高磁場中における日本酒酵母の 活動に関する研究
松下研究室
03232022 黒木 章太郎
平成 19 年 2 月 23 日
電子情報工学科
目次
第
1章 序論
1 1.1
はじめに・・・・・・・・・・・・・・・・・・・・・・・・・・・
1 1.2酒造方法・・・・・・・・・・・・・・・・・・・・・・・・・・・
1 1.3酵母の働き・・・・・・・・・・・・・・・・・・・・・・・・・・
2 1.4磁場が及ぼす影響・・・・・・・・・・・・・・・・・・・・・・・
41.4.1
磁場配向・・・・・・・・・・・・・・・・・・・・・・・・・
61.4.2
磁気力・・・・・・・・・・・・・・・・・・・・・・・・・・
71.4.3 MHD
効果・・・・・・・・・・・・・・・・・・・・・・・・
71.4.4
磁気対流・・・・・・・・・・・・・・・・・・・・・・・・・
81.5
冷凍機冷却型超伝導マグネット・・・・・・・・・・・・・・・・・
8 1.6本研究の目的・・・・・・・・・・・・・・・・・・・・・・・・・
9第
2章 実験
12 2.1試料準備・・・・・・・・・・・・・・・・・・・・・・・・・・・
12 2.2磁場装置・・・・・・・・・・・・・・・・・・・・・・・・・・・
13 2.3振とう装置・・・・・・・・・・・・・・・・・・・・・・・・・・
14 2.4全菌数測定・・・・・・・・・・・・・・・・・・・・・・・・・・
15 2.5生菌数測定・・・・・・・・・・・・・・・・・・・・・・・・・・
17第
3章 結果と検討
19 3.1静置培養・・・・・・・・・・・・・・・・・・・・・・・・・・・
193.1.1
全菌数測定・・・・・・・・・・・・・・・・・・・・・・・・
193.1.2
生菌数測定・・・・・・・・・・・・・・・・・・・・・・・・
213.2
振とう培養・・・・・・・・・・・・・・・・・・・・・・・・・・
243.2.1
温度による変化・・・・・・・・・・・・・・・・・・・・・・
273.2.2
磁場強度による変化・・・・・・・・・・・・・・・・・・・・
343.3
再現性・・・・・・・・・・・・・・・・・・・・・・・・・・・・
40第
4章 まとめと今後の課題
44 4.1まとめ・・・・・・・・・・・・・・・・・・・・・・・・・・・・
44i
4.2
今後の課題・・・・・・・・・・・・・・・・・・・・・・・・・・
45参考文献
47ii
表目次
2.1.
生菌数測定の希釈率・・・・・・・・・・・・・・・・・・・・・・
173.1. 10 T
中
15℃下における静置培養全菌数測定値・・・・・・・・・・・
20 3.2. 10 T中
15℃下における静置培養生菌数測定値・・・・・・・・・・・
21 3.3. 0 T中
25℃下における振とう培養測定値・・・・・・・・・・・・・・
253.4. 10 T
中
30℃下における振とう培養測定値・・・・・・・・・・・・・
273.5. 10 T
中
25℃下における振とう培養測定値・・・・・・・・・・・・・
293.6. 10 T
中
20℃下における振とう培養測定値・・・・・・・・・・・・・
313.7.
近似式の定数値
(1)・・・・・・・・・・・・・・・・・・・・・・・・
313.8.
各温度における増殖スピード・・・・・・・・・・・・・・・・・・・
333.9. 5 T
中
25℃下における振とう培養測定値・・・・・・・・・・・・・・
353.10. 1 T
中
25℃下における振とう培養測定値・・・・・・・・・・・・・
373.11.
近似式の定数値
(2)・・・・・・・・・・・・・・・・・・・・・・
393.12.
各磁場における増殖スピード・・・・・・・・・・・・・・・・・・
393.13. 5 T
中
25℃下における再現性実験測定値・・・・・・・・・・・・・
413.14.
同条件での増殖スピード比較・・・・・・・・・・・・・・・・・・
43iii
図目次
1.1.
日本酒の酒造工程・・・・・・・・・・・・・・・・・・・・・・・・
21.2.
酵母の電子顕微鏡写真・・・・・・・・・・・・・・・・・・・・・・
31.3.
酵母の代謝ルート・・・・・・・・・・・・・・・・・・・・・・・・
31.4.
ビオ・サバールの法則・・・・・・・・・・・・・・・・・・・・・・
51.5.
微小円柱に対する電流要素・・・・・・・・・・・・・・・・・・・・
61.6.
冷凍機冷却型超伝導マグネット・・・・・・・・・・・・・・・・・・
91.7.
高磁場下での大腸菌生菌数推移・・・・・・・・・・・・・・・・・
101.8.
超伝導マグネットの磁場勾配とシャーレの位置・・・・・・・・・・
101.9.
酵母の沈殿分布
(a)シャーレ底の吸光度
(b)沈殿パターンの様子・・・・
112.1.
ピペットマンとエッペンチューブ・・・・・・・・・・・・・・・・
122.2.
冷凍機冷却型
10 T超伝導マグネットの磁場イメージ・・・・・・・・
132.3.
振とう装置・・・・・・・・・・・・・・・・・・・・・・・・・・
152.4.
菌数計算盤・・・・・・・・・・・・・・・・・・・・・・・・・・
162.5.
酵母のコロニー形成・・・・・・・・・・・・・・・・・・・・・・
173.1. 10 T
中
15℃下における酵母全菌数の時間依存性・・・・・・・・・・
20 3.2. 10 T中
15℃下における酵母生菌数の時間依存性・・・・・・・・・・
223.3. 10 T
中
15℃下における規格化酵母全菌数の時間依存性・・・・・・・
233.4. 10 T
中
15℃下における規格化酵母生菌数の時間依存性・・・・・・・
233.5. 0 T
中
25℃下における酵母全菌数の時間依存性・・・・・・・・・・
263.6. 0 T
中
25℃下における増殖期比較・・・・・・・・・・・・・・・・
263.7. 10 T
中
30℃下における酵母全菌数の時間依存性・・・・・・・・・・
283.8. 10 T
中
30℃下における増殖期比較・・・・・・・・・・・・・・・・
283.9. 10 T
中
25℃下における酵母全菌数の時間依存性・・・・・・・・・・
30 3.10. 10 T中
25℃下における増殖期比較・・・・・・・・・・・・・・・・
30 3.11. 10 T中
20℃下における酵母全菌数の時間依存性・・・・・・・・・・
32 3.12. 10 T中
20℃下における増殖期比較・・・・・・・・・・・・・・・・
323.13.
