論文題目
赤ワイン醸造においてブドウ由来不溶性化合物が
フェノール化合物の動態に与える影響
山梨大学大学院
医学工学総合教育部
博士課程学位論文
2020 年 3 月
井上絵梨
目次
第一章 緒言 1 第二章 赤ワイン醸造時におけるフェノール化合物の抽出 4 第一節 試料と方法 4 1. 試料ブドウ 4 2. 試薬 5 3. 機器 5 4. ワイン醸造試験 6 5. 醸造中におけるマストのサンプリング 6 6. 醸造中における分析 6 6-1 アルコール濃度分析 6 6-2 ワインカラー分析 6 6-3 全フェノール濃度(フォーリン・シオカルト法) 7 6-4 BSA 結合性タンニン濃度(BSA 沈殿法) 7 第二節 結果および考察 9 第三章 赤ワイン醸造時のマスト液体および果皮中のフェノール化合物濃度の変化 14 第一節 試料と方法 14 1. 試料ブドウ 14 2. 試薬 14 3. 機器 15 4. ワイン醸造試験 15 5. 醸造中のマスト液体およびブドウ果皮のサンプリング 16 6. ブドウ果皮のアントシアニン抽出 16 7. 醸造中における分析 17 7-1 pH,糖度(Brix),アルコール濃度分析 17 7-2 ワインカラー分析 177-3 全フェノール濃度(フォーリン・シオカルト法) 18 7-4 BSA 結合性タンニン濃度(BSA 沈殿法) 18 7-5 HPLC によるアントシアニンの分析 19 第二節 結果および考察 20 1. マスト液体中の pH,糖度(Brix),アルコール濃度および色調の推移 20 2. 醸造中におけるマスト液体部分および果皮中の全フェノール量(TP) およびBSA 結合性タンニン量(PA)の推移 25 3. 醸造中におけるマスト液体部分および果皮中のアントシアニン量の推移 29 4. 醸造試験まとめ 35 第四章 ブドウ由来不溶性化合物へのフェノール化合物の吸着特性 36 第一節 試料と方法 36 1. 試料ブドウ 36 2. 試薬 37 3. 機器 37 4. 分析方法 38 4-1 ブドウ果皮由来フェノール化合物の抽出および分画 38 4-2 ブドウ果皮・果肉・種子由来不溶性化合物(IC)の調製 39 4-3 各種モデルワインの調製 41 4-4 全フェノール濃度(フォーリン・シオカルト法) 41 4-5 BSA 結合性タンニン濃度(BSA 沈殿法) 41 4-6 HPLC によるアントシアニンの分析 41 4-7 ブドウ果皮・果肉・種子由来 IC へのフェノール化合物の吸着実験 42 第二節 結果と考察 43 1. ブドウ果実からの不溶性化合物の回収率 43 2. ブドウ由来 IC のアントシアニン吸着能 44 3. ブドウ由来 IC のタンニン吸着能 46 4. 果皮 IC へのアントシアニン吸着に及ぼす pH とアルコール濃度の影響 47 5. 果皮 IC へのタンニン吸着に及ぼす pH とアルコール濃度の影響 49
第五章 総合考察 50
参考文献 53
既発表論文 58
1
第一章 緒言
フェノール化合物は赤ワインにおいて重要であり,赤ワイン1 L あたり約 3000 mg ほ ど含まれ,色調や味わいなどに影響を与えている(Sacchi et al., 2005).赤ワイン中のフ ェノール化合物は,アントシアニン,プロアントシアニジン(Proanthocyanidin: PA),非 フラボノイド,フラボノイドから構成される.Vitis. vinifera の赤ワイン用ブドウ品種に おいては,フェノール化合物は,果皮に50%,種子に 44%,その他果肉などに 6%ずつ 分布していると報告されている(Singleton and Esau, 1969).アントシアニンは赤ワイン の色を構成する主要な化合物だが,不安定な構造であるため,容易に色調が変化したり, 無色になったりする.また,PA は通称タンニンと呼ばれており,ワインの収斂味や色 調安定化に寄与することが知られている(Jackson, 2014).アントシアニン類に由来する色の濃さやPA の濃度は,ワインの価格に非常に強い正 の相関関係を有することが報告されており,色が濃い,あるいはPA 濃度の高いワイン は,高品質な赤ワインとして評価されている(Kassara and Kennedy, 2011).従って色が 濃く,PA 濃度の高いワインの製造が望まれる。実際,赤ワイン醸造において色を濃く し,PA 濃度を高める様々な手法が開発されており,例えばマセレーションの延長,サ ーモ・ヴィニフィケーションやコールドマセレーションなどが挙げられる(Sacchi and Adams, 2005).また,ブドウを破砕,搾汁後に果汁を取り除くことによりフェノール化 合物の供給源である果皮や種子の果汁に対する割合を高め,ワインのアントシアニン濃 度やPA 濃度を高くする,セニエという醸造方法も存在する. さらに, PA については含有量だけでなく,質的な違いについても注目が集まってい る(Cheynier et al., 2006).醸造では,PA はブドウ果皮および種子から抽出され,それ ぞれが構造的な特徴を持っている.PA はカテキン類が重合したもので,果皮由来の PA は,カテキン,エピカテキン,エピカテキンガレート,エピガロカテキンから構成され, 種子由来のPA は,カテキン,エピカテキン,エピカテキンガレートで構成されている. そして,ワイン中の果皮由来のエピガロカテキンと種子由来のエピカテキンガレートの 割合が,収斂味の質の違いをもたらすことが知られており,エピカテキンガレートの割 合が高くなると収斂味の荒々しさが増し,エピガロカテキンが含まれると収斂味のなめ らかさが増すことが報告された(Cheynier, 2005; Cheynier et al., 2006).また,果皮由来
2
のフェノール化合物濃度が高い赤ワインは,なめらかな収斂味を備えた高品質なワイン であると消費者に認識されており,高価格で売買されている(Kassara and Kennedy, 2011).
このように,ブドウやワインに含まれるフェノール化合物に対する関心は高く,その 関心は赤ワイン醸造におけるフェノール化合物の抽出にも及ぶ.フェノール化合物は, 醸造中にブドウ果皮と種子が果汁と接触した状態で,アルコール発酵が進むにつれて抽 出される.アントシアニンは果皮から抽出されると,その構造の不安定さから,発酵中 の果汁のアルコール濃度,pH,温度などに容易に影響を受ける(Sacchi et al., 2005).PA は果皮と種子両方から抽出され,発酵初期は果皮PA の割合が高いが,発酵後期になる と果皮 PA の割合が低くなり,種子 PA の割合が高くなることが知られている(Peyrot des Gachons and Kennedy, 2003; Koyama et al., 2007).しかしながら,醸造中にアントシ アニン類やPA 抽出がどのように起こり,赤ワインになるのかは未だ明らかにされてお らず,醸造によるフェノール化合物の抽出には未解明の部分が多い. ブドウのフェノール化合物は通常,果皮および種子細胞の液胞に存在し,赤ワイン醸 造時には,抽出中にフェノール化合物に対して化学的または物理的な親和性を有する物 質を避けつつ,液胞から液体部分に移動する(Adams, 2006).フェノール化合物は,他 の化合物と結合しやすいことが知られており,例えばPA はタンパク質や多糖類と強い 結合を生じることが知られている(Jackson, 2014).ブドウに含まれるフェノール化合物 のうち,細胞壁由来のタンパク質との親和性が高いフェノール化合物は強く結合し,醸 造時のフェノール化合物の抽出性が低下することが報告された(Springer et al., 2014). Garrido- Bañuelos et al. (2019)は,果皮細胞壁組成をビンテージや果実の熟度から比較 し,細胞壁の構成要素の一つであるペクチン含有率が,フェノール化合物の抽出率に影 響を与えることを示した.また,細胞壁等に存在する空隙の大きさや網目構造もフェノ ール化合物の吸着に大きな影響を与えることが報告された(Bindon et al., 2012). さらに,Bindon et al. (2010) は,細胞壁物質へのタンニンの吸着および脱着の抽出 モデルの仮説を提唱し,果皮や種子からマストの液体部分に抽出されたフェノール化合 物が,マスト中に存在する果皮などの不溶性細胞壁物質に再吸着され,液体部分から除 去されている可能性を示唆した.また,アントシアニン類がセルロースやセルロース・ ペクチン複合体とイオン性相互作用や疎水性相互作用によって結合していることや (Padayachee et al., 2012),細胞壁物質とタンニンやアントシアニン類が結合親和性を有
3 することが明らかになった.よって,フェノール化合物とブドウ細胞壁物質との親和性 は,ワイン醸造におけるフェノール化合物の動態に影響を与える要因となる可能性が考 えられる. したがって,赤ワイン醸造中におけるフェノール化合物が果皮および種子から液体部 分にどのように抽出され,マスト中に存在する果皮などの不溶性細胞壁物質に再吸着が 起きているかを明らかにし,未解明の部分が多い醸造中のフェノール化合物の動態を明 らかにすることを目的として本研究を行った.
