Nagoya City University Academic Repository
学 位 の 種 類 博士 (薬学) 報 告 番 号 甲第1702号 学 位 記 番 号 第348号 氏 名 小野里 太智 授 与 年 月 日 平成 31 年 3 月 25 日 学位論文の題名 人工多能性幹細胞由来腸管オルガノイドの新規分化誘導法の開発と炎症性 腸疾患モデル系の構築 論文審査担当者 主査: 頭金 正博 副査: 松永 民秀, 湯浅 博昭, 平嶋 尚英
名古屋市立大学学位論文 人工多能性幹細胞由来腸管オルガノイドの新規分化誘導法の開発と 炎症性腸疾患モデル系の構築 平成30 年度 (西暦 2019 年 3 月) 所属 臨床薬学分野 氏名 小野里 太智
一.本論文は2019 年 3 月名古屋市立大学大学院薬学研究科において審査され たものである。 主査 頭金 正博 教授 副査 平嶋 尚英 教授 湯浅 博昭 教授 松永 民秀 教授 二.本論文は、学術情報誌に掲載された次の報文を基礎とするものである。 【基礎となる報文】
1. D. Onozato, M. Yamashita, A. Nakanishi, T. Akagawa, Y. Kida, I. Ogawa, T. Hashita, T. Iwao, T. Matsunaga.
Generation of intestinal organoids suitable for pharmacokinetic studies from human induced pluripotent stem cells
Drug Metab. Dispos., 46, 1572-1580 (2018).
2. D. Onozato, M. Yamashita, R. Fukuyama. T. Akagawa, Y. Kida, A. Koeda, T. Hashita, T. Iwao, T. Matsunaga.
Efficient generation of cynomolgus monkey induced pluripotent stem cell-derived intestinal organoids with pharmacokinetic functions
Stem Cells Dev., 27, 1033-1045 (2018).
3. D. Onozato, T. Akagawa, Y. Kida, I. Ogawa, T. Hashita, T. Iwao, T. Matsunaga. Application to inflammatory bowel disease model system of human induced pluripotent stem cell-derived intestinal organoids
In preparation
三.本論文の基礎となる研究は、松永 民秀 教授の指導のもとに名古屋市立大
略語一覧 2-ME 2-メルカプトエタノール APO アポリポタンパク質 A-83-01 3-(6-methyl-2-pyridinyl)-N-phenyl-4-(4-quinolinyl)-1H-pyrazole-1-carbothioamide ABC ATP 結合カセット BCRP 乳がん抵抗性タンパク質 BMP 骨形成タンパク質 CDX 尾側タイプホメオボックス CHIR99021 6-[[2-[[4-(2,4-dichlorophenyl)-5-(5-methyl-1H-imidazol-2-yl)-2-pyrimidinyl]amino]ethyl]amino]-3-pyridinecarbonitrile CYP シトクロムP450 DAPI 4',6-diamidino-2-phenylindole
DAPT N-[(3,5-difluorophenyl)
acetyl]-L-alanyl-2-phenyl-1,1-dimethylethyl ester-glycine
DMEM Dulbecco's modified Eagle medium
DMEM/F12 DMEM/ハム F-12 培地
DMSO ジメチルスルホキシド
EGF Epidermal growth factor
EMT 上皮間葉転換
ES 細胞 胚性幹細胞
FBS ウシ胎仔血清
FGF Fibroblast growth factor
IBD 炎症性腸疾患 IFX Infliximab IL インターロイキン iPS 細胞 人工多能性幹細胞 KSR KnockOut™血清代替 L-Glu L-グルタミン溶液 LGR ロイシンリッチ反復領域を有する G−タンパク質共役型 レセプター Matrigel Matrigel™基底膜マトリックス (成長因子を減じたもの) MEF マウス胎仔線維芽細胞 NEAA 最小必須培地用非必須アミノ酸 MRP 多剤耐性関連タンパク質 MUC ムチン OATP 有機アニオン輸送ポリペプチド OLFM Olfactomedin PBS リン酸緩衝食塩水 PCR ポリメラーゼ連鎖反応 PD98059 2-(2-amino-3-methoxyphenyl)4H-1-benzopyran-4-on P-gp P-糖タンパク質 PXR プレグナンX 受容体 RIF Rifampicin RPMI ロズウェルパーク記念研究所 RT 逆転写
SB431542 4-[4-(1,3-benzodioxol-5-yl)-5-(2-pyridinyl)-1H-imidazol-2-yl]-benzamide SLC 溶質輸送体 SLC15A1/PEPT1 溶質輸送体ファミリー15 メンバー1/ペプチドトランポ ーター TGF-β 形質転換増殖因子-β TNF-α 腫瘍壊死因子-α LC/MS/MS 超高速液体クロマトグラフ–タンデム型質量分析計 VDR ビタミンD 受容体. Y-27632 (+)-(R)-trans-4-(1-aminoethyl)-N-(4-pyridyl) cyclohexanecarboxamide dihydrochloride
i 目次 第一章 序論 ... 1 第二章 多能性幹細胞由来腸管オルガノイドの新規分化誘導法の開発 ... 3 2.1 緒言 ... 3 2.2 実験方法 ... 6 2.2.1 試薬 ... 6 2.2.2 細胞 ... 7 2.2.3 培地 ... 7 2.2.4 ヒトおよびカニクイザル iPS 細胞の培養 ... 8 2.2.5 ヒトおよびカニクイザル iPS 細胞の腸管オルガノイドへの分化 ... 8 2.2.6 総リボ核酸 (RNA) 抽出 ... 10 2.2.7 逆転写反応 ... 10 2.2.8 リアルタイム PCR 解析 ... 10 2.2.9 ヘマトキシリン (HE) 染色及びアルシアンブルー染色 ... 13 2.2.10 免疫蛍光染色 ... 13 2.2.11 透過型電子顕微鏡による観察 ... 14 2.2.12 FITC-dextran 4000 (FD-4) の透過実験 ... 14 2.2.13 Rhodamine123 の取り込み実験 ... 14 2.2.14 薬物代謝実験 ... 14 2.2.15 統計解析 ... 15 2.3 結果 ... 16 2.3.1 iPS 細胞から均一な腸管スフェロイドの作製 ... 16
ii 2.3.2 腸管オルガノイドへの分化誘導に対する低分子化合物の影響 ... 17 2.3.3 iPS 細胞由来腸管オルガノイドの形態学的観察 ... 19 2.3.4 iPS 細胞由来腸管オルガノイドの免疫蛍光染色 ... 20 2.3.5 iPS 細胞由来腸管オルガノイドのバリア機能解析 ... 21 2.3.6 iPS 細胞由来腸管オルガノイドの薬物トランスポーター解析 ... 22 2.3.7 iPS 細胞由来腸管オルガノイドの CYP3A4/8 代謝活性および誘導試験 ... 23 2.4 考察 ... 24 2.5 小括 ... 28 第三章 人工多能性幹細胞由来腸管オルガノイドを用いた炎症性腸疾患モデル 系の構築 ... 29 3.1 緒言 ... 29 3.2 実験方法 ... 31 3.2.1 試薬 ... 31 3.2.2 ヒト iPS 細胞の培養 ... 31 3.2.3 ヒト iPS 細胞から腸管オルガノイドへの分化 ... 32 3.2.4 粘膜傷害モデルの作製 ... 32 3.2.5 線維化モデルの作製 ... 32 3.2.6 総リボ核酸 (RNA) 抽出 ... 33 3.2.7 逆転写反応 ... 33 3.2.8 リアルタイム PCR 解析 ... 33 3.2.9 免疫蛍光染色 ... 35 3.2.10 FD-4 の透過実験 ... 35 3.2.11 全コラーゲン量の定量 ... 36
iii 3.2.12 統計解析 ... 36 3.3 結果 ... 37 3.3.1 ヒト iPS 細胞由来腸管オルガノイド (HiOs) の特徴 ... 37 3.3.2 TNF-α 添加による粘膜傷害モデルの構築 ... 39 3.3.3 TNF-α および TGF-β 添加による線維化モデルの構築 ... 43 3.4 考察 ... 47 3.5 小括 ... 50 第四章 総括 ... 51 謝辞 ... 52 引用文献 ... 53
1 第一章 序論 新薬開発では、ターゲットの探索・スクリーニングに始まり、非臨床での様々 な試験による新薬候補の絞り込みを経て、臨床試験へと到達する。薬効が高く、 安全な新薬を患者により早く届けるためには、この創薬プロセスをいかに短く し、新薬探索の成功確率を上げるかが重要である。したがって、探索段階での化 合物の絞り込みを適切に判断する評価手法であるin vitro 細胞アッセイの重要度 が増してきている1, 2)。 腸管は医薬品の吸収、代謝、排泄や免疫機能といった様々な機能を有するとと もに、難治性腸疾患などの複雑な病態を有する疾患も多く存在する。特に、クロ ーン病や潰瘍性大腸炎では、未だ完治することができる治療薬はなく、患者数は 年々増加の一途をたどっている3)。現在、医薬品開発において有用な炎症性腸疾 患 (IBD) による消化管障害を模倣した評価系は存在していないため、この こと が新薬開発を 妨げて いる一因とな ってい る 4)。 カニクイザルは、解剖学的および生理学的にヒトと類似していることから、医 薬品開発での非臨床試験における薬物動態試験や毒性試験に最も広く使用され ている霊長類に属する実験動物である。一方で、カニクイザルとヒトでは、小腸 でのバイオアベイラビリティが異なる医薬品も多く存在する。したがって、ヒト in vivo での体内動態や安全性を正確に予測するには、カニクイザルでの in vitro の評価系とin vivo での結果との相関を検討し、さらに、ヒトとカニクイザルの 間の種差を考慮した新たなアッセイ系を構築することが望まれる。 近年、3 次元組織構造体である腸管オルガノイドは、細胞医薬だけでなく癌メ カニズムの解明、腸の病態モデル作製、薬物スクリーニングなどの分野における 研究材料としても注目を集めている5-7)。創薬スクリーニングでの腸管における
