In vivo バイオイメージングを用いた筋疲労要因の評価
稲垣 薫克
電気通信大学
2011 年 3 月
In vivo バイオイメージングを用いた筋疲労要因の評価
稲垣 薫克
電気通信大学大学院 電気通信学研究科 博士(理学)の学位申請論文
2011 年 3 月
In vivo バイオイメージングを用いた筋疲労要因の評価
博士論文審査委員会
主査 狩野 豊 准教授
委員 丹羽 治樹 教授
委員 中村 整 教授
委員 樫森 与志喜 教授
委員 白川 英樹 准教授
委員 長澤 純一 准教授
著作権所有者
稲垣 薫克
2011 年
Evaluation of the muscle fatigue factors using in vivo bio-imaging model.
Tadakatsu Inagaki Abstract
The present study attempted to elucidate skeletal muscle fatigue factors using in vivo bio-imaging model of rat skeletal muscle.
Skeletal muscle fatigue is characterized by a progressive decline of muscle force production or power output during and following prolonged or repeated muscle contractions. Although the mechanisms involved have been widely studied, detailed mechanisms are still unclear because of numerous factors influenced. Therefore, to clarify the mechanisms of muscle fatigue many studies have been performed on isolated whole muscles or single fibers. However, in vitro study has various limitations.
For instance, single fibers do not survive well at physiological temperature, there is no blood circulation and there are no interactions between different muscle fiber types, thus there are many differences compared with under in vivo conditions. Neural modulation, blood flow, muscular energy metabolism and the intracellular ion dynamics affect muscle fatigue. To study the mechanism of muscle fatigue, the complex in vivo observation model is required.
The rat spinotrapezius muscle is likened to the human quadriceps femoris with respect to containing each of the major muscle fiber types and able to observe under intravital microscopy. Recently, the bio-technology using a combined with a fluorescent substance for a specific ion has remarkable been developed. By combining these approaches, the purpose of present investigation was to test the original challenges in the rat spinotrapezius muscle during muscle fatigue, and we demonstrated following results.
To establish in vivo model of muscle fatigue, various muscle stimulations
were used. 1) Whereas repetitive isometric tetanic contractions induced
by 0.2-ms pulse stimulations did not affect arteriolar diameter or muscle
fatigability, 4-ms pulse stimulations evoked progressive arteriolar
vasoconstrictor response that is blocked by alpha adrenergic receptor
antagonists and intimately associated with fatigue process. 2) Repetitive
eccentric contractions maintained passive force production due to elevated
[Ca
2+]
ito a greater extent than isometric contractions. 3) Females
maintain force production during repetitive isometric tetanic contractions
due to [Ca
2+]
ihomeostasis was maintained, and the presence of normal
estrogen levels may not obligatory for the gender difference of muscle
fatigue. 4) Intracellular acidosis may not be a cause of muscle fatigue
because it was adjusted by monocarboxylic acid transporter that existed on
sarcolemma.
In vivo バイオイメージングを用いた筋疲労要因の評価
稲垣 薫克
概要
筋が一定の張力を継続して発揮できなくなる現象は筋疲労と呼ばれる.筋疲 労の末梢性要因には筋線維膜の興奮性や筋の収縮機構といった興奮収縮連関,
そして代謝エネルギー供給と蓄積した代謝産物の除去などの過程が阻害される ことがある.これまでの末梢性筋疲労に関する研究の多くは単離した動物の筋 束や単一筋線維標本での結果によるものが多い.しかしながら,これらの方法 は血流や細胞内外のイオン濃度勾配といった疲労に影響する因子が排除された ものである.骨格筋の疲労メカニズムの解明にはホメオスタシスが維持された 観察モデルの必要性が高い.そこで,本研究ではラット生体モデルに
in vivo
バ イオイメージング技法を適用し,末梢性疲労のメカニズムを検討するために以 下の研究課題を設定した.本論文は以下の
7
章で構成されている.第1
章では疲労のメカニズムに関す る文献研究をおこなった.第2
章ではこれまでの筋疲労研究モデルにおける問 題点を指摘し,その解決方策としてのラット生体モデルの有用性について記し た.第3
章では,研究課題1
として,バイオイメージングを活用できる筋疲労 モデルの確立のために,刺激パラメーター,収縮様式,性差について評価した.第
4
章では,研究課題2
として,筋疲労時の末梢循環動態の評価とそのメカニ ズムについて,第5
章では,研究課題3
として,筋疲労時の細胞内Ca
2+およびpH
動態に焦点を当て,疲労メカニズムの検討について記した.これらの3
つの研究課題について,その概要を以下に示す.
研究課題
1 (第 4
章):生体内環境下(in vivo)での筋疲労動態観察モデルの確 立を目的として,ラット脊柱僧帽筋における異なる収縮刺激負荷(0.2-ms vs. 4-mspulse duration)に対する発揮張力特性を検討した.その結果, 0.2-ms
刺激に対して,4-ms刺激での連続的な等尺性の強縮負荷時に顕著な筋疲労が誘発されるこ とが明らかとなった.また,伸張性収縮負荷では等尺性収縮とは異なり張力発 揮が維持されることが示され,これには伸張性収縮によって引き起こされる筋 損傷が関与することが考えられた.また,同様の刺激プロトコルにより,筋疲 労現象には性差が生じることが確かめられた.
研究課題
2 (第 5
章):課題1において観察された疲労の差が,血液循環の違い によるものかどうかを明らかにするために,連続的な筋収縮後の末梢血管動態 の評価をおこなった.その結果,4-ms刺激では筋収縮直後に細動脈の収縮が筋 収縮の頻度依存で生じ,細動脈の収縮は筋疲労が顕著になるにつれて増大する ことが明らかになった.一方,0.2-ms刺激では,細動脈の収縮応答は観察され なかった.次に,α-アドレナリン受容体を薬理学的に阻害し,交感神経系の筋疲
労への関与を検討した.その結果,連続的な筋収縮後に細動脈の血管収縮作用 が亢進する機序は,α1-ならびにα2-アドレナリン受容体を介したものであるこ
とが明らかとなった.血管緊張を決定する拡張作用と収縮作用との動的平衡は,運動の継続に伴って交感神経性の血管収縮作用が優位になり,その結果として,
運動後における血液供給量の低下を招き,筋疲労の回復が遅延することに関係 していることが考えられた.
研究課題
3 (第 6
章):筋収縮様式に関連した疲労要因としてCa
2+の関与を検討 するために,バイオイメージング法を用いて,筋細胞内のCa
2+動態を評価し,伸張性収縮では等尺性収縮と比較して顕著な
Ca
2+の蓄積が生じることが明らかとなった.さらに,疲労の性差における要因を検討し,雄と比較して雌では
Ca
2+の蓄積が生じないこと,およびエストロゲンはそれらの動態に対して関与しな いことが示された.また,筋疲労時の細胞内
pH
動態の評価とその制御機構につ いての検討をおこなった.その結果, 細胞内pH
はin vivo
条件では筋疲労時に おいても有意に低下しないことが示された.第
6
章および第7
章では各研究課題から得られた成果を検討し,本モデルの 問題点やその解決方策を述べ,末梢性疲労と循環動態および筋細胞代謝動態と の関連性について総括した.本研究の結論として,in vivoモデルの筋疲労研究によって,1)ホメオスタシ スが維持された状態で,微小循環および筋線維毎の評価が可能となった.2)筋 収縮の継続に伴って交感神経性の血管収縮作用が優位になり,その結果として,
回復期における血液供給量の低下を招き,筋疲労の回復が遅延することが明ら かとなった.3)連続的な等尺性収縮により筋細胞内
pH
は一定のままであった が,Ca2+の蓄積が生じた.また,筋細胞内のCa
2+蓄積はFemale
ラットでは起こ らず,伸張性収縮では顕著であった.末梢性疲労の発生メカニズムの解明のために,生体のホメオスタシスが維持 された状態で循環動態および筋細胞代謝動態を統合的に観察する必要性が明ら かになった.本研究におけるラット生体モデルを用いて得られた知見は,筋疲 労要因を多角的に捉えることにより,細胞機能を生体組織の統合化された状態 で明らかにすることを可能にし,今後の生体機能解明のための有用な研究技法 にも発展すると考えられる.