4
第二章 赤ワイン醸造時におけるフェノール化合物の抽出
日本で醸造されたカベルネ・ソーヴィニヨン種(CS)やメルロ種ワインのフェノール 化合物濃度は,アメリカなどで醸造されたワインよりも低く(Harbertson et al., 2008; Ichikawa et al., 2011),また,日本で最も多く生産されているマスカット・ベーリーA (MBA)種の赤ワインは,タンニン濃度が非常に低い(平均 65 mg/L)(Ichikawa et al., 2011)ことが示された.さらに,MBA では醸造中にフェノール化合物濃度が一度上昇 した後,低下する現象が認められた.一方CS では前述のような現象は認められなかっ た.赤ワイン醸造では,主にブドウ果皮および種子からフェノール化合物が抽出される. ワイン中に含まれる果皮および種子のフェノール化合物のおおよその割合は知られて いるが,醸造中におけるそれらの量や割合がどのように変化するのかは明らかにされて いない.そこで,本醸造試験ではMBA および CS の果皮と種子を別々に発酵させるこ とによって,フェノール化合物が抽出されるタイミングとブドウの部位を明らかにし, 醸造中におけるブドウ果実と液体部分でのフェノール化合物の動態を明らかにするこ とを本章の目的とした.
第一節 試料と方法
1. 試料ブドウ 試料ブドウには,山梨大学ワイン科学研究センター付設育種試験地で収穫された赤ブ ドウ2 品種を供試した(表 2-1). 表2-1 試料ブドウ 品種 収穫日 糖度(Brix) Cabernet Sauvignon (CS) 2012.10.16 21.45 2. 試薬
塩酸 特級 富士フイルム和光純薬株式会社
炭酸ナトリウム 特級 富士フイルム和光純薬株式会社
フォーリン・シオカルト試薬(F-C 試薬) 富士フイルム和光純薬株式会社 没食子酸 Extra Pure Reagent ナカライテクス株式会社
カテキン 食品分析用 富士フイルム和光純薬株式会社 牛血清アルブミン,コーンフラクションV,pH 7.0 生化学用 富士フイルム和光純薬株式会社 酢酸 特級 富士フイルム和光純薬株式会社 塩化ナトリウム 特級 関東化学株式会社 水酸化ナトリウム 特級 富士フイルム和光純薬株式会社 トリエタノールアミン(TEA) 特級 富士フイルム和光純薬株式会社 ドデシル硫酸ナトリウム(SDS) 一級 富士フイルム和光純薬株式会社 塩化第二鉄六水和物(FCR) 特級 富士フイルム和光純薬株式会社 ワイン醸造用乾燥酵母(LALVIN EC1118) Lallemand
メタ重亜硫酸カリウム 食品添加物 富士フイルム和光純薬株式会社 ドライアイス 3. 機器 簡易型アルコール分析器 理研機器 AL-3 型アルコメイト 糖度計 手持屈折計 ATAGO N1 電子天秤 SHIMAZU AUW-220D 遠心分離機 HITACHI CF-16-RN
HPLC JASCO MD-2018 PLUS PHOTODIODE ARRAY DETECTOR 吸光光度計 HITACHI SPECTROPHOTOMETER U-2900
試験管ミキサー TAITEC CO., LTD SE-04
6 4. ワイン醸造試験(Sampaio et al., 2007) 試験醸造に用いたブドウは収穫後,手で除梗を行い果実が均一になるように撹拌した. その後,3.5 kg ずつ 5 L 容ガラス製梅酒瓶に除梗した果実を分け,ピロ亜硫酸カリウム をSO2濃度で70 mg/kg になるように添加した.その後,コントロールサンプルは破砕 を行った.また,果皮と果肉を発酵させるサンプルは種子を除去し,種子と果肉を発酵 させるサンプルは果皮を除去した.この過程での酸化を極力防ぐために,適宜ドライア イスを添加しながら処理を行った.
MBA は糖度が 17.2°Brix だったので,補糖を行い 21° Brix に調整した.発酵 1 日目に 酒母としてLALVIN EC1118(Lallemand 社)を果汁に 200 mg/kg となるように添加した. また,パンチダウン法と同様の効果を得るために,朝と夕の1 日 2 回梅酒瓶を回し,内 容物を混合した.その際,酸化を防ぐために少量のドライアイスを添加した.各試験醸 造は品種および醸造方法(コントロール,果皮発酵,種子発酵)ごとに3つずつ独立系 で実施した. 5. 醸造中におけるマストのサンプリング サンプリングは発酵期間中の毎日午前9 時行った.梅酒瓶にドライアイスを入れ,ヘ ッドスペースを炭酸ガスで充満させた後に,20 mL 容遠心管にマストを 15 mL 採取し た.サンプリング後,直ちに20℃で 15,000 x g で 5 分間遠心分離を行った.そこで得ら れた上清を別の容器に移し,それをマストのサンプル試料とした. 6. 醸造中における分析 6-1 アルコール濃度分析 簡易型アルコール分析器であるアルコメイトを用いて,発酵中のマストのアルコール 濃度を3 回ずつ測定した. 6-2 ワインカラー分析(Ribéreau-Gayon et al., 2006) ① 光路長2 mm のセルを用い,逆浸透水を対照としてワインの 420,520,620 nm の 吸光度を測定した. ② 得られた値を5 倍し,A420,A520,A620 とした.また,下記の計算式より Color
7 intensity を算出した.
Color intensity = A420 + A520 + A620
6-3 全フェノール濃度(Total Phenol: TP)(フォーリン・シオカルト法)(Singleton and Rossi, 1965) A) 試薬の調製 ・ F-C 試薬(1/2 に希釈) ・ 炭酸ナトリウム溶液 炭酸ナトリウム100 g を 500 mL の逆浸透水に加え,沸騰させて溶解した.室温に冷 やし,一晩放置後濾過をして使用した. ・ 没食子酸標準水溶液(0~1000 mg/L:検量線用) 没食子酸100 mg を逆浸透水で 100 mL にメスアップした(1000 mg/L).これを希釈 し,それぞれ0,100,200,400,600,800,1000 mg/L となるように調製した. B) 操作 ① 各試験管に没食子酸標準溶液または赤ワイン試料を100 µL とり,逆浸透水 7.9 mL を添加し,攪拌した.試験管は,すべて同じものに揃えた. ② F-C 試薬 0.5 mL を加え,すぐに攪拌した. ③ 30 秒以上 8 分以内に炭酸ナトリウム溶液を 1.5 mL 加え,攪拌した. ④ 室温で2 時間静置後,765 nm の吸光度を測定した. ⑤ 没食子酸標準液の検量線より,全フェノール化合物濃度を算出した.
6-4 BSA 結合性タンニン(Proanthocyanidin: PA)濃度(BSA 沈殿法) (Harbertson et al., 2002) A) 試薬の調製 ・ Wash Buffer(200 mM 酢酸,170 mM 塩化ナトリウム, pH 4.9) ・ 900 mL の逆浸透水に 11.45 mL の酢酸を入れ,9.93 g の塩化ナトリウムを添加し, 攪拌し溶解した.水酸化ナトリウムで pH 4.9 となるように調整した後,逆浸透水 で1000 mL にメスアップした.
8
・ Protein Solution(1 mg/mL 牛血清アルブミン(BSA),200 mM 酢酸,170 mM 塩 化ナトリウム, pH 4.9)
(標準)500 mL の Wash Buffer に 0.5 g の牛血清アルブミンを添加し,攪拌溶解した. ・ Resuspension Buffer(5% トリエタノールアミン,10% ドデシル硫酸ナトリウム) 475 mL の Wash Buffer に 25 mL のトリエタノールアミンを加え,攪拌し溶解し た.ドデシル硫酸ナトリウム 50 g を少しずつ加え,攪拌して溶解した.
・ Ferric Chloride Reagent(10 mM 塩化第二鉄六水和物)
100 mL の 0.01N 塩酸に 0.2703 g の塩化第二鉄六水和物を加え,攪拌し溶解した. ・ 0.01N HCl 99 mL の逆浸透水に 1 mL の 1N 塩酸を加え,混合した. ・ カテキン標準水溶液(0~300 mg/L:検量線用) カテキン30 mg を逆浸透水で 100 mL にメスアップした(300 mg/L).これを希釈 し,それぞれ0,50,100,150, 200,250,300 mg/L となるように調整した. B) 操作 ① 1.5 mL 容のマイクロチューブに試料を 500 µL 取り,Protein Solution を 1.0 mL 加え て,静かに攪拌し,室温で15 分間静置した. ② 13,500×g(13,000 rpm)で 5 分間,遠心分離を行い,沈殿を得た. ③ 上清を捨て,Wash Buffer を 250 µL 入れ,13,500×g(13,000 rpm)で 1 分間,遠心分 離を行った. ④ 上清を捨て,Resuspension Buffer を 875 µL 加え,室温で 10 分間静置後,攪拌し, 沈殿を可溶化させた. ⑤ ④を光路長10 mm の石英セルに入れ,510 nm の吸光度を測定し,セルから溶液を マイクロチューブに戻した.この測定値をブランクとした.
⑥ 溶液を戻したマイクロチューブにFerric Chloride Reagent を 125 µL 加え,攪拌した. プロアントシアニジンを発色させるため室温で10 分間静置した.
