膜性腎症は,中高年に発症するネフローゼ症候群の原因 の第 1 位を占め,慢性の経過で発症して持続性蛋白尿から ネフローゼ症候群に進展する。腎組織では腎糸球体基底膜 の外側に沿って,IgG と補体成分 C3 の免疫沈着物が顆粒 状に沈着することを特徴とする。これに続いて顆粒状の免 疫沈着物の間に,従来の糸球体基底膜から連続するように 新たな基底膜が生成されて,糸球体基底膜のスパイク病変 と肥厚が起こる。腎糸球体基底膜に沿う免疫沈着物の出現 に伴って,腎糸球体上皮細胞の足突起に癒合と消失が,さ らに細胞間スリット膜にも形態変化が起こり,これに伴っ て大量の蛋白尿が出現する。長年にわたって不明であった ヒト膜性腎症の病因抗原として,2002 年に中性エンドペプ チダーゼが1),2009 年に M 型ホスホリパーゼ A 2 受容体 が2),2010 年にアルドラーゼ還元酵素〔アルド−ケト−還元 酵素ファミリーメンバー 1(AKR1B1)〕およびミトコンド リアスーパーオキシドジスムターゼ 2(SOD2)が3)相次いで 報告された。この発見により,Heymann 腎炎,BSA 腎炎な どの実験腎炎を用いて行われてきた膜性腎症の発症機序, 腎組織障害機序の解析の一部を,ヒトで直接に行うことが 可能となった。さらに,膜性腎症の血清診断,治療のモニ タリングも近い将来に可能となるであろう。 これまで膜性腎症の病因抗原−抗体系の解析は,ヒト膜 性腎症に酷似した糸球体病変を示すラット膜性腎症モデ ル Heymann 腎炎を用いて行われ,膜性腎症に関する多くの 知見と経験をもたらした。 本稿では,Heymann 腎炎の研究から得られた知見を中心 に,ヒト膜性腎症の研究の現状と展望について述べる。 はじめに メガリン:Heymann 腎炎抗原は,1982 年にラット近位尿 細管刷子縁蛋白中に発見され,1994 年にメガリンとして同 定された4)。メガリンは,名前の通りに巨大で,約 4,398 個 のアミノ酸で構成され,4 個のリガンド結合部位を持つ一 本鎖の糖蛋白である。メガリンは,LDL 受容体スーパー ファミリーに属する多機能受容体蛋白で,近位尿細管刷子 縁に局在し,尿中アルブミンをはじめとする多くの分子の 細胞内取り込みを担っている5)。また,メガリンはラット でのみ腎糸球体上皮細胞に発現していて,ラットに特異的 な膜性腎症抗原である。 腎糸球体上皮細胞内で産生されたメガリンは,小胞によ り細胞内を運ばれ,足突起基部で小胞と細胞膜が癒合し, 細胞膜上の小陥凹である clathrin coated pits に表出される。 この巨大蛋白の腎炎惹起抗原部位の探索では,メガリン蛋 白の細胞外ドメイン N 端 1−563(分子量 6 万)のアミノ酸 が,蛋白尿を伴う Heymann 腎炎を惹起することが明らかに された6)。さらに短いアミノ酸 1−1957),1−2367),157−2368) でも Heymann 腎炎が誘導されることから,メガリンの腎炎 惹起抗原部位は,細胞外の N 端に近いリガンド結合部の三 次構造にあると考えられる。 M 型ホスホリパーゼ A2受容体(PLA2R):Beck らによ り,ヒト膜性腎症抗原として PLA2R が報告された2)。この 蛋白はヒト型マンノース受容体ファミリー蛋白 4 個のう ちの一つである。ヒト型マンノース受容体ファミリー分子 は,通常は細胞膜上から細胞内に取り込まれてリサイクリ ングされている。PLA2R は,細胞外ドメインの N 端末に システインに富む部位を持ち,続いて 1 個のⅡ型フィブロ ネ ク チ ン ド メ イ ン, 8 個 の C 型 レ ク チ ン 様 ド メ イ ン (CTLD),細胞膜貫通部位,細胞内ドメイン,を持つ分子 量 18.5 万(非還元状態で)の一本鎖糖蛋白である。この分子 病因抗原
Pathogenic mechanism of membranous glomerulonephritis obtained from Heymann nephritis and human membranous glomerulonephritis
