レット症候群疾患特異的ヒト
iPS 細胞の樹立と
神経系細胞分化におけるエピジェネティクス
山梨大学大学院
医学工学総合教育部
論文博士学位論文
2017 年 3 月
野田 友子
目次
ページ数 序論 1 略語一覧 3 レット症候群の概要および研究の展望 第1節 レット症候群の臨床像 4 第2 節 MECP2 の機能と役割 4 第3 節 遺伝子変異と病態の関連性 6 第4 節 X chromosome inactivation とレット症候群について 7 第5 節 神経学的解析におけるレット症候群の疾患モデル 7 第1 章 レット症候群患者由来疾患特異的 iPS 細胞の樹立 第1 節 序 10 第2 節 実験材料および方法 1) 研究倫理申請 11 2) レット症候群患者の選定 11 3) 皮膚繊維芽細胞(Fibroblasts)の樹立 11 4) フィーダー細胞の調整 11 5) iPS 細胞の樹立と培養方法 11 6) X 染色体の不活化パターンの判定 12 7) 蛍光免疫染色 13 8) テラトーマ形成 14 9) 核型解析 14 第3 節 結果 1) fibroblasts の樹立とモザイク現象の確認 15 2) iPS 細胞の樹立 17 3) iPS 細胞の X 染色体の不活化パターンの判定 18 第4 節 考察 22 第2 章 X 染色体不活化解析を用いた iPS 細胞の性質安定性評価の試み 第1 節 序 23 第2 節 実験材料および方法 1) 使用した iPS 細胞について 242) 細胞の培養方法 24 3) 実験の手順 24 4) X 染色体の不活化パターンの判定 24 第3 節 結果 25 第4 節 考察 29 第3 章 レット症候群由来疾患特異的 iPS 細胞を用いた 神経系分化におけるエピジェネティック特性の解析 第1 節 序 30 第2 節 実験材料および方法 1) 使用した iPS 細胞について 31 2) 細胞の培養方法 31 3) 神経幹細胞(neurosphere)への分化誘導方法 31 4) 神経系細胞への分化誘導方法 31 5) 蛍光免疫染色 32 6) マイクロアレイ解析 32 7) 定量 RT-PCR 解析 33
8) クロマチン免疫沈降(chromatin immunoprecipitation; ChIP)
およびChIP-PCR 34 9) Bisulfite sequencing 35 第3 節 結果 1) iPS 細胞における MeCP2 発現パターンの確認 36 2) 神経分化能の確認 37 3) iPS 細胞および神経系細胞を用いた網羅的遺伝子発現解析 38 4) 変異 MECP2 発現株ではアストロサイトへの分化が促進される 41 5) MeCP2 の欠損は神経系細胞分化において GFAP への結合を失う 46 第4 節 考察 50 総括 51 参考文献リスト 55 関連論文 68 謝辞 70
- 1 -
序論
レット症候群は1966 年にウィーンの小児科医である Andreas Rett によって最 初に報告された小児期に発症する進行性の神経系疾患である[1]。その後、1983 年 にスウェーデンの Bengt Hagberg らによる 35 例の臨床像をまとめた報告がなさ れ、レット症候群が世界的に知られることとなった[2]。本疾患が広く研究される ようになった理由は、この疾患の原因遺伝子が主に X 染色体長腕末端に存在する methyl-CpG binding protein 2 (MECP2)遺伝子であることが明らかになったこと にある[3, 4]。原因遺伝子の特定ができたことで、培養細胞を用いて生体における MECP2の役割を解明するだけでなく、遺伝子ノックアウトマウスなどのモデル動 物 の 作 出 が 可 能 と な っ た[5, 6] 。 そ の 後 の 研 究 に よ り 、 リ ン 酸 化 酵 素 cyclin-dependent kinase-like 5 (CDKL5)や 転写因 子で ある forkhead box G1 (FOXG1)での遺伝子異常も、レット症候群を引き起こす遺伝子変異であることが 明らかとなった[7, 8]。しかしながら、患者の大多数はMECP2に変異を有してい る。この疾患は X 染色体連鎖優性遺伝形式に則り主に女児に発症し、その頻度は 10000 人から 15000 人に 1 人である[9]。女性には一般的に性染色体である X 染色 体が 2 本存在し、そのどちらか一方の染色体はランダムな不活化を受けることで 遺伝子発現が抑制さ れ る。この女性特有の 現 象 は X chromosome inactivation (XCI; X 染色体の不活化)と呼ばれ、ランダムに生じる不活化により女性の体細胞 は父由来X 染色体が活性化している細胞と母由来 X 染色体が活性化している細胞 の2種類がおよそ半数ずつ存在している[10-13]。MECP2はX 染色体上に存在し、 レット症候群の患者の大多数は他方の X 染色体に正常 MECP2を有する女性であ る。したがって、患者の体細胞では正常 MECP2 を有する X 染色体が不活化され ると変異 MeCP2 が産生され、変異 MECP2 を有する X 染色体が不活化を受ける と正常 MeCP2 が産生されるため、患者の体細胞は正常 MECP2 が発現する細胞 と変異 MECP2が発現する細胞の 2 種類が混在した状態である。一方で、男性の 場合はすべての細胞で変異MeCP2 が産生されるため、胎生致死または乳幼児期に 重度な脳障害が生じ死亡する場合が多数である[14-16]。 レット症候群を含む脳神経系の疾患におけるこれまでの研究手法はモデルマウ スを用いたものや、ヒトの受精卵から樹立された様々な体細胞へと分化すること ができるヒト胚性多能性幹細胞(Embryonic stem cells; ES 細胞)を利用した研究 が主であった[5, 6, 17]。特にヒト ES 細胞は高い自己増殖能と多分化能を有し、- 2 - 発生学的な研究から分化細胞の機能解析まで様々な分野での研究応用が進められ ており、同時に細胞移植治療や創薬開発のための細胞供給源として期待されてい た。しかしながら、ヒト ES 細胞は受精卵を破壊するという「命の起源はどこから 始まるのか」という問題と、実際に治療を行う際に避けては通れないであろう免 疫拒絶反応という問題が存在した。この問題点を突破したのが、2006 年にマウス で、そして 2007 年にヒトから作られた人工多能性幹細胞(induced pluripotent stem cells; iPS 細胞)である[18-20]。この細胞は、レトロウイルスを用いて体細胞 に山中4 因子と呼ばれる初期化因子(OCT4, KLF4, SOX2, and c-MYC)を導入する ことで、ヒトES 細胞と同等の特性、すなわち自己増殖能と多分化能を有する細胞 を得ることができた。この手法は神経系疾患においてもすぐさま有用性が示され、 これまで生検が困難であった患者由来の生きた神経細胞を用いた研究が活発に行 われるようになった[21-24]。本研究ではこの iPS 細胞の技術を用いて、iPS 細胞 の培養過程に引き起こされる X 染色体不活化の変遷と、神経系細胞分化における 遺伝子発現量調節機構といったエピジェネティック特性の観点から 研究を行った。
- 3 -
略語一覧
III-tubulin, neuronal class III -tubulin (チューブリンクラス III); ES cells, embryonic stem cells(胚性幹細胞);
GFAP, Glial Fibrillary Acidic Protein(グリア線維性酸性タンパク質); iPS cells, induced pluripotent stem cells(人工多能性幹細胞);
MeCP2 or MECP2, methyl-CpG binding protein 2(メチル化 CpG 結合タンパク 質);
- 4 -
レット症候群の概要および研究の展望
前述のとおり、レット症候群は主に MECP2 の変異によりもたらされる疾患で あることが判明している。そこで本節ではこれまでに明らかとなっているレット 症候群の臨床像、MECP2の役割および神経学的な解析結果について論じる。 第1節 レット症候群の臨床像 臨床経過としては、大きく4つのステージに大別される。生後6か月から18 か月までは一見すると普通の成長過程をたどるが(ステージ1)、徐々にてんかん 症状や特徴的な手もみ運動の繰り返し、それまでに獲得していた言語の喪失、 自 閉症状が観察されるようになり(ステージ 2)、急激な退行の後に多くの場合は一 部症状の改善や症状の安定化がみられる(ステージ 3)が、その後運動機能障害の 悪化がみられるようになる(ステージ 4)[25-27]。レット症候群患者の死後脳を 観察すると、健常者と比較して脳の大きさは小さく、軽 量であり、神経細胞の樹 状突起数や分枝の減少が見られた[28, 29]。レット症候群患者の多くの例で摂食嚥 下障害がみられるため胃瘻が必要となる患者もおり、患者の成長曲線は、体重、 身長、BMI の標準曲線を下回り、思春期以降に患者の身長や体重の成長が止まる と言われている[30-34]。しかしながら胃瘻を介し肺炎を引き起こすこともあるた め、患者の健康管理には慎重に取り組む必要がある[35]。 第2 節 MECP2の機能と役割 本疾患を発症する主な原因遺伝子であるMECP2は、DNA のメチル化されたシ トシンに結合するメチル化 DNA 結合タンパク質の一つとして発見された[36]。 DNA のメチル化とは、塩基配列の変化を伴わない遺伝子の発現抑制に関与する機 構であり、ヒトの発達においても重要な役割を果たしている[37]。このメチル化は 体細胞の組織によってメチル化のパターンが異なり、脳においては 神経の種類に よってメチル化のパターンが異なることが知られている[38-42]。近年の研究では、 MeCP2 がメチル化 CpG サイトだけでなくメチル化 CpH (H=A/C/T)にも結合する ことが明らかとなり、特にヒドロキシメチルシトシン(hmC)と mCpH は神経系の 細胞に多く存在することが報告されている [43-45]。加えて、年齢を重ねるととも にメチル化 CpH に MeCP2 が結合することにより、結合パターンの異常性が潜在 的にレット症候群の後期発症に関与していることが報告されている[46]。また、in- 5 -