⑦ ⑥を光路長10 mm の石英セルに入れ,510 nm の吸光度を測定した.⑤におけるブ ランクを差し引き,カテキン標準液の検量線より,プロアントシアニジン濃度を算 出した.
9
第二節 結果および考察
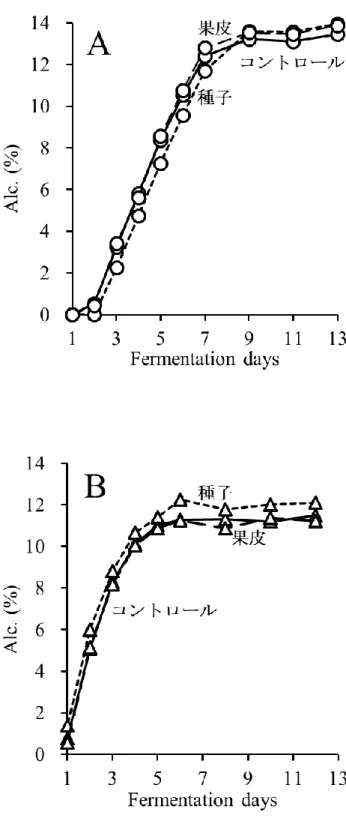
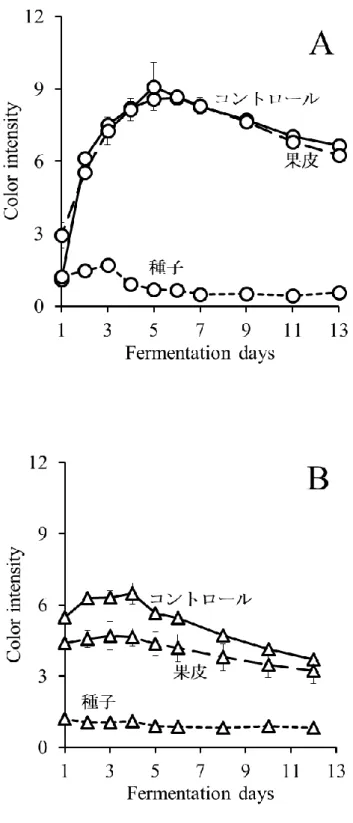
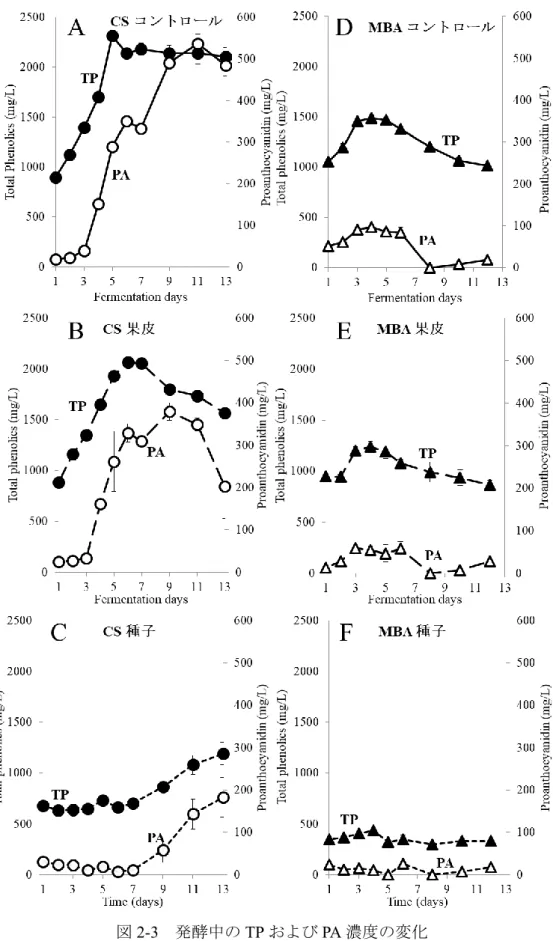
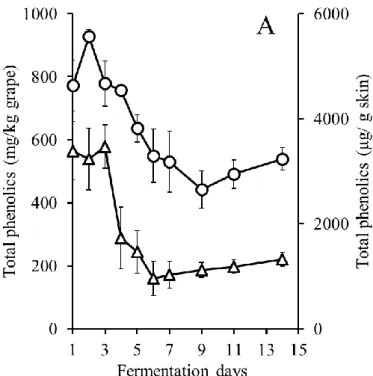
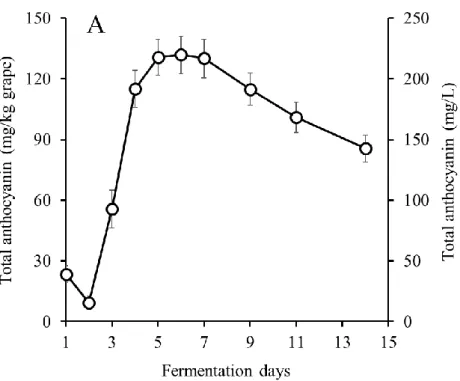
MBA において発酵後期に TP 濃度が低下することや,ワイン中の PA 濃度が顕著に低 い原因を明らかにするために,果皮と種子をそれぞれ別に発酵させる醸造試験を行った. MBA では CS よりもアルコール発酵が速く進んだが,それぞれ 3 種類の醸造方法(コ ントロール発酵,果皮発酵,種子発酵)間でほとんど差は認められず,最終的にアルコ ール濃度は約12%に達した(図 2-1-A,B).発酵期間中,コントロール発酵および果皮 発酵においてColor intensity は発酵 4〜6 日目まで増加し,その後両品種において減少し た(図2-2-A,B).この現象の一部は,コピグメンテーション効果であると考えられた (Boulton 2001).一方,種子発酵サンプルでは,アントシアニンを含む果皮が存在しな いため,両品種においてColor intensity は非常に低かった. コントロール発酵では,TP および PA 濃度は MBA で低く推移したが,両品種におい て発酵4~5 日目まで増加した(図 2-3-A,D).MBA では,PA 濃度は 4 日目にピーク (97 mg/L)に達し,その後 8 日目までに大きく減少し,この結果は,Ichikawa et al. (2012) を支持した.TP 濃度は発酵 4 日目に約 1490 mg/L に達したが,その後徐々に減少した. 一方,CS では,TP 濃度は発酵 5 日目に最大 2315 mg/L に達し,その後濃度を維持し平 衡状態となった.また,PA 濃度は,11 日目にピーク(536 mg/L)に達するまで増加し 続けた. 果皮発酵では,種子は取り除かれていたため,TP は果皮と果肉から抽出され,大部 分のTP は果皮由来だったと考えられた(図 2-3-B,E).MBA では,TP 濃度は発酵 4 日 目に1241 mg/L でピークに達し,その後徐々に減少した.PA 濃度は発酵 3 日目にピー ク(59 mg/L)に達し,その後 8 日目に 0 mg/L まで減少した.本実験における MBA の TP および PA 濃度は CS よりも低く,果皮から抽出されるフェノール化合物が,MBA において少なかったことが示された.CS でも,TP 濃度は発酵 6 日目まで増加したもの の,その後MBA 同様減少した.PA 濃度は 4 日目から増加し,9 日目にピーク(379 mg/L) に達し,11 日目に急激に減少した. 種子発酵では,果皮は取り除かれていたため,TP は種子と果肉から抽出され,大部 分のTP は種子由来であったと考えられた(図 2-3-C,F).MBA では,TP および PA 濃 度が低く,特にPA 濃度が非常に低かった.CS では,TP 濃度は発酵 7 日目から増加し10 始め,13 日目には 1191 mg/L に達した.同様に,PA 濃度も 9 日目から増加し,13 日目 には183 mg/L に達した.種子 PA が発酵後期に抽出される現象は,Koyama et al. (2007) の結果と一致した. 以上の結果より,果皮発酵および種子発酵のデータを組み合わせることで,コントロ ール発酵のTP および PA の挙動について説明が可能であった.また,MBA コントロー ル発酵終了時,ワイン中に含まれるTP の大部分は果皮に由来し,種子由来の TP は顕 著に少なかった.対照的に,CS では,コントロール発酵終了時,種子由来の TP および PA が,MBA と比較して多かった. CS ワイン中の種子 PA 濃度は,発酵中期から後期にかけて増加した.一方,果皮 PA 濃度は発酵 9 日目にピークに達し,その後減少した.従って,Peyrot Des Gachons and Kennedy (2003)でも報告されているように,果皮 PA/種子 PA 比は発酵期間に比例して減 少すると考えられる.また,果皮PA 濃度はワインの品質に密接な関係があると考えら れている(Kassara and Kennedy, 2011)ため,果皮 PA 濃度の低下を防ぐことは,ワイン 醸造において非常に重要な課題である.通常のワイン醸造では,発酵後期での果皮 PA が減少する時,種子PA が抽出され始めるため,この現象を検証することは困難であっ たが,本実験結果より,果皮PA 濃度を司る果帽管理と圧搾のタイミングは,ワインの 品質の向上を考える上で非常に重要であると考えられる. また,MBA では,果皮および種子(特に種子)から抽出された PA 濃度が全体的に低 く推移し,発酵7 日目に PA 濃度が低下し始めた.これらの結果は,MBA ワインの PA 濃度が低くなる要因であると考えられる.CS において,コントロール発酵では,TP お よびPA 濃度が連続的に増加したが,実際は果皮 TP および PA は発酵後期に減少してい ることが示された.この現象は,ワイン醸造時に発酵が進み,ブドウの細胞壁が崩れる ことによって,不溶性化合物がマスト中に露出し,発酵初期に抽出された果皮TP およ びPA が吸着するという仮説によって,一部は説明することができた(Bindon et al., 2010). さらに,より高品質な赤ワイン醸造のために,不溶性化合物への果皮TP の吸着の可能 性を含めた,果皮TP の抽出機構の解明が重要だと考えられる.
