔により体温が低下するのは,脳の体温調節中枢の機能が麻酔 により抑制されるためであると考えられる。実際,イソフルランの吸入をラットの体温 が27.5℃の時点で停止すると,ラットは震えを起こし,低温環境下で体温を回復させた。 これは,イソフルランの吸入が行われている間は体温調節中枢が抑制され,吸入が停止 されることにより,体温調節中枢の機能が回復し体温の上昇が行われることを示してい る。興味深いことに,ラットの体温が22.5℃の時点でイソフルランの吸入を停止した場 合は,吸入を停止したにも関わらず全てのラットが震えを起こすことなく体温は低下し 続けた。イソフルランの吸入を 25℃で停止した場合は,いくつかのラットは体温を回40 復させることがなかったことに加え,27.5℃で吸入を停止した場合には,全てのラット が体温を回復させたことから,体温調節中枢には体温回復行動を起こさせる下限の温度 というものが存在し,その温度が27.5℃から 22.5℃の間にあると予想される。もしこの 予想が正しいとすると,冬眠様低体温誘導を行うためにはこの下限の温度を超える体温 に誘導することが重要となる。つまり,他の手法を用いても,イソフルラン麻酔を用い る方法と同様な低体温誘導が可能であると考えられる。例えば,マウスにおいては TRPV1 チャネルの活性化が熱放散の促進と震えの抑制により体温を低下させると報告 されている(20)。このような手法を用いても,体温中枢の下限の温度を超えることさ えできれば,冬眠様低体温誘導が可能であると考えられる。 本章ではイソフルランによる吸入麻酔により低体温誘導を行ったが,体温低下の機序 はまだ明らかになっていない。前章では中枢のアデノシンA1 受容体を活性化すること により低体温誘導を行った。側脳室内に投与されたCHA は,孤束核のアデノシン A1 受 容体に作用し,熱産生を抑制することが報告されている(78)。しかし,中枢による体 温制御には,孤束核の他にも視索前野や視床下部背内側核,吻側縫線核など様々な部位 が関与し,これらの部位が神経回路を形成し体温を制御している(61)。イソフルラン による低体温誘導中にアデノシン A1 受容体のアンタゴニストである CPT を経時的に 投与しても,ラットの体温低下に影響を与えなかったことから,アデノシンA1 受容体 はイソフルランによる体温低下に関与していないと考えられる。イソフルランは抑制性 伝達物質の受容体であるGABAA受容体やグリシン受容体などを活性化する(2)ことか ら,イソフルランの吸入中の体温低下は,アデノシンA1 受容体を介さずに孤束核,あ るいは視索前野などの他の体温制御に関する脳部位を抑制したことによると考えられ る。 イソフルランによる低体温誘導では,15℃までは安定的に低体温誘導を行うことがで きたが,体温が 15℃より低下すると心停止が生じていた。吸入麻酔を停止した後に生 じていることから,低体温そのものがラットの心臓の洞調律に影響を与えることが考え
41 られる。実際に心電図波形の解析を行うと,15℃到達時では正常な洞調律を維持してい たものの,通常の体温と比べてPR 間,QRS 群,QT 間の時間の長さの有意な延長がみ られた。このような変化は不整脈発生の前兆とされている(12,32)。不整脈を起こす 要因には様々なものがあるが,その一つとして心筋細胞内のカルシウム濃度の異常な上 昇がある(62)。ラットの心筋細胞は低温に置かれると,心筋筋小胞体の Ca2+-ATPase ポ ンプの機能低下により筋小胞体へのカルシウムイオンの取り込みが低下すること(44) や,心筋細胞内のカルシウム濃度が上昇するという報告(81)もあることから,低体温 誘導により生じたPR 間,QRS 群,QT 間の延長や 15℃以降の低体温時に生じる心停止 は,低温により心筋細胞内のカルシウム濃度が上昇したことによるものであると考えら れる。 低体温の臨床応用を考えると,低体温がもたらす副作用について明らかにすることは 非常に重要である。血液生化学値における評価項目である AST,ALT の増加は主に肝 臓の,LDH は骨格筋や心臓の組織傷害を反映している(65,69,80)。また,BUN の増 加は腎機能の障害を反映するとされている(38)。本研究では低体温誘導中のラットか ら経時的に採血を行い,これら4 つの項目を測定することで低体温誘導がラットに与え る影響について評価した。興味深いことに,低体温誘導により体温が 15℃に到達した 直後では,全ての項目が低体温誘導前の通常体温と有意な差がなかった。このことは, たとえ非冬眠動物であっても,極度の低体温が与える影響は小さいことを示唆している。 しかしながら,ラットの体温が15℃に到達するまでの平均時間は 151.0 ± 7.4 分であり, 極度の低体温状態であった時間が短かったために,血液生化学値に反映されるような傷 害が生じていなかった可能性もある。実際に15℃の低体温状態を 3 時間または 6 時間 維持した場合には,全ての項目で有意な上昇が見られたことから,長時間の低体温状態 の維持はラットに非特異的な組織傷害と腎機能障害をもたらすと言える。特にLDH に おいては,低体温状態を6 時間維持した直後の値が 3 時間維持した値に比べて有意に高 かった。LDH が主に骨格筋や心臓の傷害を反映することを考慮すると,これらの臓器
