バクテリオファージ Mu のサブユニットの構造解析と 結合サブユニットの同定
Structural analysis of bacteriophage Mu subunits and identification of their bound subunits
群馬大学 研究・産学連携推進機構 高度人材育成センター
岩﨑 拓真
目次
第 1 章 研究の背景と目的.............................................. 1 1.1 バクテリオファージの研究....................................... 1 1.1.1 ファージの研究の歴史とその応用.............................. 1 1.1.2 ファージの分類と構造........................................ 3 1.1.2-1 ICTV による分類........................................ 3 1.1.2-2 テイルを持つファージである Caudovirales と、3 つの Family 4 1.1.3 Caudovirales Order に属するファージの感染と増殖の過程........ 6 1.1.4 Caudovirales に属するファージの自律的形態形成............... 8 1.1.5 ファージのネックサブユニットの数や配置のバリエーション...... 9 1.1.6 バクテリオファージの立体構造の決定.......................... 10 1.1.6-1 ファージの構造解析の進歩................................ 10 1.1.6-2 Caudovirales の 3 つの Family の間での、
サブユニットの立体構造の相関............................ 11 1.1.7 ネックサブユニットの配置は様々である........................ 14 1.1.8 ネックサブユニットの配置から得られる知見.................... 14 1.1.9 テイルファイバーの研究...................................... 15 1.1.10 構造サブユニットの配置の決定............................... 15 1.1.10-1 立体構造の決定による配置の決定......................... 15 1.1.10-2 複合体形成によるサブユニットの配置の理解............... 16 1.2 Mu ファージの研究の背景....................................... 17 1.2.1 Mu ファージ................................................. 17 1.2.2 Mu ファージの研究の歴史..................................... 18 1.2.3 Mu ファージの遺伝子......................................... 19 1.2.4 Mu ファージの構造既知のサブユニット......................... 22 1.2.5 当研究室における Mu ファージの研究の経緯と研究成果.......... 22 1.2.5-1 当研究室における Mu ファージの研究の背景................ 22 1.2.5-2 本研究を開始した当時の背景と、Mu ファージのテイルの研究 22 1.3 第 1 章のまとめと本研究の目的.................................... 24 1.4 本論文の構成.................................................... 24 1.5 参考文献 ....................................................... 25 第 2 章 Mu ファージのネックサブユニット gp36 の立体構造の決定と解析.... 30 2.1 序論............................................................ 30 2.1.1 本章の目的.................................................. 30 2.1.2 Mu ファージのネックサブユニット............................. 30
2.2 Mu ファージのネックサブユニット gp36 の大量発現と単離精製....... 31 2.2.1 Mu gp36H 発現ベクターの作成................................ 31 2.2.2 Mu gp36H 発現菌の作成と大量発現............................ 31 2.2.3 Mu p36H の単離精製......................................... 32 2.3 単離精製した gp36 の分析と X 線結晶構造解析...................... 32 2.3.1 超遠心分析による gp36H 精製物の分子量の測定................. 32 2.3.2 Mu ファージのネックサブユニット gp36H(Mu gp36H)の結晶化 33 2.4 X 線結晶構造解析................................................ 35 2.4.1 Mu p36H の結晶構造......................................... 37 2.4.1-1 Mu gp36H のα-Helix の残基側鎖間の相互作用.............. 37 2.4.1-2 Mu gp36H のタンパク質表面に局在するアミノ酸と
中心部の疎水性コア...................................... 39 2.4.2 Mu p36H の立体構造を鋳型とした相同性検索................... 40 2.4.2-1 Mu p36H の立体構造と相同性を持つ構造既知タンパク質の検索 40 2.4.2-2 Mu gp36H と構造に相同性が見られたタンパク質の概要...... 40 2.4.2-3 Mu gp36H の構造は複数のファージのネックに
共通して見られる........................................ 40 2.4.2-4
Myoviridae、Siphoviridae、Podoviridae
の間でネックサブユニットが共通していた........................ 44 2.4.3 Mu gp36 と構造が類似したタンパク質の働きと、
構造の重ね合わせ............................................ 45 2.4.3-1 HK97 gp6 との比較...................................... 45 2.4.3-2 Phi29 gp10 との比較..................................... 47 2.4.3-3 P22 gp4 との比較........................................ 48 2.4.3-4 SPP1 gp15 との比較...................................... 49 2.4.4 C 末端の比較と Mu gp36 の重合条件の考察.................... 50 2.4.5 Mu-gp36 の重合体におけるサブユニット同士の重合境界面の予想 51 2.4.6 P22-gp4 から予想される Mu-gp36 の重合条件と