北里大学医学部腎臓内科
膜性腎症の病因Ⅱ
実験腎炎からみたヒト膜性腎症
鎌
田
貢
壽
特集:膜性腎症
の腎炎抗原部位は,現在のところ CTLD の 4,5,6 番目に あると推定されている。この分子内のジスルフィド結合を 切断すると抗原性が消失するので,腎炎抗原は CTLD の三 次構造にあると考えられる。PLA2R は,ヒト腎糸球体上皮 細胞に多く表出されるとともに,尿細管にも表出されるが, その細胞内での局在や機能はまだ不明と言わざるをえな い。一方,ラットでは腎糸球体上皮細胞での PLA2R の発 現は,わずかにとどまる。 中性エンドペプチダーゼ(NEP):Devic らが,ヒト膜性 腎症抗原として報告した1)。NEP を欠損した母親が妊娠に 伴い抗 NEP 抗体を産生し,母親の抗 NEP 抗体が新生児に 移行して膜性腎症を発症させたとされている。新生児は腎 不全を伴う重篤なネフローゼ症候群に進展したが,抗 NEP 抗体の減少に伴って回復した。NEP は,9∼11 万の分子量 を持つ亜鉛依存性メタロプロテアーゼで,ネプリシン,エ ンケファリナーゼ,CD10,EC3,4,24,11 と同一である。 NEP は,細胞膜上で蛋白を切断して蛋白情報を消去する役 割を果たしている。また NEP は,ヒトの腎臓では糸球体上 皮細胞,尿細管刷子縁,血管平滑筋細胞に発現している。 アルドラーゼ還元酵素,スーパーオキシドジスムターゼ 2 (SOD2):Prunotto らにより,膜性腎症の病因抗体の標的抗 原として報告された3)。この抗原については本稿では詳述 しない。 これらの膜性腎症抗原は,いずれも糸球体上皮細胞に発 現しているが,メガリン,PLA2R,NEP は,腎糸球体上皮 細胞の足突起基部の clathrin coated pits に局在することが 共通している。Heymann 腎炎では,血流中から原尿中に 入った病因抗体は,糸球体上皮細胞足突起基部の clathrin coated pits でメガリンと会合して in situ で免疫複合体を形 成することが判明している9)。PLA2R,NEP の腎糸球体上 皮下での免疫複合体形成機序はまだ明らかにされていない が,メガリンと同様な機序が想定される。 Heymann 腎炎は,ラット腎近位尿細管刷子縁蛋白 Fx1A (メガリンを豊富に含む)ないしはメガリン蛋白をラットに 直接に投与して腎炎を誘導する能動型 Heymann 腎炎と,こ の抗原を家兎に投与して産生させた病因抗体をラットに投 与して腎炎を誘導する受動型 Heymann 腎炎とがある。 Heymann 腎炎の腎組織病変を形成し,蛋白尿を出現させ る病因抗体は抗メガリン抗体である。能動型 Heymann 腎炎 ラットで,腎糸球体上皮下沈着物中の病因抗体の IgG サブ 病因抗体 クラスを,マウス抗ラット IgG サブクラス単クローン抗体 で調べた結果では,糸球体沈着性の病因抗体はラット IgG2a 抗体が主体で,他のサブクラス抗体に比して常に優 位な沈着を示した(表)10)。ラット IgG2b 抗体と IgG1 抗体 は弱い蛍光強度で沈着するが,これらが単独で沈着するこ とはみられない。また,ラット IgG2a 抗体に加えて IgG2b 抗体あるいは IgG1 抗体の沈着を伴う場合には,尿蛋白量 が多い傾向がみられる。ラット腎糸球体への IgG2c 抗体の 沈着はみられない。糸球体に沈着する免疫グロブリン軽鎖 はκ鎖であり,λ鎖の沈着はみられない。また,蛋白尿の 出現には,糸球体基底膜に補体成分 C3 が少なくとも部分 的顆粒状に沈着することを必要としている。ラット IgG1, IgG2a,IgG2b 抗体は,補体成分 C1q と結合し,古典的経 路で補体活性化を起こすが,IgG2c には C1q の結合能力が ない。以上から,能動型 Heymann 腎炎では,古典的経路に よる補体活性化が想定される。 