目次
第 1 章 文献研究 … 1
1 .緒言
2 .骨格筋の疲労
2-1 .筋線維の分類 … 3 2-2 .筋疲労の定義 … 3 2-3 .筋疲労の発生部位 … 5 3 .筋疲労と血流量の関係 … 11 4 .筋疲労の性差 … 13 5 .筋疲労研究モデル … 15
第 2 章 研究目的および研究課題 … 16
第 3 章 ラット in vivo モデルにおける張力発揮動態 … 19
1 .背景と目的
2 .方法
3 .結果
4 .考察
5 .要約
第 4 章 筋疲労時の微小血管動態の評価 … 33
1 .背景と目的
2 .方法
3 .結果
4 .考察
5 .要約
第 5 章 筋疲労時の骨格筋細胞内イオン動態の評価 … 55
( 1 )ラット in vivo モデルにおける Ca
2+動態の評価
A .異なる収縮様式負荷時の筋細胞内 Ca
2+動態 … 55 1 .背景と目的
2 .方法
3 .結果
4 .考察
5 .要約
B .筋疲労の性差と筋細胞内 Ca
2+動態 … 68
1 .背景と目的
2 .方法
3 .結果
4 .考察
5 .要約
( 2 )ラット in vivo モデルにおける pH 動態の評価 … 80
1 .背景と目的
2 .方法
3 .結果
4 .考察
5 .要約
第 6 章 討論 … 95
第 7 章 総括 … 102
謝辞 … 105
引用文献 … 106
関連論文 … 119
略号と記号
AD analog to digital, アナログデジタル
ADP adenosine diphosphate, アデノシン二リン酸 AP action potential, 活動電位
ATP adenosine triphosphate, アデノシン三リン酸 Ca2+ calcium ion, カルシウムイオン
CaPi calcium phosphate, リン酸カルシウム CO2 carbon dioxide, 二酸化炭素
⊿ delta, 変化量
ECC eccentric, 伸張性収縮
H+ proton, 水素イオン
HR heart rate, 心拍数 ISO isometric, 等尺性収縮
K+ potassium ion, カリウムイオン
KHB Krebs-Henseleit buffer, クレブス緩衝液 MAP mean arterial pressure, 平均動脈圧 Mg2+ magnesium ion, マグネシウムイオン
MCT monocarboxylate transporter, モノカルボン酸輸送体 NHE sodium ion / proton exchanger, Na+ / H+交換体 NMR nuclear magnetic resonance, 核磁気共鳴法 NO nitric oxide, 一酸化窒素
O2 oxygen, 酸素
OVX ovariectomized female rat, 卵巣摘出ラット Pi inorganic phosphoru, 無機リン
PO2 oxygen partial pressure, 酸素分圧 PCr phosphocreatine, クレアチンリン酸 pHi intracellular pH, 細胞内pH RM repeat maximum, 最大繰り返し数
SACs stretch-activated channels, 機械受容チャネル SNA sympathetic nerve activity, 交感神経活動 SR sarcoplasmic reticulm, 筋小胞体
TnC troponin C, トロポニンC TnI troponin I, トロポニンI τ time constant, 時定数
1A first order arteriole, 第一分岐細動脈
1
第 1 章 文献研究
1 .緒言
我々が日常的におこなっている歩行や走行,荷物を持つといった身体活動は全 て骨格筋の収縮が担っている.このような動作を継続的におこなった時にみら れるパフォーマンスの低下は筋疲労と呼ばれる.筋疲労は,我々にとって身近 な現象であり,その現象を客観的に評価するために,疲労に至るメカニズムの 解明や指標を決定するため,これまでさまざまな手法や視点から数多くの研究 がなされてきたが,一致した見解が得られていない.その原因として,骨格筋 が力を産生する過程は,神経活動や血流,筋のエネルギー代謝,そして,それ らに関わる様々なイオン動態により調節されており,疲労はこれらのどの過程 にも生じる可能性があるためである.
筋疲労に関わる物質として示唆されたものの中で有名なものには乳酸がある.
乳酸は比較的強い酸であるため,乳酸から放出される
H
+ がpH
の低下(アシド ーシス)を引き起こすことで筋機能の低下が起こると考えられていた(Fitts,1994)
.このことから,アシドーシスが筋の収縮過程に及ぼす影響について,多くの研究がなされ,アシドーシスが,筋小胞体(SR)からの
Ca
2+放出低下やト ロポニンC
のCa
2+感受性の低下,またはクロスブリッジの阻害を引き起こすこ とで張力低下を招くと考えられていた.しかしながら,近年の研究では,乳酸 が遅筋線維においてエネルギー基質となることが示され(Nielsen et al 2001),アシ ドーシスが筋疲労に直接的に関与しないことも支持されつつあるが(Allen et al2008; Stary & Hogan 2005; Westerblad et al 1997),一致した見解は得られていない
(Fitts 1994; Knuth et al 2006).これらの矛盾は,実験に用いた標本の線維構成の違
2
いや刺激方法の違いにより生じているのかもしれない.これまでの末梢性筋疲 労に関する研究の多くは単離した動物の筋束や単一筋線維標本での結果による ものである.しかしながら,これらの方法は血流や細胞内外のイオン濃度勾配 といった疲労に影響する因子が排除されたものであるため,得られる情報も限 定的なものになってしまう.疲労メカニズムの解明には,実験対象として生体 恒常性の維持された観察モデルを用いる必要があると考えられる.そこで本研 究では,筋疲労のメカニズムを生理学的,生化学的および形態学的に明らかに することを目的として,ラット生体モデルにバイオイメージング技法を適用し,
筋疲労時の生体情報の可視化を試みた.
3
2 .骨格筋の疲労
2-1 .筋線維の分類
筋収縮には,筋中に存在する
ATP
がそのエネルギー源として直接的に用いら れる.最大収縮を行った場合,細胞内の貯蔵ATP
はわずか1‐2
秒間で消費され てしまうため,収縮を持続するためにはATP
を補充しなければならない.ATP の供給系としてクレアチンリン酸(PCr)の分解(ATP-PCr系),嫌気的解糖(解 糖系),酸化的リン酸化(酸化系)がある.筋線維は一般的に使用するエネルギ ー供給系の違いによって3
つのタイプに分類される(Pette & Staron 1990).ミトコ ンドリア酵素染色またはadenosinetriphosphate:アデノシン三リン酸)加水分解
酵素(ミオシンATPase)活性を用いた組織化学的手法によって,筋線維は SO
線維(Slow Oxidative Fiber),FOG線維(Fast Oxidative-Glycolytic Fiber),FG線 維(Fast Glycolytic Fiber)に分類される (Barnard et al 1971; Peter et al 1972).また,ATPase
活性のみに基づいて,遅筋線維(TypeⅠ線維),速筋線維はさらに,異なる
pH
で処理すると異なる反応性を示すことから,低いpH
の範囲で組織化学 的に同定されたミオシンATPase
の消失に対する抵抗性にもとづいてTypeⅡA,
TypeⅡB
(ⅡX)およびTypeⅡC
に分けられる(Fig.1-1)(Berchtold et al 2000).筋線維タイプの違いはミオシン重鎖のタイプ構成に特有であるだけではなく,
生理学的および生化学的な特性によっても異なる.したがって,筋疲労の研究 には筋の線維構成を明らかにすることが重要である.