2 in vitro 実験系として機能を考慮すると、初代の腸細胞や組織由来腸管オル ガノイドを使用することが理想的である。しかしながら、細胞や組織を得ること は、侵襲性が非常に高い。特に、医薬品開発では、ハイスループット性が求めら れるため、大量入手が必要となるが、その供給は困難である1)。一方、最近では ヒト人工多能性幹 (iPS) 細胞からも腸管オルガノイドの作製に成功している 8-10)。iPS 細胞は様々な組織細胞へと分化誘導できるだけでなく、健常人あるいは 患者からも作製可能であり、正常な組織細胞や疾患モデルを構築できる。また、 腸組織由来のオルガノイドとは異なり、iPS 細胞由来の腸管オルガノイドでは、 腸上皮細胞だけでなく腸管微小環境を構成している間葉系細胞も含まれている 8, 10)。したがって、多能性幹細胞由来の腸管オルガノイドは、より生体に近い腸 管モデルとして、創薬スクリーニングおよび病態生理を研究するための強力な ツールとなりうる11)。 しかしながら、既存の方法では、腸管オルガノイドを培養する際、マトリゲル 中に包埋するなど煩雑で時間が掛かる。また、方法によっては、均一なサイズの 腸管オルガノイドが大量に得にくい。さらに、多能性幹細胞由来腸管オルガノイ ドは、未成熟な性質であり、医薬品の代謝や排泄といった薬物動態学的な機能に 関する知見が不十分であることが薬物動態解析や薬物スクリーニングに用いる にあたって問題となっている12)。したがって、in vitro アッセイ系として腸管オ ルガノイドを使用するためには、薬物代謝、吸収、排泄および構造的側面などに おいて、これら機能を有する腸管オルガノイドを簡便にかつ大量に作製するこ とが不可欠である。 そこで本研究では、ヒトおよび創薬研究においてヒトと遺伝子プロファイル の相同性が非常に高い動物種として用いられるカニクイザル iPS 細胞から薬物 動態学的機能を有した腸管オルガノイドを簡便にかつ大量に作製する新規分化
3
誘導法の開発を行った。さらに、腸管オルガノイドが薬効 評価を可能 にした新
規 in vitro 評価系として応用できるか検証するため、我々はサイトカイン誘導
4 第二章 多能性幹細胞由来腸管オルガノイドの新規分化誘導法の開発 2.1 諸言 小腸は、経口投与された薬物の体内動態を評価するための重要な器官であり、 シトクロムP450 (CYP) 3A4 を含む薬物代謝酵素や取り込みおよび排出トランス ポーターは小腸で非常に多く発現している13-15)。 特に、CYP3A4 は、現在、臨 床現場で使用されている非常に多くの医薬品の代謝に関与している 16)。したが って、CYP3A4 は医薬品の初回通過代謝において重要な役割を果たしている17)。
さ ら に 、 小 腸 に お け る CYP3A4 発 現 は 、 rifampicin (RIF) お よ び
1α,25-dihydroxyvitamin D3 (VD3) によって誘導されることが知られており18, 19)、小腸に おける CYP3A4 発現の誘導は、経口投与薬のバイオアベイラビリティの低下を 招く。さらに、小腸において高発現している排出トランスポーターABCB1/P-gp もバリアとして機能する 2, 3)。多くの医薬品がCYP3A4 および ABCB1/P-gp の基 質または阻害剤であるため、これらの寄与を考慮して腸内の薬物動態を評価し なければならない。 カニクイザルは、解剖学的および生理学的にヒトと類似しているため、医薬品 開発や毒性研究、生物医学的研究において最も広く使用されている非ヒト霊長 類動物の1 つである20-22)。 カニクイザル小腸および肝臓において最も重要な薬 物代謝酵素であるCYP3A8 は、ヒト CYP3A4 に対して 90%を超えるアミノ酸配 列相同性を示すことから、ヒト CYP3A4 とほぼ同様の基質特異性を示すことが 知られている22)。 例えば、midazolam、fexofenadine、および simvastatin は、カ ニクイザルおよびヒトにおいて ketoconazole と薬物間相互作用を受けることも 知られている23, 24)。 さらに、ラット、イヌ、サル、およびヒトの薬物動態パラ メータを比較した研究では、サルから得られたデータがヒトにおける薬物動態
5 を最も正確に予測できることが示されている25, 26)。 したがって、カニクイザル はヒトの体内動態を予測する上で非常に重要な動物モデルである。 一方、ヒトにおける腸内の薬物動態 (小腸における薬物動態) を直接評価する ことは困難であるため、動物モデルにおけるin vivo と in vitro のデータ、および ヒト in vitro データをヒト in vivo データへと外挿できるか検証する必要がある 27)。具体的には、in vivo でのヒト腸内の薬物動態の予測は、カニクイザル動物モ デルからのin vitro-in vivo 相関データを用いて行うことができる。さらに、非臨 床試験段階での、種差の問題は、ヒトとカニクイザルの両方でin vitro での同様 な評価系を構築し、さらに、薬物の薬物動態プロファイルの違いをスケーリング ファクターを用いて補正することなどによって解決することが可能である。実 際、小腸ミクロソームにおける薬物の内因性代謝クリアランスは、カニクイザル とヒトとでは異なる 28, 29)。したがって、まず、ヒト腸内の薬物動態を正確に評 価するため、in vitro での腸内の薬物動態を正確に反映できる評価システムが必 要である。 近年では、新規in vitro 小腸モデルとして、腸管オルガノイドがそのような薬 物動態学的特性の予測に有用であると期待されている。腸管オルガノイドは、三 次元 (3D) 組織構造を有しており、大きさは直径約 500-1000 µm であるが、近年 では、1-2 cm 程度と大きな腸管オルガノイドも報告されている。また、一般的に は、腸管刷子縁膜側に相当する部分は内側、頂端膜側に相当する部分は外側であ り、管腔構造を有している。形も球状のものから、腸管絨毛-陰窩構造をもつ budding と呼ばれる形状が存在している。以上のことから、腸管組織を模した系 として、現在、再生医療や医薬品開発の分野で注目を集めている。Sato et al. は、 Matrigel 中に包埋培養することで、腸上皮の成長および自己再生が維持できるこ とを報告し、そしてこれらの細胞が腸細胞、杯細胞、パネート細胞および腸内分
6
泌細胞のような全ての腸管細胞を含み、絨毛陰窩構造を形成することを示した
30)。Spence et al. は、この Matrigel 包埋培養法を用いて、ヒト人工多能性幹 (iPS)
細胞から3D 腸管オルガノイドを作製できると報告している8)。作製した腸管オ ルガノイドは極性を示し、絨毛および陰窩様構造を形成し、そして様々な種類の 腸細胞を含んでいた 8)。さらに、腸管オルガノイドを利用した研究として、 Mizutani et al.は、マウス初代小腸細胞から形成された腸管オルガノイドは、 ABCB1/P-gp を介した薬物輸送の評価に有用であると報告している31)。しかしな がら、このMatrigel 中へ包埋培養するといった手法では、非常に煩雑な作業が多 く、培養体積がおよそ50-100 µL と限られていることから大量培養には不向きで ある欠点が存在した 32)。また、我々の知る限りでは、薬物動態学的評価に向け たiPS 細胞由来腸管オルガノイドの応用に関する研究は少なく、さらに、iPS 細 胞から作製された腸管オルガノイドは未熟であることから薬物動態に関連する 機能は不十分であるといった問題点も存在した12)。さらに、カニクイザルiPS 細 胞由来オルガノイドの作製に関しては今日まで報告されていない。 そこで、本研究では、ヒトおよびカニクイザルiPS 細胞から薬物動態学的機能 を有した腸管オルガノイドを簡便にかつ大量に作製する新規分化誘導法の開発 を行った。
7 2.2 実験方法
2.2.1 試薬
Activin A、fibroblast growth factor (FGF) 2 は PeproTech 株式会社 (Rocky Hill, NJ, USA)より、BD Matrigel matrix growth factor reduced (Matrigel)、40-μm nylon-mesh cell strainer は BD Biosciences 株式会社 (Bedford, MA, USA) より、6-[[2-[[4-(2,4- dichlorophenyl)-5-(5-methyl-1H-imidazol-2-yl)-2-pyrimidinyl]amino]ethyl]amino]-3-pyridinecarbonitrile (CHIR99021) 、 (+)-(R)-trans-4-(1-aminoethyl)-N-(4-pyridyl) cyclohexanecarboxamide dihydrochloride (Y-27632) は Focus Biomolecules (Plymouth Meeting, PA, USA) より、 FGF4 は BioLegend (San Diego, CA, USA)より、 EZSPHERE (#900: microwell size: diameter 500 μm, depth 100 μm) は、AGC テクノ
ガラス株式会社 (Shizuoka, Japan) より、KnockOut serum replacement (KSR)、
Advanced Dulbecco's modified Eagle medium/Ham’s F-12 (DMEM/F12) 、 N2 supplement、B27 serum-free supplement、SlowFade Diamond antifade mountant は、 Thermo Fischer Scientific 社 (Waltham, MA, USA) より、ウシ胎児血清 (FBS) は、
ニチレイバイオサイエンス社 (Tokyo, Japan) より、R-spondin 1、Noggin、epidermal