11
12
13
図2-3 発酵中の TP および PA 濃度の変化
14
第三章 赤ワイン醸造時のマスト液体および果皮中の
フェノール化合物濃度の変化
第二章では,赤ワイン醸造でCS および MBA において果皮由来フェノール化合物が 発酵後期で減少することが明らかにされた.また,果帽管理や圧搾時期がワインのフェ ノール化合物濃度に影響を与えることが報告された(Ichikawa et al., 2012).これらの現 象を説明する仮説として,Bindon et al. (2010) は,細胞壁物質へのタンニンの吸着およ び脱着の抽出モデルを提唱した.よって,果皮や種子からマストの液体部分に抽出され たフェノール化合物が,マスト中に存在する果皮などの不溶性化合物に再吸着し,液体 部分から除去されている可能性が示唆された.本章では,実際の赤ワイン醸造において, フェノール化合物がどのように抽出され,また果皮由来の不溶性化合物がどのような影 響を与えているのか明らかにすることを目的とした.第一節 試料と方法
1. 試料ブドウ 試料ブドウには,2015 年 9 月 24 日に山梨大学生命環境学部附属小曲農場(35°66'N; 138°58'E)で収穫された赤ブドウ 2 品種を供試した(表 3-1) 表3-1 試料ブドウ 品種 樹齢(年) 台木 糖度(Brix) Cabernet Sauvignon (CS) 7 Teleki 5BB 16.7Muscat Bailey A (MBA) 7 Teleki 5BB 21.8
2. 試薬
塩酸 特級 富士フイルム和光純薬株式会社
炭酸ナトリウム 特級 富士フイルム和光純薬株式会社
フォーリン・シオカルト試薬(F-C 試薬) 富士フイルム和光純薬株式会社 没食子酸 Extra Pure Reagent ナカライテクス株式会社
15 牛血清アルブミン,コーンフラクションV,pH 7.0 生化学用 富士フイルム和光純薬株式会社 酢酸 特級 富士フイルム和光純薬株式会社 塩化ナトリウム 特級 関東化学株式会社 水酸化ナトリウム 特級 富士フイルム和光純薬株式会社 トリエタノールアミン(TEA) 特級 富士フイルム和光純薬株式会社 ドデシル硫酸ナトリウム(SDS) 一級 富士フイルム和光純薬株式会社 塩化第二鉄六水和物(FCR) 特級 富士フイルム和光純薬株式会社 ギ酸 特級 富士フイルム和光純薬株式会社 メタノール 特級 富士フイルム和光純薬株式会社
ワイン醸造用乾燥酵母(LALVIN EC1118) Lallemand
メタ重亜硫酸カリウム 食品添加物 富士フイルム和光純薬株式会社 ドライアイス 3. 機器 簡易型アルコール分析器 理研機器 AL-3 型アルコメイト 糖度計 手持屈折計 ATAGO N1 電子天秤 SHIMAZU AUW-220D 遠心分離機 HITACHI CF-16-RN
HPLC JASCO MD-2018 PLUS PHOTODIODE ARRAY DETECTOR 吸光光度計 HITACHI SPECTROPHOTOMETER U-2900
試験管ミキサー TAITEC CO., LTD SE-04
pH メーター HORIBA pH/COND METER F-5
4. ワイン醸造試験(Sampaio et al., 2007)
試験醸造に用いたブドウは収穫後,手で除梗を行い果実が均一になるように撹拌した. その後,3.5 kg ずつ 5 L 容ガラス製梅酒瓶に除梗した果実を分け破砕し,ピロ亜硫酸カ リウムをSO2濃度で70 mg/kg になるように添加した.CS は糖度が 16.7°Brix と低かっ たので, 21° Brix まで上白糖で補糖を行った.発酵 1 日目に酒母として LALVIN EC1118
16 (Lallemand 社)を果汁に 200 mg/kg となるように添加した.また,果汁と果帽を混合 するために,朝と夕の1 日 2 回梅酒瓶を回した.その際,酸化を防ぐために少量のドラ イアイスを添加した.各試験醸造は品種ごとに3つずつ独立で実施した. 5. 醸造中のマスト液体およびブドウ果皮のサンプリング サンプリングは発酵期間中の毎日午前9 時行った.梅酒瓶にドライアイスを入れ,ヘ ッドスペースを炭酸ガスで充満させた後に,20 mL 容遠心管にマストを 15 mL 採取し た.サンプリング後,直ちに20℃で 15,000 x g で 5 分間遠心分離を行った.そこで得ら れた上清を別の容器に移し,それをマストのサンプル試料とした. また,発酵中の果皮分析を行うにあたり,サンプリング毎に約 5 g の果皮を採取し, 速やかに液体窒素で凍結した後,分析時まで-80℃で保存した. 6. ブドウ果皮のアントシアニン抽出 発酵中のブドウ果皮のアントシアニン抽出は,Mori et al. (2005)の方法に基づいて行った. ① あらかじめサンプリングし,-80℃で保存してあった果皮約 5 g を液体窒素中です り鉢と乳棒を使用し,粉末化した. ② 粉末状の果皮0.25 g を秤量し,15 mL 容の遠心管に採取し,抽出溶媒(ギ酸/水/メ タノール(2:28:70 (v/v/v))を 5 mL 添加した. ③ その後4℃で 24 時間,5 mL の抽出溶媒中(ギ酸/水/メタノール(2:28:70 (v/v/v))で 暗所にて抽出を行った. ④ 抽出物を15,000 x g で 5 分間遠心分離し,そこで得られた上清部分を別の容器に移 し,それを果皮のサンプルとした.
17 7. 醸造中における分析 7-1 pH,糖度(Brix),アルコール濃度分析 ・ pH の分析 pH メーターを用いて,発酵中のマストの pH を 3 回ずつ測定した. ・ 糖度(Brix)の分析 糖度計を用いて,発酵中のマストの糖度を3 回ずつ測定した. ・ アルコール濃度の分析 簡易型アルコール分析器であるアルコメイトを用いて,発酵中のマストのアルコー ル濃度を3 回ずつ測定した. 7-2 ワインカラー分析 A) 試薬の調製 ・ 1M 塩酸 110 mL の逆浸透水に 10 mL の 12N 塩酸を加えて混合した. ・ 0.1M 塩酸 90 mL の逆浸透水に 10 mL の 1M 塩酸を加えて混合した. ・ 20% メタ重亜硫酸カリウム水溶液 1000 mL の逆浸透水に 20 g のメタ重亜硫酸カリウムを添加し混合した. B) 操作:ワインカラー(Ribéreau-Gayon et al., 2006) ③ 光路長2 mm のセルを用い,逆浸透水を対照としてワインの 420,520,620 nm の 吸光度を測定した. ④ 得られた値を5 倍し,A420,A520,A620 とした.また,下記の計算式より Color intensity を算出した. Color intensity = A420 + A520 + A620
18
C) 操作:pH 2.5 でのワインカラー(Wine Color in Acid: WCA)(Bakker et al., 1986) ① 試験管に0.5 mL の試料を取り,10 mL の 1M 塩酸を加え,ただちに攪拌した. ② 色素重合体カラーで使用するため,別の試験管に6.5 mL を採取した.
③ 光路長10 mm セルを用いて 520 nm の吸収を測定した.
④ 得られた値に21 倍し,これを pH 0.25 でのワインカラー(WCA)とした.
D)操作:色素重合体カラー(Polymeric Pigment Color: PPC)(Bakker et al., 1986) ① pH 0.25 でのワインカラーで残しておいた 6.5 mL に 20%メタ重亜硫酸カリウム水 溶液を0.1 mL 加え,ただちに攪拌した. ② 1 分後,光路長 10 mm セルを用いて 520 nm の吸収を測定した. ③ 得られた値に21 倍し,これを色素重合体カラー(PPC)とした. E)操作:色差 ① ガラスセルに蒸留水を入れ,標準校正を行った. ② ガラスセルに試料を入れ,透過試料室のセルガイドにセットし,測定した. 7-3 全フェノール濃度(フォーリン・シオカルト法) 第二章 6-3 を参照. 7-4 BSA 結合性タンニン濃度(BSA 沈殿法) 第二章 6-4 を参照.