ネック領域の構造の予想...................................... 52 2.5 本章のまとめ.................................................... 53 2.6 参考文献........................................................ 54 第 3 章 Mu ファージのネックサブユニットの単離精製..................... 56 3.1 序論............................................................ 56 3.2 Mu ファージのネックサブユニットの単離精製...................... 56 3.2.1 Gp35H、gp37H、gp38H、T7-gp29、gp29H
発現ベクターと発現菌の作成.................................. 56
3.2.2 T7-gp29、gp29H、gp35H、gp37H、gp38H の大量発現と単離精製 56 3.2.2-1 Mu gp35H の単離精製.................................... 57 3.2.2-2 Mu gp37H の単離精製.................................... 57 3.2.2-3 Mu gp38H の単離精製.................................... 58 3.2.2-4 T7-gp29 の単離精製 ..................................... 59 3.2.2-5 超遠心分析による T7-gp29 の分子量の測定.................. 59 3.2.2-6 Gp29H の単離精製....................................... 60 3.2.2-7 Phenyl カラム精製によって凝集体が沈殿していた........... 61 3.2.2-8 透過型電子顕微鏡 (TEM) による T7-gp29 多量体の観察..... 62 3.2.3 ゲルろ過カラムクロマトグラフィーと超遠心分析による分子量測定 63 3.3 結晶化の検討と、第3章のまとめ.................................. 64 3.4 参考文献........................................................ 66 第 4 章 Mu ファージの複合体の形成と単離精製の試み..................... 67 4.1 序論............................................................ 67 4.2 Mu ファージのネック領域のサブユニットの複合体の単離精製........ 69 4.2.1 発現菌の混合精製による試験管内での結合の観察................ 69 4.2.1-1 発現菌の混合精製に用いる Mu gp36 発現菌の作成と大量培養 69 4.2.1-2 混合精製用の gp36 発現菌の作成と大量培養................. 69 4.2.1-3 発現菌の混合精製による
Mu gp36 と結合するネックサブユニットの探索.............. 69 4.2.1-4 Mu gp35H+ Mu gp36 複合体の単離精製................... 70 4.2.1-5 Western-blot による確認.................................. 72 4.2.1-6 共免疫沈降法による網羅的な結合の検討.................... 74 4.2.2 共発現ベクターを用いた菌体内での複合体形成.................. 75 4.2.2-1 共発現ベクターの作成.................................... 75 4.2.2-2 共発現菌の作成と発現.................................... 76 4.2.2-3 サブユニット同士の結合の観察............................ 76 4.2.2-4 透過型電子顕微鏡による gp35+gp36H 複合体の観察......... 77 4.3 本章のまとめ.................................................... 77 4.4 Mu ファージのネックにおけるサブユニットの配置の予想............ 78 4.5 参考文献........................................................ 82 第 5 章 共発現系による Mu ファージのテイルファイバーの複合体の形成.... 83 5.1 序論............................................................ 83 5.1.1 本章の目的.................................................. 83 5.1.2 Mu ファージのテイルファイバー領域のサブユニット............. 83 5.2 Mu ファージのテイルファイバーとシャペロンの単離精製............ 84
5.2.1 テイルファイバーとシャペロンの単独発現ベクターの作成........ 84 5.2.2 Mu ファージのテイルファイバーとシャペロンの発現の確認....... 85 5.2.3 Mu ファージのシャペロンの大量発現と単離精製................. 86 5.2.3-1 Mu gp50H の単離精製................................... 86 5.2.3-2 Mu H-gp51 の単離精製.................................. 87 5.3 Mu ファージのテイルファイバーとシャペロンの複合体の単離精製.... 88 5.3.1 テイルファイバーとシャペロンの発現ベクターの作成.......... 88 5.3.1-1 発現ベクターの作成.................................... 88 5.3.1-2 RT-PCR による gp49-gp50H、gp52-gp51H の mRNA の検出 88 5.3.2 Mu ファージのテイルファイバーとシャペロンの
複合体の発現の確認.......................................... 89 5.3.2-1 SDS-PAGE による観察................................... 89 5.3.2-2 Western-blot による確認.................................. 90 5.3.3 Mu ファージのテイルファイバーとシャペロンの
複合体の大量発現と単離精製.................................. 91 5.3.3-1 Mu gp49 + Mu gp50H 共発現系の精製...................... 92 5.3.3-2 Mu gp52 + Mu gp51H 共発現系の精製...................... 92 5.3.4 ヘテロな組み合わせの共発現系を用いたシャペロンの特異性の確認 93 5.3.4-1 Mu gp49 + Mu gp51H、Mu gp52 + Mu gp50H の発現の確認 93 5.3.4-2 Mu gp49 + gp51H、gp52 + gp50H の単離精製............... 95 5.4 本章のまとめ.................................................... 96 5.5 参考文献........................................................ 97 第 6 章 総括.......................................................... 98 6.1 本研究の結果.................................................... 98 6.2 参考文献........................................................ 103 Appendix 実験手順と資料の一覧....................................... 105 研究の業績............................................................ 128 謝辞.................................................................. 130
