218 米子医誌
J
Yonago Med Ass 47, 218-227, 1996フット上皮成長因子
(EGF)
受容体に関する研究
-RACE
法による
EGF
受容体
3
'
末端
cDNA
のクローニング-鳥取大学医学部生体情報学教室(主任臨連寺剛教授)佐藤幸夫-大前美加-一井昭五
S
t
u
d
i
e
s
on r
a
t
e
p
i
d
e
r
m
a
l
growth f
a
c
t
o
r
(EGF) r
e
c
e
p
t
o
r
:
R
a
p
i
d
a
m
p
l
i
f
i
c
a
t
i
o
n
and c
1
0
n
i
n
g
o
f
cDNA e
n
d
s
c
o
r
r
e
s
p
o
n
d
i
n
g
t
o
3
'
t
e
r
m
i
n
i
o
f
f
u
l
l
-
l
e
n
g
t
h
EGF r
e
c
e
p
t
o
r
mRNAs.
Yukio SATOH
,
Mika OOMAE
,
Shogo I
C
H
I
I
D
e
p
a
r
t
m
e
n
t
0
1
B
i
o
s
i
g
n
a
l
i
n
g
,S
c
h
o
o
l
0
1
L
約S
c
i
e
n
c
e
,F
a
c
u
l
ち1
0
1M
e
d
i
c
i
n
e
,T
o
t
t
o
r
i
U
n
i
v
e
r
s
i
ちんY
o
n
a
g
o
683,J
a
p
a
n
ABSTRACT
AH66 ascites hepatoma cel1s contain 9. 9, 7. 5 and 5. 4 kilobase (kb) epidermal growth fac -tor (EGF) receptor mRNAs which code for fulHength form of rat EGF receptor. cDNA corresponding to 3' termini of these mRNAs was cloned by rapid amplification of cDNA ends. Deduced amino acid sequence of cDNA contained transmembrane and tyrosine kinase domains and carboxy terminal autophosphorylation sites that were very similar to those of human EGF receptor. (Accepted on May 9, 1996)
Key
Words :
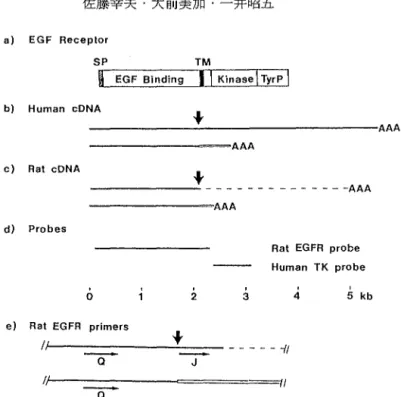
rat hepatoma cel1s, EGF receptor, EGF receptor mRNA, cDNA cloning, tyrosine kinase はじめに 上 皮 成 長 悶 子(EGF)に よ る そ の 受 容 体 (EGFR)の刺激は標的細胞の増殖を導く種々の 生理学的細胞応答を引き起こす.早期の応答とし ては,資自のチロシン残基のリン酸化,輸送系の 活発化,そして或る種の細胞においてはホスホイ ノシタイドの代謝昂進があげられ,これに続いて 数時間後に特異的な蛋白とRNAの合成が起こり, また,その後 12~18時間に DNA合成が誘起され, 細胞が成長または増殖する3,19,21)しかし, EGF がDNA合成と細抱増殖を誘起する機構は未だ十 分には解明されていない. EGFRのcDNAは,初めヒトの紹抱より単離され た11,l2,18,20) そのサイズは, 5.5 kbでその中に 3.8 kbのコード領域が含まれ,その塩基配列か ら推定されたアミノ酸配列より,①N末のシグナ ルペプチド,②細胞外のリガンド結合ドメイン,ラットEGF受察体のcDNAクロ…ニング 219 ③膜貫通ドメイン,④細胞内のチロシンキナーゼ ドメイン,⑤リン酸化されるチロシン残基を含む C末端の部分の計5つの機能領域の存在が推定さ れた.さらに, 2.7 kbのcDNAも単離された.こ の短いcDNAは細胞外リガンド結合ドメインのみ をコードしている.その蛋白産物は分泌され,生 理的なレセプターとしての作用はないと考えら れ, truncated EGFRと呼ばれている(関 1a-b). ラットにおいてもヒトと同様なtruncatedEGFR をコードする2.3kbのcDNAが単離された 13) ラ ットの全長型EGFRについても,ラットの肝臓よ り2.2kbと0.7kbのcDNAが単離された. 2.2 kbのcDNAは, EGFRの細胞外ドメインの全てと 膜貫通ドメインの一部をコードしている.それは truncated EGFRと比較すると膜貫通ドメインの 直 前 か ら 異 な っ た 配 列 を し て い る . 0.7 kbの cDNAはチロシンキナーゼドメインをコードする らしいがその配列は明らかでない.ラットの全長 型EGFRのチロシンキナーゼドメインをコードす る3'側のC末領域(上記⑤)からポリAまでの配 列に関しては全く報告されていない(国1c). 本研究は,ラットEGFRの未知C末端部分のアミ ノ酸配列を明らかにすることを目的とし,既知の リガンド結合ドメインの3'末端付近の配列をブラ イマーとしてRACE法5)を用いて,ラットの全長 型cDNAの3'末端のクローン化を試みた. 材料および方法 1 .細胞株と培養方法 AH66細胞(細胞パンク;JCRB041, AH66tc. ド ンリュウラットのヨシダ腹水癌に由来細胞9),鳥 取大学生命科学科分子生物学教室の佐藤建三教授 から供与)は, 10%新生仔牛血清(フィルトロン) を加えたダルベッコ変法イーグル培地(ニッスイ) 中で, 5%のCO2インキュベーターで
3
7
'
C
で培養 した. 2. ラット 雄の成熟ウィスターラットを用いた.肝臓は,門 脈からの氷冷生理食塩水の潅流により血液を除去 し,取り出して結合組織を除去した後,直ちに液 体窒素で凍らせ,使用するまで-80'Cで保存した. 3. RNAの分離 RNAは,酸性チオシアン酸グアニジンーフェノ-lレークロロホルム法4.14)で, AH66細胞あるいはラ ット肝から単離した.4
.