vitroの実験では DNA 中の 5-ヒドロキチメチルシトシンに MeCP2 が結合し、こ れらは活性化している遺伝子や脳において豊富であることが知られている[40, 45, 47, 48]。 これらと一致して、MeCP2 は非常に様々な組織発現しているが、特に有糸分裂 後のニューロンにおいて豊富に発現していることが知られている[49-53]。MECP2 は4 つの exon から成り立ち、N 末端が異なる複数のスプライシングバリアントが 存在する[53-55]。しかしながら、すべてのアイソフォームで Methyl-CpG binding domain (MBD; メチル化 CpG 結合ドメイン)と transcriptional repressor domain (TRD; 転写抑制ドメイン)という 2つの機能ドメインを有している[56-59]。マウ スを用いた研究では、スプライシングバリアントである MECP2e1 は MECP2e2 に比べて脳内での発現が高く、また、Mecp2e1 特異的に変異を有するマウスは異 常行動を示したのに対し、Mecp2e2 特異的に変異を有するマウスは異常行動を示 さなかった[53, 54, 60]。一方、Mecp2e2 は胎仔の胚生存率や胎盤発達に非常に重 要な役割を果たすことが知られている[61]。 また、TRD 領域は Histone Deacetylase(HDAC; ヒストン脱アセチル化酵素)を 含む SIN3A や NCoR、SMRT コンプレックスといったコリプレッサーと結合し、 転写制御の役割を果たす[58, 62-64]。MECP2中の TRD 領域は NCoR/SMRT コリ プレッサーコンプレックスと結合するのに必須の領域である。NCoR コンプレッ クスは通常、MeCP2 の 308 番目のリン酸化スレオニン残基(Thr308)と 421 番目 のリン酸化セリン残基(Ser421)と相互作用しており、この相互作用を失うことで レット症候群の表現型が現れることが判明している。より詳細に述べると、MeCP2 のThr308 および Ser421 部位がリン酸化されることで細胞膜の脱分極を誘導する。 一方で、86 番目のセリン残基(Ser86)と 274 番目のセリン残基(Ser274)のリン酸化 は、脳由来神経因子であるbrain-derived neurotrophic factor (BDNF) やアデニ ル酸シクラーゼを活性化させるフォルスコリンの刺激を受けて誘導される[65]。加 えて、MeCP2 がリン酸化されることで MeCP2 がBDNFプロモーター領域から離 れ、細胞膜の脱分極によって誘導されるカルシウム流入による神経活動依存的な 反応が生じる[51, 65-68]。近年の研究では MeCP2 の Ser421 リン酸化は、特定の ゲノム部位だけでなく全体で生じていることが報告された[69]。これらの報告から、 リン酸化が通常の神経回路発達に必要不可欠であり、MeCP2 のターゲット遺伝子 の分化に伴う 活性と抑 制に寄与する ことを指 示している。 さらに TRD 領域は AT-hook と呼ばれる特徴的なアミノ酸配列が存在し、この領域が含まれているか
- 6 - 否かでレット症候群の重症度が異なるという報告がされている[70]。MeCP2 中に は複数のAT-hook ドメインが存在し、このドメインは AT 配列が豊富な DNA 配列 を特に認識することで、この認識の結果DNA のコンパクションやオリゴマー形成 を引き起こす[70-72]。これらの発見は、変異 MeCP2 が NCoR-SMRT コリプレッ サーと結合できないために、レット症候群に至ることを示唆している。 一方、MeCP2 は当初メチル化 CpG サイトに結合する転写抑制因子として同定 されたが、cyclic AMP-responsive element-binding protein 1 (CREB1)に伴って、 転写活性因子としても働くことが明らかになっている[58, 62, 73]。この結果と一 致して、Mecp2-null マウスは転写が制御されており、MECP2に変異を加えたES 細胞由来の神経系細胞においてもまた転写の制御とタンパク質生産が低下するこ とが報告されている[74, 75]。
加えて、MeCP2 はクロマチン構造の再構成と、RNA と結合することで RNA プ ロセシングを媒介することも報告されている。例えば、異所性のMeCP2 発現は筋 分化において染色中心のクラスター形成を引き起こすことや、Dlx5-Dlx6 領域に おいて11kb のループ構造をとることなどが挙げられる[71, 76, 77]。Mecp2-null マウスを用いた研究では、RNA プロセシング中に MeCP2 が RNA と結合する Y-box 結合タンパク質 1(YB-1)と相互作用することで選択的スプライシングに関与 していることを示唆していた[78]。さらには、MeCP2 は選択的にスプライシング されたエクソンに豊富に存在し、DNA メチル化の阻害は MeCP2 のエクソンへの 特異的ターゲティングを阻害し、miRNA の産生を抑制することが明らかにされた [79, 80]。さらに近年の miRNA に関連した研究では、MeCP2 はタンパク質合成 や脂質合成を司るmTOR シグナリングを活性化させる miR-199a 産生を促進させ、 MeCP2-KO の神経細胞において miR-199a を発現させると神経学的な欠陥を回復 することが報告された[81]。 第3 節 遺伝子変異と病態の関連性 およそ90%近いレット症候群患者はMECP2に変異を有し、そのうちの65%は MBD 領域にある 4 か所の変異(R106W, R133C, T158M, R168X)と TRD 領域にあ る4 か所の変異(R255X, R270X, R294X, and R306C)に集中している[82, 83]。 また、遺伝子変異の位置も重症度に寄与することが知られており、C 末端での 変異やミスセンス変異、p.R133C、p.R294X、p.R306C などは軽症例として同定 さ れ る こ と が 多 い こ と に 対 し 、 欠 失 や 挿 入 、p.R106W、p.R168X、p.R255X、
- 7 -
p.R270X などはより重症例として報告されている[84-86]。さらに、重症例患者は 軽症例患者よりも身長、体重、BMI スコアを下回ることも報告されている[31, 32]。 第4 節 X chromosome inactivation とレット症候群について
遺伝子変異タイプに加え、レット症候群の重症度に関与するもう一つの要因と してX chromosome inactivation (XCI; X 染色体の不活化)が挙げられる。XCI と は、男性との遺伝子発現差異を埋めるために、通常 2 本存在する女性の性染色体 のうち片方の X 染色体がメチル化され不活化を受けることで遺伝子発現が抑制さ れる機構のことである[10, 11]。不活化を受ける X 染色体はランダムに決定され、 一度決定されたXCI パターンは細胞の分裂のすべての段階を通じて安定的に引き 継がれる[12, 13]。また、XCI は X-inactive specific transcript(XIST)と呼ばれる 17 kb のノンコーディング RNA によって不活化を受ける X 染色体がコーティング されることによって引き起こされる[87-89]。 ヒト ES 細胞を用いた研究から、未分化の細胞における XCI のパターンは 3 種 のクラスに分かれることが報告されている[90]。クラス 1 は、XISTの発現や転写 抑制のマーカーとして知られるヒストンH3K27 のトリメチル化(H3K27me3)など がみられない、両 X 染色体が活性化状態にある細胞株として定義される(XaXa, XIST(-))[90-93]。クラス 2 はどちらか一方の X 染色体が XIST によってコーティ ングされ、不活化されたX 染色体上に H3K27me3(XaXi, XIST(+))が現れる[90, 91, 94]。クラス 3 はクラス 2 同様に不活化された X 染色体を有するが、XISTの発現 は失われている(XaXi, XIST(-))[90-94]。クラス 1 に属する細胞群は pre-XCI とも 呼ばれる一方で、クラス2,3 に属する細胞群は post-XCI と呼ばれている。 MECP2はX 染色体上に存在するため、ランダムな XCI に伴い患者の体細胞で は正常 MECP2 を有する X 染色体が不活化されると変異 MeCP2 が産生され、変 異MECP2 を有する X 染色体が不活化を受けると正常 MeCP2 が産生される。し たがって、患者の体細胞は正常 MECP2が発現する細胞と変異 MECP2 が発現す る細胞の 2 種類が混在する。一方で、男性の場合はすべての細胞で変異 MeCP2 タンパク質が産生されるため、多数のケースで胎生致死または乳幼児期に重度な 脳障害が生じ死亡する[14-16]。 第5 節 神経学的解析におけるレット症候群の疾患モデル 前述のようにレット症候群は遺伝子変異が明らかになっているため、マウスの