growth factor (EGF) は GenScript (Piscataway, NJ, USA) より、StemSure® hPSC medium、 verapamil、RIF、VD3 は、富士フィルム和光純薬株式会社 (Osaka, Japan) より、2-(2-amino-3-methoxyphenyl)4H-1-benzopyran-4-on (PD98059)、3-(6-methyl-2-pyridinyl)-N-phenyl-4-(4-quinolinyl)-1H-pyrazole-1-carbothioamide (A-83-01) は 、 AdooQ BIOSCIENCE (Irvine, CA, USA)より、5-aza-2’-deoxycytidine は、Chem-impex international 社 (Wood Dale, IL, USA) より、N-[(3,5-difluorophenyl) acetyl]-L-alanyl-2-phenyl-1,1-dimethylethyl ester-glycine (DAPT) は、ペプチド研究所 (Osaka, Japan)
より、OCT compound は、サクラファインテックジャパン株式会社 (Tokyo, Japan)
8
量: 4000) は、Sigma-Aldrich (St. Louis, MO, USA) より、rhodamine123 は、タカ
ラバイオ社 (Shiga, Japan) より、total RNA from human small intestine samples (5 donors) は、BioChain Institute Inc. (Newark, CA, USA) より購入して用いた。その 他の試薬は全て市販の高速液体クロマトグラフ用 (LC-MS 用) もしくは特級品
を用いた。
2.2.2 細胞
ヒトiPS 細胞 (#51:Windy) は、ヒト胎児肺線維芽細胞 MRC-5 に octamer binding
protein 3/4 (OCT3/4)、sex determining region Y-box 2 (SOX2)、kruppel-like factor 4 (KLF4)、v-myc myelocytomatosis viral oncogene homolog (avian) (c-MYC) を、パン
トロピックレトロウイルスベクターを用いて導入後、ヒト胚性幹 (ES) 細胞様コ
ロニーをクローン化したものであり、国立成育医療研究センター梅澤明弘博士
よりご供与いただいた。カニクイザル iPS 細胞はカニクイザル皮膚組織より培
養 し た 線維 芽細 胞 にエ ピ ソ ーマ ルベ ク ター で あ る pCXLE-hOCT3/4-shp53-F
(OCT3/4 と p53 shRNA がタンデムに繋がったベクター)、pCXLE-hSK (SOX2 と KLF4 がタンデムに繋がったベクター)、pCXLE-hUL (L-MYC と LIN28 がタンデ
ムに繋がったベクター) をエレクトロポレーション法にて導入後、カニクイザル ES 細胞様コロニーをクローン化したものであり、当研究室で樹立した。フィー ダー細胞はマウス胎仔線維芽細胞 (MEF) を使用した。 2.2.3 培地 MEF の培養には 10% FBS、2 mM L-グルタミン (L-Glu)、1%非必須アミノ酸 (NEAA)、100 units/mL ペニシリン G、100 µg/mL ストレプトマイシンを含む DMEM を用いた。MEF の剥離液には 0.05%トリプシン-エチレンジアミン四酢酸
9
(EDTA) を、MEF の保存液にはセルバンカー1 を用いた。ヒト iPS 細胞の維持培
養には20% KSR、0.8% NEAA、2 mM L-Glu、0.1 mM 2-メルカプトエタノール
(2-ME)、5 ng/mL FGF2 を含む DMEM/F12 を用いた。カニクイザル iPS 細胞の維持
培養には20% KSR、1.0% NEAA、2 mM L-Glu、100 units/mL ペニシリン G、100
µg/mL ストレプトマイシン、0.1 mM 2-ME、5 ng/mL FGF2 を含む DMEM/F12 を 用いた。ヒトおよびカニクイザルiPS 細胞の剥離液には 1 mg/mL コラゲナーゼ IV、0.25%トリプシン、20% KSR、1 mM 塩化カルシウムを含むダルベッコリン 酸緩衝生理食塩水 (PBS) を用いた。ヒトおよびカニクイザル iPS 細胞の保存液 には霊長類ES/iPS 細胞用凍結保存液を用いた。 2.2.4 ヒトおよびカニクイザル iPS 細胞の培養
ヒトiPS 細胞はマイトマイシン C 処理を施した MEF (6×105 cells/100 mm ディ
ッシュ) 上に、カニクイザル iPS 細胞はマイトマイシン C 処理を施した MEF (1×106 cells/100 mm ディッシュ) 上に播種し、5% CO 2/95% air 条件下 CO2インキ ュベーター中 37°C にて培養した。ヒトおよびカニクイザル iPS 細胞の継代は、 3~5 日培養後、1:2~1:3 のスプリット比で行った。ヒトおよびカニクイザル iPS 細 胞は解凍48 時間後に培地を交換し、それ以降は毎日交換した。 2.2.5 ヒトおよびカニクイザル iPS 細胞の腸管オルガノイドへの分化 ヒトおよびカニクイザル iPS 細胞の腸管オルガノイドへの分化は、継代時に ヒトおよびカニクイザルiPS 細胞用培地にて 30 倍に希釈した Matrigel にてコー トした培養ディッシュに播種し、35 ng/mL FGF2 を含む StemSure® hPSC 培地に て培養し、未分化コロニーの占める割合が約80%になった状態で開始した。100
10
mM L-Glu を含むロズウェルパーク記念研究所 (RPMI) 培地で 1 日間、0.2% FBS、 100 ng/mL activin A、100 units/mL ペニシリン G、100 µg/mL ストレプトマイシン、 2 mM L-Glu を含む RPMI 培地で 1 日間、2% FBS、100 ng/mL activin A、100 units/mL
ペニシリンG、100 µg/mL ストレプトマイシン、2 mM L-Glu を含む RPMI 培地 で1 日間培養することで内胚葉に分化させた。その後、ヒト iPS 細胞では、2% FBS、3 µM CHIR99021、500 ng/mL FGF4、100 units/mL ペニシリン G、100 µg/mL ストレプトマイシンを含む RPMI+グルタマックス培地で 4 日間培養すること で腸管幹細胞へ分化させた。カニクイザル iPS 細胞では、2% FBS、6 µM CHIR99021、250 ng/mL FGF2、100 units/mL ペニシリン G、100 µg/mL ストレプ トマイシンを含むRPMI+グルタマックス培地で 4 日間培養することで中・後腸
へ分化させた。ヒトiPS 細胞では CHIR99021、FGF4 処理後、カニクイザル iPS
細胞では、FGF2、CHIR99021 処理後、細胞を 0.05%トリプシン-EDTA にて剥離
し、40 µm nylon-mesh cell strainer にて細胞塊を砕き、4.0×106 細胞を 100 mm
EZSPHERE® (#900: microwell size: diameter 500 μm, depth 100 μm) 上に播種した。
その後、2 mM L-Glu、2% B27 supplement、1% N2 supplement、100 units/mL ペニ
シリンG、100 µg/mL ストレプトマイシン、100 ng/mL EGF、100 ng/mL Noggin、
200 ng/mL R-spondin-1、10 µM Y-27632 を含む Advanced-DMEM/F12 で 3 日間培 養後、2 mM L-Glu、2% B27 supplement、1% N2 supplement、100 units/mL ペニシ
リンG、100 µg/mL ストレプトマイシン、100 ng/mL EGF、100 ng/mL Noggin、200
ng/mL R-spondin-1、Matrigel 3%を含む Advanced-DMEM/F12 で 24 日間超低接着 6 well-plate 上で浮遊培養することで腸管オルガノイドへ分化させた。薬物代謝 酵素の誘導剤処理は、2 mM L-Glu、2% B27 supplement、1% N2 supplement、100 units/mL ペニシリン G、100 µg/mL ストレプトマイシン、100 ng/mL EGF、100 ng/mL Noggin、200 ng/mL R-spondin-1、Matrigel 3%を含む Advanced-DMEM/F12
11 にRIF を 40 µM もしくは (VD3) を 1 µM となるよう添加し、回収前 72 時間培 養することで行った。また、分化開始19 日目から 34 日目まで、以前、当研究室 にて見出した低分子化合物33)であるPD98059 (20 µM)、5-aza-2’-deoxycytidine (5 µM)、A-83-01 (0.5 µM) に加え、DAPT (5 µM) を添加し、腸管オルガノイドへの 分化に及ぼす影響について検討した (Figure 1)。
Figure 1 Schematic of intestinal organoid differentiation from human and cynomolgus iPS cells.
2.2.6 総リボ核酸 (RNA) 抽出
総 RNA はヒトおよびカニクイザル iPS 細胞の分化誘導終了後、Agencourt
RNAdvence Tissue の添付マニュアルに従い抽出した。
2.2.7 逆転写反応
相補的DNA (cDNA) の合成は、ReverTra Ace qPCR RT Master Mix を使用し、
添付マニュアルに従い行った。
2.2.8 リアルタイム PCR 解析
PCR プライマーは、ヒトでは Table 1、カニクイザルでは Table 2 に示したもの
を用いた。Real-Time RT-PCR は KAPA SYBR Fast qPCR Kit を用い、cDNA を鋳
型にして、反応は添付マニュアルに従い行った。結果は内在性コントロールとし
12
ニクイザルではグリセルアルデヒド-3-リン酸脱水素酵素 (GAPDH) を用いて補
13
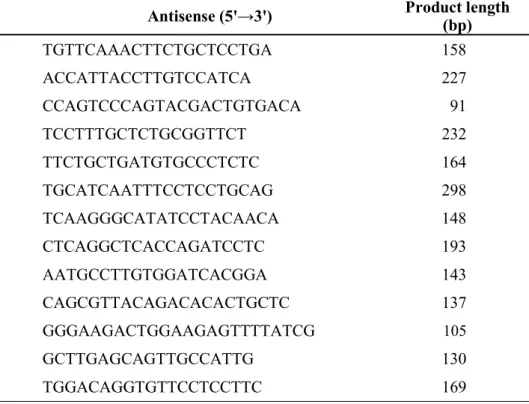
Table 1 Primers used for real-time PCR analysis of human.