一方,ヒト特発性膜性腎症の糸球体に沈着する IgG サブ クラスは IgG4 が優位であり,IgG1,IgG2 の沈着もみられ る。これらの IgG サブクラスの補体活性化経路は,IgG1 が古典的経路,IgG2 が副経路をとるとされる。一方,IgG4 は C1q の結合能がなく,その補体活性経路は不明である。 びまん性全節性糸球体病変を示す膜性腎症の糸球体には C1q の沈着はみられず,mannose-binding lectin(MBL),C3c, C4d,factor B の沈着がみられる11)。MBL は,補体活性化 レクチン経路の活性化因子であり,IgG2 は factor B を介し て副経路を活性化させることから,びまん性膜性腎症では IgG2 と IgG4 が副経路およびレクチン経路を介して補体を 活性化させると想定されている11)。これに対して,分節性 糸球体病変を示す膜性腎症では,糸球体沈着物中に IgG1, IgG3,補体 C1q がみられ,古典的経路を介した補体の活性 化が想定されている11)。 Heymann 腎炎ラットの蛋白尿出現には,腎糸球体基底膜 沈 着 物 中 に 補 体 late component C5b−9(membrane attack complex:MAC)が形成されることが必須とされている。ヒ ト膜性腎症でも,腎糸球体基底膜沈着物中に MAC が存在 することから,MAC はヒトでも蛋白尿出現に重要な役割 を演じていると考えられる。 能動型 Heymann 腎炎ラットは実験 8 週から 40 週以降 までネフローゼ症候群レベルの蛋白尿を示す(図 1)10)。こ のラット血清中の IgG 抗 Fx1A 抗体価は,4 週から 14 週ま ではコントロール群に比し有意な高値を示すが,16 週以降 は実験終了の 40 週までは腎炎と蛋白尿が持続しているに もかかわらず,コントロール群との差がみられない。また,
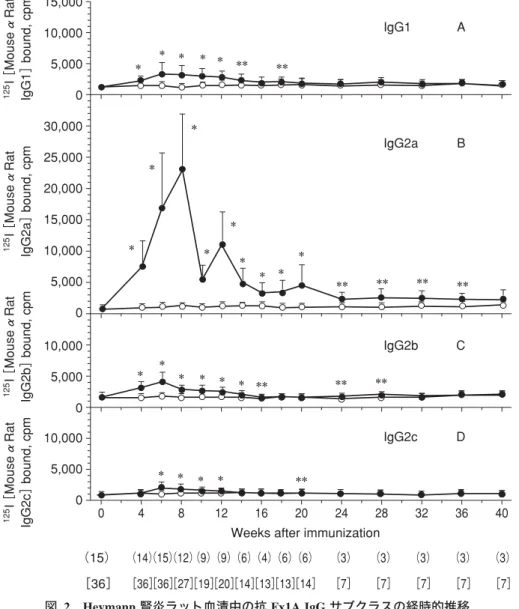
血清中の抗 Fx1A 抗体 IgG サブタイプ(図 2)は,IgG2a 抗 Fx1A 抗体価が他の IgG サブクラス抗体価に比して 6.9∼ 11.2 倍の高値を示した。8 週からのネフローゼレベルの蛋 白尿出現に伴って,血清中 IgG2a 抗 Fx1A 抗体価は低下し 始め,40 週には差がみられなくなる。ヒト膜性腎症におい ても,蛋白尿出現から一定期間が経過すると,血清中に病 因抗体が存在しても,その抗体価が低値のために検出が困 難になる場合が想定される。ヒト膜性腎症でも,すでに血 清中抗 PLA2R 抗体の測定が開始されているが2,12),まだ半 定量的な測定にとどまっており,感度,特異度の高い測定
表 マウス抗ラット IgG サブクラス,κ,λ単クローン抗体を用いた Heymann 腎炎ラット糸球体への IgG アイソタイプ,
軽鎖,C3 沈着の評価(蛍光抗体法による) 40 wks 20 wks 12 wks 8 wks 6 wks Sacrificed at: 7 7 6 7 8 Number of experiment: 3.86±0.38(7)* 1.71±0.95(3)* 2.71±0.76(6)* 1.29±0.49(2)* 0.00±0.00(0) 0.43±1.13(1) 0.00±0.00(0) 1.00±0.00(0)* 3.71±0.76(7)* 1.43±1.13(2)* 2.86±0.38(7)* 1.43±0.53(3)* 0.00±0.00(0) 1.71±1.38(4) 0.00±0.00(0) 1.00±0.00(0)* 4.00±0.00(6)* 1.17±0.98(1) 2.83±0.41(6)* 1.83±0.41(5)* 0.00±0.00(0) 2.00±0.63(5)* 0.00±0.00(0) 0.83±0.41(0) 3.71±0.49(7)* 