増殖スピード差における温度依存性・・・・・・・・・・・・・・・
343.14. 5 T
中
25℃下における酵母全菌数の時間依存性・・・・・・・・・・
363.15. 5 T
中
25℃下における増殖期比較・・・・・・・・・・・・・・・・
363.16. 1 T
中
25℃下における酵母全菌数の時間依存性・・・・・・・・・・
38iv
3.17. 1 T
中
25℃下における増殖期比較・・・・・・・・・・・・・・・・
383.18.
増殖スピード差における磁場依存性・・・・・・・・・・・・・・・
403.19. 5 T
中
25℃下における再現性酵母全菌数の時間依存性・・・・・・・
423.20. 5 T
中
25℃下における再現性増殖期比較・・・・・・・・・・・・・
42v
第 1 章 序論
1.1
はじめに
日本酒は、昔から伝統的に受け継がれて来た職人達の知識と技術の結晶であ り、日本が誇れる伝統文化の一つと言える。近年は後継者不足や近代文明によ る大量製造により、昔ながらの酒蔵は年々減少している。酒造りは自然の恵み と、大変な労力を必要とするが、それでもこうした背景の中、日本酒の伝統は 絶える事無く、日々、我々の生活を潤している。この日本酒造りに欠かせない アルコール発酵を行うのが酵母である。長い歴史の中で、酵母はあらゆる面か ら研究されてきた。
酵母の姿を人類がはじめて見たのは
1600年代の後半にさかのぼる。オランダ
の
A. van Leeuwenhoekが手製の凸レンズを用いて、発酵中のビールの中に微
小な物体を観察し、発表したのが酵母であった。その後、生物学の研究におい て重宝される事となる。日本における酒造りにおいても、酵母の存在は大変重 要な位置を占めている。それでは、こうした中で生まれる日本酒の酒造方法を まず述べる。
1.2
酒造方法
酒の原料は米・米麹であり、酒の種類によってアルコールを入れたりする。
また、酒造りにおいて重要なのは、水・米・技術・酵母・気候である。これら の要素の最適な組み合わせで、お酒のおいしさが決まる。
ではその酒造りだが、まず米の精白から始まる。米は酒造好適米と呼ばれる 日本酒造りに適した、大粒で心白と呼ばれる白色の不透明な領域が中心部にあ るものが使用される。精米された米は洗米され、水に漬けて吸水される。白米 重量の
3割ほど吸水したところで浸漬が終わり、 蒸す事により蒸米を造りだす。
次に仕込みと呼ばれる段階に移る。定温の中で蒸米に麹カビを散布して混ぜ、
米麹を作る。米麹は麹菌がつくった酵素の働きで米を溶かし、デンプンをブド ウ糖に変えて酵母が食べられるようにする役割を持っている。散布した麹菌の 胞子は、発芽して菌糸が伸び始め、約二日で蒸米を完全に覆う。この麹に水と 酵母、それに蒸米を加えて酒母というものを作る。酒母は酛(もと)ともいい、
1
まさに酒の素になる。酒母は酸性で、酵母が酸に強いのに対して、他のほとん どの有害菌が酸に弱いので、酒母の中では酵母だけが育つのである。その酒母 にさらに麹・蒸米・水を3回に分けて加え、発酵するのを待つ。3回に分ける のは、酒母に大量の物量を添加して仕込むと酒母中の酵母や酸がうすめられ、
雑菌が繁殖するおそれがあるからだ。このため、ある程度の期間をおいて酵母 の増殖をはかりながら仕込んでいく方法がとられている。米の質、麹菌が繁殖 する温度、気温の変化など、それぞれの酒蔵、それぞれの「仕込み」によって 条件・調整法は異なる。タンクの中では麹に含まれる糖化酵素の働きにより、
米のデンプンは糖分に変えられる。その後、酵母の働きにより糖分がアルコー ル発酵する。発酵は
3週間前後で終わり、熟成したもろみをしぼり、酵母など をこし取り、ろ過する事でお酒となる。これらの製法を図
1.1にまとめる。
図
1.1.日本酒の酒造工程
こうして生まれるお酒はとてもデリケートな代物で、これらの工程に少しの狂 いや変化が生じれば、その味は大きく変わってしまう。逆に言えば、その変化 を利用して様々な品質のお酒が全国各地で生まれ、その伝統の味を維持する事 が重んじられている。
1.3
酵母の働き
それでは、この工程の一つであり、酒造りの重要な要素である酵母について、
さらに詳しく述べる。酵母は菌類に属する単細胞の微生物で、細胞の一部から 芽を出して無生殖で増殖する。培養条件により異なるが、
25~
30℃位でよく成 長し、
100~
200分間に一度の割合で細胞分裂をする。大きさは直径が約
4~
5μm
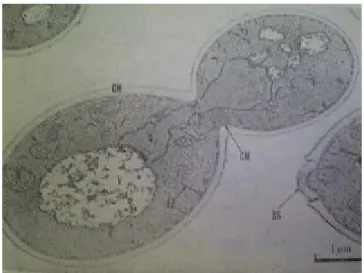
で、おおよそ球形をしている。ここで、酵母の細胞構造を図
1.2に示す
1)。
2
図
1.2.酵母の電子顕微鏡写真
細胞の外側は細胞壁でおおわれ、外層は主としてマンナン
-タンパクからでき ており、その内側にはグルカンの層がある。このグルカンの層が、細胞壁の強 さを保持するのに役立っていると考えられている。細胞壁には以上のマンナン、
タンパク質、グルカンの他に脂質やキチンその他の物質もふくまれている。細 胞分裂後に残る出芽痕にはキチンが多く含まれており、キチン合成が出芽痕形 成、及び細胞分裂に関与している事が分かる。細胞の
1回の分裂が終わってか ら次の分裂が終わるまでの間を細胞周期と呼ぶが、真核細胞である酵母は、こ の周期を正確に一定の順序で進行する。
しかし、酵母はどんな環境下でもアルコールを造り出すのかと言えば、そう ではない。酵母は生命維持のために、様々な酵素を使って糖分であるグルコー スをピルビン酸まで分解する。これは、人間から微生物までがもっている生命 維持のための代表的な代謝ルートである。
その後、ピルビン酸から2方向ヘルートが分かれる。まず、酸素欠乏状態の ときにピルビン酸はアセトアルデヒドを経て、エタノールヘと変えられる。つ まり、アルコール発酵をする。酸素がある状態では酵母は酸素呼吸をするので、