1
第 1 章 研究の背景と目的
1.1 バクテリオファージの研究
1.1.1 ファージの研究の歴史とその応用
バクテリオファージ(以下ファージ)は細菌に感染して増殖するウイルスである。それぞ れのファージが特定の細菌にのみ感染する宿主特異性を持ち、細菌が存在する環境には対 応するファージが存在すると言われている。ファージの総個体数は 1031 個と予想され、土 壌中や河川、海洋のほか、ヒトの腸内や排水中など、様々な環境中に普遍的に分布している。
海洋のファージを中心に現在でも新種の発見が続いており、また多種多様なファージが存 在する [1,2]。
ファージは現在に至るまで 100 年以上研究されてきた。1893 年に、Ivanovsky, D. が細菌 除去用のフィルター(Pasteur-Chamberland filter)を通過するタバコモザイク病の病原体
(タバコモザイクウイルス)の存在を報告した。歴史上初めてのウイルスの報告からわずか 3 年後の 1896 年に、インドのガンジス河の河水から、コレラ菌を殺菌してかつフィルター を透過する存在を、Hankin, E. H.が発見して報告した[3]。この報告がファージが初めて発 見された記録であったが注目されず、1915 年に Twort, F. によってファージが再発見され、
その後の 1917 年に d'Herelle, F. が赤痢菌ファージの発見を報告したことでようやく本格 的 な フ ァ ー ジ の 研 究 が 始 ま っ た 。 細 菌 を 溶 菌 す る ウ イ ル ス を バ ク テ リ オ フ ァ ー ジ
(bacteriophage)と命名した d'Hellele, F.は、ファージによる殺菌を細菌感染症の治療法と して利用する「ファージセラピー」を提唱した。グルジアに設立したギオルギ・エリアバ研 究所を中心に世界各地でファージセラピーの研究がなされ、臨床応用も行われた。しかしフ ァージの分類や構造解析はもちろん、分離や同定すらなされていない当時では安定した治 療は不可能であり、その後 1940 年代にペニシリンが発見されてからはファージセラピーは あまり注目されなくなった。
抗菌薬としての応用が下火となった一方で、ファージはモデル生物として盛んに研究さ れた。動物細胞や組織の培養技術は確立されていなかった当時、細菌を宿主として容易に増 殖させることができ、ヒトに直接病原性を示さないウイルスであるファージは、モデル生物 として適していた [4]。ファージの研究によって、ウイルスの研究、タンパク質の研究、遺 伝子の研究で重大な発見がいくつもなされてきた。例えば、1952 年には Hershey, A.D.と Chase, M. が、ファージの核酸またはタンパク質を 35S や 32P でラベルして宿主細菌への 感染を観察し、遺伝物質が核酸であることを突き止めた[5]。同じく 1950 年代に、λファー ジや T2 ファージの大腸菌への感染効率が菌株(例えば
E. coli
K 株とE. coli
B 株)によっ て異なる現象が発見され、制限酵素の発見につながった。Luria, S. らファージグループと 呼ばれる 3 人の研究者が、ウイルスの複製機構と遺伝的構造を明らかにしたことで、1969 年にノーベル生理学・医学賞を受賞し、また 1976 年には Fiers, F. らが、MS2 ファージの RNA から世界初の全ゲノム配列の決定を行い、1977 には φ174 ファージのゲノム DNA を2
モデルに、Sanger, F. F. らが全ゲノム配列の決定を成功させている[6,7]。このように、フ ァージの研究は、ウイルスだけでなく遺伝子やタンパク質の研究において重要な発見に繋 がり、遺伝学や初期の分子生物学の研究において大きな役割を果たしてきた。ほかにも、P1 ファージの Cre レコンビナーゼによる loxP 配列の部位特異的組換えを利用した Cre/loxP システムや、遺伝子組換えによって M13 ファージの外側にさまざまなタンパク質を連結し、
タンパク質同士や核酸などとの間の相互作用を検討するファージディスプレイ法など、研 究に用いられる実験ツールへの応用もなされてきた[8, 9]。2013 年に発表された遺伝子ノッ クアウト、ノックイン技術である CRISPER-Cas9 システムも、細菌、古細菌がファージや 外来プラスミドを Cas9 ヌクレアーゼで切断する獲得免疫であったことからファージの研究 による成果と言って良いかもしれない。
更に、ファージの構造を理解するために、1940 年代以降は透過型電子顕微鏡による観察 が盛んに行われ、1960 年代にはネガティブ染色が普及、同時期に TEM の画像から三次元 画像再構成が行われ、ファージの詳細な構造の理解が進むなど、TEM の発展とともにファ ージの理解も進んだ。また、1958 年に Kendrew, J. C. らがミオグロビンの X 線結晶構造解 析を行い、この技術は長径が 100Å~1µm 程度のファージを詳細に理解するために、現在 に至るまでファージの研究に利用されている。同様に 1992 年の 434 ファージのサブユニッ トの NMR による構造解析がなされ、また 2010 年代に T4 ファージで Cryo-EM による巨 大構造の決定がなされるなど、ファージの研究は電子顕微鏡の進歩やタンパク質の構造解 析技術の進歩において、それらの技術の発展に関わってきた。
研究の他に、工業や医療の分野などでも応用がなされてきた。例えば、T7 ファージの遺 伝子を利用した組み換え体タンパク質の大量発現は、タンパク質の研究の他に、インシュリ ンなどの有用なタンパク質の工業的な大量生産に利用されている。また、特定の宿主にのみ 感染する宿主特異性は、細菌の分類・検出法であるファージタイピング法に応用され、2011 年には FDA(Food and Drug Administration, USA)の認可を受けて、患者の血液からメチ シリン抵抗性/非抵抗性
S. Aureus
を検出する臨床検査薬が市販された [10]。ファージセラ ピーの研究も盛んである。1917 年に d'Herelle, F. によって提唱され、1942 年にペニシリン の開発によって下火になったファージセラピーの研究だが、医療現場で近年問題となって いる多剤耐性菌感染症の治療法として再度注目された。2009 年には慢性静脈性下肢潰瘍の 治療において、二次感染の原因菌に特異的なファージを混合したファージカクテルが予防 薬として臨床的に使用され、同年に多剤耐性緑膿菌による慢性耳炎に対して特異的なファ ージの投与による症状の改善が見られた[10, 11]。2016 年には、多剤耐性A. baumanii
によ る日和見感染が重篤化した患者に対して特異的なファージを腹腔投与し、ファージの全身 投与による治療成功例が報告された(パターソン症例)。米国を中心に多剤耐性菌感染症の 新規治療薬として、ファージセラピーへの関心が高まっている。加えて、ファージセラピー と同様にファージの殺菌力を応用して、食中毒菌Listeria monocytogenes
の増殖を抑える食 品添加物や、植物病原菌による作物の病害を防ぐ農薬、米国で炭そ菌やボツリヌス菌などに3
よるバイオテロ対策としての軍事利用など、さまざまな検討がなされていた[12-15]。
このように、100 年に渡って続けられたファージの研究は、遺伝子、タンパク質、ウイル スの研究の進歩に大きく貢献し、様々な応用もなされてきた。一方で、現在でも未発見のフ ァージが大量に存在すると予想される。またタンパク質の構造解析の発展に寄与したとさ れるファージの構造既知のサブユニットは、Protein Data Bank (PDB) への登録数は 2000 件を超える一方で、既知のファージが 1500 種近く、詳細が明らかになっていないサブユニ ットが多い。以上から、ファージは現状で分かっていないことが多く存在していると言える。