- 8 - 疾患モデル動物がいくつか存在する。Mecp2 欠損マウスはレット症候群の患者が 呈する症状をよく再現していることが知られている。例 えば、雄マウスにおける 短命さ、歩行障害、振戦、呼吸異常や、脳における樹状突起の数や重量の減少が 挙げられる[5, 6, 51, 95-97]。また、MeCP2 の過剰発現や発現抑制モデルのどちら でも、神経障害が引き起こされることも報告されている[98-103]。ニューロンにお いては、MeCP2 発現レベルは神経分化の最中に増加し、Mecp2変異マウスは樹状 突起やスパインの減少やシナプス形成不全、神経が未成熟であることなどが報告 されている[52, 104-110]。一方、有糸分裂終了後の神経細胞中の Tau 領域での MeCP2 の発現を誘導すると、レット症候群の表現型を緩和することができた[111]。 そのほかのマウスを用いた報告としては、 第 2 節でも述べたように BDNF は MeCP2 の票的遺伝子として知られており、BDNF の過剰発現は、Mecp2 欠損マ ウスの特徴である短命さやシナプス機能不全 といった症状を緩和させることがで きた[66, 67, 112-114]。 2007 年に、2 つの研究グループからヒト ES 細胞と同等の性質を持つヒト iPS 細胞(以下、特に注釈がない限り ES 細胞または iPS 細胞と略記する)という非 常に有益な技術が報告された[19, 20]。この技術は、ワディントンが提唱したエピ ジェネティック・ランドスケープの壁を乗り越え、終末分化した細胞が再び多能 性を獲得できるようになった。レット症候群を含む神経発達障害や神経変異疾患 の研究を行う上で神経細胞の役割を知ることは非常に有益であることに疑いはな いが、神経細胞は患者より剖検以外の手段で検体を採取することが殆ど不可能で あるため、iPS 細胞は神経疾患解析において非常に有用なツールであると期待され る。これまでに様々な研究グループからレット症候群患者由来の iPS 細胞が樹立 され、それらから誘導した神経は、これまでにマウスモデルで明らかにされた神 経学的欠陥と一致した[24, 115-121]。いくつか詳細な例を挙げると、レット症候 群患者由来の iPS 細胞を用いた初めての報告では、患者由来の神経細胞では健常 者由来の神経細胞と比較して、シナプス形成、樹状突起やスパインの数が少なく、 細胞体の大きさが小さいことを報告している[24]。この研究成果は、細胞のリプロ グラミングを経てもなお、レット症候群の表現型が維持されることを強く示唆し たものである。そのほか、MECP2e1にのみ変異を有するレット症候群患者由来の ニューロンは内在性の膜機能に異常が認められ、活動電位の低下や、ニューロン の細胞体の大きさが小さいことなどが報告されている[121]。一方、グリア細胞の 一種であるアストロサイトに視点を向けると、変異 MECP2 が発現しているアス
- 9 - トロサイトは、野生型のニューロンに対して形態的にも機能的にも悪影響を与え ることが報告されている[120]。 iPS 細胞の最大の利点は、適切な分化方法を用いることで機能的な神経やグリア 細胞へと分化させることができ、比較検討ができる点である。iPS 細胞由来のニュ ーロンサブタイプを誘導するために数多くのプロトコルが報告されており、研究 者はそれぞれの目的に応じた誘導法を選択することで研究を進めることができる。 iPS 細胞を用いたレット症候群研究の場合、他の疾患研究と比較して有利な点は、 種々のゲノム編集なしに X 染色体の不活化に伴って変異を有する疾患モデルの細 胞株と対照となるコントロールの細胞株を得ることができる点にある[115-120, 122, 123]。いずれにせよ、レット症候群を含む疾患特異的 iPS 細胞は、疾患メカ ニズムを明らかにするだけでなく、薬効評価や移植治療といった幅広い分野での 利用が期待されている。
一方で、ES 細胞や iPS 細胞には様々な XCI 状態が存在することが知られてお り、培養環境や試薬の添加によってXCI が進行または不活化を受けた X 染色体が 再活性化を受けることなどが知られている[118, 124-129]。そのため、レット症候 群のような X 染色体連鎖の疾患においては、XCI による遺伝子発現パターンによ って翻訳されるタンパク質に違いが生じるため、研究を進めるうえでゲノム安定 性の確認は重要な項目である。 本研究では、小児期に発症する進行性の神経系疾患であるレット症候群を対象 に、患者の協力を得て皮膚繊維芽細胞より iPS 細胞を樹立し、継代培養を通じた iPS 細胞の性質安定性の評価と神経系細胞への分化異様の探索を、塩基配列の変化 を伴わない X 染色体の不活化や DNA のメチル化といったエピジェネティック特 性の観点から解析を行った。
- 10 -
第
1 章 レット症候群患者由来疾患特異的 iPS 細胞の樹立
第1 節 序 2007 年に誕生した iPS 細胞は、疾患研究だけでなく創薬研究や再生医療など幅 広い分野での活躍が期待されている[19, 20]。iPS 細胞は ES 細胞同様に無限の自 己増殖能と多分化能を有する細胞と定義され、この細胞から研究対象となる細胞 を分化誘導することで、疾患の基礎研究や創薬研究に有用なツールとなるだけで なく、ES 細胞が抱えていた受精卵を破壊してしまうという倫理的問題や他家移植 による免疫不全などの問題が解決され、再生医療・細胞移植治療への応用も期待 されている。 本章では小児期に発症する進行性の神経系疾患であるレット症候群を対象に、 患者の協力を得て皮膚繊維芽細胞より疾患特異的 iPS 細胞を樹立し、将来的な疾 患解析の基となるツールを作成した。- 11 - 第2 節 実験材料および方法 1) 研究倫理申請 本論文における以下すべての研究は、十分なインフォームドコンセントの後に レット症候群患者側からの承諾を得て皮膚生検が行われた。また、樹立された皮 膚繊維芽細胞と iPS 細胞の樹立および分化誘導は、慶應義塾大学医学部倫理委員 会(承認番号 20080016)および山梨大学倫理委員会(承認番号 523 および 699) にて受理されている。なお、これらはすべてヘルシンキ宣言に則って実施された。 2) レット症候群患者の選定 本研究では、DNA 多型マーカーで一卵性双生児であること、および疾患の遺伝 子変異型が同一であることが確認されている 2 名の女性のレット症候群患者を選 定した(以下RS1, RS2 と表記する)。遺伝子変異は MECP2に生じ、806 番目の G 塩基の欠損(一塩基欠損、フレームシフトタイプ)であることが確定している [130]。皮膚生検は、患者両名が 10 歳の時点で行われた。 3) 皮膚繊維芽細胞(Fibroblasts)の樹立 患者協力のもと皮膚生検を行い、採取した皮膚組織を 1 mm 角に裁断して培養 dish に接着させ、10% FBS (Gibco), 2 mM L-グルタミン(Nacalai tesque)および 50 units および 50 mg/mL ペニシリン・ストレプトマイシン溶液(Nacalai tesque) 含有のDMEM(Wako)培地で培養し樹立した。
4) フィーダー細胞の調整
十分に増殖した SNL76/7 細胞(Wellcome Trust Sanger Institute)に対し、12.4 µg/mL となるように mitomycin C solution (Wako)を加え、3 時間インキュベータ ーにて有糸分裂的に不活化処理を行った。フィーダー細胞の培養には 7% FBS (Gibco), 2 mM L-グルタミン(Nacalai tesque)および 50 units および 50 mg/mL ペ ニシリン・ストレプトマイシン溶液(Nacalai tesque)含有の DMEM(Wako)培地で 培養した。
5) iPS 細胞の樹立と培養方法
高橋らの論文に従ってiPS 細胞の樹立及び継代培養を行った[19]。Fibroblasts に対し、マウスのレトロウイルスに対するレセプターを pLenti6/UbC-Slc7a1 プラ
- 12 -
スミドを用いて発現させたのち、初期化 4 因子(OCT3/4, SOX2, c-MYC, and KLF4)をレトロウイルスベクターによって導入した。導入後の fibroblasts をフィ ーダー細胞に播種し、iPS 細胞のコロニーの出現を待った。コロニーが現れたのち に、フィーダー細胞を播種した別の培養皿に単離し細胞株化した。インキュベー ターは、3%CO2, 20%O2の環境を用意した。培地は DMEM/F12 (Wako) に対し、
20% KSR (Gibco), 4 ng/mL Recombinant basic fibroblast growth factor, human (FGF-2) (PeproTech), 2 mM L-グルタミン(Nacalai tesque), 0.1µM
NEAA(Sigma), 0.1µM メルカプトエタノール, 50 units および 50 mg/mL ペニシ リン・ストレプトマイシン溶液(Nacalai tesque)を加えたものを利用した。継代は、 0.25% w/v トリプシン(Gibco), 0.1 mg/mL Collagenase IV (Gibco), 1 mM CaCl2,
20% KSR (Gibco)で組成される解離液を反応させることでフィーダー細胞を除去 し、iPS 細胞のコロニーを回収し適当な大きさにコロニーを砕いたのち、新しいフ ィーダー細胞上へ播種して行った。 6) X 染色体の不活化パターンの判定 X 染色体の不活化パターンは、久保田らによって構築されたヒトアンドロジェ ン 受 容 体 遺 伝 子(HUMARA) の メ チ ル 化 状 態 を メ チ ル 化 特 異 的 PCR ( 以 下 HUMARA-MSP とする)により解析し、解析結果に基づいてX染色体の不活化の 状態を検出する手法を用いた[131]。本手法は、メチル化特異的(AR-M)プライマー と非メチル化特異的(AR-U)プライマーの 2 種類のプライマーを用いることで、X 染色体の不活化状態を判定することができるものである。Xq11-12 に存在する HUMARAは、exon 1 に CAG の繰り返し塩基配列によって構成される多型領域が 存在し、通常この領域は父親由来と母親由来とでは繰り返し回数 が異なるため X 染色体の由来を識別することが可能である。メチル化特異的(AR-M)プライマーに よる増幅は、CpG ジヌクレオチドがメチル化されている場合に起こり、そして、 PCR 産物はX染色体が不活性の場合のみに得られる。同様に、非メチル化特異的 (AR-U)プライマーによる増幅は、CpG ジヌクレオチドがメチル化されていない場 合に起こり、PCR 産物はX染色体が活性の場合のみに得られる。体細胞はランダ ムな不活化を受けているため、1 種類のプライマーから父親由来および母親由来の X 染色体に由来する 2 種類の PCR 産物が得られ、その PCR 産物量の比率によっ て X 染色体の不活化状態を判断することができる。さらに、両親の協力が得られ ている場合は、父母のどちら由来の X 染色体が不活化を受けているかを明らかに
- 13 -
することができる。DNA 抽出は DNeasy (QIAGEN)を用いて行い、その後のバイ サルファイト処理はEZ DNA Methylation-Lightning™ Kit (Zymo Research)を用 いた。2 種類のプライマーを用いて PCR を行い、2.5%アガロースゲル電気泳動お よびGene Scan®ソフトウェアを備えたABI310 DNA シーケンサーを用いて、PCR
産物のピーク面積比による X 染色体の不活化パターンを判定した。プライマーは 下記の通りであり、Forward 側に FAM 蛍光を標識したものを用いた(Table 1)。 Table 1. List of PCR Primers.