ピルビン酸を炭酸ガスと水、生活に必要なエネルギーを発生させたりしながら、
循環させる。この代謝ルートを図
1.3に示す。
図
1.3.酵母の代謝ルート
3
酵母の性質の一つとして、酸素が有っても無くても生育できる点が挙げられる が、このように酵母に発酵作業を活発にさせる為には、容器上部に泡ができる 事で下部が酸素欠乏状態になる事が条件といえる。実際の酒蔵において、発酵 中の醪では炭酸ガスが発生して酸素欠乏状態となり、これに転落して死亡する 事故も昔は珍しくなかった。このような事から酒蔵には必ず一人、酸素欠乏・
硫化水素危険作業主任者の資格を持った人がいなければならない。
1.4
磁場が及ぼす影響
最近の科学技術の進歩につれて、我々が強い磁場に曝露される機会はしだい に増加している。職業人が曝露されていた加速器、核融合炉、エネルギー貯蔵 マグネット等の他、核磁気共鳴イメージング、磁気浮上超高速列車といった、
いまでは一般市民や傷病者までが曝磁される時代となっている。しかし、磁場 の生体に対する影響、特に人体影響については解明されてはいない。我々人類 は、
X線などの放射線が広く実用化されるにつれて、次第にその犠牲者が増加 してしまった歴史があり、物理的エネルギーを人体影響が解明されないまま長 時間利用すると、場合によっては大変なことになる一つの例と言える。そこで、
磁場を印加する事により生じる諸現象をまず理解する事により、磁場が及ぼす 影響の究明を行う事が必要だといえる。では、まず磁場について次に述べる。
電荷のまわりに電界ができるように、電流が流れている導線のまわりには磁 気的な磁場が生じる。定常電流からは時間的に一定な静磁場ができる。磁場は ベクトルであり、その強さを表すものとして磁束密度
Bを定義する。
Bの単位 はテスラ
(T)であり、
N/(A・
m)の次元をもつ。ここで図
1.4のように、回路
Cの 微小部分を流れる電流
(電流要素
Ids)がまわりの点
Pにつくる磁束密度は、ビ オ・サバールの法則により
2
0 d
d 4
r r s B I
×r
= π
μ
(1.1)
と表せる。ただし、
rは
Idsから点
Pに至る距離、
rrはその方向の単位ベクトル、
μ0
は真空の透磁率である
7 0 =4π×10−
μ N/Α2
(1.2)
とする。
4
I
Ids P
ds θ r
rr
dB
図
1.4.ビオ・サバールの法則
式
(1.1)より、アンペールの右ねじの法則に従って、
dBの方向は
Idsと
rrでつく る平面に垂直であり、向きは
Idsから
rrの向きに右ねじを回すときのねじの進行 方向となる。
dBの大きさは
Idsと
rrのなす角をθとすれば
π θ μ sin
4 d 0 d2
r s B= I
となる。回路
Cの全体を流れる電流が点
Pにつくる磁束密度は、電流要素
Idsからの寄与を回路全体にわたって線積分すれば求まり
∫
×= C r
r s B 0 Id 2
4
r π
μ
となる。ビオ・サバールの法則は、広がりのある空間
Vを電流が流れている場 合に一般化できる。図
1.5のように電流の流れる方向に長さ
ds、断面積
dSの微 小円柱
(体積
dV=dsdS)をとれば、この円柱に対する電流要素は電流密度
iを用い て
V i s S i s
Id =(d )d = d
となる。
5
I
ds
i
図
1.5.微小円柱に対する電流要素
dS
したがって、微小領域
dVを流れる電流要素
idVが
rだけ離れた点
Pにつくる 磁束密度は、
dVから点
Pに向かう方向の単位ベクトルを
rrとすれば、式
(1.1)よ り
2 0 d d 4
r r V B i
×r
= π μ
となり、空間
V全体にわたって体積分すれば
∫
×= V V
r r
B i d
4 2
0 r
π μ
が求まる。この磁束密度
Bと磁場の強さ
Hとの比をその物質の透磁率といい、
式
(1.2)の真空の透磁率を用いれば、次のような関係が成り立つ。磁化を
Mとす
れば、
H = B −M μ0
と表される。
H、
Mの単位は
A/mとなり、磁場の強さは単に磁場あるいは磁界 ともいう。
1.4.1
磁場配向
この磁場が引き起こす種々の現象を述べる。まず、分子や微結晶などに磁気 的な異方性がある場合には、これらの粒子には磁場方向に配向しようとするト ルクが働く。強磁性体のように永久双極子を持つ場合には、外部磁場方向に自 らの内部磁場を向けることで安定に配向する。粒子の磁気的異方性については、
分子の構造の異方性に基づくものと、粒子の形状の異方性に基づくものとに分 類できる。
磁石以外で磁場ともっともよく相互作用するものはコイルである。コイルに
6
磁石を近づけると反対方向の磁化を持とうとして誘導電流が流れる。有機化合 物の中でもっとも理想的なコイルとして働く構造はベンゼン環であるが、それ は自由に動けるパイ電子が平面上の構造をとっている為、コイルのように振る 舞うからである。しかし、分子内に誘導電流が流れているような状態は熱力学 的に不安定なので,ベンゼン環に磁場を作用させると誘導電流が流れず、ベン ゼン環の分子平面が磁場と平行になる向きに磁場配向する。タンパク質の場合 には、ペプチド結合がコイルの働きをし、ペプチド平面が磁場と平行になる向 きに磁場配向しようとする。その為、ペプチド平面がその軸方向に平行に配列 しているアルファ・へリックス構造は磁場方向に配向しようとし,ペプチド平 面が多く平行に配列しているベータ・シート構造も同様の磁場配向作用を示す。
また、芳香性アミノ酸の芳香環も同様である。これより、これら3つの構造が 分子内部で特定の方向に配列しているほど,磁気的な異方性が強いことになる。
故に、膜タンパク質などがもっとも磁気的異方性が強いタンパク質である。