1.57±0.53(4)* 2.86±0.38(7)* 1.14±0.38(1)* 0.00±0.00(0) 2.43±0.53(7)* 0.00±0.00(0) 0.86±0.38(0)* 2.63±0.92(7)* 0.13±0.35(0) 2.50±0.76(7)* 0.50±0.53(0) 0.00±0.00(0) 2.25±0.89(6)* 0.00±0.00(0) 0.63±0.52(0) IgG IgG1 IgG2a IgG2b IgG2c κ λ C3 Mean score of glomerular IF intensity IgG1 IgG2a IgG2b IgG2c κ λ Statistical table using Mann-Whitney’s U test 蛍光強度 0:陰性,蛍光強度 1:巣状分節性沈着,蛍光強度 2:軽度びまん性沈着,蛍光強度 3:中等度びまん性沈着,蛍 光強度 4:強度びまん性沈着 ( ):蛍光強度 2 以上(典型的 Heymann 腎炎)のラット数 *p<0.05 vs コントロールラット,§p<0.01,#p<0.05,Mann-Whitney U テストによる。 § § § § § § § § § § § § § # § # # § § § § § # # § # 1,000 100 10 1 0.1 Proteinuria(mg/day) 0 4 8 12 16 20 24 28 32 36 40
Weeks after immunization (15) [36] [36] [36] [28] [21] [20] [14] [14] [14] [14] [7] [7] [7] [7] [7] (15) (15) (12) (9) (9) (6) (6) (6) (6) (3) (3) (3) (3) (3) ** ** ** ** ** ** * * * * * * * 図 1 Heymann 腎炎ラット蛋白尿の推移 ●:Heymann 腎炎ラット,○:コントロールラット,↓:Fx1A 投与(1 回目 120μg,2 回目 60μg) ( ):コントロールラット数,[ ]:Heymann 腎炎ラット数,平均±SD
系の開発が待たれる。
能動型 Heymann 腎炎では,ラットの系統により明らかに 疾患感受性が異なる。Lewis(LEW)ラット,LOU ラット, Brown Norway(BN)ラ ッ ト に Fx1A 300μg を 投 与 す る と13),実験 16 週には LEW ラット 6 匹中 6 匹が大量の蛋 白尿と典型的 Heymann 腎炎組織像を呈する。一方,BN ラットは 6 匹中 6 匹が蛋白尿も Heymann 腎炎組織像も示 疾患感受性 さず,腎炎は発症しない。LOU ラットは 6 匹中 4 匹が Hey-mann 腎炎組織像を呈し,うち 2 匹が有意な蛋白尿を示し た。3 種のラット血清中 IgG の抗原認識能に差異は認めな かった。血清中抗体価は,LEW ラットが一時的に高い値を 示すものの,LOU,BN ラット間に差はなく,3 種のラッ トで十分な血清中抗体価が認められた。疾患感受性の差異 の原因として,糸球体での抗原発現量の差異,糸球体沈着 物中の IgG サブタイプの差異などが不明のまま残されて いる。また,ラットでは,各系統の主要組織適合性抗原が 一定でないために,遺伝的背景が疾患感受性に及ぼす影響 0 4 8 12 16 20 24 28 32 36 40 15,000 10,000 5,000 0 10,000 5,000 0 10,000 5,000 0 30,000 25,000 20,000 15,000 10,000 5,000 0 IgG1 A B C D IgG2a IgG2b IgG2c 125 I[ Mouse αRat IgG1] bound, cpm 125 I[ Mouse αRat IgG2a] bound, cpm 125 I[ Mouse αRat IgG2b] bound, cpm 125 I[ Mouse αRat IgG2c] bound, cpm