これまでの経緯から、未発見のファージの同定や、ファージを構成するサブユニットの構造 解析など、ファージの更なる研究を行うことで、新たな発見やファージに由来する新たな応 用につながると期待できる。本研究では、これまでに明らかになったファージの全体構造や 構造既知のサブユニットの比較から、多くのファージで外観が類似したネックやテイルフ ァイバーについて、サブユニットの配置を明らかにしてファージの間で比較することによ って新たな発見があると考えて研究を行った。古典的なファージの 1 つである Mu ファー ジのネックとテイルファイバーのサブユニットを対象に、配置の予想と構成するサブユニ ットの特定を行ったものである。第 1 章では、本研究の背景と目的の詳細を紹介した。
1.1.2 ファージの分類と構造 1.1.2-1 ICTV による分類
全ての生物は、タンパク質合成に関わり比較的進化が遅い 16SrRNA の、1500base ほど の配列の比較によって、系統的に分類されている[16]。一方で、ファージはゲノム上に rRNA の配列を無い上に、塩基配列の変異の頻度が生物と比較して高く、マーカー遺伝子 が発見されていない。そのため、ファージを塩基配列の比較のみによって分類することは 難しい。
ファージで現在主流となっている分類法は、ICTV(国際ウイルス分類委員会:
International Committee on Taxonomy of Viruses)の定める、塩基配列、形態、増殖の過 程、宿主による分類である。これまでに 1442 の Species に分類された。ICTV によって全 ウイルスは 5560 の Species に分けられているが、ファージはその 4 分の 1 を占める多様 な存在である。1442 種のファージはさらに 2 つの Order、27 の Family、その他に分類さ れている。1442 Species のファージの多くは Caudovirales Order に属する。Caudovirales はカプシドの外部にテイルと呼ばれる感染装置を持つファージであり、更に複数の Family と多数の Species に分類されている。一部の性質が近い Species は、Mu-like や T5-like な どの Genus に纏められている。(図 1.1)[2]。
4
1.1.2-2 テイルを持つファージである Caudovirales と、3 つの Family Caudovirales Order
テイルを持つファージである Caudovirales Order は、ファージの個体数の 9 割を占める と言われ、代表的なファージの分類である。Caudovirales に属するファージは、核酸を除く 全ての構造がタンパク質で構成されており、ヘッド、ネック、テイル、テイルファイバーの 構造領域を持つ(図 1.2)。ヘッドはウイルスにおけるカプシドを指し、複数のサブユニット で形成された中空の構造体で、内部にファージのゲノム DNA を格納している。ネックはヘ ッドとテイルを繋げる領域であり、ファージのゲノム DNA が出入りできる狭い穴が開い た、筒状のタンパク質複合体である。テイルは Caudovirales を定義づける構造であり、細 菌への感染を可能にする感染装置である。テイルファイバーはテイルの末端から延びる繊 維状の構造であり、宿主細菌の特異的な表面構造を認識して吸着する。図 1.1 に示した ICTV の分類によると、それぞれの Family に属するファージの Species は 340、735、169 が同定 されており、同定済みの 1442 Species に占める割合で 9 割近くを占める主要なファージで ある。T 系ファージやλファージなどが特に有名であり、研究が続けられている。
Myoviridae
Family、Siphoviridae
Family、Podoviridae
Family負染色したファージの透過型電子顕微鏡を用いた観察により、ファージのテイルの形態 が 3 つ存在することが判明した。テイルの形状から、Caudovirales Order に属するファージ は 3 つの Family に分類された。
Myoviridae
Family は、長いテイルを持ち、感染の際にテイ ルが構造変化を起こして収縮する(収縮性の)ファージである。Siphoviridae
Family は、長 いテイルを持ち、感染の際にテイルが収縮しない(非収縮性の)ファージである。Podoviridae
Family は、他の 2 つの Family のファージと比較して明らかに短いテイルを持ち、感染の際 にテイルが収縮しないファージである(図 1.2)。同じ Family に属するファージの間でテイルの外観は類似しているが、塩基配列や遺伝子 配座、宿主や近年明らかになってきた構造サブユニットの立体構造の面では異なる場合が 多々見られ、そのため現在でも、ファージの分類については議論の対象となっている[17, 18]。
5
図 1.2 Caudovirales の 3 つの Family について、透過型電子顕微鏡画像(上)と模式図(下)
で特徴を示した。
Siphoviridae
20nm
Podoviridae
テイルファイバー テイル ネック
ヘッド ゲノム DNA
Myoviridae
図 1.1 バクテリオファージの分類について、左から Order、Family、Species を記載した。Order は 当てはまる Species が多い Caudovirales と Ligamenvirales のみとし、他は Unclassified として 記載した。また、Species は数のみ記載した[2]。
6
1.1.3 Caudovirales Order に属するファージの感染と増殖の過程 溶菌性ファージと溶原性ファージ
ファージは特定の細菌にのみ感染し、宿主細菌のタンパク質合成能力を乗っ取り娘ファ ージを作らせる。作られた娘ファージは宿主細菌の細胞壁を分解して外部に放出され(溶 菌)、次の細菌に感染することで増殖を繰り返す。Caudovirales Order に属するファージの 場合、感染と増殖の過程は 2 通り存在し、どちらの過程を経るかによって溶原性ファージ と溶菌性ファージに分けられる。感染して細胞質内に侵入したファージのゲノム DNA をも とにして娘ファージが形成されて溶菌し、感染と溶菌の過程を短いサイクルで繰り返すフ ァージを、溶菌性ファージと呼ぶ。一方で、感染後に宿主細菌のゲノム DNA にファージの ゲノム DNA を組み込んで潜伏し、紫外線照射などの外部からの刺激によって娘ファージの 形成が起こるファージを、溶原性ファージと呼ぶ。溶原性ファージの中には、感染後に宿主 細菌のゲノム DNA を一部持ち去る性質を持ち、遺伝子の水平伝搬の担い手となっているも のがおり、代表的な例としては、ベロ毒素の遺伝子を運ぶ赤痢菌ファージなどが存在する。
なお、溶原性ファージと溶菌性ファージの分類は、全体的な構造の差異によって分類された
Myoviridae、Siphoviridae、Podoviridae
の 3 つの Family の分け方とは関連が見られず、そ れぞれの Family に溶菌性ファージと溶原性ファージの両方が見られる。テイルファイバーによる宿主の認識
以下の図 1.3 に、溶原性ファージの感染と増殖の過程を説明した。
宿主細菌とファージが出会うと、まずテイルファイバーが細菌の表面構造を認識して結 合する。テイルファイバーが認識できる構造は様々であり、例えば
Myoviridae
に属す T4 ファージは大腸菌の LPS と OmpC を認識して結合し、Siphoviridae
に属するλファージは LamB を認識する[19,20]。テイルファイバーが宿主を認識すると、ファージのテイルの構 造変化や、ファージのサブユニットの酵素活性、宿主の膜タンパク質の利用などによって、宿主細菌の細胞壁を通過してファージのゲノム DNA を宿主の細胞質に送り込む経路を形 成する。テイルの構造変化が最も顕著な Family は
Myoviridae
で、テイルチューブを囲んで いるテイルシースが構造変化を起こして収縮し、内部のテイルチューブが宿主細菌にむか って押し出され、機械的な圧力で細菌に穴を空ける。Siphoviridae
はベースプレートの中心 に存在するテイルスパイクの酵素活性によって、Podoviridae
はテイルチューブの酵素活性 によって、それぞれ宿主細菌の細胞壁に穴を空ける例が報告されていた[21,22]。溶原性ファージのゲノムDNAの取り込み