19 7-5 HPLC によるアントシアニンの分析
HPLC-DAD によりアントシアニンの分析を行い,全アントシアニン量を求めた. Atlantis C18 T3 カラムを用い,溶出溶媒として,移動相 A には 0.4%リン酸,移動相 B にはアセトニトリルを用いた.検出器には,JASCO MD-2018 plus Photodiode Array Detector を用いた.HPLC-DAD の諸条件は,表 3-2 に示した.また,得られたピークは Malvidin-3-O-glucoside 量として換算した. 表3-2 HPLC によるアントシアニンの分析条件 カラム Atlantis C18-T3 カラムサイズ 4.6 i. d.×150 mm, 3 µm: Waters カラム温度 40℃ カラムオーブン CO-2065 plus 溶出液 移動相A:0.4%H3PO4,移動相B:CH3CN 溶出速度 1 mL/min 試料添加量 10 µL 検出器 MD-2018 plus 検出波長 520 nm ポンプ PU-2080 plus 溶出グラジエント 時間(min) 溶液 A(%) 溶液 B(%) 0 90.0 10.0 40.0 72.3 27.7 40.1 50.0 50.0 47.0 0 100 57.0 0 100 57.1 99.0 1.0 67.0 99.0 1.0
20
第二節 結果および考察
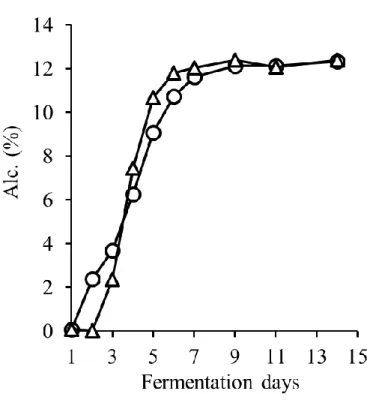
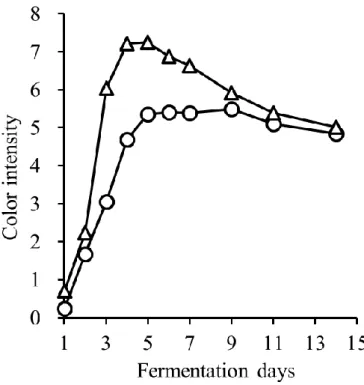
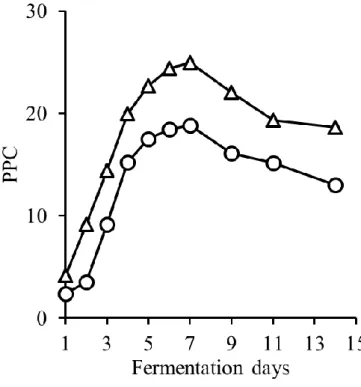
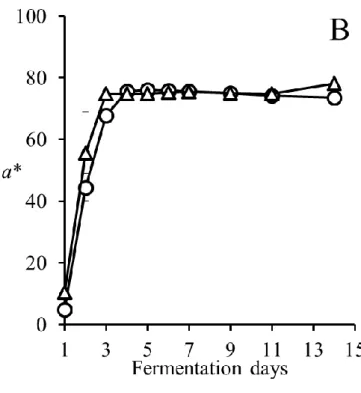
1. マスト液体中の pH,糖度(Brix),アルコール濃度および色調の推移 発酵中,CS は pH 3.7~pH 3.8,MBA は pH 3.5~pH 3.6 を推移し,発酵中に大きな変 化は認められなかった(図3-1).また,アルコールは酵母が糖を代謝することによって 生成される.そのためBrix の減少とアルコール濃度の増加は相関関係にあると言える. 従来通り2015 年度の醸造において,MBA において若干発酵が遅れ早く終了したが,両 品種とも6 日目辺りでアルコール濃度が 12%に達し,その後平衡状態になり,約 1 週間 で発酵が終了した(図3-2).本結果より,醸造試験において発酵は健全に進んだと考え られる. また,色調は赤ワインの品質に影響する重要な要素である.A420は黄色~茶色,A520 は赤色,A620は紫色である.Color intensity はこれら 3 つの値の合計値で赤ワインにおけ る色調の強度を表すものである.Color intensity は発酵初期に増加し,発酵開始後約 5 日 で最大値に達したがその後減少した(図3-3).酸性下でのワインの色を示す WCA と色 素重合体を表すPPC は,発酵開始から 7 日目で最大となり,その後減少した(図 3-4, 図3-5).さらに,色差計で L*,a*,b*を測定した(図 3-6-A,B,C).L*,a*,b*はワ インの見た目の色の強さを示す指標であり,L*は値が低いほど色が濃く,a*は値が高い ほど赤色が強く,b*は値が高いほど黄色が強いことを示す.一般的に CS は MBA に比 べ色調が濃いとされているが,本実験での醸造試験ではL*および a*に品種差は認めら れなかった.これは,CS のブドウの状態があまり良好でなかったこと,逆に MBA の状 態が良好であったことに起因すると考えられる.一方b*は CS では発酵後期に平衡状態 となったが,MBA では減少した.b*は増加すると黄色みを増し,減少すると青色みを 増していく.したがって,MBA では紫色を呈するキノイダルベースになりやすいアン トシアニンが多く含まれていた可能性が考えられる.21
図3-1 発酵中の pH の変化(CS;○,MBA;△)
22
図3-3 発酵中の Color intensity の変化(CS;○,MBA;△)
23
図3-5 発酵中の PPC の変化(CS;○,MBA;△)
24
図3-6-B 発酵中の a*の変化(CS;○,MBA;△)
25
2. 醸造中におけるマスト液体部分および果皮中の全フェノール量(Total phenolics; TP) およびBSA 結合性タンニン量(Proanthocyanidin; PA)の推移
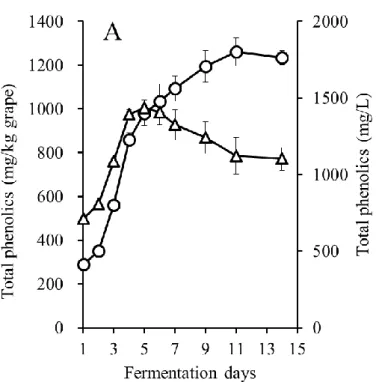
マストの液体部分に含まれるTP および PA 濃度を図 3-7-A および図 3-7-B に示した. なお本実験ではマスト液体部分と共存する果皮中に存在するフェノール化合物の全体 量を考えるため,液体部分のTP および PA 濃度(第二軸)に加え,圧搾率を 60%(1 kg のブドウから600 mL の液体が得られる)とした時に得られるフェノール化合物量を算 出しグラフの第一軸とした.なお,実際の圧搾率は,発酵中にブドウの果皮や果肉成分 が分解および溶解することで液量が徐々に増加し,発酵終了時には75~85%の圧搾率に なると推定されたが,試験醸造時の正確な液量を算出することは困難だったため,本実 験では圧搾率を一定に定め,60%として算出した. CS では TP 濃度は発酵とともに増加し続け最大 1802 mg/L となり,MBA では発酵開 始5 日目をピーク(最大 1436 mg/L)に達した.その後,MBA において TP 濃度は減少 に転じ,発酵終了時には1102 mg/L まで減少した.よって,23.3%の TP がマスト液体 中から消失し,第二章での醸造試験と同様の結果となった.PA 濃度は,CS では TP 濃 度と同様,発酵期間中上昇し続け447 mg/L だった.一方 MBA では,TP 濃度と同様, 発酵開始4 日目で最大濃度に達し 106 mg/L だったが,その後大きく減少し 31 mg/L と なった.この結果はMBA ワインには種子由来の PA が含まれていないことや,両品種 において果皮由来のPA が醸造後期に減少すること(第二章結果)が大きく関与してい ると考えられた. さらに,醸造期間中における果皮に含まれるTP,PA 量の推移を図 8-A および図 3-8-B に示した.1 kg のブドウの果皮に含まれるフェノール化合物量を算出するため,両 品種において,醸造に使用したブドウと同じブドウの果皮を手で分離し,それぞれ得ら れた果皮重量がマスト中に存在する事とし,第一軸にブドウ 1 kg あたりの果皮中のフ ェノール化合物量として示した.なお,発酵の経過とともに,果皮も徐々に分解し,果 皮重量の減少が考えられたが,マスト中の正確な果皮量を定量することが困難だったた め,発酵中の果皮量に大きな変化はないと仮定して,発酵開始から終了まで14 日間の 果皮中のフェノール化合物量を算出した. CS 果皮の TP 量は,発酵開始から 9 日目まで減少し 442 mg/kg grape となったが,そ
26 の後発酵終了時点で539 mg/kg grape まで増加した.また,PA 量の推移でも同様の推移 が見られ,発酵9 日目で 102 mg/kg grape まで減少したが,発酵終了時点では 156 mg/kg grape まで増加した.よって TP 量と PA 量の挙動が同じだったことから,CS ではフェ ノール化合物のなかでも,特にPA が果皮に再吸着したことが示唆された.そして,本 実験結果より,アルコール発酵が終了した果皮には発酵開始前の半分程度のTP が残存 することが明らかになった.MBA 果皮の TP 量は発酵開始から 6 日目まで急激に減少 し160 mg/kg grape となったが,その後は緩やかに増加し 221 mg/kg grape となった.ま た,本結果から,MBA ではアルコール発酵終了時の果皮には発酵開始前の 1/3 程度の TP が残存することが明らかになった.MBA におけるブドウ 1 kg あたりに含まれる TP 量は,発酵開始時,果汁に550 mg 程度,果皮中に 570 mg 程度含まれており,果汁と果 皮に合計1120 mg 程度の TP が含まれていたが,発酵 14 日目では果汁と果皮中の TP 量 の合計は1000 mg 程度だった.また,PA 量は発酵 3 日目から減少しはじめ,11 日目で 果皮中のPA はほとんどなくなった.従って MBA のマストおよび果皮の TP・PA 量の 結果から,果皮に再吸着が起こったのは,一度マストに抽出されたPA 以外のフェノー ル化合物であることが示唆された. 一方,CS においてブドウ 1 kg あたりに含まれる TP 量は,発酵開始時,果汁に 300 mg 程度,果皮中に 800 mg 程度含まれるため,果汁と果皮の合計で 1100 mg 程度の TP が含まれていた.一方,発酵14 日目のマストの TP 量はブドウ 1 kg あたり 1200 mg 程 度,果皮では540 mg 程度あり合計 1700 mg 以上となった.したがって,発酵初日と終 了時の差である500 mg 相当の TP が種子から抽出されたことが考えられた.果皮中の TP が増加する時期は種子由来のタンニンが増加する時期と重なることと,MBA 果皮中 のPA 量が減少することを併せて考えると,醸造時に種子由来の PA が果皮に優先的に 再吸着することが示唆された. これらの結果は,果皮の不溶性化合物(IC)はマストに抽出されたフェノール化合物 を再吸着し,また,より選択的にPA を吸着することを示した.しかし,MBA ではマス トに抽出されるPA が元々少ないので,PA 以外のフェノール化合物が果皮に再吸着し, 結果的にフェノール化合物量の少ない赤ワインとなる可能性が示唆された.