Weeks after immunization (15) [36] [36] [36] [27] [19] [20] [14] [13] [13] [14] [7] [7] [7] [7] [7] (14) (15) (12) (9) (9) (6) (4) (6) (6) (3) (3) (3) (3) (3) * * * * * * * * * * * * * * * * * * * * * * * * ** ** ** ** ** ** ** ** ** **
図 2 Heymann 腎炎ラット血清中の抗 Fx1A IgG サブクラスの経時的推移 固相ラジオイムノアッセイ。抗原量 50μg/mL Fx1A-DOC。5 倍希釈血清を用いた。
125I−モノクロナール抗ラット IgG サブクラス抗体を用いて抗体価を求めた。
パネル A:IgG1 抗 Fx1A 抗体価,パネル B:IgG2a 抗 Fx1A 抗体価,パネル C:IgG2b 抗 Fx1A 抗体価,パネル D:IgG2c 抗 Fx1A 抗体価
( ):コントロールラット数,[ ]:Heymann 腎炎ラット数,*:p<0.01,**:p<
の解析は困難である。 ヒト膜性腎症の疾患感受性は,日本人では HLA-DR2, HLA-DRB5*0101,欧米人では DR3 と関連するとされてき た。一方,今回の PLA2R1 の発見に続いて,Stanescu ら14) は,膜性腎症患者全ゲノムの単塩基多型についてフランス 人,ドイツ人,イギリス人を対象に調べたところ,膜性腎 症の発症と 2 つの対立遺伝子(PLA2R1 の遺伝子を含む染 色体 2q24 の対立遺伝子と染色体 6p21 上の HLA-DQA1 対 立遺伝子)に相関がみられた。さらに,この 2 つの対立遺 伝子に同型接合体変異がみられた場合の疾患発症リスク は 78.5 倍と報告している。この結果から Stanescu らは, PLA2R1 の変異蛋白に対する免疫応答を HLA-DQA1 対立 遺伝子の変異蛋白が促進させている可能性を指摘してい る。 Heymann 腎炎抗原メガリンは,腎糸球体上皮細胞足突起 の基部の小陥凹である clathrin coated pits で細胞膜上に表 出される。一方,血中から糸球体基底膜を通過して糸球体 上皮細胞下に到達した抗メガリン抗体は,clathrin coated pits でメガリンと出会い免疫複合体を作る。この免疫複合 体は,糸球体基底膜外透明層に放出されるが,一部は coated pits と接している。この状態で免疫複合体の形成が 繰り返されて免疫複合体が成長し,上皮細胞足突起間ス リット膜の直下に到達する9)。この免疫複合体は,腎糸球 体基底膜の構成成分であるⅣ型コラーゲン,フィブロネク チン,ラミニンと強く結合し,腎糸球体基底膜に固着して 免疫沈着物となる。 ヒト膜性腎症でも,同様の免疫沈着物形成機序が想定さ れるが,今後は PLA2R1 と抗 PLA2R1 抗体による腎糸球体 基底膜上皮側の免疫沈着物形成機序の解析が待たれる。 ヒト膜性腎症抗原が発見された今,実験腎炎から得られ た情報をヒト膜性腎症で検証することが可能となった。さ らに,膜性腎症の血清診断,血清モニタリングの実現など が期待できる。 利益相反自己申告:申告すべきものなし 文 献
1.Devic H, Guigonis VG, Mougenot B, Decobert F, Heymann JP, Bensman A, Deschenes G, Ronco PM. Antenatal membra-nous glomerulonephritis due to anti-neutral endopeptidase 腎糸球体基底膜上皮側での免疫沈着物の形成機序
antibodies. N Engl J Med 2002;346:2053−2060.