ノーザンプロット解析 RNA(20μg)は,変性 (50% ホルムアミド/2M ホルムアルデヒド ,65'C, 5分)させ, 2Mホルム アルデヒドを加えた1%アガロースゲlレで電気泳 動した10) ナイロンメンブレンフィルター(ハイ ボンド、N+,アマシャム)にRNAをプロットし固 定した15) EGFRのmRNAを検出するプロープの cDNA(関 1d,EGFR probe)は, Dr. Earp HS(University of North Carolina, Chapell 耳ill,NC, USA)より供与されたC10ne4.2 DNA の2.2kb EcoRI断片で,ラット EGFRの細胞外 ドメインを全てコードしている13) DNAプロー プは,ランダムプライマーラベリングキット(宝 酒造)を用いて1.85MBqのα_32P-dCTP(比活性 111 TBq/mmol, ICN)で標識した.フィルター は, 10%ポリエチレングリコール2)を含む42'Cの 諮液中で標識プロープとハイブリダイゼーショ ン17)の後,50'Cのo.
1XSSC/O. 1 % SDSで洗い17), 増感紙を用いて-80'CでX線フィルムに露光させ た.mRNAサイズマーカーとして0.24-9.49kb Ladder(GIBCO BRL)を用いた.フィルターは, 95'Cの0.1%SDSiこよりプロープを剥し,加のプ ロープとのハイブリダイゼーションに用いた. 5.サザンプロット解析 DNAは, 1 %アガロースゲルで電気泳動の後,ナ イロンメンブレンフィルターにプロットし固定し た15) EGFRの チ ロ シ ン キ ナ ー ゼ ド メ イ ン の cDNAを 検 出 す る プ ロ ー プ ( 図 1d,ヒト T K probe)cDNAは, Dr. Merlino GT (Nationa1 Can-cer Institute, Bethesda, Maryland, USA)より 供与されたpE7の0.72kb EcoRI断片で,これは ヒトEGFRのチロシンキナーゼドメインをコード している12) プロープの標識とハイブリダイゼー ションは,ノーザンプロット解析と向じ方法で行 なった.DNAのサイズは,.
i
.
DNAの狂indIII断片 を用いて決定した.6. RACE法
RACE (rapid amplification of cDNA ends)法5)は, 3'RACE System(GIBCO BRL)を用いた. AH66 細 胞 のR N A(1μg)か ら プ ラ イ マ -P[5'-GGCCACGCGTCGACT AGT AC (T) 17 V -3']を用 いて一本鎖cDNAを合成した.アンチセンスプラ イマーI(プライマーPに特異的)とセンスプライ マーQ(図1e,ラット EGFRI3)の塩基配列2021-2042に対応)を用いてTaKaRaEx Taq DNAポ
220 佐藤幸夫・大前美加・一井昭五 a) EGF Receplor
SP TM
H E G F Bindingll Kinase
I
TyrPI
b) Human cDNAt
AAA AAA c) Rat cDNA AH A A -A H -A H 司 AH 品 v d) ProbesRat EGFR probe Human TK probe
o 2 3 4 5主b e) Rat EGFR primers
t
- 0
一
J ー信一ー司イ/1'J 11
-a
図 EGFRcDNAの模式図. (a)全長型ヒトEGFRタンパク.SP;シグナルペプチド.EGF Bin -ding;EGF結合ドメイン.TM;膜貫通ドメイン.Kinase;チロシンキナーゼドメイン.TyrP;C末 端の自己リン酸化領域.(b)ヒトEGFRcDNA. EGFRタンパクに対応させて示す.黒線;全長型 EGFRのcDNA.矢印;全長型EGFRとtruncatedEGFRの毘列の分岐点.白線;全長型EGFRと異 なるtruncatedEGFR配列 AAA;ポリAテール. (c)ラットEGFRcDNA.点線;全長型EGFRの cDNAの未知の配列(長さも未知).白線;全長型EGFRと異なるtruncatedEGFR配列(ヒト配列 と異なる). (d)本研究に用いたcDNAプロープ.TK;チロシンキナーゼ. (e)ラットEGFR cDNAの増幅に用いたセンスプライマーQとJの位置.図lcのー音
s
を拡大して示す . Qは全長型 EGFRとtruncatedEGFRに共通.Jは全長型EGFRに特異的.リメレースにより第一段階の増幅を行なった.ブ ライマー
-
1
と内側のプライマ-J(図le,ラット EGFR塩基配列2076-20951こ対応)を用いて第一 段措の増幅を行なった. 7. cDNAクローニングとコロニーハイブリダイ ゼーションによるスクリーニング 増幡産物は,プラスミドベクタ-pBluescriptII SK+
(Stratagene)に連結し,大腸菌JMI09の形 質 転 換7)に用いた.形質転換体のクローンは, LB/ Amp/1PTG/X-Galプレート15)上の白いコロ ニーとして同定した.コロニーの一部をナイロン メンブレンフィルターに移し,コロニーのDNA を変性しフィルターに臨定した.フィルターは, サザンプロット解析と同じ手法で, 32p標識TK probeとのハイブリダイゼーションを行なった. EGFR cDNAを含むクローンを同定し,それら のプラスミドDNAを調製15)した.8
.