2-2 .筋疲労の定義
骨格筋における疲労,すなわち筋疲労は,最大筋力の低下や最大短縮速度の
4
Fig.1-1. Serial cross sections of human deltoid muscle stained for myofibrillar ATPase (mATPase) after preincubation at pH 10.5 or 4.3, cytochrome c oxidase (Cyt C ox), and α -glycerophosphatase dehydrogenase ( α -glyc).
The fiber types are indicated; for type I, IIA, IIB(IIX) and IIC fibers. They
show a largely reciprocal staining pattern after acid and alkaline
preincubation, respectively, as well as in the reaction for cytochrome c
oxidase and α-glycerophosphatase dehydrogenase, respectively
(Berchtold
et al. 2000).
5
低下と定義される(Bigland-Ritchie 1984; Sogaard et al 2006).筋疲労の程度には,
運動の強度やその頻度,持続時間,そして,運動間の休息時間といった多くの 条件が関与している(Enoka & Duchateau 2008).Fig.1-2は,単離したマウスの短 趾屈筋に対し,
70Hz
で4
秒に1
回の電気刺激を2
分間連続的に負荷したときの 張力変化を示している(Lannergren & Westerblad 1991).このように,最大筋力に おける低下は,連続的な強縮刺激中に最大等尺性張力を測定したときに明らか であり,一連の強縮の2
回目においてもみられるような進行的な低下が観察さ る.また,疲労動態は収縮頻度の増加によって異なり,筋線維のエネルギー供 給系の違いによっても異なる.さらに,筋線維の疲労動態は,興奮収縮連関の 過程における疲労発生部位によって3
つの相に分けられるが,詳細は次の節で 述べる.2-3 .筋疲労の発生部位
随意収縮において,筋は,大脳皮質から始まり脊髄における下位運動ニューロ ンの興奮をもたらす複雑な経路によって活性化される.下位運動ニューロンの 軸索は活動電位を筋の神経筋接合部まで伝える.脊髄内部と上部の過程は中枢 性と定義され,末梢神経,神経筋接合部および筋における過程は末梢性として 定義される.高強度での運動を継続すると最大発揮張力をはじめとした筋収縮 性が低下する.現象として,発揮張力の低下や短縮速度の低下,筋弛緩時間の 延長が観察されるようになる.筋疲労は中枢神経系の障害や骨格筋内の末梢性 因子の障害によって生じる.すなわち筋の収縮過程のすべてが疲労発生部位の 候補となるのである.Fig.1-3 に興奮収縮連関で筋疲労が生じる可能性のある部 位について示した.さらに複雑なことに,筋疲労に関与する物質も
pH
やCa
2+, 無機リン(Pi),活性酸素など複数あり,それらが作用する部位も上記のように6
Fig.1-2. Force records during fatigue produced by repeated short tetani in an isolated mouse single fibre under aerobic (A) and anaerobic (B) conditions.
Triangles (V) indicate times when the tetanus interval was shortened. In the
top panel, the phases of fatigue have indicated. Both runs obtained from the
same fibre, in A under aerobic and in B under anaerobic conditions (N
2+
CN poisoning) (Lannergren & Westerblad 1991).
7
Fig.1-3. Diagram of major components of muscle fiber involved in excitation-contraction coupling.
Numbers indicate possible sites of muscular fatigue and include the following: 1) surface menbrene, 2) T tublar charge movement with sarcoplasmic reticulm (SR) Ca
2+release, 4) SR Ca
2+release, 5) SR Ca
2+reuptake, 6) Ca
2+binding to troponin, and 7) myosin binding to actin, ATP
hydrolysis, and cross-bridge force development and cycle rate (Fitts, 1994).
8
複数存在し,作用する物質も
1
つではないため,非常に複雑になっている.こ こでは筋の収縮機構にそって,疲労発生部位について述べる.運動神経からのインパルスによって筋細胞膜に発生した活動電位は,膜表面を 伝播して横行小管を伝導し筋細胞の深部へと伝わる(Fig.1-3 ①‐③).筋小胞体
(SR)からの
Ca
2+は横行小管で活動電位が適切に生じなければならない.高強 度の運動によって筋疲労が生じている場合は,細胞外のK
+濃度増加が生じるた め(Clausen 2003).In vitroの単離筋において,5分から10
分の繰り返しの収縮に よって細胞内のK
+濃度([K+]
i)が30%低下し,静止膜電位が 10~18 mV
ほど脱 分極する(Balog & Fitts 1996).繰り返しの収縮によって細胞内のATP
濃度が低下 することにより,ATP感受性のK
+チャネルが開口してK
+の漏出が生じ,横 行小管の興奮性を低下させる.筋収縮による筋細胞内からのK
+流出量は遅筋線 維に比べ速筋線維で多いことが報告されている(Cairns et al 1997).したがって,持続的な筋収縮が筋力の低下を引き起こす要因として,活動筋から放出される
K
+が細胞膜の興奮性の低下を惹起し,その結果としてSR
へのシグナル伝達を抑 制してSR
のCa
2+放出を抑制するためだと考えられる.横行小管と
SR
の接合部では(Fig.1-3 ③‐④),横行小管膜に存在するジヒドロ ピリジン(DHP)受容体が横行小管の膜電位変化をSR
に伝える.DHP 受容体 からの情報伝達がMg
2+のリアノジン受容体への親和性低下というかたちで生じ,Mg
2+がリアノジン受容体から遊離することによってCa
2+が放出されると考えら れている.Fryerら(Fryer et al 1995)は,単一筋線維を50 mM
のPi
に曝露するとSR
のCa
2+放出低下が生じることを報告し,それが筋形質のPi
濃度上昇により一 部がSR
へと流入することによってCaPi
を形成することによると推測した.後 に,Allen
ら(Allen & Westerblad 2001)によってPi
濃度が6 mM
以上になるとCaPi
が形成されることが考えられている.これらは,クレアチンキナーゼ欠損マウ9
スの筋では
SR
のCa
2+放出低下が生じないことにより確認されている(Dahlstedt& Westerblad 2001).したがって, SR
のCa
2+放出低下は高強度運動によって生じ たPi
の蓄積が起因すると考えられている.SR
から放出されたCa
2+はアクチンフィラメントのトロポニンC(TnC)に結合
してミオシンフィラメントとの相互作用により張力が発生する(Fig.1-3 ⑤‐⑦). トロポニンはT, I, C
の3
成分で構成され,通常TnI
の一部がミオシン頭部とア クチンとの間に入り込むように位置し,クロスブリッジ形成を抑制するように 機能する.Ca2+がTnC
に結合すると抑制部位の位置変化によりTnI
の抑制作用 が失われクロスブリッジ形成が可能になる.pH
が低下すると,TnC
のCa
2+ 感受 性の低下が生じる(Ball et al 1994).筋原線維が収縮を起こすために必要なエネル ギーはミオシンATPase
の触媒作用により得られるが,その活性はTnC
に結合す るCa
2+量に依存するため,Ca
2+の不足により低下する.筋の弛緩で,筋形質内のCa
2+濃度が低下しTnC
からCa
2+が解離すると,アクチンからミオシン頭部が離 れ,筋原線維は弛緩する.筋形質内のCa
2+濃度の低下はSR Ca
2+-ATPase
により1mol
のATP
により2mol
のCa
2+が取り込まれることから,この酵素活性が弛緩 速度を規定している.Dukeらは単一筋線維を用いてPi
濃度上昇によりSR
からCa
2+の漏出することを示した.筋収縮時の安静期間中にみられるCa
2+濃度の増加 はPi
が原因と考えられている(Duke & Steele 2000).以上のことから,Ca2+や
H
+,Mg2+などといった多くのイオンや,ATP やADP,
グリコーゲンなどのエネルギー基質の作用により興奮収縮連関のそれぞれの過 程が阻害され(Fig. 1-4),これらの影響が
Fig.1-2
での筋疲労動態の変化をもた らしている.また,これらの代謝産物の変化は,運動中の複合的なイオン交換 によって影響を受ける.したがって,筋疲労時の筋線維レベルでのこれらのイ オン濃度の変化を捉えることは,筋疲労メカニズムの解明に重要である.10
Fig. 1-4. Schematic diagram illustrating the major mechanisms that contribute to muscle fatigue.