Primer Sequence (5’ to 3’)
AR-M Forward (methylated) GTT GTG AGT GTA GTA TTT TTT GGT AR-M Reverse (methylated) CAA ATA ACC TAT AAA ACC TCT ACA AR-U Forward (unmethylated) GCG AGC GTA GTA TTT TTC GGC
AR-U Reverse (unmethylated) AAC CAA ATA ACC TAT AAA ACC TCT ACG
7) 蛍光免疫染色 リン酸緩衝液(PBS; Nacalai tesque)を溶媒とし、4%パラホルムアルデヒド溶液 を作成し細胞を固定する。培養液と等量の 4%パラホルムアルデヒド溶液で 10 分 室温にて固定し、上清をアスピレート後、再度 4%パラホルムアルデヒド溶液を加 え、10 分間室温にて固定した。PBS で 2 回洗浄後ブロッキング溶液(PBS, 0.3% triton-X100 および 5% FBS)を加え 1 時間静置した。次に、ブロッキング溶液で 希釈した 1 次抗体を加え、冷蔵庫内で一晩一次抗体反応を行った。一次抗体反応 後PBS を用いて 2 回洗浄し、ブロッキング溶液で希釈した一次抗体のホストと対 応する二次抗体 (Invitorogen)および Hoechst 33342 を用いて室温で 1 時間二次 抗 体 反 応 を 行 っ た 。 二 次 抗 体 反 応 後 PBS で 洗浄し、マウント材(Lab Vision™ PermaFluor™ 水性封入剤; Thermo Fisher Scientific)を用いてスライドガラスへ とマウントした。作成したスライドは蛍光顕微鏡(Apotome; Zeiss)で観察した。抗 体は下記のリストを用いた(Table 2)。
Table 2. List of Antibodies.
Antibody Dilution Source Catalogue number
- 14 -
Phalloidin 1:2000 Dyomics 490-33 TRA-1-81 1:1000 Millipore MAB4381 Nanog 1:100 COSMO BIO Co., LTD. RCAB0004P-F Alexa Fluor® 488
ヤギ抗マウス IgG 二次抗体
1:1000 Thermo Fisher Scientific A11029
Alexa Fluor® 555 ヤ ギ 抗 ラ ビ ッ ト IgG 二次抗体
1:1000 Thermo Fisher Scientific A21429
8) テラトーマ形成
免疫不全動物である NOD.CB17-Prkdcscid/J、通称 NOD/SCID mouse の精巣 に iPS 細胞を注入しテラトーマ形成を行うことで、三胚葉分化の確認を行った。 購入時は6 週齢のオスのマウスを用意した。術後 8 週を目途に 4%パラホルムアル デヒドを用いて還流固定を行いサクリファイスし、精巣を取り出し切片を作成す る。作成した切片は組織染色としてヘマトキシリン・エオシン染色(HE 染色)を行 った。 9) 核型解析 ギムザ染色を用いた G-バンド法にて核型解析を行った。
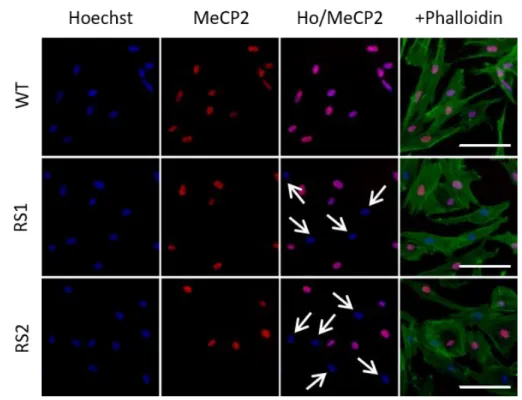
- 15 - 第3 節 結果 1) fibroblasts の樹立とモザイク現象の確認 MECP2に変異を有するレット症候群患者の場合、体細胞においてMeCP2 のモ ザイク現象が生じていると考えられる。樹立したfibroblasts におけるモザイク現 象とMeCP2 陽性細胞の存在比率を確かめるために、抗 MeCP2 抗体を用いて免疫 染色を行った。細胞骨格はアクチン繊維と結合する蛍光標識ファロイジンを用い て標識した。健常者由来の fibroblasts では、すべての細胞で MeCP2 と核染色で あるHoechst の共染色を認めた。一方、レット症候群患者由来の fibroblasts (RS1 およびRS2)では MeCP2 では染色されない核が存在した(Figure 1)。MeCP2 陽性 細胞の数をカウントした結果、RS1 では約 64%、RS2 で約 60%の細胞のみが MeCP2 陽性細胞であった。以上のことから、樹立された fibroblasts は正常MECP2 発現細胞と変異 MECP2 発現細胞の 2 種類の混在状態であることが確認できた (Figure 2)。
Figure 1. Immunostaining for MeCP2 (red) and phalloidin (green) along with Hoechst staining (blue) of wild-type MECP2- and mutant MECP2-expressing fibroblasts. Arrows indicated nuclei stained with Hoechst only. WT, wildtype. Scale bar, 100 μm.
- 16 -
Figure 2. Fraction of MeCP2-positive cells among wild-type MECP2- and mutant MECP2-expressing fibroblasts (n = 5 experiments; > 500
Hoechst-positive cells per experiment; *p < 0.05). (関連論文 (4) より転載)
- 17 -
2) iPS 細胞の樹立
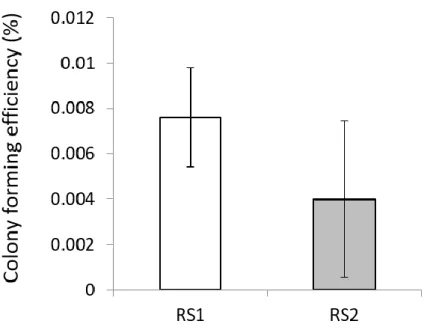
正常MECP2発現細胞と変異MECP2発現細胞の2 種類の混在状態である fibroblasts よりレトロウイルスを介して初期化 4 因子(OCT3/4, SOX2, c-MYC, KLF4)を導入することで、レット症候群患者由来疾患特異的 iPS 細胞の樹立を試 みた。ES-like な形態を示した株をクローン化し、RS1 からは 44 クローン、RS2 からは37 クローンが樹立された。両者の iPS 細胞クローン樹立効率は、統計学的 な有意差は存在しなかった(Figure 3)。
Figure 3. iPS cell formation efficiency of 50,000 infected fibroblasts derived from Rett syndrome patients at 28 days after reprogramming.