27
図3-7-A 発酵中のマスト液体中に含まれる TP 量の変化(CS;○,MBA;△)
28
図3-8-A 発酵中の果皮に含まれる TP 量の変化(CS;○,MBA;△)
29 3. 醸造中におけるマスト液体部分および果皮中のアントシアニン量の推移 マスト液体部分に含まれる全アントシアニン量は,CS,MBA 両品種において発酵開 始後6 日目まで増加し,最大濃度(CS で 220 mg/L,MBA で 231 mg/L)に達し, その後 減少し,CS で 143 mg/L,MBA で 121 mg/L であった(図 3-9-A,図 3-9-B).また,果 皮中の全アントシアニン量は,発酵開始後5 日目まで急激に減少し,その後平衡状態の まま推移した(図3-10-A,図 3-10-B). さらに疎水性の異なるアントシアニン類でかつ全体のアントシアニン量に対する比 率が高かった3 種類のアントシアニン[CS;Cyanidin-3-glucoside (Cy-3-glc), Malvidin-3-glucoside (Mv-3-glc), Malvidin-3-acetyl glucoside (Mv-3-Acglc)], [MBA;Peonidin-3-glucoside (Pn-3-glc), Mv-3-glc, Malvidin-3-(6-p-coumaroyl)-glucoside-5-glucoside (Mv-3-(6-p-coumaroyl)-glc-5-glc)]の発酵中におけるマストおよび果皮中での推移を図 3-11 お よび図3-12 に示した.疎水性の低い Cy-3-glc や Pn-3-glc などのアントシアニンは,発 酵初期にマストへ抽出され,その後大きく減少した(図3-11-A,図 3-12-A).Mv-3-glc はCS および MBA ワイン中に最も多く含まれるアントシアニン類であることが知られ ており(Ryan and Revilla, 2003; Koyama et al., 2017),Cy-3-glc および Pn-3-glc よりも疎 水性が高いアントシアニンである.CS および MBA 両品種における Mv-3-glc は発酵開 始から一週間ほどかけて抽出され,その後CS で 39%,MBA で 52%減少した.よって 果皮中のMv-3-glc も抽出され減少し,その後平衡状態となった(図 3-11-B,E,図 3-12-B,E).一方,疎水性の高い Mv-3-Acglc および Mv-3-(6-p-coumaroyl)-glc-5-glc は発酵初 期の抽出では,Mv-3-glc と同様の挙動を示したが,マストへ抽出された後の減少率が低 く,液体中に残存し,その割合はそれぞれCS で 22%,MBA で 15%だった(図 3-11-C, F,図 3-12-C,F).このことから,疎水性の低いアントシアニン類は容易に抽出されや すく,疎水性の高いものは少し遅れて発酵中期にかけて抽出される傾向にあること,疎 水性の高いアントシアニンは低いものと比較すると,マスト中に残存していることが示 された.しかし,発酵中の果皮に含まれるアントシアニン量は品種や組成に関係なく, 発酵開始から一週間ほどで抽出され,その後変化は認められなかった.よって,発酵期 間中に抽出されたPA は果皮に再吸着したが,アントシアニン類は果皮への再吸着が起 こらなかったか,吸着と抽出が同時に起き,見かけは平衡状態となっていたことが考え
30 られた. また,ブドウ 1 kg あたりのアントシアニン量において,マストのアントシアニン量 が果皮に比べて少なかった(図3-11,図 3-12).その要因として,pH 上昇による無色化 やコピグメンテーション,種子PA による色素安定化(Sparrow et al., 2015)に加え,発 酵中にブドウや酵母由来の多糖類やタンパク質と結合し安定化しており,従来の分析方 法では検出できない形でワイン中に存在している可能性が考えられた.
31
図3-9-A CS における発酵中のマスト液体中に含まれる全アントシアニン量の変化
32
図3-10-A CS における発酵中の果皮に含まれる全アントシアニン量の変化
33 図3-11 CS における発酵中のマスト液体中(A,B,C)および果皮中(D,E,F)の アントシアニンの変化
34 図3-12 MBA における発酵中のマスト液体中(A,B,C)および果皮中(D,E,F) のアントシアニンの変化
35 4. 醸造試験まとめ 以上の結果より,醸造中に一度果皮から抽出されたフェノール化合物,特にPA は果 皮に再吸着し,ワインのフェノール化合物濃度を低下させている可能性があることが示 唆された.アントシアニンに関しては,疎水性の低いものほど発酵初期に抽出され,そ の後マストからも消失したが,疎水性の高いアントシアニンは,抽出された後もマスト 中に残存することが明らかになった.また,一度抽出されたアントシアニンは果皮に再 吸着せず,無色化や安定化などの影響を強く受けたことが推測された. 本実験では,醸造中のマスト液体部分および果皮中のフェノール化合物の抽出挙動を 分析し,果皮からどのようにフェノール化合物が抽出され,一部は再吸着することを明 らかにした.また,赤ワイン醸造において,圧搾時点の果皮には相当量のTP や PA(CS で約1/2,MBA で約 1/3)が存在し,果皮中の TP および PA 量が発酵中に増加すること が明らかになった.従って,液体であるワイン中のフェノール化合物濃度を高めるため には,果皮中のフェノール化合物が最低となる時期に圧搾を行うことが,再吸着を防止 し,ワイン中のフェノール化合物濃度を最大にする上で,最良の方法であると考えられ た.今回の結果では,CS では 9 日目,MBA では 6 日目で圧搾することが適当だと考え られた.しかし,種子や果肉にも吸着は起こりうるため,これらを含めた実験が必要で ある.また,パンチダウンやポンピングオーバーなど果帽の処理方法やその強さも大き な影響を与えることが考えられた.しかし,発酵が終わり,ワインになってからの長期 的な色調の安定性などを考慮した,果皮,種子,果肉などの重要性についても報告され ており(Sparrow et al., 2015),抽出や吸着の現象を調べる以外にもさらに多くの実験が 必要である.今後,赤ワイン醸造のためのさらなるフェノール化合物抽出機構が明らか になることが期待される.