Heading in each box identifies subcellular function, and the subsequent list
indicates cellular changes occurring during fatigue that influence the
subcellular function. SM, surface membrane; TT, t tubule; SR,
sarcoplasmic reticulum; AP, action potential (Allen et al. 2008).
11
3 .筋疲労と血流量の関係
筋への血液循環の低下は,筋内の酸素供給不足を引き起こし,筋疲労を増大す る.また,異なる運動強度での筋収縮中における運動単位の動員の機序や,筋 線維組成の違いなどに起因する筋の酸化的能力によって,血液供給の低下が筋 持久能力に及ぼす影響に差が生じることも知られている.これまでに,このよ うな影響については,閉塞性動脈硬化症などの病態や阻血下での運動中の血流 動態の差異について研究されてきた.閉塞性動脈硬化症の患者では,活動筋へ の十分な血流量の確保が困難であるため,軽度の歩行運動をおこなった場合で も, 疲労感や痛みを感じることが報告されている (Safar et al 1987).また,外 部圧迫による血流制限を施すことにより,健常者においても閉塞性動脈硬化症 と同様に著しい筋力の低下や運動回数の減少が報告されている.例えば,Wern
bom et al. (Wernbom et al 2006)は,最大挙上重量(1RM)の 20~50%での膝伸展運
動によって,運動強度の高い50%強度では収縮期血圧の 130%の外部圧迫による
影響を受けないが,低強度になるにつれて反復回数が低下することを報告して いる.また,運動時の動脈血圧上昇が活動筋の灌流圧を上げ,筋力発揮に有利にはた らくことが,筋疲労時において示されている (Wright et al 2000).一方,平均血 圧 を 低 下 さ せ た 場 合 で は , 筋 力 発 揮 の 低 下 が 生 じ る こ と も 示 さ れ て い る
(Fitzpatrick et al 1996).平均血圧が筋力発揮を変動させる詳細な機構は明らかで
はないが,神経刺激によって誘発される活動電位波形が血圧変動により影響さ れないことから(Fitzpatrick et al 1996; Wright et al 2000),筋内には血圧に反応して 張力を変動するという機構が存在すると考えられている.一方,活動筋において,骨格筋血流は運動中に代謝需要に応じて増加する
12
(Andersen & Saltin 1985).運動中や運動後での骨格筋微小循環における血管緊張
は,交感神経性血管収縮作用と局所で放出される血管拡張性物質との動的平衡 により決定されている(Thomas & Segal 2004).運動強度の増加とともに骨格筋血 流は増大するが,筋交感神経活動もまた運動強度にともない増加する(DiCarlo etal. 1996, O'Leary et al. 1997).交感神経活動の増加は非活動部位における血管収縮
を生じ運動中の血流再分布において重要な役割を果たしている.交感神経活動 の効果器であるα-アドレナリン受容体は動脈にそって階層的に分布し,α1
およ びα2
受容体は筋収縮の血管調節に関与している.Buckwalterら(2001)は運動 がα1
およびα2
媒介の血管収縮を減弱化することを明らかにし,その応答は運 動強度により各受容体で異なり,α1 媒介の血管収縮は激運動時でのみ減弱する が,α2
媒介の血管収縮は全ての運動強度で減弱することを示している.さらに,α2
媒介の血管収縮はα1
媒介のものと比べて,PO
2(70から10 mmHg)や pH
(7.4 から7.0)の低下(McGillivray-Anderson and Faber, 1990, Tateishi and Faber, 1995),
K
ATPチャネルの活性化を通じたNO(Thomas et al. 1997)などの代謝的な変化に
影響を受けやすいことが明らかになっている.特に,小細動脈に分布するα2- アドレナリン受容体媒介の血管収縮作用が収縮筋で産生される物質による代謝 性阻害を受けやすいことが起因すると考えられる.したがって,筋疲労メカニ ズムの解明には筋疲労時の活動筋における血流動態,また,それを制御する末 梢の細動脈の血管動態を明らかにすることも重要となる.13
4 .筋疲労の性差
興味深いことに,ヒトや動物を用いた研究によって筋疲労には性差が存在する ことが示されている.前述のように,筋疲労は筋の最大張力発揮能力の低下と して表される.疲労耐性の能力は筋持久力と表現され,標的とする張力を維持 できなくなるまでの時間として表される.男性と比べて女性では疲労耐性があ るといった,筋疲労の性差に関する多くの報告がある.この筋疲労の性差は,
運動強度の増加とともに観察されなくなる傾向がある(Hicks & McCartney
1996).例として,膝伸展運動における 1
回の最大運動(1RM)の80%の強度
や(Laforest et al 1990; Maughan et al 1986),母指内転筋(adductor pollicis
muscle)における間欠的な最大収縮時において性差が観察されなくなる(Ditor
& Hicks 2000).また,最大筋力の一致した男性と女性では性差が観察されなく
なることも報告されていることから (Hunter et al 2004),絶対筋力の差も性差 の要因の一つであると考えられる.また,絶対筋力の差は筋収縮中の血液潅流 に影響する.筋収縮中に血管は筋内圧の上昇により圧迫され,血流が停止する.したがって,女性と比べて男性では筋収縮中の血液循環が低下し,酸素運搬や 代謝産物の除去などが障害され,結果として早い筋疲労が生じると考えられる.
一方,女性ホルモンであるエストロゲンにおいても,運動中,特に長時間運動 中のエネルギー代謝に影響する可能性があることから,筋疲労の性差の要因の 一つとして考えられている.エストロゲンが女性において,長時間運動中の糖 代謝が男性と比べて利用率が低いことが報告されており(Tarnopolsky 2000),こ れは動物を用いた実験でも確認され,卵巣摘出した雌ラットに対しエストラジ オールを投与すると,トレッドミル走行後のグリコーゲン枯渇が低下し,疲労 困憊に至るまでの時間も延長することが報告されている(Kendrick et al 1987).
14
しかしながら,グリコーゲン枯渇がみられない短期間の運動においても性差が みとめられることや(Kendrick et al 1987),閉経後の女性においても男性と比べ て疲労耐性があるといった報告(Hicks & McCartney 1996)から,筋疲労の性差 におけるエストロゲンの関与には議論の余地がある.