42 が特に低体温の影響を受けやすいと言える。また,時間経過に伴い有意な増加を示した ことから,この影響はより長時間の維持をした場合には更に大きくなることが予想され る。つまり,非冬眠動物には低体温状態を維持することができる時間に上限があること が推測できる。冬眠動物であるシリアンハムスターは極度の低体温状態を24 時間以上 維持することが可能(59,76)であることから,冬眠動物と非冬眠動物の最も大きな違 いは,低体温状態を安全に維持する能力にあると考えられる。冬眠動物は冬眠期間中に 高い低体温耐性を示すことが知られており(19,35,49),この冬眠期間中には,RNA の転写やタンパクの発現が時期特異的に制御されている(16,66,83)。もし,冬眠時 に起きる制御を非冬眠動物においても再現することができれば,より長時間の低体温状 態を傷害なく維持することが可能となるだろう。 また,低体温療法では低体温に誘導するのと同様に,低体温からの回復が重要である。 そこで,低体温の維持時間の違いが,低体温からの体温回復にどのような影響を与える かについても評価した。二元配置分散分析により,全ての血液生化学値の項目において 低体温状態の維持時間と体温の回復の間に有意な交互作用があったことから,低体温の 維持時間の長さが低体温からの回復後の血液生化学値に影響を与えていることが明ら かとなった。低体温状態の維持時間に関わらず,体温の回復後の値が回復前の値より有 意に高かったことから,低体温状態の維持は体温回復時に生じる血液生化学値の上昇の 程度に影響を与えていることが予想される。この影響は相加的,相乗的に作用し,血液 生化学値の上昇をより大きくする,あるいは拮抗的に作用して血液生化学値の上昇を抑 えている可能性が挙げられる。しかし,体温回復前に見られた各低体温維持群間の有意 な差が,体温回復によって消失あるいは逆転することはなかったことから,拮抗的に作 用してはないと考えられる。相加的あるいは相乗的に作用するかをより詳細に検討する ために,各条件での血液生化学値の平均値の差を求めることで,低体温維持と体温回復 が血液生化学値に与える影響を評価した。図15 に示すように,3 時間維持と 6 時間維 持の両方の場合において,低体温維持とその後の体温回復による影響が,低体温維持単
43 独の影響と体温回復の単独の影響の和より大きかった。各条件間の平均値の差を求めた ため,統計学的解析を行うことはできなかったが,もし低体温の維持時間と体温の回復 が相加的に影響を与えているのであれば,これらの値は等しくなると考えられるため, 低体温維持の影響と体温回復の影響は,相加的というよりは相乗的に影響を与えている と考えられる。このことは,低体温の維持時間が長時間になるほど,体温回復時に生じ る影響が大きくなることを示唆しているため,極度の低体温を医療に応用するためには, この体温回復時に生じる変化をできるだけ抑える必要がある。冬眠動物では体温の回復 時にも様々な遺伝子の発現が変化する(66,83)ことから,このような変化を再現する ことで軽減することができると考えられる。 血液生化学値の変化より,ラットは長時間の低体温状態の維持あるいは低体温状態か らの回復により組織傷害や腎機能障害が生じることが明らかとなった。このような変化 は冬眠動物にはみられないことから,より安全な低体温誘導法を確立するためには改良 するべき点ではある。しかし,6 時間の低体温状態を維持したラットの血液生化学値が 1 週間後には正常な値に回復していたことから,本研究で明らかとなった体温誘導法に よる影響は一過性のものであり,重大な後遺症をもたらすものではないと考えられる。 このことは,本章で確立したイソフルランによる低体温誘導法を医療応用する点で非常 に重要な点となる。 本章では,外科手術を必要としないイソフルランによる麻酔と冷却を組み合わせた冬 眠様低体温誘導法を確立した。本章で得られた主な結果は1)イソフルランの吸入麻酔 をラットの直腸温度が22.5℃の時点で停止することにより,ラットに心停止を起こさせ ることなく直腸温度 15℃の状態に誘導することが可能であることを明らかにした。2) この低体温状態を6 時間まで維持することができた。3)低体温誘導したラットの体温 は,加温することにより回復した。4)AST,ALT,LDH,BUN は低体温状態の維持と 低体温からの体温回復により上昇した。5)上昇した値は 1 週間以内に回復した。これ らの結果は,確立したイソフルランによる冬眠様低体温誘導法により,非冬眠動物であ
44
るラットに重大な後遺症をもたらすことなく低体温状態に誘導し,少なくとも6 時間ま