Gene name Sense (5'→3') Antisense (5'→3') Product length (bp) ABCB1/P-gp CCCATCATTGCAATAGCAGG TGTTCAAACTTCTGCTCCTGA 158
ABCC2/MRP2 ACAGAGGCTGGCAACC ACCATTACCTTGTCCATCA 227
ABCG2/BCRP AGATGGGTTTCCAAGCGTTCAT CCAGTCCCAGTACGACTGTGACA 91
CDX2 ACCTGTGCGAGTGGATGC TCCTTTGCTCTGCGGTTCT 232
Chromogranin A TCCGACACACTTTCCAAGCC TTCTGCTGATGTGCCCTCTC 164 CYP3A4 CTGTGTGTTTCCAAGAGAAGTTAC TGCATCAATTTCCTCCTGCAG 298
HPRT CTTTGCTTTCCTTGGTCAGG TCAAGGGCATATCCTACAACA 148
LGR5 TGCTCTTCACCAACTGCATC CTCAGGCTCACCAGATCCTC 193
Lysozyme TCAATAGCCGCTACTGGTGT AATGCCTTGTGGATCACGGA 143 MUC2 AGAAGGCACCGTATATGACGAC CAGCGTTACAGACACACTGCTC 137 SLC15A1/PEPT1 CACCTCCTTGAAGAAGATGGCA GGGAAGACTGGAAGAGTTTTATCG 105 SLCO2B1/OATP2B1 CTTCATCTCGGAGCCATACC GCTTGAGCAGTTGCCATTG 130 Villin AGCCAGATCACTGCTGAGGT TGGACAGGTGTTCCTCCTTC 169
14
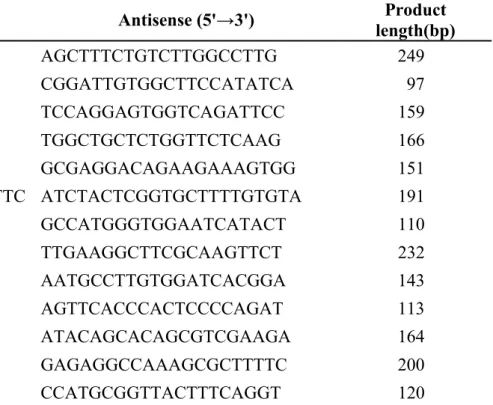
Table 2 Primers used for real-time PCR analysis of cynomolgus monkey.
Gene Name Sense (5'→3') Antisense (5'→3') length(bp) Product ABCB1/P-gp TGGTTTCGATGATGGAGTCA AGCTTTCTGTCTTGGCCTTG 249 ABCC2/MRP2 GACACATCTGCCATTCGACAT CGGATTGTGGCTTCCATATCA 97 ABCG2/BCRP GGTCACGTAACCTTGGGGTA TCCAGGAGTGGTCAGATTCC 159 Chrmogranin A CACTCCGAGGAGATGAACGG TGGCTGCTCTGGTTCTCAAG 166
CDX2 CCCCACAACCCAGCTAGTTA GCGAGGACAGAAGAAAGTGG 151
CYP3A8 CCAAGAAGCTTTTAAGATTTGATTTC ATCTACTCGGTGCTTTTGTGTA 191 GAPDH TCACCAGGGCTGCTTTTAAC GCCATGGGTGGAATCATACT 110
LGR5 CAGCGTCTTCACCTCCTACC TTGAAGGCTTCGCAAGTTCT 232
Lysozyme TCAATAGCCGCTACTGGTGT AATGCCTTGTGGATCACGGA 143
MUC2 ACACCTACACGCACATCGAG AGTTCACCCACTCCCCAGAT 113
SLC15A1/PEPT1 CAATGTTCTGGGCCTTGTTT ATACAGCACAGCGTCGAAGA 164 SLCO2B1/OATP2B1 AAGGAAACCAAGGCCACGAT GAGAGGCCAAAGCGCTTTTC 200 Villin GGAACGTGGTAGCTGGAGAG CCATGCGGTTACTTTCAGGT 120
15 2.2.9 ヘマトキシリン-エオシン (HE) 染色及びアルシアンブルー染色 分化誘導終了後、腸管オルガノイドを 4%パラホルムアルデヒドにて固定し、 OCT compound にて凍結包埋した。厚さ 10 µm の凍結切片を作製後、スライドガ ラスに貼り付け、HE 染色では、マイヤーヘマトキシリン及びエオシンアルコー ルを使用して染色した。アルシアンブルー染色では、pH 2.5 におけるアルシアン ブルーを染色し、核染色としてヌクレアファストレッドを使用した。 2.2.10 免疫蛍光染色 分化誘導終了後、腸管オルガノイドを 4%パラホルムアルデヒドにて固定し、 OCT compound にて凍結包埋した。厚さ 10 µm の凍結切片を作製後、スライドガ ラスに貼り付け、抗原の賦活化を行った。5% FBS 溶液にて 30 分間ブロッキン グし、一次抗体を4℃で 1 晩反応させた。その後、スライドガラスを洗浄し、二 次抗体を室温で 1 時間反応させ、核染色として 4',6-diamidino-2-phenylindole
(DAPI) を用いた。用いた抗体は Table 3 に示した。封入作業を行い、Zeiss LSM510 共焦点レーザー顕微鏡を用いて、蛍光を観察した。
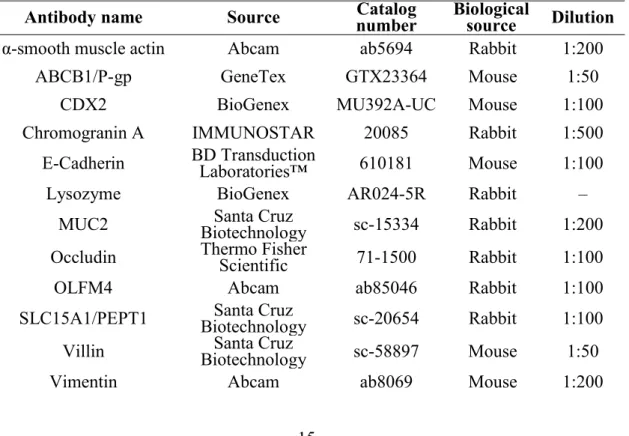
Table 3 Primary and secondary antibodies.
Antibody name Source number Catalog Biological source Dilution
α-smooth muscle actin Abcam ab5694 Rabbit 1:200
ABCB1/P-gp GeneTex GTX23364 Mouse 1:50
CDX2 BioGenex MU392A-UC Mouse 1:100
Chromogranin A IMMUNOSTAR 20085 Rabbit 1:500
E-Cadherin BD Transduction Laboratories™ 610181 Mouse 1:100
Lysozyme BioGenex AR024-5R Rabbit –
MUC2 Biotechnology Santa Cruz sc-15334 Rabbit 1:200
Occludin Thermo Fisher Scientific 71-1500 Rabbit 1:100
OLFM4 Abcam ab85046 Rabbit 1:100
SLC15A1/PEPT1 Biotechnology Santa Cruz sc-20654 Rabbit 1:100
Villin Biotechnology Santa Cruz sc-58897 Mouse 1:50
16 Alexa Fluor® 488
donkey Anti-Rabbit IgG (H&L)
Thermo Fisher
Scientific A-21206 Rabbit 1:200
Alexa Fluor® 488 goat
anti-Mouse IgG (H&L) Thermo Fisher Scientific A-11001 Mouse 1:200 Alexa Fluor® 568 goat
anti-Mouse IgG (H&L) Thermo Fisher Scientific A-11004 Mouse 1:200 2.2.11 透過型電子顕微鏡による観察 分化誘導終了後、腸管オルガノイドを2.5%グルタルアルデヒドにて前固定し、 4℃で一晩インキュベーションした。その後、1%四酸化オスミウムにて 4℃で 2 時間、後固定を行い、エタノールによる脱水作業を行った。プロピレンオキサイ ドに置換した後、樹脂内に包埋した。0.1 µm の切片を作製し、酢酸ウラニルに て電子染色を行った後、JEM-1400 Plus 透過型電子顕微鏡を用いて、微絨毛及び タイトジャンクションの観察を行った。 2.2.12 FITC-dextran 4000 (FD-4) の透過実験 分化誘導終了後、腸管オルガノイドを FD-4 を含む HBSS (ハンクス緩衝塩類 溶液) で 37°C にてインキュベーションした。HBSS は、137 mM 塩化ナトリウ ム、5.4 mM 塩化カリウム、0.81 mM 硫酸マグネシウム、0.44 mM リン酸二水素 カリウム、0.34 mM リン酸水素二ナトリウム、1.3 mM 塩化カルシウム、4.2 mM 炭酸水素ナトリウム、5.6 mM D-グルコース、10 mM HEPES を含む pH 7.4 のも のを用いた。インキュベーション終了後、氷冷したHBSS で細胞を洗浄した。そ
の後、Nikon ECLIPSE Ti-S 顕微鏡を用いて、蛍光を観察した。
2.2.13 Rhodamine123 の取り込み実験 分化誘導終了後、腸管オルガノイドを rhodamine123 を含む HBSS で 37°C に てインキュベーションした。インキュベーション終了後、氷冷した HBSS で細 胞を洗浄することにより取り込みを停止させた。その後、Zeiss LSM510 共焦点 レーザー顕微鏡を用いて、蛍光を観察した。 2.2.14 薬物代謝実験 分化誘導終了後、腸管オルガノイドを5 µM midazolam を含む培地 2 mM L-Glu、
17
2% B27 supplement、1% N2 supplement、100 units/mL ペニシリン G、100 µg/mL ス
トレプトマイシンを含むAdvanced-DMEM/F12 で 37°C にてインキュベーション し、24 時間経過後、培地をサンプリングした。代謝活性は、液体クロマトグラ フィー–マススペクトロメーター (LC–MS/MS) を用いて測定した培地中の 1’-OH midazolam の量より算出した。代謝実験終了後、タンパク定量を行い、代謝 活性をタンパク量で補正した。 2.2.15 統計解析 「n」は biological replicate の数を表している。それぞれの実験で 3 サンプルず つ回収した。それぞれの実験は少なくとも 2 回繰り返し行われた。定量的なデ ータはmeans ± S.D.として表した。多重比較は、一元配置分散分析を行った後、
two-tailed Dunnett’s test あるいは Bonferroni’s correction を用いて実行した (Figure 3, 4 and 9b)。また、2 群間比較は、Student’s t-test を用いて解析した (Figure 9a)。 その際の統計分析は、 PASW Statistict 18 system software を用いて行われた。
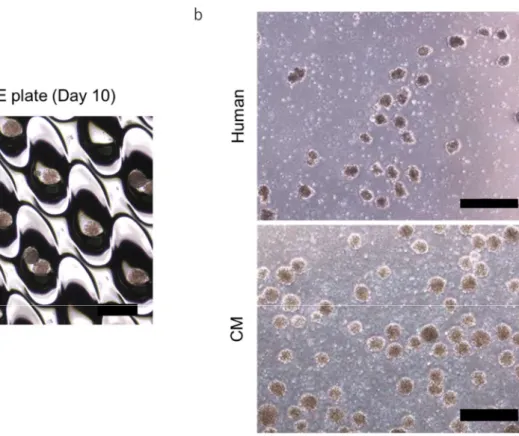
18 2.3 結果 2.3.1 iPS 細胞から均一な腸管スフェロイドの作製 腸管オルガノイドへの分化誘導は、ヒトおよびカニクイザルiPS 細胞に activin A で 3 日間処理することで内胚葉へと誘導し、さらに 4 日間ヒトでは、CHIR99021 およびFGF4 を、カニクイザルでは、CHIR99021、FGF2 を処理することで中・ 後腸へ分化させた。その後、EZSPHERE プレートを用い、均一なスフェロイド を形成させた。続いて、腸管への分化培地に切り替え、分化工程の19 日目から 34 日目まで低分子化合物を添加することで、腸管オルガノイドの作製を行った (Figure 1)。その結果、EZSPHERE プレートを使用することで均一なサイズの腸 管オルガノイドを簡便かつ大量に作製することが可能となった (Figure 2a)。ま た、浮遊培養させることで、容易に大量培養することができた (Figure 2b)。
Figure 2 Morphology of human and cynomolgus monkey (CM) iPS cell-derived intestinal spheroids.