15
5 .筋疲労研究モデル
疲労研究における最高の判断基準は,中枢制御下での灌流されたインタクト な筋を用いること,つまり生体機能が統合化された状態のままで観察すること が理想である.これまでの末梢性筋疲労に関する研究の多くは単離した動物の 筋束や単一筋線維標本での結果によるものであるが,これらの方法では血流や 細胞内外のイオン濃度勾配といった疲労に影響する因子が排除されている.ま た,単離した筋は生理的温度である
37℃付近において生存できないため,20℃
程度の低温下で用いられる.Westerblad ら(Westerblad et al 1997)は
pH
低下 に伴う発揮張力の低下は温度依存的であり,筋疲労は生理的温度ではアシドー シスの影響を受けにくいことを示している.これらのことは,これまでに蓄積された
in vitro
での知見がin vivo
では必ずしも生じない可能性を示唆するものである.さらに,近年の研究では疲労物質とされていた速筋線維から排出され た乳酸が遅筋線維においてエネルギー基質となることが示されており,筋線維 間の相互作用も加味する必要がある.
これまでの
in vivo
条件での疲労研究は,筋電図や磁気共鳴分光法(NMR)を 用いたものがほとんどであったが,これらの方法では筋線維毎の様々なイオン や代謝産物などの動態を評価することはできない.さらに,意識下では,中枢 性の疲労要素も考慮しなければならない.最近では,特定のイオンと結合する ことで蛍光を発する物質や蛍光タンパク質の開発が盛んにおこなわれてきてお り,細胞内のイオン動態を直接的に観察するバイオイメージング技法が確立さ れてきた.この方法をホメオスタシスが維持されたモデルに適用することによ って,複雑な筋疲労メカニズムの解明に繋がることが考えられる.16
第 2 章 研究目的および研究課題
高強度での運動を継続すると最大発揮張力をはじめとした筋収縮性が低下す る.筋が一定の張力を継続して発揮できなくなる現象は筋疲労と呼ばれる.通 常の運動における筋疲労では末梢性要因である筋線維膜の興奮性や興奮収縮連 関,筋の収縮機構,そして代謝エネルギー供給と蓄積した代謝産物の除去など の過程が阻害されることが要因となることが示唆されているが,一致した見解 は得られていない.この要因として,生体恒常性が維持されたモデルを用いて いないことが考えられる.近年,特定のイオンと結合することで蛍光を発する 物質や蛍光タンパク質の開発が盛んにおこなわれてきており,細胞内のイオン 動態を直接的に観察するバイオイメージング技法が確立されてきた.この方法 をホメオスタシスが維持されたモデルに適用することによって,非常に複雑な 筋疲労メカニズムの解明に繋がることが考えられる.そこで,本研究では,末 梢性疲労のメカニズムを生理学的,生化学的および形態学的に明らかにするこ とを目的として,ラット生体モデルにバイオイメージング技法を適用した筋疲 労動態の可視化を試みるため,以下の研究課題を設定した.
研究課題 1 :ラット in vivo モデルにおける張力発揮動態
in vivo
での筋疲労動態観察モデルの確立を目的として,ラット脊柱僧帽筋における異なる収縮刺激負荷に対する発揮張力特性を検討する(実験
1)
.本モデル における張力発揮動態を,実験1-1
では刺激パルス幅の違いがについて,実験1-2
では等尺性(ISO)収縮と伸張性(ECC)収縮といった収縮様式の違いにつ いて検討し,実験1-3
では筋疲労に性差が生じるのかについて検討する.17
研究課題 2 :筋疲労時の微小血管動態の評価
筋疲労時の骨格筋における微小循環動態と筋疲労との関連性を検討するために,
連続的な筋収縮後の末梢血管動態の評価をおこなう.実験
1-1
において観察さ れた疲労の差が,血液循環の違いによるものかどうかを明らかにするために,同様の異なるパルス幅を用いた連続的な筋収縮後の末梢血管の内径動態を評価 する(実験
2)
.次に,α-アドレナリン受容体を薬理学的に阻害し,筋疲労と交 感神経系の関係性を検討する(実験3).
研究課題 3 :筋疲労時の筋細胞代謝動態の評価
ラット生体モデルに蛍光指示薬を用いたバイオイメージング法を適用し,筋疲 労時の筋細胞代謝動態の評価を,細胞内の
Ca
2+およびpH
動態に焦点を当てて検 討する.( 1 )ラット in vivo モデルにおける Ca
2+動態の評価
実験
2
の結果を受け,Ca
2+感受性蛍光色素を用いることによって,ISO
収縮およ びECC
収縮負荷による筋細胞内のCa
2+動態を評価する(実験4)
.実験1-3
で 観察された連続的なISO
収縮負荷時によるFemale
における筋疲労耐性には,エ ストロゲンや筋線維タイプ構成,または細胞内のCa
2+恒常性の違いにより生じ た可能性を検討するために,エストロゲン枯渇モデルとして卵巣摘出(OVX)の
Female
ラットを用いることによって,Male,FemaleおよびOVX female
との 比較をおこなう(実験5)
.( 2 )ラット in vivo モデルにおける pH 動態の評価
本モデルにおける筋細胞内の
pH(pHi)の変化を検討するために,pH
感受性の18
蛍光指示薬を用いたバイオイメージング法を適用することによって,1)ラット 生体モデルにより筋細胞内
pH
におけるin vivo
キャリブレーションにより蛍光強 度の変化を確認する,2)連続的な筋収縮後の筋疲労時の細胞内pH
動態を明ら かにする,3)本モデルにおけるpH
制御機構の役割を検討するために,筋細胞 膜に存在するNa
+/H
+交換体と乳酸‐H+共輸送体に着目し,薬理学的な阻害下に おけるpHi
動態の評価をおこなう.19
第 3 章 ラット in vivo モデルにおける張力発揮動態
(研究課題 1 )
1 .背景と目的
近年の研究により,これまで疲労物質とされていた乳酸は遅筋線維においてエ ネルギー基質となること(Bergersen et al 2006),張力低下の引き金となると考えら れていたアシドーシスは単離筋において温度依存で張力に及ぼす影響が異なる ことが報告されている(Ranatunga 1987).これらの新しい知見は,筋疲労メカニ ズムについて再考する機会をもたらした.これまでの筋疲労メカニズムに関す る研究の多くは,単離した動物の筋束や単一筋線維標本での結果によるもので ある.In vitro条件下の単離した筋は生理的温度である
37℃付近において生存で
きないため,20℃程度の低温下で用いなければならない.したがって,これら の実験対象から得られた生体情報が実際にin vivo
で生じているものなのかにつ いては議論の余地がある.また,筋疲労は神経性調節や血液循環,そして筋収 縮に関与する様々なイオン動態の影響を受ける.それらは単一に変動するので はなく,相互作用しながら同時発生的に生じる(Cairns & Lindinger 2008).また速 筋‐遅筋線維間のエネルギー基質のやり取りが存在する.したがって,筋疲労 メカニズムの解明のためには,生体環境が統合された実験系を用いる必要があ る.この解決方策として,ラットの背中部に位置する脊柱僧帽筋を対象にした 生体顕微鏡観察法(Gray 1973)を用いることが考えられる.この筋はヒトの大腿四 頭筋と類似した筋線維組成であり(TypeⅠ: 40%, TypeⅡA : 7%, TypeⅡD/X : 17%,TypeⅡB : 35% , Delp & Duan 1996)
,最薄部位で筋線維数本程度の厚さしかもた ず,環状の血管ネットワークが筋の剥離後も保持されるという特徴を持つため,20
筋微小循環の研究に用いられてきた(Gray 1973).本研究の目標は,この筋を対象 とした筋疲労モデルを確立することである.