また、分子自体が磁気的異方性を示さない場合でも、粒子の形状が異方的な 場合には磁場配向する。例として、磁気的に等方的な分子でも、その結晶の形 が針状や板状など異方的ならば,結晶自体の帯磁率が方向によって異なるため、
磁場配向する。
1.4.2
磁気力
磁場が一様でなく、位置
zにより磁場の強さ
Hが違うような磁場勾配が存在 する下では磁気力が発生する。物質の統計を
M、モル濃度の磁気の感受性を とすれば、
XM
z H H MX
F M
∂
= ∂
で表される。磁場勾配を上ってゆく方向に磁気力が働く物質は帯磁率が正の強 磁性体および常磁性体で、逆に磁場勾配を下ってゆく物質が反磁性体である。
タンパク質や水をはじめほとんどの有機物は反磁性体であるが、タンパク質結 晶を結晶化容器の器壁に接触させることなく成長させることができれば、結晶 中へのひずみの導入などを防ぐ事ができる為、有力な結晶化方法であると考え られている。
1.4.3 MHD
効果
電荷
Qを持つ荷電粒子が速度 で磁束密度
Bの磁場の中を運動しているとき、
磁場より受ける力は、
v
7
F =Q(v×B)
となり、この力をローレンツ力という。磁場中で電解質溶液に電流を流した場 合、磁場と直角方向に電流が流れるとこのローレンツ力が働くため、溶液には 大きな流れが生じる。これを
MHD(Magneto hydrodynamic)効果と呼ぶ。ロー レンツ力は大きな力なので、多くの磁場効果の中でも早くに見つかった効果で ある。この効果の例として、タンパク質の結晶化溶液に磁場下で電流を流すよ うな場合には、十分大きな影響を及ぼすと考えられている。
また、溶液に電流を流さずとも、溶液中のイオンなどの電荷を持った化学種 が一定の方向に運動することでも、これらの化学種にローレンツ力が働く。溶 液中に引加された電流ではなく、特定化学種のイオン個々にローレンツ力が働 くため、上述の
MHD効果とは区別され、マイクロ
MHD効果と呼ばれている。
例として、結晶が成長している場合には、沖合の溶液から結晶表面へ溶質分子 の流れができる。そのため、溶質分子が電荷を持っている場合には個々の溶質 分子にローレンツ力が働き、分子の拡散運動はローレンツ力の方向に曲げられ る。
1.4.4
磁気対流
1.4.2
で説明した磁気力の他に、系内の帯磁率に不均一がある場合、及び系内
の帯磁率が均一でも磁場に不均一がある場合には,これらに基づき磁気力が働 く。そのため、磁気力に基づいた磁気対流が発生することになる。この効果を うまく応用する事ができれば、系内の対流を抑制できたり、望みの向きに磁気 対流を流し、系内を攪拌させたりすることができる。
以上述べてきたような磁場下での種々の影響がこれまで分かってきたが、生 物に与える影響のように、未だに解明されていない部分もある。
1.5
冷凍機冷却型超伝導マグネット
近年の超伝導分野の発展により、冷凍機冷却型超伝導マグネットを用いて、
10 T
の高磁場が長時間安定して印加できるようになった為、高磁場中での生物 活動を調べる事ができるようになった。図
1.6にこの装置を示す。従来の超伝導 マグネットでは通常、液体ヘリウムで冷却されてきたが、本実験で用いるよう な冷凍機冷却型のマグネットでは、エポキシ樹脂などで含浸して安定性を強化 したものであり、液体ヘリウムによる浸漬冷却は必要としない。この為、この ような冷凍機などによる伝導冷却法は、安定性が増すと共に運転コストが大幅 に削減される。これにより高磁場下での研究が容易になった。
8
図
1.6.冷凍機冷却型超伝導マグネット
1.6本研究の目的
現在、細胞の機能や構造の基本的な調節機構を解明するために、分子生物学 や生理学、さらに遺伝学の研究材料として広く利用されているのが酵母である。
酵母の細胞の構造や、生活環は高等生物に近い共通性をもち、増殖や死滅とい ったサイクルが一定して行われる酵母は、外部影響の効果を確認し易い。こう いった真核生物を研究する事で基礎生物学の発展につながる。この酵母に上述 の冷凍機冷却型超伝導マグネットにより高磁場を印加する。実際に微生物分野 での研究として、図
1.7に示すように、大腸菌を
43℃の
5.2~
6.1 Tの磁場下で
1.5 g/L
のグルタミン酸を加えた
25%の
LB培地中で好気的培養した時、安定期
の細胞数が地磁場に比べて
10万倍多かったという
Horiuchiらの磁場の印加に よる大腸菌死滅の抑制効果の研究や
2)、
9
図
1.7.高磁場下での大腸菌生菌数推移
20 40 60 80 t(h)
Ikehata
らによる、出芽酵母に
5 Tの磁場を
2時間かけた時、図
1.8に示す超伝 導マグネット内の位置により図
1.9で表すような沈殿パターンに違いが見られ た
3)といった報告がされている。
図
1.8.超伝導マグネットの磁場勾配とシャーレの位置
10
図
1.9.酵母の沈殿分布
(a)シャーレ底の吸光度
(b)沈殿パターンの様子 これまで述べてきた酵母や醸造条件の少しの違いで味が変わるお酒の性質上、
高磁場を印加する事で酵母に変化が起これば味が変化する可能性は高く、それ によるお酒造りの新たな品質が生まれる可能性が期待できる。これを踏まえ、
本研究では高磁場を印加して酵母が受ける影響や活動を調べ、研究する事が目 的である。
11
第 2 章 実験
2.1
試料準備
現在の酒造りでは、協会酵母と呼ばれる各地の酒造場から優秀な酵母を分離 し、純粋に培養した酵母を使用している。協会酵母にも様々な種類が有るが、
最初の静置培養実験では呼吸機能が比較的弱く、発酵機能が強く、
10~
15℃の 低温でもよく発酵する協会酵母7号
(k-
7)と呼ばれる酵母を使用した。
福岡県工業技術センターにおいて、最初に、通常
1/mLで飽和する協
会酵母
7号を