宿主細菌の細胞壁に穴が空くと、それまでヘッドに押し込められていたファージのゲノ ム DNA が穴を通って宿主細菌の細胞質に送り込まれる。このとき、ファージのネック領域 のサブユニットも構造変化を起こしてファージのゲノム DNA の放出を可能にすると言わ れるが、はっきりと確認はされていない。ファージのゲノム DNA が細菌の細胞質に取り込
7
図 1.3 溶原性ファージのライフライクル
まれることで、感染が成立する。取り込まれたファージのゲノム DNA は、宿主細菌のゲノ ム DNA に組み込まれる。この状態のファージのゲノム DNA をプロファージと呼び、プロ ファージは宿主細菌の細胞分裂の際にコピーされて受け継がれる。プロファージを持つ細 菌を溶原菌と呼び、感染からプロファージの形成の流れを溶原サイクルと呼ぶ。
娘ファージの自律的形態形成と溶菌
溶原菌の培養液に抗菌薬や熱を加えるなど、溶原菌の周辺環境が生育に適さない状況に なると、プロファージのサブユニットが宿主のタンパク質合成能力によって発現する。図 1.3 の左側のサイクルに移行し、これを溶菌サイクルと呼ぶ。一斉に発現したファージの構 造サブユニットは決まった順序で自律的に結合し、娘ファージが形成される。この発現から 娘ファージの形成の過程を、自律的形態形成と呼ぶ。構造サブユニットと同時に、溶菌に関 する酵素(細胞膜の内膜に細孔を形成するHolin、ペプチドグリカンを分解する Lysin、細 胞膜の外膜を分解する Spanin など)が発現する。娘ファージの自律的形態形成と同時に、
これらの蓄積や酵素活性の制御によって、宿主細菌の溶菌が発生する。外部に放出された娘 ファージが新たに宿主細菌に感染し、溶原化する。溶菌までのサイクルを繰り返すことによ り、もしくは溶原菌の細胞分裂によってプロファージが伝播することで、溶原化ファージは 増殖する。
8
1.1.4 Caudovirales に属するファージの自律的形態形成
変異体解析やゲノム解析、電子顕微鏡による観察やサブユニットの立体構造の決定など、
これまでのファージの様々な研究から、ファージの自律的形態形成の過程は、Caudovirales の間で大まかな流れが共通していることがわかっている。一例として
Myoviridae
の自律的 形態形成の過程を図 1.4 に示した[23,24]。宿主細菌のタンパク質合成能力によって一斉に 合成されたサブユニットは、ヘッドとテイルに分かれて別個に形態形成する。テイルの末端 の繊維状の構造をテイルファイバーと呼び、ヘッドとテイルを連結する円筒状の構造をネ ックと呼ぶ。本論文では、ファージの構造を、上記のヘッド、ネック、テイル、テイルファ イバーの 4 つの構造領域に分けて扱った。ヘッドの形態形成は、ネックサブユニットであるポータルサブユニットの形態形成によ って開始される。ポータルサブユニットが重合してリングを形成し、このリングを土台とし て、足場タンパク質と呼ばれるサブユニットが形態形成し、足場を覆うようにカプシドのサ ブユニットが重合する。その後足場タンパク質は分解され、ポータルに結合したパッケージ ングモーターによって、中空のカプシドにファージのゲノム DNA が詰められる。規定の長 さのゲノム DNA がカプシドに詰められたら、パッケージングモーターが解離して、ネック の構造サブユニットであるコネクターがポータルに結合する。コネクターはポータルと同 様にリング構造であり、しかし何らかの仕組みによって中心の孔からゲノム DNA が漏出す ることを防いでいる。以上の経緯で
Myoviridae
のヘッドが形態形成される。Myoviridae
のテイルの形態形成は、最初にベースプレートと呼ばれる円盤状のタンパク質複合体の形成され、ベースプレートから複数本の細い繊維状のテイルファイバーが伸び、
また円盤の中心軸からテイルチューブと呼ばれる構造が伸びる。テイルチューブは多数の サブユニットが重合した筒状の構造であり、その長さはテープメジャーと呼ばれるサブユ ニットによって決定される。テイルチューブの外側を覆うように、更にテイルシースのサブ ユニットが筒状に重合する。テイルシースは
Myoviridae
に属するファージの感染の際に構 造変化を起こして収縮し、内側の円筒であるテイルチューブを宿主細菌に向かって機械的 に押し出す働きを持つ。決められた長さまで重合が進んだテイルチューブやテイルシース の末端に、ネックの構造であるテイルターミネーターが結合して、テイルが形成される。テ イルターミネーターはテイルチューブやテイルシースの伸長を停止する働きを持つため、テイルターミネーターを欠損した変異体ファージは、形態形成したヘッドと、長さが一定で ないテイルを形成する。
ファージの形態形成の流れは大まかには以上のように進行する。ただし
Siphoviridae
に はテイルシースは存在せず、Podoviridae
にはテイルチューブが無く、ベースプレートに対 応する構造も他の 2 つの Family のような円盤状の構造は形成しない。同じ Family のファ ージであっても、例えばMyoviridae
である T4 ファージと P2 ファージの間で、T4 ファー ジは 5 つのネックサブユニットがポータル、コネクター、テイルターミネーターの役割に 分かれてネックを形成するが、P2 ファージはポータルに対応するサブユニットのみでネッ9
クを構成している。このように、ファージの形態形成は Species ごとに多様性があることが わかっている。
1.1.5 ファージのネックサブユニットの数や配置のバリエーション
ファージはヘッド、ネック、テイル、テイルファイバーの構造領域で構成される。この うちヘッド、テイル、テイルファイバーの外観は 3 つの Family において多様であり、一 方でネック領域だけは、共通して細い円筒構造である。ネックの中心軸には、感染の過程 で宿主の細胞内にファージのゲノム DNA を送るために穴が開いている。
近年の Cryo-EM の進歩に代表されるような構造解析技術の進歩や、変異体解析とゲノ ム解析を含む 100 年近いファージの研究により、ネックを構成するサブユニットの配置や 数には、様々なバリエーションがあることがわかってきた。例えば同じ
Myoviridae
Family に属する T4 ファージと P2 ファージのネックにおいて、構造サブユニットの数や配置は大 きく異なっていた(図 1.5)[23, 26, 27, 28]。全体の構造が円筒構造でどのファージでも共 通しているにも関わらず、構成するサブユニットは P2 ファージの 1 種類、P22 ファージ や T7 ファージの 2 種類、SPP1 ファージやλファージの 4 種類、T4 ファージの 5 種類と 多様であった。1 種類のサブユニットで形成できる構造を多種のサブユニットで形成する 意味や必要性は大きな疑問であり、その解決は、ファージの構造の多様性への理解につな がると思われる。また、円筒構造という決まった構造を様々なパターンで形成しているネ ックを研究することは、タンパク質間相互作用のモデルとしての意義があるだろう。立体構造については、既知の構造は少ないため法則を見出すには至らないが、共通した 部分が見られた。まず、ヘッドの末端に位置するポータルサブユニットは、既知の構造全
図 1.4
Myoviridae
の自律的形態形成の一例10
てで相同性が見られた。PDB には、
Myoviridae
である T4 ファージ(T4 gp20)、Siphoviridae
である G20C ファージ、SPP1 ファージ(SPP1 gp6)、HK97 ファージ、Podoviridae
である T7 ファージ、P22 ファージ(P22 gp1)、Phi29 ファージのポータルサブユニットの構造が登録されており、Dali server で検索したところ T4 gp20 の構造に対し て Cα rmsd が 4~6 程度と、有意な相同性があることがわかった。一方で、ポータルの 直下に位置するサブユニットについて T4 ファージは構造が不明であったため SPP1 ファ ージのネックサブユニット gp15 を対象に相同性検索を行ったところ、HK97 gp6 のみがヒ ットし、同じ
Siphoviridae