- 18 -
3) iPS 細胞の X 染色体の不活化パターンの判定
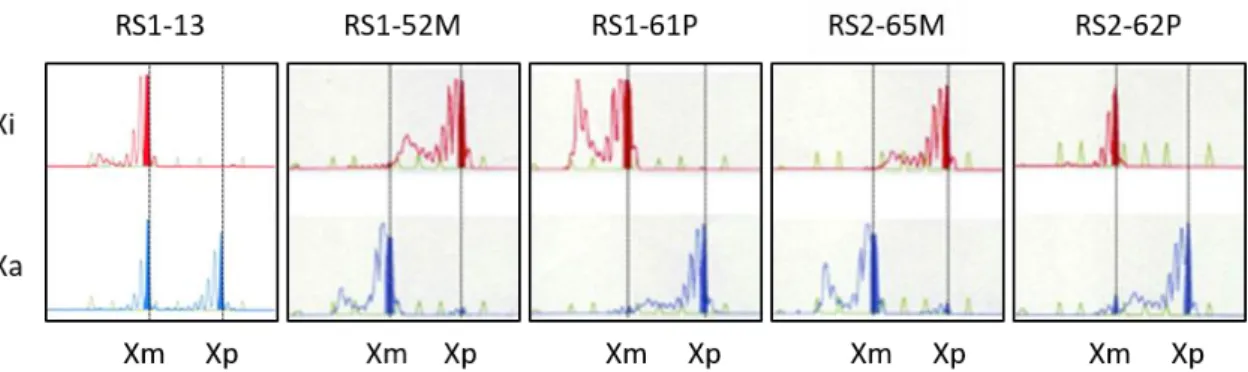
レット症候群の研究においては、XCI による X 染色体の発現パターンがMECP2 の発現状態を左右するため、樹立された iPS 細胞の XCI パターンを
HUMARA-MSP を用いて解析を行った。はじめに、HUMARA-MSP で得られた PCR 産物を電気泳動したところ、大多数の iPS 細胞株は AR-M プライマーと AR-U プライマーのそれぞれから1つずつのPCR 産物が得られた。これはすなわち、ど ちらかのX 染色体が不活性化を受け、1 本の活性化 X 染色体が存在することを示 唆している(活性化X 染色体; Xa、不活性化 X 染色体; Xi と表記し、以下 XaXi 株とする)。一方、一部の株では不活化を受けているX 染色体が同時に活性化状態 にあるという、両X 染色体部分活性状態である株も得られた(以下、partial-XaXa 株とする)。しかしながら、AR-U のみで増幅された株、つまりマウス ES 細胞で 見られるような両X 染色体が活性化している株は存在しなかった(Table 3)。 また、今回のレット症候群患者の場合は両親の協力が得られているため、どち らのX 染色体が不活化を受けているか判定することが可能であり、さらには、変 異のMECP2は父親由来の X 染色体に由来するということが明らかになっている [130]。そこで、樹立した株の中から partial-XaXa 株から 1 株(RS1-13), XaXi 株 より母由来X 染色体発現株(RS1-52M および RS2-65M), および父由来 X 染色体発 現株(RS1-61P および RS2-62P)の合計 5 株について、iPS 細胞の特性解析を行っ た。5 株はすべて未分化マーカー(NANOG および TRA-1-81)を示し、テラトーマ 形成による三胚葉分化能を有することが確認できた(Figure 4, Figure 5)。また、 染色体異常の有無を確認するためにこれらを核型解析したところ、すべての株で 正常な核型であることが示された(figure 6)。以上の結果より、レット症候群患者 由来のfibroblasts より 3 種類の iPS 細胞が樹立できたと結論付けられた。
Table 3. Classification of XCI statuses in of Rett syndrome iPSCs*
Patient Maternal X active clones
Paternal X active
clones Partial-XaXa clones
RS1 3 36 5
RS2 32 1 4
- 19 -
Figure 4. XCI patterns in the five hiPSC lines, as assessed by the
methylation-specific PCR-based HUMARA assay. Xi, X inactivation pattern based on the inactive X chromosome; Xa, X inactivation pattern based on the active X chromosome; Xm, X chromosome inherited from the mother; Xp, X chromosome inherited from the father.
- 20 -
Figure 5. Characterization of Rett syndrome iPSCs.
iPSCs demonstrate similar embryonic stem cell-like morphology and stain positively for the pluripotency markers, NANOG and TRA-1-81. In addition, the teratomas corresponded to well-defined, cystic tumors containing tissues of all three germ layers (endoderm, mesoderm, and ectoderm). Scale bar for phase contrast images (PH), 500 μm; scale bar for NANOG, and TRA-1-81
immunostaining, 50 μm. Scale bar for teratomas, 100 µm. (関連論文 (4)より一部改変の上転載)
- 21 -
Figure 6. Images of Rett syndrome iPSC karyotypes. Standard G-banding analysis was performed in the five iPSC lines.
- 22 - 第4 節 考察 レット症候群患者の皮膚繊維芽細胞から疾患特異的 iPS 細胞を樹立することが できた。また、得られたiPS 細胞の大多数は、リプログラミングが生じた皮膚繊 維芽細胞のX 染色体のメチル化状態、すなわち X 染色体の不活化パターンを維持 したものと考えられる。一方で、不活化を受けつつも 2 本の X 染色体が活性化し ている株の存在から、iPS 細胞の樹立時のリプログラミングは、少なくとも 2 本 のX 染色体が再活性化を受けるところまで初期化されることが判明したが、完全 な両X 染色体活性化状態の株は得られなかった。この結果より過去の知見でも明 らかになっていた通り、ES/iPS 細胞の XCI 状態はマウスのそれとは性質が異なる ことを改めて証明した[124]。 加えて、モノクローナル的に得られたMECP2に変異を有する細胞は、XCI に よってMECP2の発現状態が変わるため、X 染色体の不活化パターンを判定する ことは非常な重要な意義を有する。実験の目的に応じて適切な細胞株を選ぶため に、HUMARA-MSP などの手法を用いて X 染色体の不活化パターンを判定するこ とが必須である。
- 23 -
第
2 章 X 染色体不活化解析を用いた iPS 細胞の性質安定性評価の試み
第1 節 序
X 染色体の不活化(X chromosome inactivation; XCI)は、男性と女性の性染色 体による遺伝子発現の差を埋めるために生じる、エピジェネティックな現象の一 つである[10, 11]。XCI は X-inactive specific transcript (XIST)と呼ばれる 17 kb のノンコーディング RNA によって不活化を受ける X 染色体がコーティングされ ることによって引き起こされる[87-89]。ES 細胞を用いた研究から、未分化の細胞 における XCI のパターンは 3 種のクラスに分かれることが報告されている[90]。 クラス1は、XIST の発現や転写抑制のマーカーとして知られるヒストン H3K27 のトリメチル化(H3K27me3)などがみられない、両 X 染色体が活性化状態にある 細胞株として定義される(XaXa, XIST(-))[90-93]。クラス 2 はどちらか一方の X 染 色 体 が XIST に よ っ て コ ー テ ィ ン グ さ れ 、 不 活 化 さ れ た X 染 色 体 上 に H3K27me3(XaXi, XIST(+))が現れる[90, 91, 94]。クラス 3 はクラス 2 同様に不活 化されたX 染色体を有するが、XISTの発現は失われている(XaXi, XIST(-))[90-94]。 クラス1およびクラス2の細胞に属する細胞の XCI 状態は不安定で、培養を経る ことでクラス3 へと移行しやすいことが明らかになっている[90-94]。 これまでに述べてきたように、MECP2に変異を有するレット症候群の疾患解析 を行う上では、X 染色体の不活化パターンは MECP2 の発現に直結するため注視 する必要がある。本章では、第1 章で得られた不活性化を受けている X 染色体が、 同時に活性化も受けているという両X 染色体部分活性状態である株の XCI 状態が 培養中に変化し得るか否かを判定するために、経時的に HUMARA-MSP を行うこ とでXCI パターンを判定した。
- 24 - 第2 節 実験材料および方法 1) 使用した iPS 細胞について 第1 章で樹立した iPS 細胞の中から、樹立時に partial-XaXa 株と判定された株 3 株(RS1-13, RS1-44, および RS1-76)を選択した。加えて、健常者の皮膚繊維芽 細胞由来のiPS 細胞(WD5 および WD37)の 2 種類を今回の解析に加えた。 2) 細胞の培養方法 第1 章第 2 節 5)を参照のこと。 3) 実験の手順 本実験は、同一条件での培養下における XCI 状態の変遷を確認するために行っ た。継代ごとに経時的に DNA を回収し、HUMARA-MSP を用いて XCI パターン の解析を行った。
4) X 染色体の不活化パターンの判定 第1 章第 2 節 6)を参照のこと。
- 25 -
第3 節 結果
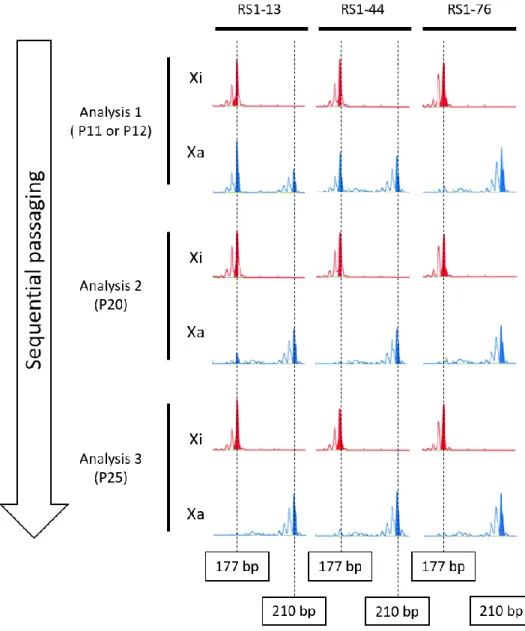
樹立時に不活性化を受けている X 染色体が、同時に活性化も受けているという 両X 染色体部分活性状態である株 3 種類(RS1-13, RS1-44, および RS1-76)につい て、経時的にDNA を回収して HUMARA-MSP を行い、XCI パターンを解析した。 樹立時からいくつか継代を経た RS1-13 および RS1-44 は AR-M プライマーで 1 種類、AR-U プライマーで 2 種類の PCR 産物が得られたのに対し、RS1-76 は AR-M およびAR-U のどちらからも 1 種類の PCR 産物しか得られなかった(Figure 1)。 これらのiPS 細胞の継代を続けて、Gene Scan®を用いてX 染色体の不活化パター
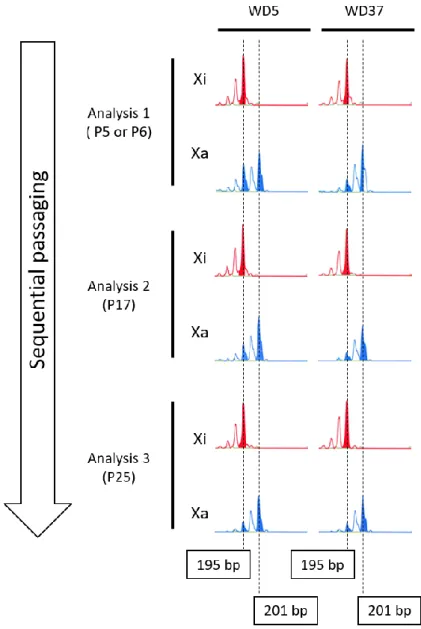
ンについて詳細な検討を行った。Gene Scan®での解析は、PCR 産物がピークとし て表示される。P11(または P12)では AR-U では 2 種類のピークが存在していた が、継代を経るごとにピークが減少していくことが示された。同時に、RS1-76 の 結果から、一度不活化が決定されたX 染色体は、少なくとも P25 までの間に再活 性化を示すことはなかった(Figure 2)。 健常者由来の iPS 細胞における XCI パターンについても検討するため、WD5 とWD37 という新たな細胞株を樹立した。健常者の iPS 細胞は、父由来 X 染色体 と母由来X 染色体の間で CAG リピートがわずか 6 bp の差しか存在しなかったた め(195 bp および 201 bp)、電気泳動による判定は不可能であった。一方、Gene Scan®での解析は、電気泳動では判定できない細かな差を明らかにすることができ た。解析の結果、健常者由来の iPS 細胞株においてもピーク面積の減少が確認で きた(Figure 3)。以上の結果から、一般的な iPS 細胞培養環境下では FGF-2 とい う未分化維持因子を添加していても、XCI が進行し不安定な不活化状態から安定 的な状態へと変遷することが証明できた。
- 26 -
Figure 1. Electrophoretic separation of the HUMARA-MSP products in established Rett syndrome iPSCs. Lanes 1,3, and 5, PCR products amplified from methylated primers (i.e. inactive X). Lanes 2,4, and6, PCR products amplified from unmethylated primers (i.e. active X). Genomic DNA of RS1-13 and RS1-76 were sampled from P11 iPSCs, and RS1-44 was sampled from P12 iPSCs.