1.25 mL採取し、米麹と水で作成した酵母などを培養するときに
培地として用いられるグルコース濃度
15%の麹汁
100 mLに入れることにより、
酵母濃度
1/mLを作り出した。これを
30℃で三日間寝かせ、お酒の密
度と同じになるようにした。この液体試料をピペットマンにより
625μ
L採取 し、エッペンチューブに
2回入れる事により、
1.25 mLの試料を用意し、これ を
14個作る事で
2パターンの一週間分の試料の用意とした。このピペットマン は
70%のエタノールによって消毒され、先端のチップ、及びエッペンチューブ は
121℃、高圧状態に設定したオートクレイブにより殺菌し、無菌状態としてい る。図
2.1の上部にピペットマンと下部にエッペンチューブを示す。
108
0 . 2 ×
106
5 . 2 ×
図
2.1.ピペットマンとエッペンチューブ
また、二回目以降の振とう培養実験では、日本酒酵母に変わって実験酵母を 使用した。この酵母を富栄養培地である
YPD(Yeast extract Peptone Dextrose)12
合成培地を用いて培養した。これはイーストエキス、ポリペプトン、グルコー スの栄養素をそれぞれ
1:2:2の割合で調合し、この溶液
5 mL中に実験酵母をお
およそ
1/mL程度の濃度になるよう加え、この試料を磁場印加用と対照
の二つをコニカルチューブに入れて用意した。
106
0 . 5 ×
2.2
磁場装置
福岡大学にあるジャパンマグネットテクノロジー社製の冷凍機冷却型
10 T超 伝導マグネットを用いて磁場を印加した。
磁場をかけた状態についての説明の前に、この装置について述べる。円柱形の 機器の中央部に円筒形の入れ物が有り、中には水が入っている。最初の実験で はこの水の水位を
28.5 cmに調節して、サンプルに均一な静磁場が発生するよ うにした。水はジェネレーターにより温度管理を行い、一定に保つように設定 した。この装置の磁場の状態を表したものを図
2.2に示す。この状態の装置で 磁場をかけた。
28.5 cm
磁場安定領域
図
2.2.冷凍機冷却型
10 T超伝導マグネットの磁場イメージ
同じように円筒形機器に水を張り、磁場のかかっていない定常状態における 対照試料として、同じく試料と装置の準備をした。静置培養実験では、これら の試料を一日おきに磁場中と地磁場中からそれぞれ一つ取り出し、
7日間繰り返 す事により、一週間分の実験試料を得た。磁場をかけた試料を
A、磁場をかけ なかった試料を
Bとし、経過日数に応じて、
A1、
A2…A6、
A7、
B1、
B2…B6、
13
B7
というように区別しておく。
また、振とう培養実験では水の水位をコニカルチューブの下部が浸かる程度 に調整し、温度一定に保つように設定した。この状態で振とう装置を作動させ、
規定経過時間毎に磁場中と地磁場中からそれぞれ
100 Lの試料を取り出した。
ここでも同様に、磁場をかけた試料を
a、磁場をかけなかった試料を
bとし、
経過時間に応じて、
a0、
a1…a9、
a10、
b0、
b1…b9、
b10というように区別し ておく。
μ
2.3
振とう装置
最初に行った静置培養実験では、後述するように培養環境の問題があった。
加えて、一週間という実験時間の長さも短所であった。これらを改善する為に、
培養方法を振とう培養に切り替えた。振とう培養する利点として、溶液中の液 体が攪拌され、十分に養分が行渡り、エアレーションも行う事ができる点であ る。静置培養で起こっていたエッペンチューブ底部での酵母や栄養分・老廃物 の蓄積が防げ、培養環境面の改善が期待できる。さらに振とう実験では温度を
20~
30℃と高く設定し、静置培養では一週間かかった培養時間を一日に早める 措置を取り、繰り返し実験を行い易くした。これらの培養環境改善により、同 条件下での実験を繰り返し行う事が可能となり、再現性の高い実験が行える。
次に、振とう装置自体について述べる。
本実験で使用する振とう装置は、空気の出し入れによってピストン運動を行 う、アルバック九州社製のエアーシリンダー型振とう装置である。ストローク 幅は
1 cmで、
0.4秒で一往復するように設定した。この先端にコニカルチュー ブを挿入するのだが、エッペンチューブから容器を変更した理由は、小さなエ ッペンチューブを7個用意した場合、それぞれで培養環境が違ってくる恐れが ある為に、大きな一つの容器にまとめる事とした。振とう装置を図
2.3に示す。
図
2.3の右図で示すような縦長い器具の先端にコニカルールチューブをはめ、空 気の出し入れによりピストン運動を行う。
14
図
2.3.振とう装置
2.4全菌数測定
次に、実験試料の測定方法を述べる。酵母数の測定にはその試料毎に適切な 希釈を行い、菌数計算盤上の酵母を顕微鏡により観察し、
1 mL中の数を割り 出す。まず、菌数計算盤について説明する。
この装置はカバーグラスとの間に一定の深さがあり、図
2.4のような方眼を 刻んだスライドグラスに菌浮遊液をとり、顕微鏡下で区画内の全菌数が計算で きる。この時、採取する容器であるエッペンチューブの底に酵母が固まってい る場合がある為、試料液内が均一になるように振って良く混ぜる。スライドグ ラスの方眼は、深さ
0.1 mm、方眼間隔
0.05 mmとなっている。つまり、
1区 画の菌数を
xとして、次のような計算により
1 mL中の全菌数 が分かる。
n15
図
2.4.菌数計算盤
まず、図
2.4で示した部分である
1区画の体積
Vを求める。
V =0.05mm×0.05mm×0.1mm
mL 10 5 . 2
cm 10 5 . 2
mm 10 5 . 2
7 3 7
3 4
−
−
−
×
=
×
=
×
=
よって、
1区画の菌数を
xとして、
1mL中の全菌数 は
n10 5 .