36
第四章 ブドウ由来不溶性化合物へのフェノール化合物の
吸着特性
第三章では,赤ワイン醸造においてフェノール化合物,特にタンニン(PA)が果皮由 来の不溶性化合物に再吸着することが明らかにされた.赤ワインを醸造する際,原料ブ ドウの品種や産地,熟度などにより,マストのpH は変化する.また,醸造中に生成す るエタノールが様々な化合物の抽出機構に影響を与えることは容易に想像できる.しか し,フェノール化合物の吸着活性を調べた先行研究では,pH やエタノール濃度を固定 して行った報告はあるものの,醸造によって変化するpH やアルコール濃度などの変化 が,吸着活性にどのような影響を与えるのかは明らかにされていない.吸着活性に影響 を与える要因を特定できれば,フェノール化合物の抽出機構の解明や,ワイン製造に適 した果汁の調整技術の開発および科学的理解に大きな知見を与えると考える. 本章では,CS や MBA の不溶性化合物(果皮・果肉・種子由来)のタンニンやアント シアニン類であるフェノール化合物に対する最大吸着能と吸着条件を明らかにするこ とを目的としてモデル系実験を行った.第一節 試料と方法
1. 試料ブドウ 試料ブドウには,2015 年 9 月 24 日に山梨大学生命環境学部附属小曲農場で収穫され た赤ブドウ2 品種を供試した(表 4-1). 表4-1 試料ブドウ 品種 樹齢(年) 台木Cabernet Sauvignon (CS) 7 Teleki 5BB Muscat Bailey A (MBA) 7 Teleki 5BB
37 2. 試薬 2-[4-(2-ヒドロキシエチル)-1-ピペラジニル] エタンスルホン酸(HEPES) 特級 ナカライテスク株式会社 アセトン 特級 富士フイルム和光純薬株式会社 2-アミノ-2 ヒドロキシメチル-1,3-プロパンジオール(Tris) 特級 富士フイルム和光純薬株式会社 フェノール 特級 富士フイルム和光純薬株式会社 エタノール 特級 富士フイルム和光純薬株式会社 メタノール 特級 富士フイルム和光純薬株式会社 クロロホルム 一級 関東化学 ヘキサン 特級 富士フイルム和光純薬株式会社 酒石酸水素カリウム Guaranteed Reagent ナカライテクス株式会社 塩酸 特級 富士フイルム和光純薬株式会社 炭酸ナトリウム 特級 富士フイルム和光純薬株式会社 フォーリン・シオカルト試薬(F-C 試薬) 富士フイルム和光純薬株式会社 没食子酸 Extra Pure Reagent ナカライテクス株式会社 牛血清アルブミン,コーンフラクションV,pH 7.0 生化学用 富士フイルム和光純薬株式会社 酢酸 特級 富士フイルム和光純薬株式会社 塩化ナトリウム 特級 関東化学株式会社 水酸化ナトリウム 特級 富士フイルム和光純薬株式会社 トリエタノールアミン(TEA) 特級 富士フイルム和光純薬株式会社 ドデシル硫酸ナトリウム(SDS)一級 富士フイルム和光純薬株式会社 塩化第二鉄六水和物(FCR) 特級 富士フイルム和光純薬株式会社 3. 機器 電子天秤 SHIMAZU AUW-220D 遠心分離機 HITACHI CF-16-RN ホモジナイザー POLYTRON PT 10-35 GT
38 グラインダー NATIONAL MX-C-X103-D
振とう機 YAMATO SCIENTIFIC CO., LTD BW-400 凍結乾燥機 TAITEC VD-800F FREEZE DRYER ウォーターバス YAMATO THERMO-MATE BF400
HPLC JASCO MD-2018 PLUS PHOTODIODE ARRAY DETECTOR エバポレーター EYELA ROTARY EVAPORATOR N-1000
アスピレーター EYELA A-1000S
pH メーター HORIBA pH/COND METER F-54
吸光光度計 HITACHI SPECTROPHOTOMETER U-2900 試験管ミキサー TAITEC CO., LTD SE-04
4. 分析方法
4-1 ブドウ果皮由来フェノール化合物の抽出および分画(Kennedy and Jones; 2001) A) 試薬の調製 ・ アセトン溶液(2:1 (v/v) acetone:water) アセトン2000 mL,逆浸透水 1000 mL を混合し,メンブランフィルターでろ過を した. ・ エタノール溶液(13% (v/v)) エタノール130 mL に逆浸透水を加え 1000 mL にメスアップし,メンブランフィ ルターでろ過をした.
・ 1:1 (v/v) MeOH:water containing 0.1% TFA
メタノール1000 mL,逆浸透水 1000 mL を混合し,TFA を 2 mL 加え,よく混合 し,メンブランフィルターでろ過をした.
・ 2:1 (v/v) acetone:water containing 0.1% TFA
メタノール2000 mL,逆浸透水 1000 mL を混合し,TFA を 2 mL 加え,よく混合 し,メンブランフィルターでろ過をした.
39 B) 操作 ① ブドウ果皮が浸るようにアセトン溶液を入れ,室温で24 時間撹拌と振とうを行った. ② ロータリーエバポレーターを用いて,35℃にてアセトンを除去し,その後少量の メタノールを添加した. ③ Toyopearl HW 40-F をメタノールおよび逆浸透水にて洗浄し,カラムに充填した. ④ Toyopearl HW 40-F は 1:1 (v/v) MeOH:water containing 0.1% TFA 溶液をカラム体積
の3 倍量流し平衡化した.
⑤ ②を1:1 (v/v) MeOH:water containing 0.1% TFA 溶液に溶解し,平衡化後のカラムに アプライした.
⑥ 1:1 (v/v) MeOH:water containing 0.1% TFA 溶液を流し,アントシアニン画分を溶出 した.
⑦ 続いて2:1 (v/v) acetone:water containing 0.1% TFA 溶液を流し,タンニン画分を溶 出した. ⑧ それぞれの画分についてロータリーエバポレーターを用いて,メタノールまたは アセトンを除去し,それぞれアントシアニン画分,タンニン画分とした. 4-2 ブドウ果皮・果肉・種子由来不溶性化合物(IC)の調製(Bindon et al., 2010) A) 試薬の調製 ・ 40 mM HEPES 溶液(pH 7.0) 9.53g の HEPES を 1 L の逆浸透水に溶解した. ・ 70 % (v/v) アセトン溶液 アセトン70 mL,逆浸透水 30 mL を測りとり,混合した. ・ フェノール-0.2 M トリス‐塩酸緩衝溶液(pH 6.7) 2.4g のトリスを 60mL の逆浸透水に溶解させて,塩酸で pH 6.7 にメスアップした. フェノールを1/4 量混合し,激しく撹拌したのち遠心分離を行い,下層部を使用した. ・ 80 % (v/v) エタノール溶液 エタノール80 mL,逆浸透水 20 mL を測りとり,混合した. ・ 1:1 (v:v) メタノール:クロロホルム溶液 メタノール50 mL,クロロホルム 50 mL を測りとり混合した.
40 B) 操作(果皮由来 IC の精製) ① 凍った果皮を70 % (v/v) アセトン溶液に浸漬した(24 時間). ② アセトン溶液を除いた後,蒸留水で洗浄した. ③ 果皮をホモジナイズ(8000rpm,20 分間)した. ④ フェノール-0.2 M トリス‐塩酸緩衝溶液を加え洗浄した. ⑤ ④の不溶性画分に80 % (v/v) エタノール溶液を加え,2 回洗浄した. ⑥ ⑤の不溶性画分にアセトンを加え,石炭酸を取り除く作業を3 回行った. ⑦ ⑥の不溶性画分に1:1 (v:v) メタノール:クロロホルム溶液を加え,ゆっくり 30 分 間振とうさせた. ⑧ ⑦の不溶性画分を凍結乾燥し,液体窒素中で粉末化して果皮由来不溶性化合物とし て以降の実験に用いた. C) 操作(果肉由来 IC の精製) ① 冷凍状態のブドウ果肉をホモジナイズ(8000rpm,20 分間)し,液体状にした. ② 直ちに40 mM HEPES 溶液を①で得られた液体状のものと同量添加した. ③ 撹拌(4℃,15 分間)した. ④ 遠心分離(4℃,8000g,20 分間)した. ⑤ 70 % (v/v) アセトン溶液に④で得られた沈殿物を加えた. ⑥ ⑤のアセトン溶液を除いた後,蒸留水で洗浄した. ⑦ ⑥の不溶性画分にフェノール-0.2 M トリス‐塩酸緩衝溶液を加え洗浄した. ⑧ ⑦の不溶性画分に80 % (v/v) エタノール溶液を加え,2 回洗浄した. ⑨ ⑧の不溶性画分にアセトンを加え,石炭酸を取り除く作業を3 回行った. ⑩ ⑨の不溶性画分に1:1 (v:v) メタノール:クロロホルム溶液を加え,ゆっくり 30 分 間振とうさせた. ⑪ ⑩の不溶性画分を凍結乾燥し,液体窒素中で粉末化して果肉由来不溶性化合物と して以降の実験に用いた.
41 D) 操作(種子由来 IC の精製) ① 乾燥したブドウ種子を70% (v/v) アセトン溶液に浸けた(3 日間). ② ①の種子をグラインダーで粉末化した. ③ ②の粉末種子をヘキサン,70% (v/v) アセトン溶液,1:1 (v:v) メタノール:クロロ ホルム溶液で複数回洗浄した. ④ ③の粉末種子を,さらに液体窒素中で粉末化して種子由来不溶性化合物とした. 4-3 各種モデルワインの調製 A) 試薬の調製 ・ モデルワイン 120 mL のエタノールと 800 mL の逆浸透水を混合し,5.0 g の酒石酸水素カリウム を添加し,スターラーで攪拌し溶解させた.1N 塩酸で pH 3.3 となるように調整し た後,逆浸透水で1000 mL にメスアップした. 上記のモデルワインの調製法を基本として,pH 調製時に後の実験の反応時の pH を 考慮した上で,pH 3.2~6.0 の pH のそれぞれ異なるモデルワインを調製した.また, エタノールと逆浸透水の比率を変えることで,アルコール濃度が0%~14%のモデル ワインを調製した.pH の調製は,適宜塩酸水溶液または塩化カリウム水溶液を用い た. 4-4 全フェノール濃度(Total Phenol: TP)(フォーリン・シオカルト法) 第二章6-3 を参照.
4-5 BSA 結合性タンニン(Proanthocyanidin: PA)濃度(BSA 沈殿法) 第二章6-4 を参照.
4-6 HPLC によるアントシアニンの分析 第三章 7-5 を参照.