身体活動は筋の発揮する力が起因となるが,筋力は筋の収縮様式によって大き く変化する.筋の収縮様式は静的収縮と動的収縮に分類され,静的収縮は筋長 が維持された状態で張力が発揮される等尺性(アイソメトリック;ISO)収縮で あり,動的収縮は筋が短縮しながら張力が発揮される短縮性(コンセントリッ ク;CON)収縮と伸張されながら張力を発揮する伸張性(エキセントリック:
ECC)に分類される. ISO
収縮は長さ変化を伴わないため,エネルギーは仕事ではなく熱として発揮される.また,ISO収縮は持続的な筋収縮運動であるため,
高強度の
ISO
運動では筋収縮に伴う機械的圧迫により活動筋血流が阻害される.Ichinose
ら(Ichinose et al 2006)はヒトの静的なハンドグリップ運動(ISO収縮)時 において,最大随意収縮(MVC)の30%で筋交感神経活動の増加および血圧の
上昇を示している.一般的に,随意運動時では交感神経活動の増加が生じ,そ の増加は動脈血管の収縮を生じる.活動筋では,交感神経活動が抑制され,血 流を増加させると考えられている.一方,ECC 収縮は他の収縮様式と比べて酸 素摂取量やATP
消費などの代謝コストが低く,筋放電量が小さいにもかかわら ず発揮張力が大きいという特性を持っている(Seger & Thorstensson 2000; Smith1991).したがって,筋疲労は ECC
収縮と比べるとISO
収縮において顕著になることが予想される.また先行研究から,筋疲労には性差が生じることが報告さ れているが(Hunter et al 2004; Hunter & Enoka 2001; Wust et al 2008),その要因は明 らかではない.そこで本研究では,in vivo での筋疲労動態観察モデルの確立の ため,表面電極による筋収縮負荷での刺激条件について,刺激パルス幅,収縮 様式,筋疲労の性差に着目して,これらの異なる刺激負荷による筋の発揮張力 動態の比較および検討をおこなった.
21
2 .方法
1.被験動物
本実験では,9-16週齢の
Wistar
系ラット(日本SLC)を用いた.全てのラッ
トは,室温23±2
℃,湿度55±10%で 12
時間の明暗サイクルに管理された飼育室 において餌と水を自由摂取できる状態で飼育した.全ての実験は電気通信大学 動物実験委員会の承認を得たものであり,本学動物実験指針に沿って行なわれ た.2.外科的処置
本実験では,ラット脊柱僧帽筋を対象とした.脊柱僧帽筋は薄い膜状筋であり,
神経活動や血流が維持された状態での顕微鏡観察が可能である(Gray 1973;
Sonobe et al 2008).外科手術のために,ラットにペントバルビタールナトリウム
(共立製薬,70mg/kg/Body weight)を腹腔内注入(i.p.)して麻酔をした.麻酔 は必要に応じ適宜追加した.処置中のラットは,体温を保持するため,37℃に 維持したホットプレート(BWT-100,バイオリサーチセンター)上に静置した.
すべての外科手術は実体顕微鏡下でおこなった.
麻酔下において,ラットの正中線から
3 cm
程度切開し,右背柱僧帽筋の組織を 傷つけないよう慎重に組織から剥離し,表面を覆う結合組織を取り除いた.脊 柱僧帽筋に特徴的なアーチ状の細動静脈システムを維持するため,筋の近位部 において主となる供給動脈を傷つけないよう注意しながら筋の一部分を剥離し た.剥離した筋は馬蹄型の針金のリングに筋長を維持するように縫合糸(Sigma)で固定された.筋収縮刺激負荷のために,筋の両端に電極を結びつけて電気刺 激装置(3F46,NEC 三栄)に接続し,収縮中の筋の発揮張力を測定するため筋
22
の端を運動負荷装置(RU-72 形,NEC メディカルシステムズ)にワイアーで結 びつけた.
外科手術の間,対象筋表面の乾燥を防ぐため,95% N2
-5% CO
2混合ガスによりpH 7.4
に平衡化し,37℃に維持された恒温槽に入れて保温したバッファー(Krebs-Henseleit Buffer: KHB ,132 mM; NaCl(Wako), 4.7mM; KCl(Wako),
21.8mM; NaHCO
(関東化学)3, 2mM; MgSO
4(Wako), 2mM;CaCl
2*2 H
2O
(Wako)) により常に保湿した.外科手術終了後,ラットを
37℃に維持したガラスホットプレート(Kitazato Supply)に固定し,脊柱僧帽筋部分は,呼吸や拍動以外の要因による観察部位の
ズレを防ぐためにガラスプレート上で固定した(Fig.3-1).3.実験手順
実験は,以下の
3
つの実験条件下にておこなった.・実験
1-1.電気刺激パラメーターが張力発揮に与える影響
上記の電気刺激装置により,実験開始前に電気刺激負荷の状態と筋の収縮状態 を確認した後,6-8 V,100Hz,刺激持続時間
0.7 sec
の等尺性(アイソメトッリ ク:ISO)強縮刺激を3
秒に1
回の頻度で負荷し,50
回(2.5 min)を1
セットと してこれを5
分間の安静期間をおいて10
セット行なった.電気刺激のパルス幅 の違いが張力発揮に与える影響を検討するために,上述の電気刺激をⅰ)4-ms の矩形波(N = 4),ⅱ)0.2-msの矩形波(N = 5)の2
条件で負荷した.・実験
1-2.様式の異なる筋収縮負荷が張力発揮に与える影響
本実験では,異なる収縮様式が張力発揮に及ぼす影響を検討するために,ⅰ)
23
ISO
収縮(N = 3),および,ⅱ)伸張性収縮(エキセントリック収縮:ECC,N = 4)
負荷時における張力発揮の比較をおこなった.ISO
収縮負荷条件は上述の4-ms
条件と同様におこなった.筋が伸張されながら収縮するECC
収縮を負荷するた め,脊柱僧帽筋遠位部に繋いだワイアーの一端を電気刺激装置と連動したひず み計を有する運動負荷装置に固定した.筋の伸張は安静時のサルコメア長に対して
10%程度の伸張(2.7µm
からおよそ3.0µm
まで)を伴うように回転角度を30 degree,角速度を 100 degree / sec
に設定し,電気刺激開始から0.2 sec
後に伸 張刺激を負荷した.・実験条件
1-3.性差が張力発揮に与える影響
本実験では,ⅰ)16週齢の雌ラット(N = 4),および雌ラットとの体重差を考 慮して,ⅱ)10-12週齢の雄ラット(N = 3)を用いた.電気刺激条件は上述の
4-ms
のISO
収縮負荷条件(実験条件1)でおこなった.
4.張力測定
上述のそれぞれの刺激負荷時における発揮張力は,運動装置のひずみ計を介 したコンピューター(Power Book,1400c/133)と解析装置(Mac Lab/8s,AD
Instruments Pty. Ltd.)にてモニターし,解析ソフト(Chart 3.5.6/s, AD Instruments
Pty. Ltd.)を用いて定量した.筋疲労の指標として,50
回の収縮のうち刺激開始後
5
回と終了前の5
回の張力を平均し,1,5,10セット時における1
セットの 初期張力からの変化を相対値化してグラフ化した.5.統計処理
本実験における統計量は,平均値 +/- 標準誤差(SE)として示した.統計処理
24
は全て,Prism ver.4(GraphPad Software)を用いておこなった.グループ内の初 期張力からの変化には繰り返しのある一元配置の分散分析によりおこない,有 意性が示されたときに
Bonferroni
の多重比較検定をおこない,5%未満の危険率 を有意水準とした.25
Fig.3-1. A schema of direct microscopic observation for spinotrapezius
muscle and its force development in vivo.