(a) Uniform spheroids on an EZSPHERE plate at day 10. Scale bar, 500 µm. (b) Spheroids in suspension culture. Scale bar, 500 µm.
19 2.3.2 腸管オルガノイドへの分化誘導に対する低分子化合物の影響 ヒトおよびカニクイザル iPS 細胞から腸管オルガノイドへの分化誘導の際に添 加する低分子化合物の影響について調べた。その結果、A-83-01 (0.5 µM)、PD98059 (20 µM)、5-aza-2’-deoxycytidine (5 µM) を添加した群 (A/PD/5-aza) で腸管関連遺 伝子や CYP3A 等をはじめとする様々な薬物動態関連遺伝子の mRNA 発現量は
増加した (Figure 3 and 4)。さらに、DAPT (5 µM) を加えた群 (A/PD/5-aza/DAPT)
でヒト腸管オルガノイドにおいては、低分子化合物非添加群であるnon treatment
群に比べ、腸管での主要な薬物代謝酵素であるCYP3A4 で約 2,000 倍、ペプチド
の取り込みトランスポーターであるSLC15A1/PEPT1 で約 10 倍、排出トランス
ポーターであるABCB1/P-gp で約 36 倍、ABCG2/BCRP で約 125 倍、ABCC2/MRP2
で約47 倍など多くの薬物動態関連遺伝子の mRNA 発現が上昇した (Figure 3)。
また、腸管を構成している細胞マーカーである villin1 (吸収上皮細胞)、ムチン
(MUC) 2 (杯細胞)、chromogranin A (腸管内分泌細胞)、尾側タイプホメオボックス (CDX) 2 (腸管系譜の細胞)、LGR5 (腸管幹細胞) は non treatment 群に比べ、同程
度もしくはそれ以上の mRNA 発現を示した (Figure 3)。カニクイザル腸管オル
ガノイドにおいてもヒトと同様の傾向を示し、non treatment 群に比べ、CYP3A8
で約 1,400 倍、SLC15A1/PEPT1 で約 2,200 倍、ABCB1/P-gp で約 18 倍、
ABCG2/BCRP で約 2.8 倍、ABCC2/MRP2 で約 27 倍など多くの薬物動態関連遺
伝子のmRNA 発現が上昇した (Figure 4)。また、villin、MUC2、chromogranin A、
lysozyme、CDX2 は non treatment 群に比べ、同程度もしくはそれ以上の mRNA 発 現を示した (Figure 4)。これまでに、A-83-01、PD98059、5-aza-2’-deoxycytidine、 DAPT の組み合わせの他に、cAMP シグナル経路を活性化させる 8-Br-cAMP や cAMP の 分 解 を 抑 制 す る IBMX 、 Wnt シ グ ナ ル 経 路 を 活 性 化 さ せ る 6-bromoindirubin-3′-oxime、CHIR99021、また DAPT のみを添加したが、A-83-01、 PD98059、5-aza-2’-deoxycytidine、DAPT 群のような mRNA 発現上昇効果は認め られなかった (データは示していない)。しかし、これらの化合物について添加 濃度、添加期間、組み合わせに対する詳細な検討はなされていないため、今後、 より最適な分化誘導法を開発するために再度検証する必要があると考えられる。
20
21
Figure 3 mRNA expression of intestinal markers and pharmacokinetic-related genes in human iPS cell-derived intestinal organoids. 0.25 % DMSO (Non treatment group), A-83-01/PD98059/5-aza-2′-deoxycytidine aza) or A-83-01/PD98059/5-A-83-01/PD98059/5-aza-2′-deoxycytidine/DAPT (A/PD/5-aza/DAPT) were added on days 19-to-34. After 34 days of differentiation, total RNA was extracted, and mRNAs were analyzed using SYBR Green real-time PCR. Data are represented as means ± S.D. (n = 3). All mRNA expression levels were normalized relative to the mRNA level of HPRT. Gene expression levels are represented relative to the level in the adult small intestine, which was set to 1. Statistical analysis was performed using Bonferroni’s correction: *P <0.05, #P<0.01.
Figure 4 mRNA expression of intestinal markers and pharmacokinetic-related genes in the CM iPS cell-derived intestinal organoids.
0.25 % DMSO (Non treatment group), A-83-01/PD98059/5-aza-2′-deoxycytidine (A/PD/5-aza) or A-83-01/PD98059/5-aza-2′-deoxycytidine/DAPT (A/PD/5-aza/DAPT) were added on days 19-to-34. After 34 days of differentiation, total RNA was extracted, and mRNA expression was analysed by real-time PCR. Data are represented as the mean ± S.D. (n = 3). mRNA expression levels were normalised to the expression of GAPDH. Gene expression levels are represented as relative to the levels in the small intestine, set to 1. Statistical analysis was performed using Bonferroni’s method, *P < 0.05, #P < 0.01.
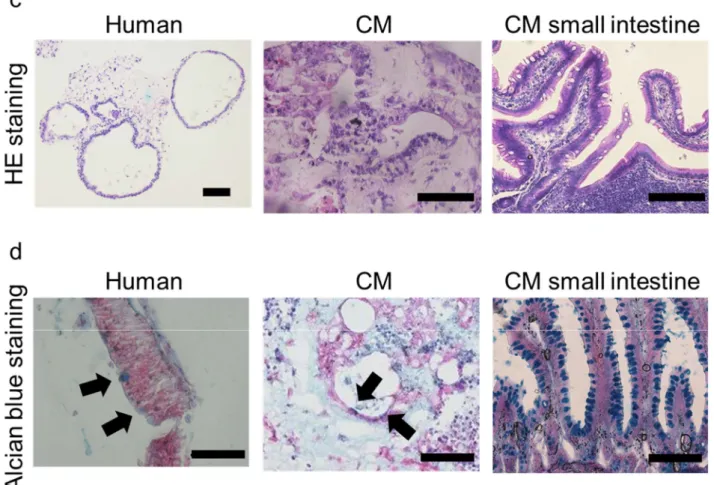
22 2.3.3 iPS 細胞由来腸管オルガノイドの形態学的観察 低分子化合物の添加により分化誘導したヒトおよびカニクイザル iPS 細胞由 来腸管オルガノイドは球状の形をしていた (Figure 5a)。透過型電子顕微鏡の観 察から腸管オルガノイドの内側に腸管刷子縁膜側に存在する微絨毛およびタイ トジャンクションの形成が確認された (Figure 5b)。また、HE 染色やアルシアン ブルー染色の結果から、腸管オルガノイドは分泌細胞を含む細胞集団であるこ とが分かった (Figure 5c and d)。
23
Figure 5 Morphology of human and CM iPS cell-derived intestinal organoids.
(a) Images acquired by phase contrast microscopy. Scale bar, 500 μm. (b) TEM images: brush border with microvilli (MV, white arrow) and tight junction (TJ, white arrow head). Scale bar, 1 μm. (c) HE staining of human and CM iPS cell-derived intestinal organoids and CM small intestine. Scale bar, 100 µm. (d) Alcian blue staining of human and CM iPS cell-derived intestinal organoids, and CM small intestine. Black arrow shows Alcian blue-positive cells. Scale bar, 100 µm.
24
2.3.4 iPS 細胞由来腸管オルガノイドの免疫蛍光染色
免疫蛍光染色により、腸管を構成している様々な細胞(吸収上皮細胞、腸管幹 細胞、杯細胞、腸管内分泌細胞、パネート細胞、間葉系細胞)のマーカーの発現
25
Figure 6 Immunofluorescence staining of intestinal cell markers in human and CM iPS cell-derived intestinal organoids
Immunofluorescence staining for villin, OLFM4, CDX2, MUC2, E-cadherin, chromogranin A, lysozyme, vimentin, and α-SMA in human and CM iPS cell-derived intestinal organoids and CM small intestine. Scale bar, 50 µm. Nuclei were counterstained with DAPI.