で構造既知のλ gpW は前述のように相同性が見られなかっ た。これについて、ヘッドを構成する構造既知のサブユニットでは、T4 ファージ、HK97 ファージ、T5 ファージ、T7 ファージなどで構造に相同性が見られた。また、テイルは 3 つの Family で大きく構造が異なる。以上から、構造に相同性があるヘッドサブユニットと 隣接しているポータルサブユニットは多くのファージにおいて構造が相同であり、一方で 全体に大きく構造が異なるテイルに近いネックサブユニットには多様な構造が見られるの ではないかと予想した。1.1.6 バクテリオファージの立体構造の決定 1.1.6-1 ファージの構造解析の進歩
ファージの構造を理解する研究は、1930 年代に透過型電子顕微鏡が開発されて大まかな 外観を知ることができるようになって以来、技術発展と共に進められてきた。当初はヘッド とテイルを判別することが出来た程度であったが、1960 年代に負染色したファージの観察 が可能になり、ネックやテイルファイバーの存在が確認できたことで、ファージを 3 つの
図 1.5 ネックを構成する構造サブユニットの数や配置は多様である
11
Family に分類することができた。1980 年代以降は X 線結晶構造解析、少し遅れて NMR に よるタンパク質の構造解析が可能になり、T7 発現システムの開発などによりファージを構 成するサブユニットの原子レベルでの構造解析が可能になった。近年では電子顕微鏡や検 出器、解析法の進歩から、従来の X 線結晶構造解析などの個々のサブユニットの構造解析 と組み合わせることで、ファージのヘッド全体やテイル全体のような、巨大なタンパク質複 合体の構造解析が可能になってきた。
これまでにファージのサブユニットは、Protein Data Bank (PDB) に 2000 個程度が登録 された。同定済みの 1500 Species がそれぞれ数個~数百個の ORFs を持つため、代表的な ファージである T-even ファージ(T1~T7 ファージ)やλファージなどの一部のファージ を除いて、構造解析はあまり進んでいないと言える。しかし、ファージの個々のサブユニッ トや複合体の立体構造の蓄積によって、ファージ全体が持つ性質を(ごく一部だが)予想す ることが可能になってきた。
1.1.6-2 Caudovirales の 3 つの Family の間での、サブユニットの立体構造の相関 ヘッドのサブユニットの立体構造は一致している
これまでに構造が決定された 2000 ほどのファージ由来の構造サブユニットを比較するこ とによって、Caudovirales 全体としてヘッドの構造サブユニットの立体構造がほぼ一致して いることがわかってきた。T4 ファージ(
Myoviridae
)の gp23 は、ヘッドの大部分を構成す るサブユニット(Major head subunit と呼ばれる)である。この T4 gp23 は、Siphoviridae
である T5 ファージ、Podoviridae
である T7 ファージのそれぞれで構造に相同性が見られ た。また同じくヘッドを構成するサブユニット(Head vertex protein)は T4 ファージ、HK97 ファージ、T5 ファージで構造に相同性が見られた(図 1.6)。構造解析がなされたヘッドサ ブユニットは少ないが、ヘッドを構成するサブユニットは Family を跨いで構造に相同性が ある例が複数存在していることがわかっていた。テイルのサブユニットの立体構造は一致する例が少ない
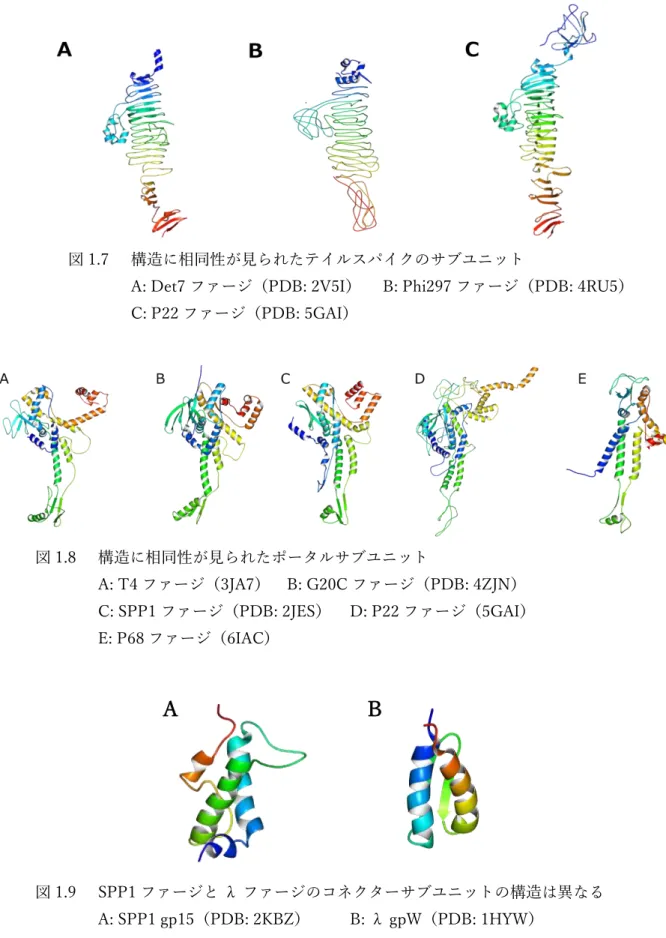
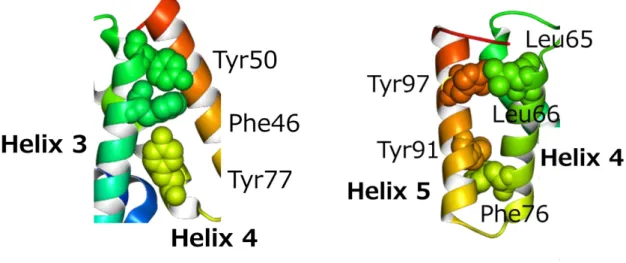
一方で、テイルを構成するサブユニットには Family を超えた構造の相同性が見られる例 は(現状では)少ない[25]。3 つの Family はテイルの構造の比較から分類されており、全 体構造として共通点が少ないためであると思われる。3 つの Family で共通した構造が見ら れた例としては、Det7 ファージ(
Myoviridae
)、Phi297 ファージ(Siphoviridae
)、P22 ファージ(
Podoviridae
)の間で、サブユニットの構造に相同性が見られた。テイルスパイクは、テイルの構造の中で例外的に 3 つの Family に共通して存在する構造であることから、
テイルスパイクを構成するサブユニットの立体構造が一致していることは納得できる(図 1.7)。
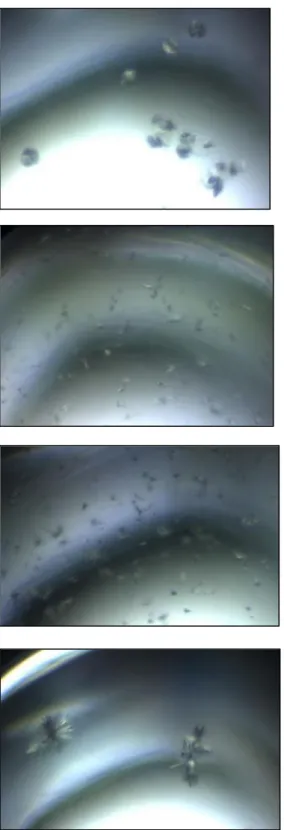
12 ネックはポータルサブユニットの構造が一致
異なる Family に属するファージの間で、ヘッドの構造サブユニットは立体構造が一致す る例が多く報告されており、一方でテイルの構造サブユニットは例が少ない。全体的な構 造を見ても、ヘッドは正二十面体が殆どであり、一方でテイルの構造は 3 つの Family に 分けられる。同じ構造のヘッドと様々な構造のテイルを、ネックはどのような仕組みで連 結しているのだろうか。
構造既知のネックサブユニットは、ほとんどがポータルサブユニットであった。
Myoviridae である T4 ファージ、Siphoviridae である G20C ファージ、SPP1 ファージ、
Podoviridae である P22 ファージ、P68 ファージの立体構造が PDB に登録されており、全 て構造に相同性が見られた(図 1.8)。このことから、3 つの Family で同じ構造のサブユニ ットで構成されたヘッドと隣接したポータルサブユニットは、ほぼ同一の構造である可能 性が高いだろう。一方で、コネクターやターミネーターで構造既知のサブユニットは比較 できるほどの数が無く、しかし SPP1 ファージとλファージのコネクターサブユニットは 異なる立体構造が PDB に登録されていた(図 1.9)。このことから、ポータルサブユニッ トの構造は一致し、ポータルサブユニットと連結するコネクターから、ファージごとに構 造の差が発生する可能性があった。しかし前述のとおり構造既知のサブユニットが少な く、何かしらの法則を見出すことができない。
図 1.6 構造に相同性が見られたヘッドサブユニット
A~C:Major head subunit D~F:Head vertex protein
A: T4 gp23 (PDB: 5VF3) B: T5 gp10 (PDB: 5TJT) C: T7 ファージ (PDB: 3J7W) D: T4 gp24 (PDB: 1YUE) E: HK97 (PDB: 3QPR) F: T5 ファージ (PDB: 6OKB)
13
図 1.7 構造に相同性が見られたテイルスパイクのサブユニット
A: Det7 ファージ(PDB: 2V5I) B: Phi297 ファージ(PDB: 4RU5)
C: P22 ファージ(PDB: 5GAI)
図 1.8 構造に相同性が見られたポータルサブユニット
A: T4 ファージ(3JA7) B: G20C ファージ(PDB: 4ZJN)
C: SPP1 ファージ(PDB: 2JES) D: P22 ファージ(5GAI)
E: P68 ファージ(6IAC)
図 1.9 SPP1 ファージと λ ファージのコネクターサブユニットの構造は異なる A: SPP1 gp15(PDB: 2KBZ) B: λ gpW(PDB: 1HYW)
A B
14 1.1.7 ネックサブユニットの配置は様々である
ヘッドに隣接するポータルサブユニットは立体構造が一致していた。一方で、テイルに近 いネックサブユニットの立体構造は、3 つの Family の間だけでなく、Species ごとに違いが ある可能性が示された。私は、この構造の違いから、同じような構造のヘッドを多様なテイ ルに連結するために、ファージごとにどのよう仕組みを持っており、ファージの間でどのよ うな法則性があるのか知りたいと考えている。ネックの構造の差から何らかの法則性を知 るには、多数のファージのネックを構成するサブユニットの立体構造を決定し、比較する必 要がある。しかし、現在の技術では多数の構造解析を行うには 10 年単位の時間や人手が必 要である。
一方で、ネックに存在するファージの Species ごとの違いは、サブユニットの立体構造だ けではない。前述したように、先行研究の変異体解析やゲノム解析、その他の電子顕微鏡に よる観察などから、ネックを構成するサブユニットの数や配置が Species ごとに多様である ことがわかっている(図 1.5)。例えば
Myoviridae
である T4 ファージは 5 種類のサブユニ ットから構成されたネックを持つ一方で、同じMyoviridae