2.Beck LH, Bonegio RGB, Lambeau G, Beck DM, Powell DW, Cummins TD, Klein JB, Salant DJ. M-type phospholipase A2
receptor as target antigen in idiopathic membranous nephropathy. N Engl J Med 2009;361:11−21.
3.Prunotto M, Carnevali ML, Candiano G, Murtas C, Bruschi M, Corrandini E, Trivelli A, Magnasco A, Petretto A, Santucci L, Mattei S, Gatti R, Scolari F, Kador P, Allegri L, Ghiggeri GM. Autoimmunity in membranous nephropathy targets aldose reductase and SOD2. J Am Soc Nephrol 2010;21:507−519. 4.Saito A, Pietromonaco S, Loo AK, Farquhar MG. Complete
cloning and sequencing of rat gp330/“megalin,”a distinctive member of the low density lipoprotein receptor gene family. Proc Natl Acad Sci USA 1994;91:9725−9729.
5.Saito A, Sato H, Iino N, Takeda T. Molecular mechanisms of receptor-mediated endocytosis in the renal proximal tubular epithelium. J Biomed Biotechnol 2010;2010:403272. 6.Oleinikov AV, Feliz BJ, Makker SP. A small N-terminal 60−
kD fragment of gp600(Megalin), the major autoantigen of active Heymann nephritis, can induce a full-blown disease. J Am Soc Nephrol 2000;11:57−64.
7.Tramontano A, Knight T, Vizzuso D, Makker SP. Nested N-ter-minal megalin fragments induce high-titer autoantibody and attenuated Heymann nephritis. J Am Soc Nephrol 2006;17: 1979−1985.
8.Tazaki H, Kamata K, Naito S, Okamoto T, Kobayashi K, Yamanaka N, Aoyama T. Passive Heymann nephritis induced by genetic immunization using megalin cDNA fragments. Kitasato Med J 2011;41:69−75.
9.Kerjaschki D, Farquhar MG. Immunocytochemical localization of Heymann nephritis antigen(gp330)in glomerular epithelial cells of normal Lewis rats. J Exp Med 1983;157:667−686. 10.内田満美子,鎌田貢壽.Heymann 腎炎の病因抗体 IgG アイ
ソタイプの長期観察.北里医学 1994;24:357−367. 11.Segawa Y, Hisano S, Matsushita M, Fujita T, Hirose S,
Take-shita M, Iwasaki H. IgG subclasses and complement pathway in segmental and global membranous nephropathy. Pediatr Nephrol 2010;25:1091−1099.
12.Stahl R, Hoxha E, Fechner K. PLA2R autoantibodies and recurrent membranous nephropathy after transplantation. N Engl J Med 2010;363:496−498.
13.西村真里子.膜性糸球体腎炎の疾患感受性機序の解析.北 里医学 1995;25:468−470.
14.Stanescu HC, Arcos-Burgos M, Medlar A, Bockenhauer D, Kottgen A, Dragomirescu L, Voinescu C, Patel N, Pearce K, Hubank M, Stephens HAF, Laundy V, Padmanabhan S, Zawadzka A, Hofstra JM, Coenen MJH, Heijer M, Kiemeney LALM, Bacq-Daian D, Stengel B, Powis SH, Brenchley P, Fee-hally J, Rees AJ, Debiec H, Wetzels JFM, Ronco P, Mathieson PW, Kleta R. Risk HLA-DQA1 and PLA2R1 alleles in
idio-pathic membranous nephropathy. N Engl J Med 2011;364: 616−626.