26
3 .結果
実験
1-1.刺激パラメーター
図
3-2
は0.2-ms
と4-ms
のパルス幅での電気刺激時における連続的なISO
収縮中の相対的な発揮張力の変化を示している.0.2-ms のパルス幅の条件下では,セ
ット内で
20-40%の張力低下が観察され,安静期間の後に初期張力付近まで回復
することが示された.一方,4-ms の条件下では,セット内における張力低下は
0.2-ms
条件と同様に20-40%程度であったが,セット間では安静期間を置いても
初期張力までの回復は観察されず,セットを繰り返すごとに初期張力の進行的 な低下が観察され,特に
10
セット時では68 +/- 8%の有意な低下が観察された(P
< 0.01)
.実験
1-2.筋収縮様式
図
3-3
はISO
収縮とECC
収縮プロトコル中に測定した筋発揮張力の相対変化 を示している.ISO
収縮群ではセット内で20-30%程度の張力低下がみとめられ,
10
セットの終了時では初期値から42 +/- 7%まで低下した(P < 0.01)
.一方で,ECC
収縮群では,セット内で張力が低下する傾向がみられたが,10セットを通 じて有意な張力低下は示されなかった.実験
1-3.筋疲労の性差
図
3-4
はMale
とFemale
のISO
収縮プロトコル中に測定した発揮張力の相対変化を示している.Maleのデータは条件
2
におけるISO
収縮群の値を用いた.Female
ではセット内で20%程度の張力低下が観察されたが有意なものではなく,
その低下は
Male
と比較すると小さく,10
セットの終了時において39 +/- 7%ま
での低下であった.27
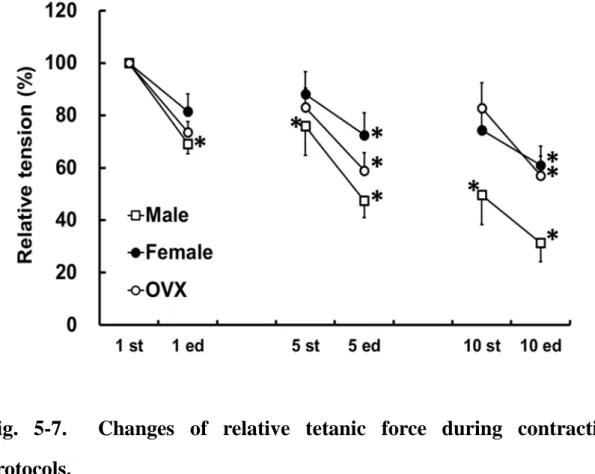
Fig.3-2. Changes of relative tetanic force during contraction protocols.
Force normalized to initial (100%) denotes the average of first and last 5
contractions of sets 1, 5, and 10 from 0.2-ms pulse duration condition
(closed circles) and 4-ms pulse duration condition (open circles). Values are
means ± SE (0.2-ms: N = 5, 4-ms: N = 4). Significance compared with
initial value for each condition: *P < 0.01.
28
Fig.3-3. Changes of relative tetanic force during ISO and ECC protocols.
Force normalized to initial (100%) denotes the average of first and last 5
contractions of sets 1, 5, and 10 from isometric (ISO, open circles) and
eccentric contraction protocol (closed circles). Values are means ± SE
(ISO: N = 3, ECC: N = 4). Significance compared with initial value for
each condition: *P < 0.05, **P < 0.01.
29
Fig.3-4. Changes of relative tetanic force during contraction protocols in male and female rats.
Force normalized to initial (100%) denotes the average of first and last 5 contractions of sets 1, 5, and 10 from male (open circles) and female rats (closed circles). Values are means ± SE (Male: N = 3, Female: N = 4).
Significance compared with initial value for each condition: *P < 0.05, **P
< 0.01.
30
4 .考察
本研究では,電気刺激条件や収縮様式,そしてラットの性別が連続的な筋収 縮刺激による筋の張力発揮に及ぼす影響を検討した.本研究により得られた知 見は以下の通りである.1) 電気刺激パルス幅が
4-ms
では0.2-ms
のものと比べ て顕著な張力低下(筋疲労)を生じる.2) ECC条件ではISO
条件と比べて張力 低下が小さい.3) Femaleと比べてMale
では顕著な筋疲労が生じた.電気刺激パラメーターが張力発揮に及ぼす影響について
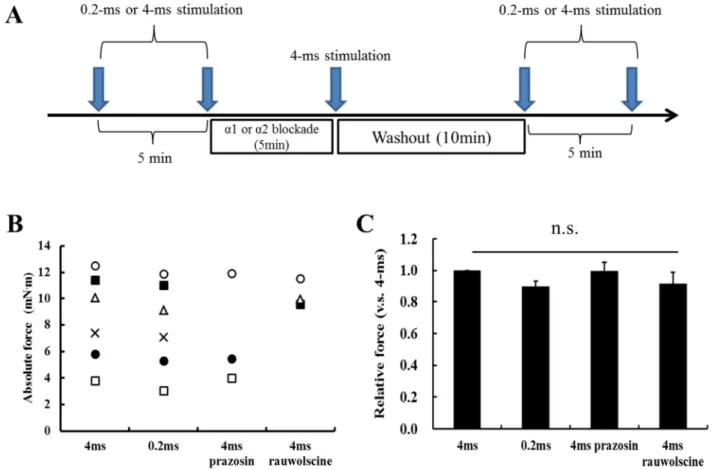
電気刺激のパルス幅の違いが筋の発揮張力動態に差を生じさせた要因として 考えられるものには,交感神経性の血管収縮が生じたかどうかにあると考えら れる.