である P2 ファージは 1 種類の サブユニットで構成されたネックを持っている。Siphoviridae、Podoviridae
にもそれぞれ特 徴的なサブユニットの構造が見られる。1.1.8 ネックサブユニットの配置から得られる知見
ヘッドとテイルの形態形成の最終段階を担うネックの理解は、ファージの形態形成を理 解するうえで重要である。様々なファージにおいて、ファージの構造中でのネックサブユニ ットの配置を知ることで、その共通点から各ファージの間でネックの形態形成が類似した 過程であるか、予想することが出来ると考えた。
また、多くのファージの間で立体構造の決定ではなく、サブユニットの配置の差を比較す ることで、何らかの法則性の発見に繋がるかもしれない。例えば、ファージは塩基配列の変 異が発生しやすいため、塩基配列から遺伝的な距離が理解しづらいが、サブユニットの立体 構造には保存が見られる。ネックサブユニットの数や配置の類似性も保存されると思われ ることから、ファージの進化について知見が得られるかもしれない。
以上の知見がネックサブユニットの配置から得られると考えたこと、加えて、テイルと異 なり Family 間で外観が円筒構造と類似していたことから、サブユニットの配置の比較が Family を超えて可能であったことが、ネックを研究対象とした理由の 1 つであった。更に、
多様なファージのネックの配置の決定によって、ネックサブユニットの構造解析の際に効 率的にネックサブユニットを研究できるはずであり、将来的には構造解析の進歩によって、
筆者が本来知りたかった「同じ構造のヘッドと異なる構造のテイルを連結する円筒構造が ファージごとにどのような仕組みになっているのか」の疑問が解ける日が来ると期待し、ま ずはファージの 1 種、Mu ファージについて、ネックの配置の検討を試みることにした。
15 1.1.9 テイルファイバーの研究
多くの Caudovirales Order に属するファージは、テイルファイバーが宿主細菌の表面構 造を特異的に認識して結合することで感染する。また、テイルファイバーは繊維状の三量体 構造を形成しており、ファージ由来のシャペロンか分子内のシャペロンドメインの存在に よって、発現の際に球状にならずに正確なフォールディングを行う[32-52]。テイルファイ バーの宿主認識部位はファージごとに異なっており、細菌に特異的に結合できる。構造解析 はドラッグデリバリーシステムへの応用や、タンパク質間(またはタンパク質 - LPS 間)
の相互作用のモデルとしての意義がある。このように、テイルファイバーはファージの感染 機構を知るうえで重要な研究対象である。
本研究でネックサブユニットの配置を決定することに決めた。その理由は前述したよう に、どの Family のファージでも類似した構造を持つネックのサブユニットの数やその配置 がファージによって異なることについて、この差が発生する理由や仕組みを知りたいと考 えたことである。テイルファイバーも同様に、どのファージにおいても繊維状の類似した構 造を持ち、Caudovirales の殆どで比較検討が可能であった。また、T4 ファージは gp34~
gp37 の 4 つのサブユニットから構成されたテイルファイバーを持ち、T5 ファージは pb1 と pb132 の 2 つのサブユニットから構成されたテイルファイバーを持つように、構成する サブユニットの数や配置の違いがあることがわかっており、その比較からネックと同様の 知見が得られる可能性があった。以上から、テイルファイバーを構成するサブユニットにつ いても研究することにした。
1.1.10 構造サブユニットの配置の決定 1.1.10-1 立体構造の決定による配置の決定
ファージの構造サブユニットは多種多様な原子構造を持つが、異なるファージの間で一 部の構造サブユニットが同じである例が多数報告されていた。以下の図 1.10 に示した SPP1 ファージとλファージのサブユニットの構造はその一例である。SPP1 ファージとλ ファージはともに
Siphoviridae
に属するファージであり、一方で宿主は異なっている。SPP1 ファージは
Bacillus subtilis
に感染し、λファージはEscherichia coli
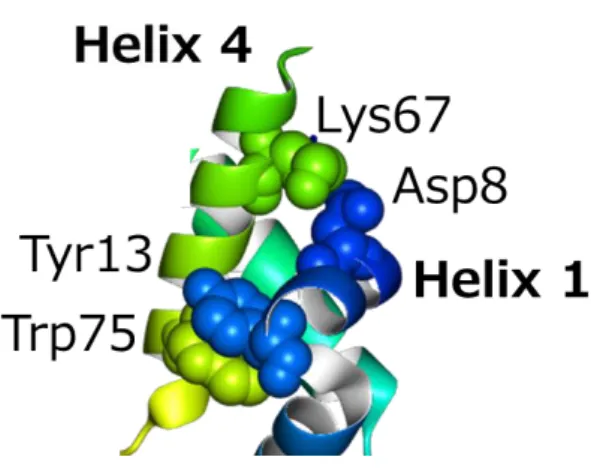
に感染する。SPP1 ファージはネックを構成する全ての構造サブユニットの立体構造が決定され(図 1.10 左に示した)、このうち gp17 と呼ばれるサブユニットは、λファージのネックサブユ ニット gpU と構造に相同性があることがわかっていた。SPP1 gp17 とλ gpU はともにテ イルと隣接しており、またどちらも重合して 6 量体のリングを形成するなど、サブユニッ トが同じ配置にあった[29, 30]。SPP1 gp17 とλ gpU の例のように同じ構造を持つサブユ ニットが同じ配置にあることは一般に見られることであるため、本研究において Mu ファ ージの個々のサブユニットの立体構造を決定し、構造既知のファージのサブユニットの中 から構造に相同性があるサブユニットを見つけることにより、それぞれのサブユニットが ネックにおいてどのような配置にあるかを決定することに繋がる。
16
1.1.10-2 複合体形成によるサブユニットの配置の理解
個々のサブユニットの構造解析を行う一方で、本研究では複合体の形成を試みた。ファ ージの感染の過程で、一斉に発現した構造サブユニットは自律的形態形成を経て娘ファー ジを形成する[31]。サブユニットの結合の順序はタンパク質間相互作用によって制御され ており、非特異的な結合はほぼ発生しないと言われている。この性質から、結合が見られ るサブユニット同士は、ファージの構造において隣接したサブユニットであると言える。
さらに、安定な複合体の構造解析に成功すれば、単独のサブユニットの構造解析よりも天 然に近い状態の構造を得ることができる。これにより構造サブユニットの配置を知ること ができるほか、サブユニット同士の相互作用部位の決定、サブユニットの重合度の決定な ど、様々な情報を得ることができる。
一方で、複合体の構造解析は一般に難しいとされる。図 1.11 に概念図を示した。自律的 形態形成において、最初の段階である単量体は不安定であり、他のサブユニットとの結合 による安定化が最終産物である娘ファージの形成の動力であると言われる。実際に、単量 体のサブユニットは時間経過によって変性や凝集を起こしやすい一方で、最終産物である 娘ファージは、非常に安定である。同様に、自律的形態形成の途上にある会合体や準安定 な複合体も、解離や凝集が発生する。結晶化が可能なほどに安定な複合体がどのサブユニ ットの組み合わせによって得られるかはファージごとに異なる。
以上の経緯から、本研究では複合体を形成するネックサブユニットの組み合わせを探索 し、これによって隣接したサブユニットを決定し、ネックにおけるサブユニットの配置を 予想することを目標とした。加えて、結晶化可能な複合体を得る前段階として、安定な複 合体を形成する実験的な手法の検討を行った。
図 1.10 SPP1 ファージと λ ファージの間で共通した原子構造を持つ 構造サブユニットが存在する
17 1.2 Mu ファージの研究の背景
1.2.1 Mu ファージ
Myoviridae
に属するファージは有名なもので T4 ファージ、P2 ファージ、P1 ファージ、Sf6 ファージ、Det7 ファージなどが存在し、Mu ファージもまた代表的な
Myoviridae
ファージである。当研究室では Mu ファージの全構造サブユニットの立体構造の決定を目 指し、20 年近く研究を続けている。テイルのサブユニットを中心に研究しており、これま でにほぼ全てのテイルサブユニットの単離精製を行い、その一部は立体構造を決定するこ とに成功した。Mu ファージは、Myoviridae
Family に属するファージの 1 つである(図 1.12 左)。電子顕微鏡画像による観察から、Mu ファージは図 1.12 右に示した構造を持つ ことがわかっていた。正二十面体のヘッド、ヘッドとテイルを連結する円筒状のネック、収縮性の長いテイル、短いテイルファイバーを持っている。Mu ファージは変異体解析と ゲノム解析から 1 部のネックサブユニットの役割と配置が予想されており、しかし全体の 配置は不明であった。