2 × −7
= x
n
6 10 0 . 1
0 . 4
× −
= x
=4.0x×106 1/mL
(2.1)
となり、以上より
1区画の菌数を測定すれば全菌数が求まる事が分かる。
また、振とう培養実験では一律に
10倍希釈ではなく、酵母菌数が
100前後に なるように試料各々に適した希釈を行い、精度の高い測定を行った。ここで、
後の議論として を磁場を印加して培養した試料の酵母数、 を対照として地 磁場で培養した試料の酵母数と定義しておく。
nM nc
16
2.5
生菌数測定
生菌数を測定するには、酵母がコロニーを形成したかどうかを見れば分かる。
コロニーを形成していれば、その酵母は生きていた事になるからである。図
2.5にこの様子を示す。
1cm
図
2.5.酵母のコロニー形成
この方法を平板培養法と呼び、予め底皿に薄く寒天培地を張り、そこに試料を 混ぜる。ここで、シャーレに
300前後の酵母菌が散布されるように希釈を行っ た。
希釈率を表
2.1に示す。シャーレを
4皿用意し、それぞれに
#1、
#2、
#3、
#4の記号を割り振った。 今回の生菌数測定では
#1、
#3にはシャーレに
50 L。
#2、
#4
にはシャーレに
100 Lの希釈した試料を、エタノールとガスバーナーによ る熱で殺菌したガラス棒により、シャーレ上に薄く延ばし散布した。
μ μ
表
2.1.生菌数測定の希釈率
A1、B1 A2、B2 A3、B3 A4、B4 A5、B5 A6、B6
#1、#2 3万倍 5万倍 7万倍 10万倍 10万倍 12万倍
#3、#4 10万倍
希釈のやり方についてだが、数万倍という希釈の為に二度に分けて希釈を行っ た。例を挙げると、
A1-#1の場合試料から
10 Lμを取り出し、水
990 Lμと合
17
わせて
100倍希釈の試料を作り出す。この
100倍希釈試料から
2 Lを取り出
し、水
600 Lと混ぜて
3万倍希釈の試料を得た。このように希釈した資料を
散布したシャーレ
48個を、
30℃一定で三日間培養を行い、コロニーの生育を促 した。この時シャーレを引っくり返して、雑菌が培地表面に落ちる事がないよ うに考慮した。
μ μ
18
第 3 章 結果と検討
3.1
静置培養
実験の記録を以下に述べる。
2006年
9月
14日
16時に磁場をかけ、
15日の
15時
30分に最初の試料を取り出した。続けて
16日には
15時
25分に、
17日 には
14時
5分に試料を取り出したが、
17日の
19時頃に台風による停電で、
18日の
16時
9分まで磁場のかかっていない状態が続いた。
19日、
20日、
21日は
16時に試料を取り出したが、停電による
21時間のロスにより、用意した試料 一週間分ではなく
6日分の計
12試料のみを得た。 実験は
10 Tの静磁場で行い、
酵母の培養温度を
15℃とした。
3.1.1
全菌数測定
次に、光学顕微鏡により菌数計算盤上
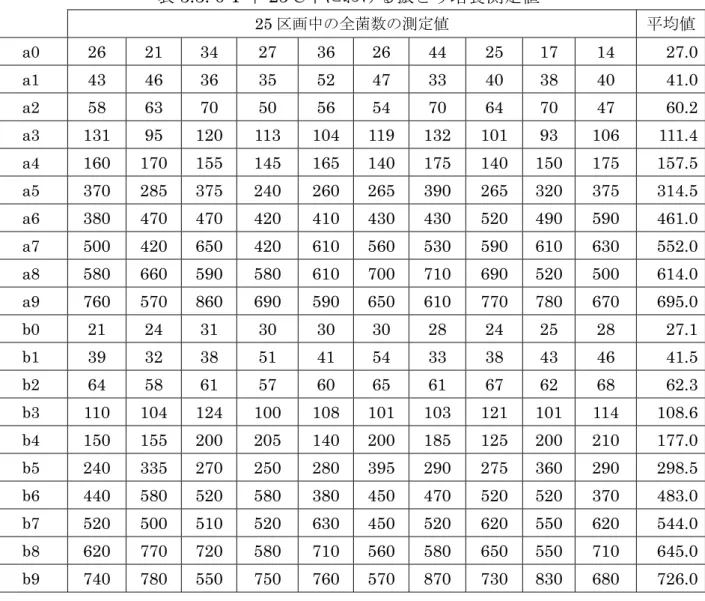
25区画中の全菌数を測定した結果を、
表
3.1に示す。 これらの値の平均値に、 式
(2.1)を用いて得た
1 mL中の全菌数と、
時間経過による変化をグラフで表したものを図
3.1に示す。ここで、近似式とし て
nM =6.616×105t+2.5×106および、
nc=9.456×105t+2.5×106を用いた。
19
表
3.1. 10 T中
15℃下における静置培養全菌数測定値
25区画中の全菌数の測定値 平均値
A1 14 12 15 17 13 15 10 14 18 14.2 A2 29 15 11 18 16 24 24 19 17 19.2 A3 13 18 17 12 18 5 4 11 12 12.2 A4 14 25 18 17 14 13 21 19 16 17.4 A5 21 15 26 16 17 9 15 16 21 17.3 A6 18 24 17 13 28 28 16 19 15 19.7 B1 7 13 7 5 6 4 11 10 12 8.3 B2 18 18 37 24 33 27 16 36 27 26.2 B3 22 15 21 13 16 20 19 13 16 17.2 B4 49 40 35 27 38 40 37 32 40 37.5 B5 33 41 28 49 37 30 22 35 30 33.8 B6 23 19 35 33 28 28 27 34 37 29.3
0 50 100 150
106 107 108
Yeast number n ,n (1/ml)
t(h)
MF(10 T) contrast
Mc
図
3.1. 10 T中
15℃下における酵母全菌数の時間依存性
図
3.1におけるグラフにより、時間経過と共に酵母が増えているのが分かる。
さらにこの増殖は勢いが衰え、ある値で収束している。これは酵母が試料の栄 養分を食い尽くし、増殖に歯止めがかかった為であり、生物学的観点から見て
20
妥当であり、この事を確認できた。ここで最も注目すべき点は磁場による影響 があるのかどうかとう事だが、グラフより磁場をかけた試料の方が増殖活動が 抑えられている事が読み取れる。
3.1.2
生菌数測定
次に、生菌数測定で得た値を表
3.2に示す。この値から明らかに
#3、
#4は希 釈の段階で失敗し、十分なデータが得られなかったので、後からの考察に考慮 しないものとする。残りの
#1、
#2を平均し、
1 mL中の生菌数と、時間経過に よ る 変 化 を グ ラ フ で 表 し た も の を 図
3.2に 示 す 。 こ こ で 、 近 似 式 と し て
および、 を用いた。
6 5
M =2.053×10 t+2.5×10
n nc =3.617×105t+2.5×106
表
3.2. 10 T中
15℃下における静置培養生菌数測定値