塩基配列決定 クローン化したEGFRcDNAを含む約2.4kbの BstX1-Kpn1ないしSalI断片をHaeillないしAlu1 消化により断片化し,プラスミドベクターに連結して大腸菌を形質転換してcDNA断片を含むク ローンを単離し,各クローンのプラスミドDNA を調製した1)α_32P-dCTP用BcaBESTジデ、オキ シシークエンシンク鳴キット(宝酒造)またはAL-Fred DNAシークエンサー用Cy5オートリード シークエンシングキット(ファルマシア)を用い て,各クローンのプラスミドDNAの塩基配列を 決定した.これらの配列をつなぎ合わせて元の cDNAの塩基毘列を決定した. 結 果 1. AH66細胞の4種のEGFRmRNA
ラット
EGF
受容体のc
D
NA
クローニングa
kb
9.9
~7.5
~5.4
~2.6
.~c
1
.
2
「.
.
.
.
.
2.
4
kb
2
2
1
b
1
1
4
4
4
3 4
.
.
.
.
.
5
k
b
.
.
.
.
.
2
.
4
図2
A
H
6
6
細胞からのEGFR
m
RN
A
とE
GF
R
c
DN
A
の増幅.(
a
)
E
G
FR mRN
A
の発現.AH
6
6
細胞のRNA
(
2
0
μ
g
)
をEGFR
c
DN
A
プロープを用いてノーザンプロット解析した.mR
NA
のサイズ(
k
b
)
を左に示す.(
b
)
a
のフィルターよりプロープを剥し,1.4
k
b
のDNA
断片(図4
c
に後述) をプロープとして検出した.(
c
)
増幅したEGFR
c
DN
A
の検出.PCR
増幅産物をTK
プロープを 用いてササ'ンプロット解析した.レーンl
と2
;
第一段階R
A
C
E
の産物.レーン3
と4
;
第二段階RACE
の産物.サイズ(
k
b
)
を右に示す. ンプロット解析により検討した(図2
a
)
.
ラットE
G
FR
の細胞外ドメインをコードするc
DN
A
プ ロープを用いて4
つのバンドが検出された.その サイズは9
.
9
,7
.
5
,5
.
4
,2
.
6
k
b
であった.これ らのサイズは,ラット肝のE
GF
RmRN
A
のサイ ズ6,8,13,16)と一致していた.従って,AH
6
6
細胞は, ラット肝と同様な全長型E
G
FR
のmRNA
(
9
.
9
,7
.
5
,5
.
4
k
b
)
とt
r
u
n
c
a
t
e
dEGFR
の
mRN
A
(
2
.
6
k
b
)
を発現しており,cDNA
クローニングの材料 とした.2
.
R
A
CE
法によるmRN
A
のポリA
末端と内部のE
GF
R
配列を用いたPCR
増幅 検出された2
.
6
k
b
のmRNA
は,t
r
u
n
c
a
t
e
d
EGFR
のmRNA
である(図l
c
)
.
全長型EGFR
の3
'
側の 未知の部分のc
D
NA
をクローニングする目的で3
'
R
A
CE
法を行なった(方法参照).A
H
6
6
細胞のRN
A
より一本鎖c
DN
A
を合成し,二段階のP
C
R
増幅を行なった.増幅産物中にE
G
FR
c
D
NA
を 検出するためにサザンプロット解析を行なった. 第一段階と第二段階のPCR
増幅産物中にそれぞ れ約2~3k
b
(
図2
c
, レーント2
)
と約5
k
b
付近と2
.
4
k
b
のサイズをもっバンド(図2
c
,レーン3
-
4
)
が検出された.従って,目的とする全長型EGFR
のチロシンキナーゼドメインをコードするc
DN
A
が増幅されたものと推定された.3
.
c
DN
A
のクローン化 検出したc
DN
A
をプラスミドベクターに連結し, 大腸菌を形質転換した.2
0
3
個の形質転換大腸菌 コロニーより目的とするEGFR
c
DNA
を検出す るために,コロニーハイフリダイゼーションを行222 佐藤幸夫・大前美加 一井昭五
図3 EGFR cDNA クローンのスクリーニング.RACE産物をプラスミドに連結し,大腸菌を形質転換 し,形質転換した菌のコロニーをフィルター上に並べ,TKプロープを用いてコロニーハイブリ ダイゼーションを行なった.10コロニーがハイブリダイゼーション陽性を示した. なった.陽性を示す10個のコロニーを同定した(図 3).これらのクローン化されたcDNAを確認し, そのサイズを知るために,陽性クローンのプラス ミドを調製し,制限酵素 CSalI,SacI)で二重消 化し電気泳動した(図4a).大部分のクローンに ついて,1.4 kbと1.0kbの二本のバンドが検出 された.ただし,クローン7および90は, 1.4 kb より少し短いサイズのバンドを示した.またク ローン35は,おそらく不完全消化によると思われ る2.4kbのうすいバンドも見られた.以上のこ とから,このクローン化されたcDNAは2.4kbの 長さを有し,SalIないしSacIによって切られて 1.4kbと1.0kbの断片を生じるものと考えられ た.それを確認するためにTKプロープを用いて サザンプロット解析を行なった(図的).1.