- 27 -
Figure 2. XCI patterns in Rett syndrome iPSC lines (RS1-13, RS44, and RS1-76). Red peaks indicate PCR products amplified with AR-M primers (i.e., inactive X chromosome; Xi); blue peaks indicate PCR products amplified with AR-U primers (i.e., active X chromosome; Xa). The X-axes of several peaks shows the number of base pairs, which is calculated by the ABI 310 DNA sequencer equipped with Gene Scan® software; The Y-axes indicate the peak
heights as determined by the area derived from PCR products. Xa and Xi peak area ratios represent XCIpatterns.
- 28 -
Figure 3. XCI patterns in control iPSC lines (WD5 and WD37). Peak colors and axes are same in figure 2.
- 29 - 第4 節 考察 以上の結果より、partial-XaXa 株の XCI パターンは継代を重ねることで変化し、 時間の経過とともにXCI が進行する様子を明らかにした。これらの結果は既報の 内容と合致しており、樹立直後のiPS 細胞の XCI 状態は不安定であると考えられ る[91, 94, 124, 132, 133]。レット症候群を含む X 染色体に原因遺伝子を有する疾 患について研究を進める場合、X 染色体の発現パターンによって責任遺伝子の発 現が変化しうる。幸運なことに、X 染色体連鎖疾患の場合はモノクローナルな体 細胞から樹立されることで、ゲノム編集なしに疾患タイプの細胞と対照のコント ロール細胞の 2 種類の株を得ることができるが、それらを用いて遺伝子変異によ り引き起こされる影響を検討する上ではゲノム状態の安定性が担保されていなけ ればならない。iPS 細胞の XCI 状態はどちらかの X 染色体が不活化を受けている 状態が一般的であるとされる一方で、いくつかの研究グループから不活化を受け たX 染色体の再活性化が報告されている[118, 124-129]。したがって、若い継代数 時を含む、iPS 細胞の維持培養中に引き起こされる XCI パターンの変化を慎重に 評価する必要がある。そのXCI パターン判定法として、HUMARA-MSP は簡便で 確実な方法として有用であると言える。
- 30 -
第
3 章 レット症候群由来疾患特異的 iPS 細胞を用いた神経系分化におけ
るエピジェネティック特性の解析
第1 節 序 序論で述べたように、レット症候群は小児期に発症する進行性の神経系疾患で ある。第1 章において、母由来 X 染色体発現 iPS 細胞株(RS1-52M および RS2-65M) と父由来 X 染色体発現 iPS 細胞株(RS1-61P および RS2-62P)の iPS 細胞を樹 立し特性評価を行った。これまでの研究で、本研究に協力いただいた患者の変異 は父由来X 染色体に生じていることが判明している[130]。すなわち、母親由来 X 染色体が発現している細胞では正常 MECP2が、父親由来 X 染色体が発現してい る細胞では変異 MECP2が発現していると考えられる。そこでこれらの iPS 細胞 を用いて、神経分化におけるMECP2発現の有無による影響を調べた。- 31 - 第2 節 実験材料および方法 1) 使用した iPS 細胞について 第1 章で樹立した iPS 細胞の中から、樹立時に母由来 X 染色体発現 iPS 細胞株 (RS1-52M および RS2-65M)と父由来 X 染色体発現 iPS 細胞株(RS1-61P およ びRS2-62P)と判定された株を選択した。 2) 細胞の培養方法 第1 章第 2 節 5)を参照のこと。 3) 神経幹細胞(neurosphere)への分化誘導方法 神経幹細胞分化誘導法は松本らの論文の手法に従った[134]。基本培地は media hormone mix (MHM)と呼ばれる培地を用いた。MHM の組成は DMEM/F-12 (1:1) (Gibco), 0.6% glucose, 2 mM glutamine, 3 mM sodium bicarbonate, 5 mM HEPES, 25 mg/ml insulin, 100 mg/ml transferrin, 20 nM progesterone, 30 nM selenium chloride and 60 mM putrescine (DMEM/F12 を除きすべて Sigma)であ る。MHM に 2% B27® Serum-Free Supplement (Invitrogen) および 20 ng/ml
FGF-2 (PeproTech) を 加 え た も の を 、 神 経 幹 細 胞 の 誘 導 培 地 と し て 用 い た 。 TrypLE™ Select (Gibco) を用いて 5 分間 iPS 細胞を処理し、コロニーをシングル セルにした。T75 フラスコ(Nunc)に 1×104 cells/mL となるように細胞を調整し、
神経幹細胞誘導培地に 10 µM の Y-27632 (Wako)および 10 ng/ml となるように Leukemia Inhibitory Factor Protein, Recombinant human (Millipore)を加えて 低酸素インキュベーター(4%O2)にて培養を行った。14 日間ごとに neurosphere
をTrypLE™ Select を用いてシングルセルにし、この時点から 5×104 cells/mL に
細胞濃度を変更して継代を繰り返した。 4) 神経系細胞への分化誘導方法
神経幹細胞から神経系細胞への終末分化は、次の通り行った。poly-L-ornithine (Sigma)および Fibronectin (Sigma)(以下 PO/FN と略記)でコーティングを行っ た 6 well plate に対して、neurosphere を分散させて 1.2×106 cells 播種し、
Falcon® 1.0μm ポア透明 PET メンブレン付き 6 ウェルプレート用インサート
(Corning)を用いてインサート側に初代培養で得られたマウスアストロサイトを 敷き、マウスアストロサイトとの共培養を行った。免疫染色用には 24 well plate
- 32 -
の well 内に PO/FN コート処理を行った 14 mm のカバースリップ(Matsunami) を備え、その上に neurosphere をシングルセルの状態で 1.0×105 cells 播種し、
Falcon® 1.0μm ポア透明 PET メンブレン付き 24 ウェルプレート用インサートを
用 い て マ ウ ス ア ス ト ロ サ イ ト と 共 培 養 を 行 っ た 。 培 地 は MHM に 2% B27®
Serum-Free Supplement (Invitrogen)および 50 units および 50 mg/mL ペニシリ ン・ストレプトマイシン溶液(Nacalai tesque)を添加した培地で 30‐32 日間培養 を行った。
5) 蛍光免疫染色
細胞の固定方法、染色方法はは第1 章第 2 節 7) を参照のこと。一次抗体および 二次抗体は下記のものを使用した(Table 1)。
Table 1. List of Antibodies.