#1 #2 #3 #4 A1 25 32 7 20 A2 62 119 6 27 A3 71 132 12 25 A4 38 97 3 26 A5 39 88 2 14 A6 33 95 7 19 B1 20 26 5 20 B2 69 129 21 38 B3 74 158 20 39 B4 59 117 10 22 B5 48 90 80 154 B6 78 144 17 27
21
0 50 100 150 106
107 108
t(h)
Yeast number n ,n (C.F.U./ml)
MF(10 T) contrast
cM
図
3.2. 10 T中
15℃下における酵母生菌数の時間依存性
ここまでの実験を通して、全体的な実験手法として改善の余地があると考え た。まず、試料を磁場にかける際に
14個の試料の状態が水に浮いているもの、
沈んでいるものでバラバラだったという事が挙げられる。この状態の悪さを顕 著に示したのが、前述の全菌数測定の時で、試料によってはエッペンチューブ の底に酵母が固まっていたり、固まっていなかったりした為である。また、菌 数測定においても測定力の無さを痛感した。チップやエッペンチューブの蓋内 部は、雑菌を付けない為にも余計な物に触れさせてはいけないのだが、実験中 触れてしまった事が多々あった。同様に正確な測定について、シャーレに試料 を散布する際に他の雑菌が混入しないように蓋を少しだけ開け、すばやく処理 しないといけないのだが、明らかに目に見える他の菌が生育していたシャーレ が有った。また、図
3.1、
3.2から読み取れるように生菌数が全菌数をはるかに 上回るという本来ならば有り得ない実験結果となった。しかしながら、磁場を 印加した影響については、両者共に磁場を印加した試料では増殖が抑えられて いる事で一致する。ここで、全菌数と生菌数のそれぞれの最大値で規格化した グラフを図
3.3と図
3.4に示す。ここで、近似式として およ び、 を用いた。両グラフにおいて、磁場を印加した試料の規
530 . 0 2 M 6.592 10 t
n = × −
595 . 0 2 c 4.328 10 t
n = × −
22
格化した数値が、対照と比べて高い事が見てとれ、全菌数・生菌数共にこの傾 向がある事を示している。
0 100
10–2 10–1 100
Standardization Yeast number n ,n
t(h)
MF(10 T) contrast
Mc
図
3.3. 10 T中
15℃下における規格化酵母全菌数の時間依存性
0 100
10–2 10–1 100
t(h)
Standardization Yeast number n ,n
MF(10 T) contrast
Mc
図
3.4. 10 T中
15℃下における規格化酵母生菌数の時間依存性
23
3.2
振とう培養
これらの事から、正確な実験データを得る為の改善策として、磁場中に投入 する試料を同条件にする為に
7個に分けていた試料を一つにまとめ、振る事が 望ましいと考え、振とう実験へと移行する。
また、磁場を印加して培養する試料
aと、対照として地磁場中で培養する試 料
bの酵母の発育状態が、磁場を印加しない
0 Tにおいては差が無い事をまず 証明する。温度を
25℃一定で固定して、
0 T下で培養を行った最初の試料
a0、
b0は等しく、後の
a1、
b1は実験開始より
1時間後に採取し、
a2、
b2は
2時間 後、
a3、
b3は
4時間後、
a4、
b4は
6時間
15分後、
a5、
b5は
9時間後 、
a6、
b6は
12時間後、
a7、
b7は
16時間後、
a8、
b8は
21時間後、
a9、
b9は
26時 間後に採取した。これらの試料を光学顕微鏡により菌数計算盤上
25区画中の全 菌数を測定した結果を、表
3.3に示す。これらの値の平均値に、式
(2.1)を用い て得た
1 mL中の全菌数と、時間経過による変化をグラフで表したものを図
3.5に示し、 増殖期での分裂スピードを表すグラフを図
3.6に示す。 図
3.5では、
t =0での
nM、 の値をそれぞれ
nc nM0、
nc0と定義し、
γを定数とすれば、 お
よび、 の近似式を用いた。以下の全ての実験において、これらの近似 曲線を導入する。
tγ
n nM = M0
tγ
n nc = c0
24
表
3.3. 0 T中
25℃下における振とう培養測定値
25区画中の全菌数の測定値 平均値
a0 26 21 34 27 36 26 44 25 17 14 27.0 a1 43 46 36 35 52 47 33 40 38 40 41.0 a2 58 63 70 50 56 54 70 64 70 47 60.2 a3 131 95 120 113 104 119 132 101 93 106 111.4 a4 160 170 155 145 165 140 175 140 150 175 157.5 a5 370 285 375 240 260 265 390 265 320 375 314.5 a6 380 470 470 420 410 430 430 520 490 590 461.0 a7 500 420 650 420 610 560 530 590 610 630 552.0 a8 580 660 590 580 610 700 710 690 520 500 614.0 a9 760 570 860 690 590 650 610 770 780 670 695.0 b0 21 24 31 30 30 30 28 24 25 28 27.1 b1 39 32 38 51 41 54 33 38 43 46 41.5 b2 64 58 61 57 60 65 61 67 62 68 62.3 b3 110 104 124 100 108 101 103 121 101 114 108.6 b4 150 155 200 205 140 200 185 125 200 210 177.0 b5 240 335 270 250 280 395 290 275 360 290 298.5 b6 440 580 520 580 380 450 470 520 520 370 483.0 b7 520 500 510 520 630 450 520 620 550 620 544.0 b8 620 770 720 580 710 560 580 650 550 710 645.0 b9 740 780 550 750 760 570 870 730 830 680 726.0