0
kb と2.4kbのバンドにはチロシンキナーゼの配列 が含まれることが確認された.1.0 kbバンドは 全てのクローンにおいて観察された.2.4 kbの バンドは大部分のクローンにおいて観察された しかし, クローン7と90においては2.4kbより少 し短いサイズのバンドが観察され,クローン17に おいては1.0kb以外のバンドは全く観察されな かった.1.4 kbバンド(図4a)には,いずれのク ローンでも,チロシンキナーゼの配列が全く検出 されなかった(図4b).1.4 kbバンドは2.4kbパ ンドの断片であると考えられたので,確認のため, 1.4 kb断片をプロープと したサザンプロット解 析を行なった(図4c). その結果,大部分のクロー ンについて, 1.4 kbと2.4kbのバンドが,プロー プとした1.4kb DNAの配列を有することが確認 された.クローン?と90では, 1.4 kbと2.4kbよ り少し短いサイズのバンドが検出された.クロー ン17では,1.4 kbや2.4kbのバンドは観察され ず,かわりに0.4kbと1.0kbの強いバンドと2.0 kbの弱いバンドが観察された.これらの結果よ り,クローン化したcDNAは,大部分が約2.4kb のサイズであり,SalI/SacIM
化により1.0kbと 1.4
kbの断片を生じるものと結論された.ただ し,クローン?と90では,2.4 kbより少し短いサ イズであり ,1.0 kbと1.4kbより少し短い消化 産物を生じた.クローン17では,2.4 kbのサイ ズであるが,消化により二本の1.0kb断片と一 本の0.4kb断片を生じた.いずれのクローンも 一本の1.0 kb断片はチロシンキナーゼドメイン に対応していた. 4. 1.4kb断片をプロープに用いたノーザン解 析による全長型EGFRmRNAの検出 我々がクローン化した, 2.4 kbのcDNAが, 本当 にラッ トEGFRのmRNAより由来したものであ ることを確認するために,図2aのフィルターを,223 1.4kb 1.0 ラットEGF受容体の
c
DNA
クローニングb
789094 ...:.... 2.4kb・
・
・
・
・
・
‘
1.0 2430313539ー
・
相
同
.
一
‘
‘
6 7 13 17.
.
.
.
.
.
・
.
,
‘
ー
‘
c
l‘
2.4kb.
.
.
.
1
.4 789094.
.
.
.
‘
2430313539 4‘
6 7 13 17 陽性クローンのプラスミドDNA
の解析.(
a
)
制限消化後の電気泳動像.プラスミドDNA
を SalI/SacI消化後, 10/0アガロースゲル上で電気泳動し,エチジウムブロミドで染色した. M;,l/HindIlI .レーンの上の数字;クローンの名.クローン6と24;陰性クローン.DNA
のサイズ (kb)は右に示す.2.9kbバンドはベクターである. (b)チロシンキナーゼ配列の検出. aのサザ ンプロット解析をした.TK
プロープを用いた.(c) 1.4
kbのDNA断片をプロープとするサザン プロット.aのクローン30の1.4 kbDNA
断片を回収してプロープとし, bのフィルターを解析し た. 図4
同 T m m 叩 , 4 ~\OO ←~今~→←」比~ 1-企L参 HZη コ~ 」丘→ トー且L→ トA止→ 」立→ ←」盟→ ト」止→~込→ト...M....→ ~←--.ð.L→←
Aヱ
ー
→
←....dL....←」盆---< ~→ ←」旦→←~ ←一主lー--j ~ぷ旦→ ←ー出吋 ←~←i止→ , q4.R.I ←...M!.→ ~oo 3000塩基配列決定の概略.上の線;ラット EGFR
cDNA
(
決定した 2096-4733の2638bp).H;HaeIlI/SalI断片.