Kjellmer
の早期の知見(Kjellmer 1964)では,短いパルス幅(0.1-ms)の刺激で は交感神経系が活性化しないことを示している.また,Honig
とFrierson (Honig
& Frierson 1976)はイヌの骨格筋において, 0.2-および 0.5-ms
のパルス幅での閉鎖 神経刺激では最大張力発揮および持続性の血管拡張が惹起されたが,2-および5-ms
のパルス幅での刺激は同様の最大張力発揮と動脈の血管収縮が惹起される ことを示している.これらの先行研究から,本実験における4-ms
のパルス幅を 用いた電気刺激では,交感神経性の血管収縮が生じ,その結果,安静期におけ る血流量の低下を招いたことで張力発揮の回復不全が生じたのではないかと考 えられた.一方で,高強度運動では,筋内圧持続的な最大収縮中において,血 管は筋内圧の上昇による機械的な圧迫を受けるために止まることは古くから知 られており(Barcroft & Millen 1939),このような状況下では酸素供給が阻害され,顕著な疲労を引き起こす.以上のことから,4-ms のパルス幅を用いた電気刺激 によって顕著な疲労が生じた要因を明らかにするためには,活動筋における血
31
液循環およびそれを制御する末梢血管動態を明らかにする必要があると考えら れる
様式の異なる筋収縮負荷が張力発揮に及ぼす影響について
これまでの先行研究から,ECCは
ISO
などの他の収縮様式と比較して,筋損 傷を誘発することが知られている.ECC 後の骨格筋において,興奮収縮連関の 異常,サルコメア構造の崩壊,細胞膜透過性の上昇を引き起こすことが示され ている. これらの障害には細胞内の[Ca2+]([Ca
2+]i)が関与することが Allen
ら により報告され,彼らはECC
収縮による機械的ストレスにより細胞膜上の機械 受容チャネル(SAC)由来の細胞内へのCa
2+流入が生じ,上昇した[Ca2+]i
が筋 損傷を誘発することを示唆している.安静時の[Ca2+]i
はECC
収縮後に10
分以 内で上昇することが単一筋線維で示されている.したがって,本実験で観察さ れた連続的なECC
収縮負荷にもかかわらず張力が維持されていた要因として,[Ca
2+]i
の上昇により筋拘縮が生じ,筋の受動張力が増加したことが考えられる.以上のことから,
ECC
収縮では筋疲労のみならず筋損傷が関与するために,ISO
収縮と比べ張力低下に差が生じたことが考えられた.性差が張力発揮に及ぼす影響について
骨格筋の疲労耐性における性差はこれまでによく議論されており,男性と比 較して女性では疲労しにくく,疲労からの回復も早いことが示されている.本 研究においても,10セットの
ISO
収縮で生じた張力の低下が雄ラットと比較し て雌ラットでは小さいことが観察された(Fig.3-6).これまでに,骨格筋において みられる性差には,エネルギー代謝や筋線維タイプ構成などが挙げられている が,一致した見解は得られていない.これまでの研究では,女性ホルモンのエ32
ストロゲンが筋の張力発生や疲労耐性に影響するという報告(Gigli & Bussmann
2001; McGoldrick et al 1998; Phillips et al 1996; Sarwar et al 1996)と影響はない (Bassey et al 1996; Greeves et al 1997; Tiidus et al 1999)という相反する報告がなさ
れており,一致した見解は得られていない.エストロゲンは運動誘発性の酸化 ストレスやそれに伴う筋疲労および筋損傷を低下させること(Tiidus 1995; Tiiduset al 1999)により脂質の代謝やグリコーゲンの保持作用といった筋の代謝を変化
させることによって,筋疲労に影響することが示唆されている(Kendrick et al1987).したがって,本研究で観察された疲労の性差を検討するためには,卵巣
摘出手術を施した雌ラットや,雄ラットに対するエストロゲン処置などの方法 を用いることが必要である.5 .要約
生体内環境下(in vivo)での筋疲労動態観察モデルの確立を目的として,ラット 脊柱僧帽筋における異なる収縮刺激負荷に対する発揮張力特性を検討した.そ
の結果,
0.2-ms
刺激と比べて,4-ms
刺激での連続的な等尺性の強縮負荷時に顕著な筋疲労が誘発されることが明らかとなった(実験
1)
.また,この4-ms
刺激 を用いた伸張性収縮負荷では,疲労の顕著な等尺性収縮とは異なり,張力発揮 が維持されることが示された(実験2)
.また,実験1
と同様の刺激プロトコル により,筋疲労には性差が生じることが示された(実験3)
.33
第 4 章 筋疲労時の末梢血管調節と交感神経系の関 与 ( 研究課題 2)
1 .背景と目的
骨格筋への血液供給の低下は,1)筋内の酸素供給不足,2)無酸素性代謝に よる代謝産物除去の停滞などを生じ,筋疲労要因の一つとして考えられる.し たがって,筋疲労時の血流応答を明らかにすることは重要である.
骨格筋血流は運動中に代謝需要に応じて増加する(Andersen & Saltin 1985).筋 収縮開始時の血流増加は筋ポンプ作用(Sheriff et al 1993)と即時の血管拡張
(Tschakovsky et al 1996)が起因すると考えられている.運動中や運動後での骨格
筋微小循環における血管緊張は,交感神経性血管収縮作用と局所で放出される 血管拡張性物質との動的平衡により決定される(Thomas & Segal 2004).運動強度 の増加とともに骨格筋血流は増大するが,筋交感神経活動もまた運動強度にと もない増加する(DiCarlo et al 1996; O'Leary et al 1997).交感神経活動の増加は非 活動部位における血管収縮を生じ,運動中の血流再分布において重要な役割を 果たしている.さらに,交感神経活動の増加は呼吸筋活動量の増加を引き起こ し,呼吸筋の機能低下が生じることで換気機能低下を惹起するため,下肢筋に おける血流の低下,およびパフォーマンスの低下を引き起こす要因となりうる(Amann et al 2010; Dempsey et al 2008; Dempsey et al 2006; Harms et al 2000; Romer
et al 2006; Sheel et al 2001).活動筋では局所的な血管拡張作用が交感神経性血管
収縮作用を抑制するため,これらの競合する作用の平衡を拡張方向へ変化させ ることで血流増加が生じると考えられている.この抑制作用は機能的交感神経 遮断と呼ばれ,1962年にRemensnyder
らにより提唱された.しかしながら,こ34
の機能的交感神経遮断にもかかわらず,交感神経性血管収縮が運動中に活動筋 における血流増加を抑制するという報告もある(Buckwalter & Clifford 1999;
DiCarlo et al 1996; Hamann et al 2002; Rowell et al 1986).微小循環調節における交
感神経系の作用には議論の余地があるものの,活動筋において筋代謝産物や一 酸化窒素(NO)がα
アドレナリン受容体の賦活による血管収縮作用を打ち消す ように作用するため,活動筋では血管収縮を生じる交感神経の活性化が減弱す ることを示した研究もある(Anderson & Faber 1991; Buckwalter et al 2001; Thomaset al 1994; 1997; Tschakovsky et al 1996).しかしながら,連続的な強縮刺激による
筋疲労時において観察される血管拡張応答の減弱が,交感神経活動の増加によ るのか,または,機能的交感神経遮断の減弱化によるのかどうかについては明 らかにされていない(Anderson & Faber 1991; Buckwalter et al 2001; Thomas et al1994; 1997; Tschakovsky et al 1996).
運動中の機能的交感神経遮断において
α
アドレナリン受容体が重要な役割を 果たすことが先行研究により示されている(Buckwalter et al 2001; Thomas et al1994).また, DeLorey
ら(DeLorey et al 2006; DeLorey et al 2007)はイヌにおける長 時間のトレッドミル走行中に交感神経性の血流抑制が生じていることをα
アド レナリン受容体阻害剤の大腿動脈内注入により確認し,長時間運動中にα
アド レナリン受容体の応答性が変化しないことも確認している.一方で,Tschakovsky
ら(Tschakovsky et al 2002)はα
アドレナリン受容体作動薬の動脈内注入は交感神 経系の全身的な活性化を生じてしまうことを指摘している.接合部後のα
アド レナリン受容体は細動脈の平滑筋上に存在するため,内腔への薬剤付加が対象 とする末梢血管のα
受容体に限局して作用するとは考えにくい.さらに,抵抗 血管の径の大きさと交感神経性血管収縮の感受性の間には逆の相関があること も示されている(Boegehold & Johnson 1988; Marshall 1982; Ohyanagi et al 1991).35



![Fig. 5-1. Schematic showing direct microscopic observation for spinotrapezius muscle and [Ca 2+ ] i in vivo](https://thumb-ap.123doks.com/thumbv2/123deta/7734920.1711770/73.892.128.785.199.688/fig-schematic-showing-direct-microscopic-observation-spinotrapezius-muscle.webp)
![Fig. 5-4. Changes of [Ca 2+ ] i in spinotrapezius muscles in vivo during 10 sets of isometric (ISO) and eccentric (ECC) contractions](https://thumb-ap.123doks.com/thumbv2/123deta/7734920.1711770/77.892.135.758.171.634/fig-changes-spinotrapezius-muscles-vivo-isometric-eccentric-contractions.webp)