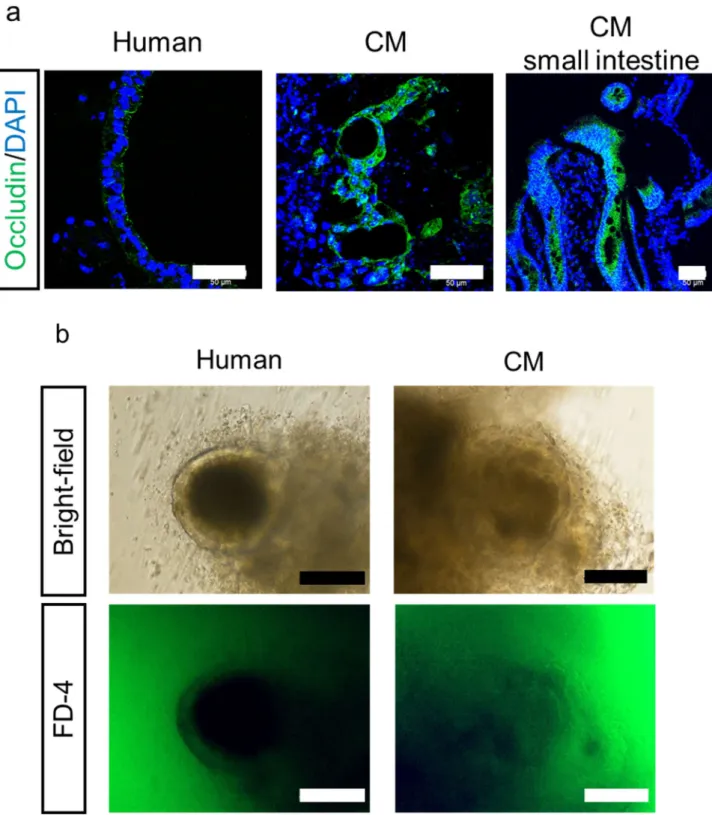
26 2.3.5 iPS 細胞由来腸管オルガノイドのバリア機能解析 機能的なタイトジャンクションが形成されているか確認するため、まず免疫 蛍光染色にて、タイトジャンクションマーカーである occludin のタンパク質発 現を確認した (Figure 7a)。この結果から、タイトジャンクションは、腸管オルガ ノイドの管腔側に沿って発現していることが示唆された。また、透過型電子顕微 鏡の観察から、腸管オルガノイドの管腔内側が腸管刷子縁膜側に相当すること が明らかとなった (Figure 4b)。そこで、非吸収性マーカーである FD-4 を用いた 腸管オルガノイド内への透過試験を行ったところ、オルガノイド内への蓄積は 認められなかった (Figure 7b)。
27
Figure 7 Analysis of barrier function.
(a): Immunofluorescence staining of tight junction marker occludin (green). Scale bar, 50 µm. Nuclei were counterstained with DAPI (blue). (b): FD-4 permeability in human iPS cell-derived intestinal organoids. Organoids were incubated with FD-4 (1 mg/mL) for 1 hour at 37°C. Bright-field image and fluorescence image. Scale bar, 50 µm.
28
2.3.6 iPS 細胞由来腸管オルガノイドの薬物トランスポーター解析
ヒトおよびカニクイザル腸管オルガノイドでは、小腸に特異的に発現してい
るSLC15A1/PEPT1、および腸管で主要な排泄トランスポーターである
ABCB1/P-gp のタンパク質発現が腸管オルガノイドの内側管腔に沿って認められた (Figure 8a)。そこで、排出トランスポーターである ABCB1/P-gp の基質である rhodamine123 および阻害剤である verapamil を用いて、ABCB1/P-gp の機能評価
を行った。rhodamine123 の腸管オルガノイド内への排出が認められ (Figure 8b)、
29
Figure 8 The expression of drug transporters and efflux transport of rhodamine123.
(a): Immunofluorescence staining of SLC15A1/PEPT1 (A, green), ABCB1/P-gp (B, green). Scale bar, 50 µm. Nuclei were counterstained with DAPI (blue). (b): Efflux transport of rhodamine123 through ABCB1/P-gp. Organoids were incubated with rhodamine123 (10 µM) for 1 hour at 37°C in the absence or presence of verapamil (100 µM). Bright-field images. Fluorescence images. Scale bar, 100 µm.
30
2.3.7 iPS 細胞由来腸管オルガノイドの CYP3A4/8 代謝活性および誘導
CYP3A4/8 の基質である midazolam とその阻害剤である ketoconazole を用いて、 CYP3A4/8 の代謝活性を測定した。ヒトおよびカニクイザル iPS 細胞由来腸管オ
ル ガ イ ド に お い て 、midazolam の 代 謝 活 性 が 認 め ら れ 、 そ の 残 存 活 性 は
ketoconazole の添加により、ヒトでは約 38%、カニクイザルでは約 5%にまで、
有意に阻害された (Figure 9a)。また、CYP3A の誘導剤である RIF および VD3
を用いて、CYP3A4/8 の代謝活性の誘導について検討した。ヒトでは RIF 添加群
で1.5 倍、VD3 添加群で 2.0 倍有意に誘導された。一方、カニクイザルでは RIF
添加群で7.5 倍有意に誘導されたが、VD3 添加群では 3.5 倍であり、有意な変化
31
Figure 9 Activity and induction of CYP3A4/8.
(a) Metabolic activity of CYP3A4/8 in the human and CM iPS cell-derived intestinal organoids. For the analysis of metabolic activity, human and CM iPS cell-derived intestinal organoids were incubated with 5 µM midazolam for 24 h at 37 °C in the absence or presence of ketoconazole. The supernatant was recovered and metabolites were analysed by LC–MS/MS. Data are represented as the mean ± S.D. (n = 4). The statistical analysis was performed using a two-tailed Student’s t-test; *: *: P < 0.05,#: P < 0.01. (b) Induction of metabolic activity of CYP3A4/8 in the human and CM iPS cell-derived intestinal organoids: 40 µM rifampicin or 10 µM 1α,25-dihydroxyvitamin D3 (VD3) were added in the final 72 h of incubation. For the analysis of metabolic activity, human and CM iPS cell-derived intestinal organoids were incubated with 5 µM midazolam for 24 h at 37 °C after treatment with the inducers. The supernatant was recovered and metabolites were analysed by LC–MS/MS. Data are represented as the mean ± S.D. (n = 4). The activity is represented relative to the level in control group, which was set to 1. The statistical analysis was performed using the two-tailed Dunnett’s test; *: P < 0.05,#: P < 0.01.
32 2.4 考察 本研究では、A/PD/5-aza/DAPT の低分子化合物の組み合わせを分化誘導時に使 用することでヒトおよびカニクイザル iPS 細胞から薬物動態学的機能を有する 腸管オルガノイドを作製することに成功した。iPS 細胞由来腸管オルガノイドの 作製は、以前にいくつか報告されている8, 9, 34, 35)。しかし、これら既存の方法で は、Matrigel 中へ包埋するなど煩雑で時間がかかり、そして培養体積が限られる ことから多くの腸管オルガノイドを一度に培養することが困難であった 32)。本 研究では、EZSPHERE plate を使用して均一なスフェロイドを大量に作製した後 に、浮遊培養を行ったため、多数の均一なスフェロイドを簡便に作製し、大量培 養することに成功した (Figure 2)。また、Matrigel 包埋法のような以前の方法で は、培養体積が限られることから薬物スクリーニングおよびハイスループット スクリーニングへの応用は困難であったが、我々の方法では大量で均一な腸管 オルガノイドを必要とする解析への応用を可能にする。 腸管分化を促進するために、いくつかの化合物を試験し、A/PD/5-aza/DAPT 添 加群が薬物動態関連遺伝子のmRNA 発現が著しく増加することを明らかにした
(Figure 3 and 4)。また、A/PD/5-aza/DAPT 添加群ではヒトおよびカニクイザル両
方でMUC2、chromogranin A、CDX2 などの様々な腸細胞マーカーの mRNA 発現
に顕著な増加が認められた (Figure 3 and 4)。一方、ヒトではパネート細胞マーカ
ーであるlysozyme の mRNA 発現は、non treatment 群の mRNA 発現レベルと比
較して有意に減少したが、吸収上皮細胞マーカーである villin1 および腸管幹細
胞マーカーである LGR5 の mRNA 発現レベルは変化しなかった (Figure 3)。カ
ニクイザルでは、lysozyme の mRNA 発現は non treatment 群に比べて有意に増加 したが、LGR5 の mRNA 発現レベルは減少した。これらの結果から、ヒトでは A/PD/5-aza/DAPT により、腸管上皮細胞自身の分化、並びにパネート細胞および 腸管幹細胞というよりもむしろ分泌細胞を含む腸管細胞への分化が促進された こと、カニクイザルでは腸管上皮細胞への分化誘導が促進されたことが示唆さ れた。また、これらmRNA 発現変動の違いは、iPS 細胞の株間による分化指向性 の違いに起因したことである可能性が示唆された。以前、我々は、A-83-01、 PD98059、および 5-aza-2’-deoxycytidine の組み合わせが、ヒト iPS 細胞の腸管上
33 皮細胞様細胞への分化を促進することを報告した33)。また、DAPT および wnt ア ゴニストは、ヒトES 細胞の腸管上皮細胞への分化を誘導することが報告されて いる36)。さらに、DAPT が notch 活性化を阻害することで、腸管上皮細胞の成熟 または分泌細胞への分化を促進することも報告されている 37)。これらの結果に 基づいて、我々はこれらの低分子化合物が相加的に効果を発揮することによっ て、iPS 細胞由来腸管オルガノイドの分化を促進することを示唆した。しかし、 将来的には詳細なメカニズムをさらに検証する必要がある。 ヒトおよびカニクイザルiPS 細胞由来腸管オルガノイドでは、villin1、occludin、 SLC15A1/PEPT1、および ABCB1/P-gp のタンパク質発現は、腸管オルガノイド 内腔に沿って局在しており、透過型電子顕微鏡観察から微絨毛およびタイトジ
ャンクションも腸管オルガノイドに存在していた (Figure 5b, 6, 7a, 8a)。これら
の結果から、腸管オルガノイドの管腔内側が腸管刷子縁膜側に相当することが 示唆された。さらに、非吸収性マーカーであるFD-4 の腸管オルガノイド内への 透過は認められず (Figure 7b)、ABCB1/P-gp を介した排出方向の輸送が検出され た (Figure 8b)。 近年 、マウ スや ヒト組 織 由来腸 管オ ルガノ イ ドを用 いて ABCB1/P-gp の排泄機能を定量化する研究が報告され、それらは ABCB1/P-gp 阻 害剤スクリーニング系として応用されることが期待されている31, 38, 39)。 本研究