A
;
A
l
u
I/SalI断片. K;KpnI/HaeIlI断片.数字;断片のクローンの名.矢印;配列決定した方向と長さ. 図5
2
2
4
佐藤幸夫・大前美加・一井昭五 4〆 「今ユ.96'"" CTGCCACCTCTGCCAτGCAAACTGTACCTAτGGATGTGCTGGGCCAGGCCTTAAAGGATGTCAACAACCAGAAGG~CCAAAGATCCCATCCATCGCCACT 2100 C I I L C H A N C T Y G C A G P G L K G C Q Q P E G P K P 5 A T ..-2104'" GGG以TTGTGGGTGGCCTCCTCTTCATAGTAGTGGTGGCCCTTGGGATCGGCCTCTTCATGCGAAGGCGTCACATTGTCCGAAAACGTACACTACGGCGCC G V G G L L F V V V A L G G L F M R R R H V R K R T L R R L TGCTTCAAGAGAGAGAGCTCGTGGAACCTCTCACACCCAGCGGAGAAGCTCCGAACCAAGCCCACTTGAGGATATTAAAGGAAACAGAATTCAAAAAGAT L Q E R E L V E P L T P 5 G E A P N Q A H L R L K E T E F K K CAAAGTTCTGGGTTCAGGAGCATTTGGCACAGTGTATAAGGGTCTCTGGATCCCAGAAGGCGAGAAAGTGAAAATCCCTGTGGCCATCAAGGAGTTAAGA K V L G 5 G A F G T V Y K G L W P E G E K V K P V A K E L R GAAGCCACATCTCCCAAAGCCAACAAGGAAATCCTTGATGAAGCCTACGTGATGGCCAGTGTGGACAACCCTCATGTATGCCGCCTCCTGGGCATCTGTC E A T 5 P K A N K E L D E A Y V M A 5 V D N P H V C R L L G C L TGACCTCCACTGTCCAGCTCATTACACAACTCATGCCCTATGGTTGCCTCCTGGACTATGTCCGAGAACATAAGGACAACATTGGCTCCCAGTACCTACT T 5 T V Q L T Q L M P Y G C L L D Y V R E H K D N G 5 Q Y L L CAACTGGTGTGTGCAGATTGCAAAGGGCATGAACTACCTGGAAGACCGGCGTTTGGTACACCGTGACTTGGCAGCCAGGAATGTACTGGTAAAGACACCA N W C V Q A K G M N Y L E D R R L V H R D L A A R N V L V K T P CAGCATGTCAAGATCACAGATTTTGGACTGGCCAAACTGCTTGGTGCTGAGGAGAAAGAATACCATGCAGAGGGGGGCAAAGTGCCTATCAAGTGGATGG Q I I V K T D F G L A K L L G A E E K E Y H A E G G K V P K W M A CTTTGGAATCAATTTTACACCGAATTTATACACACCAAAGCGACGτCTGGAGCTATGGAGTCACCGTGTGGGAACTGATGACCTTTGGGTCCAAGCCTTA L E 5 L H R Y T H Q 5 D V W 5 Y G V T V W E L M T F G 5 K P Y TGATGGGATCCCTGCAAGTGAGATCTCATCCATCCTAGAGAAAGGAGAGCGCCTTCCACAGCCACCTATCTGCACCATCGACGTCTACATGATCATGGTC D G P A 5 E 5 5 L E K G E R L P Q P P C T D V Y M M V AAGTGCTGGATGATAGATGCTGATAGCCGCCCAAAGTTCCGAGAGTTGATTCTCGAATTCTCCAAAATGGCCAGAGACCCACAGCGCTACCTTGTTATCC K C W M D A D 5 R P K F R E L L E F 5 K M A R D P Q R Y L V Q AGGGGGATGAAAGGATGCATTTGCCGAGCCCTACAGACTCCAACTTTTACCGAGCCCTGATGGAGGAGGAGGACATGGAAGACGTAGTTGATGCTGATGA G 0 E R 同 H L P 5 P T 0 5 N F Y R A L M E E E 0 M E D V V 0 A D E ATACCTCATCCCACAGCAAGGCTTCTTCAACAGCCCATCCACGTCACGGACTCCACTCTTGAGCTCTCTGAGτGCAAATAGCAACAGTTCCACTGTGGCT Y L P Q Q G F F N 5 P 5 T 5 R T P L L 5 5 L 5 A N 5 N 5 5 T V A TGCATTAATAGAAATGGGAGCTGCCGTGTCAAAGAAGACGCCTTCTTGCAACGGTATAGCTCCGATCCCACCAGCGTCCTGACAGAGGACAACATAGATG C N R N G 5 C R V K E 0 A F L Q R Y 5 5 0 P T 5 V L T E D N D D ACACATTCCCTCCCGTGCCTGAATATATAAACCAATCTGTTCCCAAGAGGCCGGCTGGCTCTGTGCAGAACCCAGTCTATCACAATCAGCCCCTGCATCC T F P P V P E Y N Q 5 V P K R P A G 5 V Q N P V Y H N Q P L H P AGCTCCTGGAAGAGACCTGCATTATCAAAATCCCCATAGCAATGCGGTGAGCAACCCTGAGTATCTCAACACTGCCCAGCCGACCTGCCTCAGTAGTGGG A P G R 0 L H Y Q N P H 5 N A V 5 N P E Y L N T A Q P T C L 5 5 G rTTGACAGCTCTGCCCTCTGGATCCAGAAAGSCAGCCACCAAATGAGCCTGGACAACCCTGACTACCAGCAGGACTTCTTTCCCAAAGAAGCCAAGCCGA F D s s A L W I Q K G s H Q H s L D N P D Y Q G D F F P K E A K P H ATGGCATCTTTAAGGGCCCCACAGCTGAAAATGCAGAGTACCTGCGGGTGGCACCGCCAAGCAGTGAGTTTATTGGAGCATGACATTGAAGAGGCATTGT G F K G P T A E N A E Y L R V A P P 5 5 E F G A