42 4-7 ブドウ果皮・果肉・種子由来 IC へのフェノール化合物の吸着実験 4-2 で精製したブドウ由来 IC(50 mg)とフェノール化合物(10 mg)を異なる pH(反 応時pH 2.9~4.2)およびアルコール濃度(0~14%)のモデルワイン(10 mL)中で 32℃ にて 1 時間反応させ,遠心分離を行い,得られた上清の全フェノール濃度を F-C 法, BSA 結合性タンニン濃度を BSA 沈殿法,アントシアニン組成の分析を高速液体クロマ トグラフィー(HPLC)で分析を行った.
43
第二節 結果と考察
1. ブドウ果実からの不溶性化合物の回収率 CS および MBA の果皮,果肉,種子の生果重量および乾燥重量を表 4-2 に示した.CS はMBA よりも果粒が小さい特徴があるため,同じ重量だと CS は MBA よりも果粒数 が多くなり,結果として果皮や種子が多かった.果皮と種子由来IC の乾燥重量は,MBA よりもCS で多かった.果肉の生果重量は,果実重量の 80%以上を占めた.しかし,生 果の果肉には多量の果汁が含まれているので,果肉から得られるIC の量は,両品種に おいて果皮または種子から得られるものより少なかった. 表4-2 ブドウ果実の重量および不溶性化合物の乾燥重量 CS MBA 生果重量 (g/kg berry) IC (dry g/kg berry) 生果重量 (g/kg berry) IC (dry g/kg berry) 果皮 果肉 種子 129.4±8.7 829.2±24.1 41.4±3.6 8.7±1.1 2.7±1.4 10.4±0.8 101.7±9.0 871.8±19.2 26.5±5.5 6.1±0.9 4.6±1.5 6.1±1.844 2. ブドウ由来 IC のアントシアニン吸着能 アントシアニンの果皮・果肉・種子から得られた IC への吸着実験を行った.本実験 で得られたアントシアニンの総量は,ブドウ1 kg あたり CS で 911 mg/kg,MBA で 782 mg/kg であった.さらに表 4-2 の結果に基づいて,果皮中の全アントシアニンに対する 果皮・果肉・種子におけるIC の比率を算出した.これらの比率を用いて,果皮中に含 まれる全アントシアニン量に対するIC の吸着能を比較した(図 4-1). CS において,標準モデルワイン溶液(pH3.3,12%エタノール)では,果皮 IC は全ア ントシアニンのうち12%を吸着し,果肉 IC と種子 IC はそれぞれ 4%の全アントシアニ ンを吸着した.MBA では標準モデルワイン溶液において,全アントシアニン量のうち 果皮IC は 20%,果肉 IC は 5%,および種子 IC の 3%の吸着能を有していた.合計では, CS の IC は全アントシアニンのうち 20%の吸着能があり,MBA の IC は全アントシア ニンのうち28%の吸着が認められた. CS と MBA を比較すると,アントシアニンに対 してMBA 果皮 IC は約 8%高い吸着能が認められたが,果肉および種子 IC は両品種に おいて吸着能は5%未満であった.よって MBA 果皮 IC の高い吸着能は MBA の特性で あることが考えられる. また,ODS カラムを用いたアントシアニンの HPLC 分析では,溶出時間の遅いアン トシアニンがIC に対して,吸着しやすい傾向があり,この結果はアントシアニンの疎 水性がIC への吸着に影響を及ぼす可能性があることを示唆した.CS と MBA のアント シアニン組成は異なることが知られており(Koyama et al., 2017),これらの違いが IC と の結合親和性に影響を与えている可能性が示唆された.しかし本実験では,アントシア ニンの構造同定を行わなかったため,アントシアニンの組成および構造が吸着に及ぼす 影響について結論は出なかった. また,ムラサキニンジンを用いた実験において,セルロースやペクチンといった細 胞壁物質の曝露が,アントシアニンの吸着に影響を及ぼすことが報告された
(Padayachee et al., 2012).IC を構成する不溶性多糖類の組成の違いは,アントシアニ ン吸着能に影響を与える可能性があるが,CS および MBA の IC を構成する不溶性化 合物の組成について,詳細はほとんど知られていないので,IC の組成がどの程度アン トシアニン吸着能に影響を与えているかは不明である.また,本実験ではIC を粉末化
45
し,アントシアニンに対する最大吸着能を測定した.したがって,結果で得られたIC へ吸着するアントシアニン量は,実際のワイン製造条件下よりも多かったと推察される.
46 3. ブドウ由来 IC のタンニン吸着能 アントシアニン吸着実験と同様に,果皮由来のタンニンに対する果皮・果肉・種子の IC の比率を算出し,皮膚タンニンに対する IC の吸着能を計算した.得られた果皮タン ニンは,ブドウ1 kg あたり CS で 2071 mg/kg,MBA で 1255 mg/kg であった.CS およ びMBA 由来 IC の果皮タンニン吸着率(%)を図 4-2 に示した. CS および MBA 両品種において,果皮 IC は果皮タンニンの 30%を吸着した.対して 種子IC は CS で 18%,MBA で 45%吸着した.果皮タンニンは分子量が比較的大きく, ブドウ品種によって平均重合度(mDP)や組成が異なることが報告された(Koyama et al., 2017).さらに,分子量が大きいタンニンは IC に容易に吸着することが報告された
(Bindon et al., 2014; Le Bourvellec et al., 2004).一方,本実験では IC のタンニン吸着能 はブドウの部位によっても異なることが示された.また, MBA 種子 IC における高い 吸着能は,MBA 赤ワイン醸造においてタンニンが低濃度になってしまう(Ichikawa et
al., 2011; Ichikawa et al., 2012; Okuda et al., 2014)要因のひとつとなる可能性が示唆された.
47 4. 果皮 IC へのアントシアニン吸着に及ぼす pH とアルコール濃度の影響 赤ワイン醸造時,発酵後期にアントシアニン濃度が低下する現象(Okuda et al., 2014) と,果皮IC のアントシアニンとの高い吸着能(図 4-1)との関係が重要であると考えら れた.そこで,さらに果皮IC における pH(図 A)とエタノール濃度(%)(図 4-3-B)の条件を変化させたうえでアントシアニンの吸着を測定した.本実験は,2.5〜5.0 の pH 範囲および 0〜14%のエタノール濃度で行った. pH の変化に関係なく,MBA 果皮 IC においてアントシアニンの吸着率は,CS 果皮 IC よりも高く,この結果は図4-1 を支持した.また,アントシアニン吸着率は pH の変化 によって変動し,CS では pH 3.2 および pH 4.0 で,MBA では pH 3.0 と pH 4.4 で極大の 吸着率が見られたが,CS における pH による変化は明確ではなかった.また,pH 3.5 付 近で吸着率の極小が認められた.アントシアニンはいくつかの形態が存在するが,フラ ビリウムのみが正電荷を有するので,IC による吸着に影響を与えている可能性が考え られた.アントシアニンのIC への吸着に極大および極小の吸着条件が存在する理由は 本実験では明らかにされなかったが,IC に含まれるタンパク質の等電点(pI)やカルボ キシ基の酸解離定数(pKa)がアントシアニンとの相互作用に影響を及ぼしている可能 性が示唆された.現在,IC の化学的・物理的性質についてはほとんど知られておらず, さらなる研究を進める必要がある. 一方,アントシアニン吸着率はエタノール濃度の増加に伴って減少し(図4-3-B),果 皮IC へのアントシアニン吸着が疎水性相互作用を介して起こることを示唆した.実際, より疎水的なアントシアニン(アシル化アントシアニン)は,より吸着しやすい傾向が あり,親水的なアントシアニンはエタノール濃度の増加に伴って吸着しにくくなる現象 が見られた.
48
図4-3 pH(A)とエタノール濃度(B)が果皮 IC のアントシアニン吸着率に与える 影響.(○:CS,△:MBA)
49 5. 果皮 IC へのタンニン吸着に及ぼす pH とアルコール濃度の影響 アントシアニン吸着実験と同様に pH(図 4-4-A)およびエタノール濃度(図 4-4-B) に着目し,果皮IC への果皮タンニンの吸着条件を検討した.果皮タンニンの吸着率は pH の影響を強く受け,pH3.3 付近で極大が認められた.MBA では果皮タンニンの吸着 率はpH 4.2 より高い pH で再び増加した.一方,エタノール濃度の増加に伴い,果皮タ ンニンの吸着率はMBA と CS の両方で減少した.この結果は,アントシアニン吸着実 験で認められた現象と同じであり,果皮タンニンの吸着は疎水性相互作用を介して起こ ることが示唆された.プロシアニジンとリンゴ由来の細胞壁物質との相互作用は,弱い 水素結合と疎水性相互作用によって引き起こされることが報告された(Le Bourvellec et al., 2004). 第二章および第三章で,赤ワイン醸造中に果皮フェノール化合物の濃度が低下し,そ の一部が果皮に再吸着することが明らかになり,本章では,アントシアニンおよび果皮 タンニンに対するIC の最大吸着能が示された.また,アントシアニンと果皮タンニン の吸着および抽出におけるpH の影響が明らかになったことは,赤ワイン醸造において 重要だと考えられる. 図4-4 pH(A)とエタノール濃度(B)が果皮 IC のタンニン吸着率に与える影響. (○:CS,△:MBA)