Antibody Dilution Source Catalogue number
MeCP2 1:200 Cell Signaling Technology #3456S OCT4 1:200 Santa Cruz Biotechnology, Inc. Sc-5279 III-tubulin 1:2000 Sigma T8660 MAP2 1:250 Sigma M4403 GFAP 1:500 Invitrogen 13-0300 Alexa Fluor® 488 ヤギ抗マウス IgG 二次抗体
1:1000 Thermo Fisher Scientific A11029
Alexa Fluor® 555 ヤ ギ 抗 ラ ビ ッ ト IgG 二次抗体
1:1000 Thermo Fisher Scientific A21429
Alexa Fluor® 555 ヤギ抗ラット IgG 二次抗体
1:1000 Thermo Fisher Scientific A21434
Alexa Fluor® 647 ヤギ抗マウス IgG 二次抗体
- 33 -
6) マイクロアレイ解析
RNA の回収は、iPS 細胞は継代のタイミングで、neurosphere は継代のタイミ ングで、神経系細胞は培養開始から30‐32 日のタイミングで、それぞれを 1 mL のTRIzol Regent (Life Technologies)に溶解し回収した。回収したサンプルにクロ ロホルムを200µL 加え、激しく混合させたのちに 15,000 rpm、4℃の条件で遠心 分離を行った。遠心後、上清に対し等量の 70%エタノールを加え、その混合溶液 をRNeasy (QIAGEN)のカラム上に添加した。その後の RNA 抽出は RNeasy のプ ロトコルに従った。2100 Bioanalyzer (Agilent Technologies)を用いて RNA のク オリティチェックを行ったのち、100 ng の RNA を用いて GeneChip® Human
Genome U133 plus 2.0 Array (Affymetrix)とハイブリダイズした。ハイブリダイ ズしたマイクロアレイはGeneChip® Scanner 3000 (Affymetrix)でスキャンされ、
MAS 5.0 アルゴリズムにてシグナル強度を数値化した。シグナル強度 50 以上と判 定されたサンプをlog 値に変換し、Cluster 3.0 Software を用いて階層型クラスタ リングを作成した[135]。PCA プロットは TIGR MeV (Multiple Experimental Viewer) Software を用いて作成し、スキャッタープロットは GeneSpring GX Software (ver. 12.6.1)を用いて作成した[136]。
7) 定量 RT-PCR 解析
RNA の回収は、マイクロアレイ解析時と同じ手法で行った。SuperScript III First-Strand Synthesis System for RT-PCR (Life Technologies) と oligo-dT primers を用いて、メーカーの手順書に従って逆転写を行い、cDNA 合成を行った。 定量RT-PCR は SYBR Premix ExTaq Tli RNaseH Plus (Takara)を用いて反応液 を 調 整 し ABI PRISM® 7900HT Sequence Detection System (Applied
BioSystems)にて解析を行った。定量 RT-PCR の反応は、下記のプライマーを用 いて行った(Table 2)。
Table 2. List of PCR Primers.
Primer Sequence (5’ to 3’)
GFAP-Forward TGTGAGGCAGAAGCTCCAGGATGA GFAP-Reverse AGGGTGGCTTCATCTGCTTCCTGT S100-Forward GTGGCCCTCATCGACGTTTT
- 34 - S100-Reverse ACCTCCTGCTCTTTGATTTCCTCT TUBB3-Forward ATTTCATCTTTGGTCAGAGTGGGGC TUBB3-Reverse TGCAGGCAGTCGCAGTTTTCAC MAP2-Forward GGCCCAAGCTAAAGTTGGTTCTC MAP2-Reverse GCAGTGACATCCTCAGCCAAAG ACTB-Forward TGAAGTGTGACGTGGACATC ACTB-Reverse GGAGGAGCAATGATCTTGAT
8) ク ロ マ チ ン 免 疫 沈 降 (chromatin immunoprecipitation; ChIP) お よ び ChIP-PCR ChIP は木村らが報告した手法を参考に、若干の変更を加えながらクロスリンク 法を用いて行った[137]。常温にて neurosphere に対し 1%ホルムアミド(Nacalai tesque)をもちいて、撹拌させながら 10 分間クロスリンク処理を行った。その後 終濃度200 mM となるようにグリシン溶液を加えて、撹拌させながら 5 分間反応 させた。その後遠心を行い、上清をアスピレートし使用するまで-80℃で保管した。 プロテアーゼインヒビターであるcOmplete, EDTA-free (Roshe)を含む溶解液(50 mM Tris-Hcl(pH 8.0), 1%SDS, 10mM EDTA)を用いて細胞を溶解後、Bioruptor UCD-300 (Diagenode, Inc)を用いてクロスリンクされた DNA の超音波破砕を行 った。超音波破砕は 30 秒処理、60 秒未処理を 6 サイクル行った。断片化された DNA を プ ロ テ ア ー ゼ ン イ ン ヒ ビ タ ー 含 有 の ChiP dilution buffer (50 mM Tris-Hcl(pH 8.0), 167 mM NaCl, 1.1% Triton-X 200, 0.11% sodium deoxycholate) と RIPA buffer-150 mM(50 mM Tris-Hcl(pH 8.0), 150 mM NaCl, 1mM EDTA, 0.1% SDS, 1% Triton X-100, 0.1% sodium deoxycholate)で適当量に 希釈した。希釈したDNA の一部をインプットとして回収し、−30℃で保存した。 続いて、希釈したDNA と Normal IgG および目的の抗体と反応させた磁気ビーズ (Dynabeads® M-280 Sheep anti-Rabbit IgG; Life Technologies)を、4℃で一晩反
応させた。翌日、磁気ビーズをRIPA buffer-150 mM(ただし、プロテアーゼンイ ンヒビター不含)およびRIPA buffer-500 mM(50 mM Tris-Hcl(pH 8.0), 500 mM NaCl, 1mM EDTA, 0.1% SDS, 1% Triton X-100, 0.1% sodium deoxycholate)で洗 浄後、TE buffer (10 mM Tris-Hcl(pH 8.0), 1 mM EDTA)で洗浄した。その後 ChIP 溶出buffer (10 mM Tris-Hcl(pH 8.0), 300 mM NaCl, 5mM EDTA, 0.5% SDS)を 200 µL 加え、インプットに対しても総量が 200 µL で終濃度 0.5% SDS となるよ
- 35 -
うにChIP 溶出 buffer を加えて、65℃で 8 時間以上インキュベートした。その後、 RNA 分解酵素である RNaseA (20 mg/mL; MACHEREY-NAGEL)を 2 µL 加えて 37℃で 30 分加温後、Proteinase K (20mg/mL; Sigma)を 5 µL 加えて、55℃で 1‐ 3 時間インキュベートした。その後、フェノール・クロロホルム・イソアミルアル コールを用いてDNA 抽出を行い TE buffer にて DNA を溶解した。ChIP-PCR 反 応は定量 RT-PCR における逆転写後と同様の手法を用いて行った。プライマーリ ストをTable 3 に、使用した抗体を Table 4 に記載した。
Table 3. List of ChIP-PCR Primers.
Primer Sequence (5’ to 3’)
STAT3 binding site-Forward TCATGCCCAGTGAATGACTC STAT3 binding site-Reverse AGATGCCAGGCTGTCAGG hGFAP exon1-Forward AGAGCAGGATGGAGAGGAGA hGFAP exon1-Reverse CCTTGAAGCC AGCATTGAGT
Table 4. Primary antibodies used for ChIP.
Antibody Source Catalogue number
MeCP2 Cell Signaling Technology #3456S normal rabbit IgG Santa Cruz Biotechnology, Inc. Sc-2027
9) Bisulfite sequencing
DNeasy (QIAGEN) を 用 い て DNA 抽 出 を 行 っ た の ち 、 Methylamp DNA Modification Kit (Epigentek)を用いてバイサルファイト処理を行った。STAT3 結 合サイトを含むGFAP プロモーター領域をPCR で増幅させた。増幅させた PCR 産物はpT7Blue T-vector (Novagen)を用いてクローニングを行い、12 個以上のシ ーケンス解析を行った。PCR のプライマーは次のとおりである(Table 5)。 Table 5. List of bisulfite sequencing Primers.
Primer Sequence (5’ to 3’)
hGFAP promotor-Forward TTGGGGAGGAGGTAGATAGTTAGGTTTT hGFAP promotor-Reverse CATCCCCTAATCCCCTTTCCTAAA
- 36 - 第3 節 結果 1) iPS 細胞における MeCP2 発現パターンの確認 第 1 章で樹立された母由来 X 染色体発現株(RS1-52M および RS2-65M)と父由 来X 染色体発現株(RS1-61P および RS2-62P)の iPS 細胞における MeCP2 発現パ ターンについて、未分化マーカーである OCT4 と MeCP2 の共染色を行った。母 由来X 染色体発現株では MeCP2 陽性であり、父由来 X 染色体発現株では MeCP2 陰性であることが確認できた(Figure 1)。
Figure 1. Characterization of iPSCs derived from the Rett syndrome monozygotic twins. MeCP2 (red) and OCT4 (green) expression in iPSC lines. Maternal wild-type MECP2 was preferentially active, exhibited MeCP2 expression in the nuclei, whereas the RS1-61P and RS2-62P iPSC lines, in which paternal mutant MECP2 was preferentially active, did not. Scale bar, 150 μm.