ACCAGCTACAAACCGGACTTTCCAGAAGCCCAGGACCAAGCCATGGCAGCACCTCTGCTCCTGACAGCCATGTCCACATTGTGTCAAATGTCAAACCCTC AGACTGGCTTTAAAGCATAACTCTGACGGGCTTTGTCACTGAGCCAAGAAGTGGGCCCTCCCCTGATGCTCTTTGGGAAGTTGAAGGTATATCTATTGGT CTTCGAACTGTGAAGATTCCACTGAAAGGTATCCATCGAGAACATTGTCCTTTTGGAACAGAAGGTTGCCGTCATGGTGAGGTACATAGGGGGAAAAAAA ACAGACCTATGGCGCTTGCTGCATAGGGAACTCTGGGATTCTTGTCTTTATTGATTTGATTCATGCACTCTTCCATAAAGGAAGAAGCTTGCCCGTAGGG TGTATTACACAGAGTTGCCTGGAGCCAACTGACCAGACAGTTGGTTCTAGCAGCTCTGCATCAAGACACTTCCATGGCAAGATAACTACATGCACAAGAA GTCCTGGATGTGCTCAGCAGGCCACACTTGTACAGCATTAAACCATGGCAGGTACAATTGGATAAGCCACTTTGTTACTTACTGGGACTGGGAGAAGAAT GACTGGGTAGAAT77TCCCTCAGACTTACTTTTTTTGTATAAAT^TGTCCCTGATACT7AACACACGごTAG7TTACCAGTGTTTTCTGACTATTAGACTA C己TTTTATGTT7TCTGTTTCATTGTTTTGAGTTTTAATATGC7TTCCTG7TTTCATTTCATGAAGTAAACAAACAAACAAACAAGCAAAAAAAAAAAAAC AG7ATTATTATCAAAGAACAACCATGATTCAAACCCATTCGAACCATCAGTATTGTAACCCAAAGCCTTTATCTAAGCTGGAGTAACCATGCAAAAATTC CTAGAAGAAATTTAACCCAAAAAAAAAAAAAAA 220U 2300 2400 2500 2600 2700 2日00 2!lOO 3000 3100 : 1200 33()1J 34UO 350U 36()1J 3700 3A()0 39UO 4000 4100 4200 4:l00 44011 4500 4600 4700 4733 図6
全長型ラットEGFRcDNA
の塩基配列と推定アミノ酸配列.2
0
0
1
-
4
7
3
3
の塩基配列を示す.2
1
0
3
までは既知の配列13). 今回明らかにしたのは2
0
9
6
-
4
7
3
3
.
塩基配列の下に推定されるアミノ酸配 列を示す.下向きの矢印;
t
r
u
n
c
a
t
e
dEGFR
と異なる配列が始まる点.ラット EGF受容体の cDNA~ノローニング 225 J , r6~1 "",
R
.
a
t
601・GENNTLVWKFADANNVCI1LCHANCTYGCAGPGLKGCQQPEG'I'KJ PS 1 A TGド
VG己a.LF1 VV 司<>ド〉ド******.***..*
*
*
*
*
*
.
*
*
*
*
*
*
.
*
*
*
*
.
*
*
...*本*本******.**.**...*H
u
m
a
n
601" GENNTLVWKYADAGHVCHLCHPNCTYGCTGPGLEGC-PTNGPKIPSIATGMVGALLししLV 661 ' 66 ()"' V八LGIGLFMRRRliIVRKRTLRRLLQERELVEPLTPSGEAPNQAHLRULKETEFKKIKVしG ****.**本**本**水*京*****本**市本来************本*** **'14****水*******埼〈 V ^LGTGLFMRRRHIVRKHTLRRLLQERELVEPLTPSGEAPNQALLRIILKETEFKK 1 KVLG 721 ' SGAFGTVYKGLWIPEGEKVKIPVAIKELREATSPKANKEILDEAYVMASVDNPHVCRLLG *****同ド**持<*****本〉ド*****本*~ド〉ド***************************>ドホ*>ド**** 720・・I
SGAFGTVYKGLWIPEGEKVKIPVAIKELREATSPKANKEILDEAYVMASVDトIIコ日VCRLLG 781 ' I 1 CLTSTVQLI TQLMPYGCLLDYVREHKDN 1 GSQYLLNWCVQ 1 AKGMNYLEDRRしVHRDLA **************本.**********>ド****>ド埼<*****>ド**埼<********,ド******埼<** 780"1 ICLTSTVQLITQLMPFGCLLDYVREHKDNIGSQYLLNWCVQIAKGMNYLEDRRLVHRDLA 841'IARNVLVKTPQHVKITDFGLAKLLGAEEKEYHAEGGKVPIKWMALESILHRIYTHQSDVWS 本本**>ド*******>ド***********神:******本*******水****本*************** 840"IARNVLVKTPQHVKITDFGLAKLLGAEEKEYHAEGGKVPIKWMALESILHRIYTHQSDVWS 901・
IYGVTVWEしMTFGSKPYDGIPASEISSILEKGERLPQPPICTIDVYMIMVKCWMIDADSRP *>宮本********本**>ド**>ド〉ドホ*****************>ド*水*本***>ド**>ド*******持<**>ド 900" 1 YGVTVWELMTFGSKPYDGIPASEISSILEKGERLPQPPICTIDVYMIMV KCWMIDADSRP 961 ' 960" KFRELILEFSKMARDPQRYLVIQGDERMHLPSPTDSNFYR八LMEEEDMEDVVDADEYLIP*
*
*
*
*
*
.
**********ホ****埼1*
*
*
*
司〈本***********斗<**.時<***.********司<** }(FRELIIEFSKMARDPQRYLVIÇ~GDERMHLPSPTDSNFYRALMDEEDMDDVVDADEYLIP 1021 ' QQGFFNSPSTSRTPLLSSLSANSNSSTVACINHI¥IG--SCRVKEDAFLQRYSSDPTSVLTE *****.****米3事*********.**.******.米***
*
.
.
*
*
*
.
*
*
*
*
*
*
*
*
*
*
.
.
**>ド 1020" QQGFFSSPSTSRTPLLSSLSATSl¥INSTVACIDRNGLQSCPIKEDSFLQRYSSDPTGALTE 1079' DNIDDTFPPVPEYINQSVPKRPAGSVQNPVYHNQPLHPAPGRDLHYQNPHSNAVSNPEYL 米.****本 **************>ド*******>ド***本*.*米*.*
*
*
*
*
.