Mu ファージのネックサブユニットは、Mu gp29、Mu gp35、Mu gp36、Mu gp37、Mu gp38 の 5 つから構成されると予想されていた。T4 ファージのネックサブユニットが 5 つ であり、現在ネックサブユニットが判明しているファージの中では最多である。T4 ファ ージのネックサブユニットのうち、コネクターを構成する T4 gp13 と T4 gp14 は、交互に 円状に結合することでリングを形成するという、他のファージでは見られない独特な形態 形成をすることがわかっており、T4 ファージと同じく 5 つのサブユニットで構成される Mu ファージのネックサブユニットも、やはり独特の形態形成を見せるのではないかと期 待した。
図 1.11 自律的形態形成によって単量体が安定な複合体を形成する概念図
18
また、Mu ファージのテイルファイバーは、2 つ存在し、どちらかが発現することで Mu ファージは大腸菌を含む広範囲の腸内細菌に感染することができる[31, 42-45]。この 2 つ のテイルファイバーの遺伝子は gp49 と gp52 であると言われ、どちらかのテイルファイバ ーが発現する際に同時に gp50 または gp51 が発現することがわかっていた。2 つのサブユ ニットが同時に発現することで、Mu ファージのテイルファイバーは 2 つのサブユニット で形成されるのか、もしくは片方のサブユニットは他のファージで見られるようなシャペ ロンであるのか、本研究で明らかにすることにした。
以上の経緯から、Mu ファージのネックとテイルファイバーを対象に、構造解析と複合 体の形成を試みることにした。以下に Mu ファージの性質や研究の歴史、Mu ファージに ついてこれまでに当研究室で明らかにしたことをまとめた。
1.2.2 Mu ファージの研究の歴史
Mu ファージは、1950 年にカリフォルニア大学バークレー校の学生だった Taylor, L. に よって、P1 ファージの溶原菌から偶然発見された。Taylor, L. らの研究によって、Mu フ ァージが自身のゲノム DNA を宿主細菌のゲノム上のランダムな位置に組み込む性質があ ることが判明した。トランスポゾンの最初の発見例として研究対象になったほか、Mu フ ァージの遺伝子導入の位置によっては欠損変異体が作られることから、自然宿主である大 腸菌において変異体作成に利用された歴史がある。Mu ファージの名称の由来はこのよう な Mutator としての性質から来たものである [44-46]。1985 年に M. M. Howe らが変異 体解析によって 24 の必須遺伝子を同定したほか、2003 年には G. J. Morgan らが Mu ファ ージの 36,717bp に渡る全塩基配列を決定し、55 個の ORFs の同定とゲノム解析によるア ノテーションを行った[47-49]。当研究室では 2000 年代前半から Mu ファージの全構造サ 図 1.12 バクテリオファージ Mu の全体構造
(左)負染色した Mu ファージの電子顕微鏡画像[45]
(右)Mu ファージの模式図
1.0Å
19
ブユニットの立体構造の決定を目指して研究を開始した。Mu ファージのテイルの構造サ ブユニットを中心に、組み換え体タンパク質として網羅的に単離精製を行い、2005 年には テイルの構造サブユニットである gp44 の立体構造を高解像度で決定し、2013 には同じく テイルの構造サブユニットである gp45 の立体構造の決定に成功した。本研究は 2013 年か ら Mu ファージのネックに注目して、ネックの構造サブユニットを対象に研究を開始し、
研究の過程でテイルファイバーも含めて網羅的な単離精製と構造解析を試みた。
1.2.3 Mu ファージの遺伝子
Mu ファージは、先行研究によって多数の変異体が分離されており、必須とされる遺伝 子が同定されていた[47]。また全塩基配列が決定され、構造既知のファージのアミノ酸配 列との間で有意な相同性が(一部の ORFs を除いて)見られないことがわかっていた。ゲ ノム解析によって 55 個の Open Reading Frames(ORFs、gene1~gene55)が同定済みで
(表 1.1)、アノテーションが行われた[48, 49]。遺伝子産物は gene product に番号をつけ て gp1~gp55 と呼ばれる。図 1.13 のように Mu ファージの遺伝子マップを線状で示した 際の各構造領域の位置を記載した。
図 1.13 Mu ファージの遺伝子マップ
全塩基長は 36,717bp であり、55 個の ORF が同定されていた。図に示 した遺伝子の配置は、溶原化した Mu ファージの遺伝子配座を 5’末端側を左、3’
末端側を右にして記載したもので、5’末端に AttL、3’末端に AttR が存在する。
20 表 1.1 Mu ファージの遺伝子の一覧
ネックを構成すると予想された遺伝子を黄色で示し、テイルファイバーを構成すると 予想された遺伝子は緑色で示した。
21 表 1.1 Mu ファージの遺伝子の一覧
ネックを構成すると予想された遺伝子を黄色で示し、テイルファイバーを構成すると 予想された遺伝子は緑色で示した。
22 1.2.4 Mu ファージの構造既知のサブユニット
構造未知サブユニットについて立体構造を決定することは、Mu ファージの形態形成や、
感染の作用機序などの生命現象を詳細に理解することに繋がる。しかし、Mu ファージの全 体構造はある程度予想されていた一方で、個々のサブユニットの詳細な構造や機能はほと んど分かっていなかった。研究当初、構造が決定されていたサブユニットは、転写制御因子 gp1 と gp2 [50,51]、転移酵素 gp3、gp4 [52,53]、中期プロモータータンパク質 gp17[54]、
ベースプレートの構造サブユニット gp44、gp45 [55, 37]の 7 サブユニットだけであった。
7 つの構造既知のサブユニットのうち、構造サブユニットは gp44 と gp45 の 2 つだけであ り、また Mu gp44 と Mu gp45 は当研究室で構造解析に成功したサブユニットである。
1.2.5 当研究室における Mu ファージの研究の経緯と研究成果 1.2.5-1 当研究室における Mu ファージの研究の背景
当研究室では現在に至るまで 20 年近く Mu ファージの研究を続けている。当研究室で Mu ファージの研究を始めた当時は、ヒトゲノム計画の完了の前後の時期であり、全塩基 配列が決定されたファージは多くなかった。Mu ファージは全塩基配列の決定がなされた 数少ないファージの 1 つであった。Mu ファージの自然宿主に大腸菌が含まれ、組み換え 体タンパク質として Mu ファージのサブユニットを大量発現することが容易であった。更 に前述したように、多数の変異体が分離され、変異体解析とゲノム解析によって遺伝子の アノテーションがある程度なされていた。このことから、当時はファージの構造解析のモ デル生物として Mu ファージは適当なターゲットだったと言える。
1.2.5-2 本研究を開始した当時の背景と、Mu ファージのテイルの研究
2005 年以降に次世代シーケンサーが開発されたことで、ウイルスの全塩基配列の決定と ゲノム解析は容易となった。本研究を開始した 2013 年当時は、発見済みのファージはほ ぼ全塩基配列が決定され、データベースで簡単に検索することができた。2000 年代前半に あった Mu ファージのアドバンテージは、多数の変異体が存在すること、自然宿主である 大腸菌によって容易に組み換え体タンパク質を得られることを除いて、あまり大きくない と言って良いだろう。
一方で、当研究室では当時までに 15 年近く Mu ファージを研究してきたことで、ノウ ハウや設備が揃っていた。特に、宿主細菌に Mu ファージのゲノム DNA を取り込ませる 働きをするテイルについては、テイルを構成するサブユニットは大半が組み換え体タンパ ク質として大量発現する系が構築済みであり、単離精製条件も多くが決定されていた(図 1.14)。感染の際にテイルの 2 重円筒構造の外側の円筒であるテイルシースが構造変化を起 こして収縮し、内側のテイルチューブが押し出される機械的な動きによって宿主細菌の細 胞壁を貫通するテイルの動作は、タンパク質性のナノマシンのモデルとして注目してい た。テイルの仕組みを理解するため、当研究室では構造サブユニットの立体構造の決定を