- 37 -
2) 神経分化能の確認
得られた4 種類の iPS 細胞から神経幹細胞を経て神経分化誘導を行ったところ、 ニューロンマーカーであるMicrotubule-associated Protein 2 (MAP2)陽性細胞、 およびアストロサイトマーカーであるGlial fibrillary acidic protein (GFAP)陽性 細胞を得ることができた。これらの神経系細胞は、iPS 細胞で示した MeCP2 発現 パターンを維持しており、X 染色体の不活化パターンは分化を通じても維持され ていることが証明された。加えてGFAP 陽性細胞では MAP2 陽性細胞と比較して MeCP2 の発現強度が弱いことが示された(Figure 2)。これは、既存のマウス神経 細胞の報告と一致していた[138, 139]。
Figure 2. Immunostaining of neural cells derived from Rett syndrome iPSCs. Immunostaining was performed to evaluate expression levels of MeCP2 (red), the neuronal marker, MAP2 (green), and the astrocytic marker, GFAP (magenta) in Rett syndrome iPSC-derived neural cells. Ho, Hoechst (blue). Arrowhead indicated MAP2, MeCP2 and Hoechst positive cells. Scale bar, 50 μm.
- 38 - 3) iPS 細胞および神経系細胞を用いた網羅的遺伝子発現解析 MeCP2 の有無が未分化の iPS 細胞および神経系細胞の転写ネットワークにどの ような影響を与えるかを特定するために、iPS 細胞と誘導した神経系細胞の網羅的 遺伝子発現解析を行った。PCA 解析の結果、iPS 細胞時は一つの領域に収束する も の の 、 神 経 系 細 胞 に お い て は X 染色体 の発現 パター ンに応 じて、 すなわ ち MeCP2 の発現パターンに応じて 2 つの領域に分かれた(Figure 3)。正常MECP2 発現株と変異 MECP2 発現株を比較するためにスキャッタープロットを描いたと ころ、未分化の iPS 細胞ではどの株同士を比較しても非常に高い相関係数(R2 > 0.99)を示した。一方で、神経系細胞へと分化させた場合、MECP2 の発現パター ンが同一の株同士を比較すると比較的高い相関係数(R2 > 0.99)を示したが(例; RS1-52M v.s. RS2-65 M または RS1-61P v.s, RS2-62P)、MECP2の発現パターン が異なる株同士を比較した場合、若干低い相関係数(R2 < 0.98)を示した(Figure 4)。 同様に、クラスター解析を行ったところ、未分化の iPS 細胞の状態では 4 つの細 胞株間に有意な差が認められなかったが、神経分化誘導後には MECP2 の発現パ ターンによって区別された(Figure 5)。
Figure 3. Results of PCA (performed by using MeV software (TIGR) Software) of microarray gene expression in Rett syndrome iPSC lines and neural cells differentiated from Rett syndrome iPSC lines. N, neural cells differentiated from iPSC lines.
- 39 -
Figure 4. Scatter plots of microarray gene expression in Rett syndrome iPSC lines and neural cells differentiated from Rett syndrome iPSC lines. Neural cells were co-cultured for ~30 days with mouse astrocytes.
- 40 -
Figure 5. Global gene expression/comparative microarray analyses of undifferentiated iPSCs and differentiated neural cells. N denotes neural cells. (関連論文 (4)より転載)
- 41 - 4) 変異MECP2発現株ではアストロサイトへの分化が促進される これまでの神経分化に関するMeCP2 の報告において、MeCP2 はアストロサイ ト関連遺伝子の発現を制御していることが報告されている[140-142]。また、Gfap および S100は MeCP2-null マウス ES 細胞から誘導されたアストロサイトにお いて、正常マウス ES 細胞と比較して高発現していることも報告されている[143, 144]。加えて、MeCP2 に R168X 変異を有するマウス神経幹細胞では、神経分化 が抑制される一方で、異常なアストロサイト分化が起こることが報告されている [145]。そこで、神経分化誘導をマウスアストロサイトと約 30 日共培養を行い、 抗neuronal class III -Tubulin (III-tubulin)抗体および抗 GFAP 抗体を用いて 免疫染色を行った(Figure 6)。GFAP 陽性細胞をカウントした結果、正常MECP2 発現株と比較して変異 MECP2発現株(RS1-61P, RS2-62P)では GFAP 陽性細胞が 有意に存在することが明らかとなった(Figure 7)。III-tubulin と GFAP が共陽性 の細胞は存在しなかったため、GFAP 陽性細胞はアストロサイト系統の細胞であ ると結論づけた。これらの結果は、MeCP2 がGfapやS100といったアストロサ イト関連遺伝子のプロモーターに結合することで、ニューロンからアストロサイ トへの変換を抑制するという報告と合致している[141, 142, 145-147]。MeCP2 が アストロサイト関連遺伝子の発現に影響を及ぼしているか否かを特定するために、 得られた神経系細胞をサンプルとして定量 RT-PCR を行った。その結果、免疫染 色 の 画 像 と 同 様 に 変 異 MECP2 を 発 現 株(RS1-61P および RS2-62P)では正 常 MECP2発現株(RS1-52M および RS2-65M)と比較してGFAPとS100の遺伝子発 現量が有意に高いことが明らかとなった(Figure 8)。また、ニューロンマーカーで あるTUBB3やMAP2 は発現が低下傾向にあったが、TUBB3はRS2 のみ、MAP2 はRS1 のみ有意差がついた(Figure 9)。
- 42 -
Figure 6. Immunostaining images of neural cells. Double labeling for GFAP (red) and III-tubulin (green) is shown along with Hoechst staining (Ho, blue) in RS1-52 M, RS1-61P, RS2-65M, and RS2-62P Rett syndrome iPSC-derived neurons. Scale bar, 200 μm.
- 43 -
Figure 7. Fraction of GFAP-positive cells relative to Hoechst-positive cells in neural cells differentiated from Rett syndrome iPSC lines. Data were analyzed by Student’s t-test and Welch’s t-test (*p<0.05). n = 5 experiments.
- 44 -
Figure 8. A qPCR-facilitated comparison of gene expression for astrocytic markers (GFAP, S100) in neural cells differentiated from Rett syndrome iPSC lines.Relative gene expression levels were normalized to ACTB expression and standardized by the RS1-52M value, which was set to 1. Data were analyzed by Student’s t-test and Welch’s t-test (*p<0.05). n = 4 experiments.
- 45 -
Figure 9. A qPCR-facilitated comparison of gene expression for neuronal markers (TUBB3, MAP2) in neural cells differentiated from Rett syndrome iPSC lines. Relative gene expression levels were normalized to ACTB expression and standardized by the RS1-52M value, which was set to 1. Data was analyzed by Student’s t-test and Welch’s t-test (*p<0.05). n = 4 experiments.
- 46 -
5) MeCP2 の欠損は神経系細胞分化においてGFAPへの結合を失う
マウスのニューロンにおいて、MeCP2 はGfapの高メチル化されたexon1 に結 合することが報告されている[141, 146]。そこで、ヒトの神経幹細胞においても同 様に GFAP の exon1 領域に MeCP2 が結合するか否か、クロマチン免疫沈降法 (ChIP)を用いて解析した。はじめに神経幹細胞をサンプルとして ChIP を行い、 GFAPプロモーター領域のSTAT3 binding site を含む領域とGFAPのexon1 を含 む領域について、定量RT-PCR 法を用いて MeCP2 結合能の検討を行った(Figure 10, indicated by blue bidirectional arrows)。STAT3 は転写活性因子として働き、 GFAP を高発現させる因子として知られている[148, 149]。両者の領域において、 正常 MECP2 発現細胞株では MeCP2 の結合が確認できたが、変異 MECP2 発現 細胞株では結合が見られなかった(Figure 11)。次に、GFAPが高発現する理由が、 GFAP プロモーター領域におけるメチル化率の差によるものか特定するために、 神経幹細胞を用いてbisulfite sequencing を行った(Figure 10, indicated by black bidirectional arrows)。その結果、正常MECP2発現株と変異MECP2発現株のど れもが高メチル化状態にあり、この領域におけるメチル化の差は存在しなかった (Figure 12)。以上の結果より、変異MECP2発現株におけるアストロサイトマー カーである GFAP の高発現は、GFAP プロモーター領域がメチル化されていても MeCP2 が変異型であるために結合できず、GFAPの発現が抑制されていないこと によって、結果的に発現が上昇していることが示唆された。言い換えると、異常 な遺伝子発現状況は、GFAPプロモーターおよびGFAP exon 1 に結合する MeCP2 量の変化に起因し得るものである。
- 47 -
Figure 10. Schematic representation of the GFAP genomic locus. ChIP/qPCR analysis was performed for the genomic regions (i.e., the STAT3 binding site and GFAP exon 1) indicated by the blue bidirectional arrows. Bisulfite sequencing was performed for the genomic region indicated by the black bidirectional arrows.
- 48 -
Figure 11. MeCP2 binding analysis and DNA methylation status of the GFAP promoter region. MeCP2 binding to the STAT3 binding site within the GFAP promoter region was quantified via ChIP/qPCR analysis by using an anti-MeCP2 antibody in neural cells differentiated from Rett syndrome iPSC lines. Data were analyzed by Student’s t-test and Welch’s t-test (*p<0.05). (関連論文 (4)より転載)
- 49 -
Figure 12. DNA methylation status of the GFAP promoter region. Methylation frequencies of (1) the CpG site within the STAT3 recognition sequence and (2) seven other CpG sites around this sequence were analyzed in iPSC-derived neurosphres via bisulfite sequencing.