***.本司<.*
*
*
*
*
1080・・ DSIDDTFLPVPEYINQSVPKRPAGSVQNPVYHNQPLNPAPSRDPHYQDPHSTAVGNPEYL 1139' NTAQPTCLSSGFDSSALWIQKGSHQMSLDNPDYQQDFFPKEAKPNGIFKGPTAENAEYLR 本*.****..*
.
*
*
*
.
*
*
**>ド***.*本***持<>ド****************埼< >ド******** 1140"且
エ
VQPTCVNSTFDSPAHWAQKGSHQISLDNPDYQQDFFPKEAKPNGIFKGSTAENAEYLR 1199' V八PPSSEFIGA*
*
*
.
*
*
*
*
*
*
*
1200" VAPQSSEFIGA 図 7 ラットとヒトの全長型EGFRのアミノ酸配列の比較.601-1209のアミノ酸配列を示す.上段;ラ ットEGFR.下段;ヒトEGFR. *;一致.一;ギャップ.箱区画;チロシンキナーゼドメイン.アン ダーライン;膜貫通ドメインと 6個のチロシンリン酸化部位. 1.4 kb断片をプロープとしてノーザンプロット 解析を行なった(図2b).その結果, 9.9, 7.5, 5.4 kbの3本のバンドが検出され,その位置は図 2aの3本の全長型EGFRmRNAの位置と完全に一 致した.しかし, 2.6 kbのtruncated EGFR mRNAは全く検出されなかった.従って,この 1.4 kbのプローブは全長型EGFRmRNAのみを 特異的に検出するプロープであった.以上の結果 より我々はラットの全長型EGFRのcDNAの3
'
側 の末端部分を単離できたと考えた.AH66細胞か ら単離したcDNAは,ラットの組織の豆GFR mRNAも検出できるかどうかを確認するため, ラット肝からのRNAサンプルをノーザンプロッ ト解析した.その結果, 1. 4 kbプローブは, A H 66細胞のRNAでもラット肝のRNAでも同様に, 9.9 kb, 7 kb, 5 kbの3本のmRNAのみを検出し た(データ示さない).従って単離したcDNA断 片は, AH66細胞とラットの肝臓に共通するラッ トのEGFRのcDNA配列を含むと考えられた. 5. クローン化したEGFRcDNAの塩基配列と226 佐藤幸夫・大前美加・一井昭五 推定アミノ離配列 ク口ーン化した2.4kbのEGFRcDNAを,制限 消化により断片化し,断片をプラスミドベクター によりクローン化した.各クローンのプラスミド DNAの塩基配列を決定し,これらの配列をつな ぎ合わせてEGFRcDNAクローンの塩基配列を 決定した(図5).今回明らかにした配列 (2096 -4733)は,既知の配列(1-2103)13)にin-frameで 接続する2630bp長の新しい配列を含んでいた(図 6) .それは559残基 (65l!-1209A)のアミノ酸配 列をコードしており,膜貫通ドメインの一部,チ ロシンキナーゼドメイン,および6個のチロシン リン酸化部位を含むC末領域であった(図7).こ の自己列を全長型ヒトEGFRのそれ18)と比較する と,ラットEGFRに2アミノ酸(1056-1057)のギ ャップが見いだされ, 94. 1 %のアミノ酸配列が一 致した.この配列をコードする塩基配列 (2104 -3783)は,ヒトと比べ,上記2アミノ酸ギャップ に対応する6bpのギャップが見いだされ, 86.0 %の一致を示した.…方 3'非翻訳領域の塩基配 列 (3784-4733)は,ヒトと比べ59.8%の一致し か示さなかった.合わせて2104-4733の配列はヒ トと比べ76.3%が一致していた.これらの結果よ り,我々はラットEGFRの細胞内のドメインの全 てをコードするcDNAクローンを単離出来たと考 える. 考 察 我々の研究の目的は,ラットのEGFRの未知のC 末端のアミノ酸配列を明らかにすることであっ た.そのアプローチとして, 3'RACE法を用い, 目的のcDNAを増幅した.クローン化したcDNA は, 2.4 kbのサイズであり,制限酵素消化によ って1.0kbと1.4kbの断片に分けられた.1. 0 kbの断片は,チロシンキナーセ
1
こ特異的なcDNA プローブとハイブリダイズしたので,チロシンキ ナーゼ特異的cDNA配列を含むものと患われる. 1.4 kb断片は,全長型EGFRの3つのサイズの mRNA(10, 7.5, 5.4 kb)を検出したのでこれら の全長型EGFRのmRNAに共通な3'末端の配列を 含むものと考えられる.今回明らかにしたcDNA の配列は2630bpであり,推定アミノ酸配列は全 長型ヒトEGFRの膜貫通ドメインよりC末端部分 までの配列とよく似ている.ラットEGFRの既知 の部分的アミノ酸配列13)と合わせると,ラット EGFRの全長は1209アミノ酸残基となる.この全 長型EGFRをコードするcDNAは長さが合計4.7 kbとなり,サイズ的に見て5.4kbのmRNAにほ ぼ完全に対応する可能性が考えられる.一方, 9. 9 kbと7.5kbのmRNAは,この配列も含むもの の他に未知の配列も含むものと思われる. 結 圭五 回ロ ラット腹水肝癌由来AH66細越より全長型EGF受 容体のmRNAに対応するcDNAの3'末端の約2.6 kbの部分をRACE法により単離した.その塩基 配列は,ヒトEGF受容体の膜貫通ドメイン,チ ロシンキナーゼドメイン, C末白日リン酸化領域 と類似したアミノ酸配列をコードしていた. 本論文の概要は,平成7年度生命科学科卒業研究とし て発表した.西連寺剛教授のご校関に感謝いたします. 文 献 1) Akiyama H, Kawai S(1995) A strategy for one week sequencing. Protein, Nucleic acid and Enzyme 40, 173-178. 2) Amasino R M(1986) Acce1eration of nucleic acid hybridization rate by polyethylene glycol.Anal Biochem 152, 304-307. 3) Carpenter G, Cohen S (1990) Epidermal growth factor. J Biol Chem 265,7709-7712. 4) Chomczynski P, Sacchi N (1987)Single -step method of RNA isolation by acid guanidinium thiocyanate-phenol-chloroform extraction. Anal Biochem 162,
156-159. 5) Frohman M A, Dush M K, Martin GR(1988)Rapid production of fulHength cDNAs from rare transcripts: Amplification using a sigle gene-specific oligonucleotide primer. Proc Natl Acad Sci USA 85, 8998-9002. 6)
I
c