25
0 10 20 30 107
108
t(h)
Yeast number n ,n (1/ml)
MF(0 T) contrast
Mc
図
3.5. 0 T中
25℃下における酵母全菌数の時間依存性
0 2 4 6
107 108
t(h)
Yeast number n ,n (1/ml)
MF(0 T) contrast
Mc
図
3.6. 0 T中
25℃下における増殖期比較
26
図から読み取れるように、試料
aと
bに違いは無く、図
3.6の試料
aと
bの 傾きの差が
0.002 1/mL・
hとほぼ無い事からも、磁場をかけるマグネット内で の
0 Tでの培養と、地磁場中での培養に違いは見られなかった。
3.2.1
温度による変化
10 T
中
30℃下で試料を培養する。まず、最初の試料
a0、
b0は等しく、後の
a1、
b1は実験開始より
1時間後に採取し、
a2、
b2は
2時間後、
a3、
b3は
4時 間後、
a4、
b4は
6時間後、
a5、
b5は
9時間後 、
a6、
b6は
12時間後、
a7、
b7は
24時間後、
a8、
b8は
27時間後に採取した。これらの試料を光学顕微鏡によ り菌数計算盤上
25区画中の全菌数を測定した結果を、表
3.4に示す。これらの 値の平均値に、式
(2.1)を用いて得た
1 mL中の全菌数と、時間経過による変化 をグラフで表したものを図
3.7に示し、増殖期での分裂スピードを表すグラフを 図
3.8に示す。
表
3.4. 10 T中
30℃下における振とう培養測定値
25区画中の全菌数の測定値 平均値 25区画中の全菌数の測定値 平均値 a0 47 58 47 44 49.0 b0 47 58 47 44 49.0 a1 104 61 91 81 84.3 b1 110 98 96 96 100.0 a2 189 168 152 174 170.8 b2 174 145 154 173 161.5 a3 51 49 30 50 450.0 b3 63 48 45 55 527.5 a4 90 91 92 123 990.0 b4 66 119 108 122 1037.5 a5 141 127 98 113 1197.5 b5 175 137 130 153 1487.5 a6 149 142 148 165 1510.0 b6 185 163 181 185 1785.0 a7 223 248 262 230 2407.5 b7 226 173 206 209 2035.0 a8 229 219 249 237 2335.0 b8 263 268 250 311 2167.5
27
0 10 20 30 107
108
Yeast number n ,n (1/ml)
MF(10 T) contrast
t(h)
Mc
図
3.7. 10 T中
30℃下における酵母全菌数の時間依存性
0 2 4 6
107 108
Yeast number n ,n (1/ml)
MF(10 T) contrast
t(h)
Mc
図
3.8. 10 T中
30℃下における増殖期比較
28
同様に
10 T中
25℃下で試料を培養する。まず、最初の試料
a0、
b0は等しく、
後の
a1、
b1は実験開始より
1時間後に採取し、
a2、
b2は
2時間後、
a3、
b3は
4時間後、
a4、
b4は
6時間後、
a5、
b5は
9時間後 、
a6、
b6は
12時間後、
a7、
b7は
15時間後、
a8、
b8は
21時間後に採取した。これらの試料を光学顕微鏡 により菌数計算盤上
25区画中の全菌数を測定した結果を、表
3.5に示す。これ らの値の平均値に、式
(2.1)を用いて得た
1 mL中の全菌数と、時間経過による 変化をグラフで表したものを図
3.9に示し、増殖期での分裂スピードを表すグラ フを図
3.10に示す。
表
3.5. 10 T中
25℃下における振とう培養測定値
25区画中の全菌数の測定値 平均値 25区画中の全菌数の測定値 平均値 a0 25 27 22 34 27.0 b0 35 22 33 22 28.0 a1 27 38 26 41 33.0 b1 37 36 37 35 36.3 a2 43 58 34 43 44.5 b2 45 51 52 58 51.5 a3 59 65 79 70 68.3 b3 61 93 137 71 90.5 a4 103 147 177 169 149.0 b4 217 198 173 163 187.8 a5 323 343 356 358 345.0 b5 418 337 396 402 388.3 a6 760 710 630 680 695.0 b6 890 610 106 830 609.0 a7 1200 1100 1280 760 1085.0 b7 920 960 840 930 912.5 a8 1450 1230 1230 1310 1305.0 b8 1330 990 1100 1290 1177.5
29
0 10 20 30 107
108
t(h)
Yeast number n ,n (1/ml)
MF(10 T) contrast
Mc
図
3.9. 10 T中
25℃下における酵母全菌数の時間依存性
0 2 4 6
107 108
t(h)
Yeast number n ,n (1/ml)
MF(10 T) contrast
Mc
図
3.10. 10 T中
25℃下における増殖期比較
30