では、A/PD/5-aza/DAPT を用いてヒト iPS 細胞から ABCB1/P-gp を介した排泄機
能 を 持 つ 腸 管 オ ル ガ ノ イ ド を 作 製 す る こ と に 成 功 し た こ と か ら 、 こ れ も ABCB1/P-gp を介した排泄トランスポーター阻害剤スクリーニングまたは評価 のために利用可能であると考えられた。
ヒトおよびカニクイザル腸管オルガノイドは多様な薬物動態関連遺伝子を発 現することに加えて、CYP3A4/8 活性を示し、この代謝活性は ketoconazole によ って顕著に阻害された (Figure 9a)。 また、CYP3A4/8 の代謝活性は、それぞれ
プレグナンX 受容体 (PXR) およびビタミン D 受容体 (VDR) を介して CYP3A4
を誘導することが知られている18, 19)。ヒト腸管オルガノイドではRIF および VD3
によって有意にCYP3A4 の代謝活性が誘導されたが (Figure 9b)、カニクイザル
ではVD3 では有意な誘導は確認できなかった (Figure 9b)。カニクイザル iPS 細
34 化として未だ不十分だと考えられた。また、ヒトiPS 細胞由来腸管オルガノイド でも、CYP3A4 代謝活性の誘導倍率は、2D 培養細胞および動物モデルなどの他 のモデルと比較して低かった。 特に、PXR および VDR の mRNA 発現レベル は、成人小腸におけるmRNA 発現レベルと比較して低かったことから(データは 示していない)、このことが、CYP3A4 誘導能が十分に検出されない要因である と考えられた。 したがって、小腸における薬物動態を正確に評価するために、 iPS 細胞由来腸管オルガノイドにおける CYP3A4 の代謝活性および誘導能を改 善する分化プロトコールをさらに、開発する必要がある。 腸管オルガノイドの内側は腸管刷子縁膜側であることから、腸管管腔側から の薬物吸収過程を簡便に評価するのは難しい。したがって、小腸における初回通 過効果を調べるためには、吸収過程を評価することができる新しい方法を開発 しなければならないと考える。また、腸内クリアランスを評価するためには、小 腸でのアベイラビリティーが既に知られている様々な医薬品を用いてin vivo と 腸管オルガノイドとの相関を明らかにする必要があると考えられる。特に、ヒト と薬物代謝酵素基質特異性が非常に関連しているカニクイザルは、薬物の薬物 動態学的特性を評価するため、非臨床の動物モデルとして創薬研究において使 用されている。カニクイザルの薬物動態学的パラメータは、マウス、ラット、お よびイヌのそれらと比較して、ヒトにより類似しているとの報告がある 25, 26)。
また、Kim et al.は、カニクイザル in vitro および in vivo 両方の CYP3A 誘導デー
タから、ヒトCYP3A4 の in vitro および in vivo での誘導を予測することができ
ると報告している 40)。したがって、カニクイザルのin vitro 小腸モデルを作製す ることは重要であると考える。したがって、カニクイザルのin vitro 小腸モデル を作製することは重要であると考える。また、近年、in vitro データからヒト in vivo の動態予測を行うことが可能となり、最近では、その外挿の精度が向上し、 医薬品開発に活用されている。カニクイザルでは、非臨床試験段階でin vivo と in vitro の両方のデータを入手することが可能であることから、数学的モデリン グシミュレーション技術等を用いてin vivo-in vitro 相関パラメータを正確に評価 することが可能である。そのため、ヒトin vitro から in vivo を正確に予測するた めに、カニクイザルでのin vivo-in vitro 相関パラメータがヒトへと外挿可能か検
35 証する必要がある。しかしながら、カニクイザルでは、ヒトと遺伝的に類似して いるにもかかわらず、いくつかの医薬品の経口バイオアベイラビリティは、ヒト での値よりも低いことが報告されている 28, 29)。このことから、カニクイザルと ヒトとの間にはさまざまな種差があるため、カニクイザル実験データを直接外 挿して、ヒトにおける体内動態を予測することは困難であると考えられる。した
がって、将来的には、ヒトin vitro とカニクイザル in vivo-in vitro の実験データか
ら種差等による薬物動態プロファイルの違いをスケーリングファクターを用い て補正することなどによって、カニクイザルにおけるin vivo-in vitro 相関をヒト in vivo-in vitro 相関に外挿できるシステムを構築することが望まれる。現在のと ころ、腸管オルガノイドを用いて、薬物の小腸でのアベイラビリティーを予測す ることは困難であるが、ミニチュア化した Ussing チャンバーを利用すること、 また、上皮細胞を単離し、セルカルチャーインサート上に播種することで、薬物 が代謝酵素やトランスポーターの基質、阻害剤、誘導剤となるかどうか、小腸で 生成した代謝物がこれらの阻害剤となるかどうかなどを判断することは可能だ と考えられる。 ヒトおよびカニクイザル iPS 細胞由来の腸管オルガノイドは、小腸に存在す る様々な細胞 (腸細胞、腸幹細胞、杯細胞、腸内分泌細胞、パネート細胞、およ び間葉細胞) を含んでいた。特に、間葉系組織は腸幹細胞の維持および自己再生 に重要である 41)。さらに、間葉系組織はクローン病において、組織の線維化に 関与していることが報告されている 42, 43)。ヒトおよびカニクイザル iPS 細胞由 来腸オルガノイドは腸上皮細胞および間葉系細胞を含むので、ヒト腸管線維症 のin vitro モデルとして使用することができる可能性が考えられる。 iPS 細胞由来腸管オルガノイドが IBD の細胞移植療法の開発に役立つことも 期待されている。 iPS 細胞由来腸管細胞は、粘膜修復のための自家腸管上皮細 胞の潜在的な供給源であり、そして他の細胞を超える大きな利点として、免疫拒 絶を避けられる。 いくつかの研究では、IBD における移植治療のため腸管オル
ガノイドの利用が検証されている 7)。 さらに、Shiba et al. はカニクイザル iPS
細胞の同種異系移植はカニクイザル心筋梗塞モデルにおける心臓の収縮性を改
36
ノイドは、ヒトにおける同種異系細胞移植のための重要な研究ツールとなると 考えられる。
37 2.5 小括 本研究では、EZSPHERE plate と浮遊培養により、腸管オルガノイドの大量培 養に成功した。また、ヒトおよびカニクイザルiPS 細胞から腸管オルガノイドへ の分化促進に有効な低分子化合物の組み合わせを新たに見出すことができた。 さらに、この方法によって作製した腸管オルガノイドは、機能的なタイトジャン クション、排出トランスポーターの輸送機能に加え、薬物代謝酵素活性および誘 導能を有するなど、腸管に特徴的な様々な薬物動態学的機能を有していること が明らかとなった。
38 第三章 多能性幹細胞由来腸管オルガノイドを用いた炎症性腸疾患モデル系の 構築 3.1 諸言 クローン病および潰瘍性大腸炎に代表される慢性疾患である IBD は世界中で 患者数が増加している 3)。 IBD は、免疫細胞の過活性化およびサイトカインの 過剰な分泌と関連している45, 46)。 特に、クローン病において、異常に活性化さ れたマクロファージによって産生される腫瘍壊死因子-α (TNF-α) のような炎症 性サイトカインは、腸上皮細胞の損傷を引き起こす7, 47)。腸内細菌によるその後 の侵襲に関連した免疫応答は、疾患の進行をさらに加速する 48)。実際に、上皮 細胞の破壊は、現在、この疾患の重要な特徴と考えられている49)。近年、抗
TNF-α 抗体である infliximab (IFX) は、活動期にある IBD 患者の腸炎症および上皮損
傷を軽減するということが報告されている 50)。しかし、腸内の創傷治癒を促進 する薬剤は上市されておらず、その開発が急務となっている 7)。 慢性炎症による腸管組織の線維症も、一般的なIBD 合併症である51, 52)。トラ ンスフォーミング成長因子-β (TGF-β) は、重要な線維化に関与するサイトカイ ンであり43, 53)、上皮間葉転換 (EMT) を促進させる43, 52)。実際、TGF-β は EMT を引き起こし、コラーゲンに富んだ細胞外マトリックスを産生する活性化筋線
維芽細胞を生成する54)。EMT の間、上皮細胞は vimentin および α-SMA のよう
な間葉細胞マーカーを獲得し、上皮細胞マーカーを失うことで、さらに、コラー ゲン産生性線維芽細胞・筋線維芽細胞へ移行することが知られている 55)。最終 的に、過剰な線維化は線維性狭窄症をもたらし、生活の質を損なうばかりでなく、 外科的手術を必要とする 56)。従って、創傷治癒を促進するための治療薬を開発 するため、腸の線維症の適切な実験モデルも必要である。 現在の IBD 粘膜損傷および線維症のモデル、特に、薬物スクリーニングにお いてはさまざまな問題点が存在する。例えば、Caco-2、T-84、および CCD-18co 細胞などのin vitro モデルは、複雑な腸環境を模倣していない57, 58)。一方、in vivo 動物モデルでは、ヒトの疾患を完全には再現できず、さらに、医薬品開発におけ る大規模スクリーニングには制限がある 59)。したがって、前述したように、近
39 年、腸管オルガノイドは、腸疾患の研究および治療のための重要なツールとして 非常に期待されている5, 60, 61)。特に、侵襲性が低く、病態モデルとしても構築可 能なヒトiPS 細胞由来腸管オルガノイド (HiOs) が、注目されている8, 10, 32)。例 えば、Rodansky et al.は、ヒト ES 細胞由来腸管オルガノイドを用いて、腸線維症 の新しいモデルを作製した 59)。この腸管オルガノイドでは、筋線維芽細胞と多 様な腸上皮細胞が含まれているが、Matrigel 中に包埋培養されているため、ハイ スループットの薬物スクリーニングへの応用は困難である32)。 本研究では、HiOs を用いて、TNF-α および TGF-β の処理により IBD モデルを 作製し、その有用性を検証した。さらに、薬剤スクリーニングおよびハイスルー プットスクリーニングに応用するために、前述のように作製した浮遊培養によ る腸管オルガノイドを使用した。
40 3.2 実験方法
3.2.1 試薬
Activin A、FGF2 は PeproTech 株式会社 (Rocky Hill, NJ, USA) より、BD Matrigel matrix growth factor reduced Matrigel 、 40-μm nylon-mesh cell strainer は BD Biosciences 株式会社 (Bedford, MA, USA) より、6-[[2-[[4-(2,4-dichlorophenyl)-5-(5-methyl-1H-imidazol-2-yl)-2-pyrimidinyl]amino]ethyl]amino]-3-pyridinecarbonitrile (CHIR99021) 、 (+)-(R)-trans-4-(1-aminoethyl)-N-(4-pyridyl) cyclohexanecarboxamide dihydrochloride (Y-27632) は Focus Biomolecules (Plymouth Meeting, PA, USA) より 、 FGF4、TNF-α は BioLegend (San Diego, CA, USA) より、EZSPHERE (#900:
microwell size: diameter 500 μm, depth 100 μm) は、AGC テクノガラス株式会社 (Shizuoka, Japan) より、KSR、Advanced-DMEM/F12、N2 supplement、B27 serum-free supplement、SlowFade Diamond antifade mountant は、Thermo Fischer Scientific
社 (Waltham, MA, USA) よ FBS は、ニチレイバイオサイエンス社 (Tokyo, Japan)
より、R-spondin 1、Noggin、EGF は GenScript (Piscataway, NJ) より、StemSure® hPSC medium、SB431542 は、富士フィルム和光純薬株式会社 (Osaka, Japan) よ り、2-(2-amino-3-methoxyphenyl)4H-1-benzopyran-4-on (PD98059)、3-(6-methyl-2-pyridinyl)-N-phenyl-4-(4-quinolinyl)-1H-pyrazole-1-carbothioamide (A-83-01) は 、 AdooQ BIOSCIENCE (Irvine, CA, USA)より、5-aza-2’-deoxycytidine は、Chem-impex international 社 (Wood Dale, IL, USA) より、IFX は Novus Biologicals (Littleton, CO, USA) より、TGF-β1, SinoBiological 株式会社 (Beijing, China) より、4-[4-(1,3-benzodioxol-5-yl)-5-(2-pyridinyl)-1H-imidazol-2-yl]-benzamide (SB431542) は Cayman Chemical 社 (Ann Arbor, MI, USA) より、OCT compound は、サクラファ インテックジャパン株式会社 (Tokyo, Japan) より、FD-4 は、Sigma-Aldrich (St. Louis, MO, USA) より、total RNA from human small intestine および colon samples (それぞれ 5 donors) は、BioChain Institute 株式会社 (Newark, CA, USA) より購入 して用いた。その他の試薬は全て市販の高速液体クロマトグラフ用もしくは特 級品を用いた。
41 ヒト iPS 細胞 (#51:Windy) は、2.2.2 で述べたように国立成育医療研究セン ター研究所の梅澤 明弘博士らにより、ご供与いただいた.未分化のヒト iPS 細 胞の維持培養はMEF 上で行い、2.2.2 で述べた方法と同様に行った。 3.2.3 ヒト iPS 細胞から腸管オルガノイドへの分化 ヒトiPS 細胞から腸細胞への分化は 2.2.3 で述べた方法と同様に行った。簡単 に述べると、 ヒト iPS 細胞を 100 ng/mL activin A によって 72 時間処理する ことによって内胚葉へ分化させた。次に内胚葉様細胞に 3 µM CHIR99021、500 ng/mL FGF4 を 96 時間処理することによって中・後腸へと分化させた。次に、 40 µm ナイロンメッシュのセルストレーナーにて細胞塊を砕き、3.0×106細胞を
100 mm EZSPHERE®上に継代し、100 ng/mL EGF、100 ng/mL Noggin、200 ng/mL
R-spondin-1、10 µM Y-27632 を含む Advanced-DMEM/F12 で 3 日間培養した。そ
の後、100 ng/mL EGF、100 ng/mL Noggin、200 ng/mL R-spondin-1、Matrigel 3%を
含むAdvanced-DMEM/F12 で 24 日間超低接着 10 cm dish 上で浮遊培養すること
で腸管オルガノイドへ分化させた。また、20 μM PD98059、5 μM
5-aza-2’-deoxycytidine、および 0.5 μM A-83-01 を分化開始 19 日目 から 34 日目まで添加
した。培地は3 日おきに交換した。
3.2.4 粘膜傷害モデルの作製
分化誘導34 日終了後、2 mM L-Glu、2% B27 supplement、1% N2 supplement、
100 units/mL ペニシリン G、100 µg/mL ストレプトマイシン、100 ng/mL EGF、 100 ng/mL Noggin、200 ng/mL R-spondin-1、Matrigel 3%を含む Advanced-DMEM/F12
で3 日間浮遊培養した。その後、粘膜傷害を評価するため TNF-α (30 ng/mL) を
96 時間処理した。必要に応じて、抗 TNF-α 抗体 (infliximab, 1 µg/mL) を同時に 添加した。培地は毎日交換した。
3.2.5 線維化モデルの作製
分化誘導34 日終了後、2 mM L-Glu、2% B27 supplement、1% N2 supplement、
42
100 ng/mL Noggin、200 ng/mL R-spondin-1、Matrigel 3%を含む Advanced-DMEM/F12
で3 日間浮遊培養した。その後、線維化モデルを構築するため、TNF-α (30 ng/mL)
と TGF-β (2 ng/mL) を 48 時間処理した。必要に応じて TGF-β 阻害剤である
SB431542 (SB, 20 µM) を同時に添加した。培地は毎日交換した。
3.2.6 総リボ核酸 (RNA) 抽出
総RNA はヒト iPS 細胞から腸管オルガノイドへの分化誘導終了後、Agencourt
RNAdvence Tissue の添付マニュアルに従い抽出した。
3.2.7 逆転写反応
相補的DNA (cDNA) の合成は、ReverTra Ace qPCR RT Master Mix を使用し、
添付マニュアルに従い行った。
3.2.8 Real-Time PCR 法
PCR プライマーは、Table 4 に示したものを用いた。Real-Time PCR は KAPA SYBR Fast qPCR Kit を用い、cDNA を鋳型にして、反応は添付マニュアルに従い
43
Table 4 Primers used for real-time PCR.
Gene name Sense (5'→3') Antisense (5'→3') Product length (bp)
α-SMA GCCTGAGGGAAGGTCCTAAC CAGAGCCATTGTCACACACC 227
APOB TTGCTGAAGAAAACCAAGAACTC CCCTCTTGATGTTCAGGATGTAA 142
APOC3 CTCAGCTTCATGCAGGGTTAC TAACGGTGCTCCAGTAGTCTTTC 148
Aquaporin 3 GATCAAGCTGCCCATCTACACC CCATTGATCATATCCAAGTGTCCAG 186
CDX2 ACCTGTGCGAGTGGATGC TCCTTTGCTCTGCGGTTCT 232
Chromogranin A TCCGACACACTTTCCAAGCC TTCTGCTGATGTGCCCTCTC 164 Collagen type 1 AAGAGGAAGGCCAAGTCGAG AGATCACGTCATCGCACAAC 155 Fibronectin TATGTGGTCGGAGAAACGTG TCCTTGTGTCCTGATCGTTG 129
HPRT CTTTGCTTTCCTTGGTCAGG TCAAGGGCATATCCTACAACA 148
IL-1β GTGGCAATGAGGATGACTTGTTC TAGTGGTGGTCGGAGATTCGTA 124
LGR5 TGCTCTTCACCAACTGCATC CTCAGGCTCACCAGATCCTC 193
Lysozyme TCAATAGCCGCTACTGGTGT AATGCCTTGTGGATCACGGA 143
MUC2 AGAAGGCACCGTATATGACGAC CAGCGTTACAGACACACTGCTC 137
MUC5B GTTCCACCCGTCACTGTCTT TCAGAGAACACGTAGTTGCAAAG 149
OLFM4 CAGACACCACCTTTCCCGTG CCTTCTCCATGATGTCAATTCGG 171
SATB2 AACTGCTCAAAGAGATGAACCAG ATGGCCCTCAGGTTTACTAGAAG 136
TNF-α GGCAGTCAGATCATCTTCTCG GCTGGTTATCTCTCAGCTCCAC 146
Villin AGCCAGATCACTGCTGAGGT TGGACAGGTGTTCCTCCTTC 169
44 3.2.9 免疫蛍光染色 分化誘導終了後、腸管オルガノイドを 4%パラホルムアルデヒドにて固定し、 OCT compound にて凍結包埋した。厚さ 10 µm の凍結切片を作製後、スライドガ ラスに貼り付け、抗原の賦活化を行った。5% FBS 溶液にて 30 分間ブロッキン グし、一次抗体を4℃で 1 晩反応させた。その後、スライドガラスを洗浄し、二 次抗体を室温で 1 時間反応させ、核染色として DAPI を用いた。用いた抗体は Table 5 に示した。封入作業を行い、Zeiss LSM510 共焦点レーザー顕微鏡を用い て、蛍光を観察した。
Table 5 Primary and secondary antibodies.
Antibody name Source Catalog number Biological source Dilution
α-SMA Abcam ab5694 Rabbit 1:200
Caspase-3 Cell Signaling Technology 45 Rabbit 1:100
Chromogranin A IMMUNOSTAR 20085 Rabbit 1:500
CDX2 BioGenex MU392A-UC Mouse 1:100
E-Cadherin Transduction BD
Laboratories™ 610181 Mouse 1:100
LGR5 ABGENT AP2745d Rabbit 1:100
Lysozyme BioGenex AR024-5R Rabbit ―
MUC2 Santa Cruz sc-15334 Rabbit 1:200
Occludin Thermo Fisher Scientific 71-1500 Rabbit 1:100
Villin Santa Cruz sc58897 Mouse 1:100
Vimentin Abcam ab8069 Mouse 1:200
ZO-1 Thermo Fisher Scientific RF236801 Mouse 1:100
Alexa Fluor® 488 donkey anti-Rabbit IgG
(H&L)
Thermo Fisher
Scientific A-21206 Rabbit 1:200
Alexa Fluor® 568 goat
anti-Mouse IgG (H&L) Thermo Fisher Scientific A-11004 Mouse 1:200 3.2.10 FD-4 の透過実験