hii S, Satoh Y, Hoshikawa y, Yoshida A (1991)Re-investigation of ontogenesis of epidermal growth factor receptor mRNA in the liver of rats: Quantitative evaluation of Northern blot analysis. Endocrinol Japon 38, 511-516.7) Inoue H,Nojima H,Okayama H(1990) High efficiency transformation of
Es-ラットEGF受容体のcDNAクローニング 227 cherichia co1i with p1asmids. Gene 96, 23 -28. 8) Johnson AC, Garfie1d SH, Mer1ino GT, Pastan I (1988) Expression of epiderma1 growth factor receptor proto-oncogene mRNA in regenerating rat 1iver. Biochem Biophys Res Commun 150, 412-418. 9) Katuta H, Takaoka T, Yasumoto S(1973) Toxic metabo1ites re1eased from hepatoma cells in cu1ture. 1.effects of metabo1ites of hepatomas on various cells. J Nat1 Cancer Inst 5,1 1841-1844.
10) Lehrach H, Diamond D, Wozney J1ι Boedtker H (1977) RNA mo1ecu1ar weight determinations by ge1 e1ectrophoresis under denaturing conditions, a critica1 reex -amination. Biochemistry 16, 4743-4751. 11)Lin CR, Chen WS, Kruiger W, Sto1arsky
LS, Weber W, Evans RM, Verma IM, Gill GN, Rosenfe1d M G(1984) Expression doning of human EGF receptor comp1emen-tary DNA: gene amplification and three re1ated messenger RNA products in A431 cells. Science 224, 843-848.
12) Merlino GT, Ishii S, Whang-Peng J, Knutsen T, Xu YH, C1ark AJL, Stratton RH, Wilson RK, Ma DP, Roe BA, Hunts JN, Shimizu N, Pastan I(1985) Structure and 1ocalization of genes encoding aberrant and norma1 epiderma1 growth factor receptor RNAs from A431 human carcinoma cells. Mo1 Cell Bio1 5, 1722-1734.
13) Petch LA, Harris J, Raymond V W, B1asband A, Lee DC, Earp HS(1990) A truncated, secreted form of the epiderma1 growth factor receptor is encoded by an a1 -ternative1y sp1iced transcript in norma1 rat tissue. Mo1 Cell Bio1 10
,
2973-2982. 14) Puissant C, Houdebine LM(1990) An im -provement of the sing1e-step method of RNA iso1ation by acid guanidinium thioc・ yana te-pheno1-ch1oroform extraction. Biotechniques 8, 148-149. 15) Sambrook J, Fritsch EF, Maniatis T(1989) Mo1ecu1ar cloning: a 1aboratory manual. Co1d Spring Harbor Laboratory Press, New York16) Satoh Y, Hoshikawa y, Ichii S(1990) Epiderma1 growth factor mRNA in livers from newborn, adu1t and partially hepatoec -tomized rats. Endocrino1 Japon 37, 430 -435.
17)Thomas PS (1980) Hybridization of dena -tured RNA and smal1 DNA fragments trans司
ferred to nitrocellu1ose. Proc Nat1 Acad Sci USA 77, 5201-5205.
18) Ul1rich A, Coussens L, Hayflick JS, Dull TJ, Gray A, Tam A W, Lee J, Yarden Y,
L
i
bermann TA, Sch1essinger J, Down-ward J, Mayes EL V, Whitt1e N, Water幽fie1d M D, Seeburg PH (1984) Human epiderma1 growth factor receptor cDNA se -quence and aberrant expression of the am-p1ified gene in A431 epidermoid carcinoma cells. Nature 309, 418-425. 19) Ullrich A, Sch1essinger ](1990) Signa1 transduction by receptors with tyrosine kinase activity. Ce1l206, 203-212. 20) Xu YH, Ishii S, C1ark AJL, Sul1ivan M, Wilson RK, Ma DP, Roe BA, Merlino GT, Pastan I(1984) Human epiderma1 growth factor receptor cDNA is homo1ogous to a variety of RNAs overproduced in A431 carcinoma cells. Nature 309, 806-810. 21)Yarden y, Ullrich A(1988) Growth factor
receptor tyrosine kinases. Ann Rev Biochem 57, 443-478.