Nagoya City University Academic Repository
学 位 の 種 類 博士 (薬科学) 報 告 番 号 甲第1766号 学 位 記 番 号 第363号 氏 名 小枝 暁子 授 与 年 月 日 令和 2 年 3 月 25 日 学位論文の題名 カニクイザル新鮮及び凍結肝細胞における薬物動態関連遺伝子発現の経時 変動並びに誘導プロファイルの比較 論文審査担当者 主査: 頭金 正博 副査: 松永 民秀, 湯浅 博昭, 井上 靖道
1 名古屋市立大学学位論文 カニクイザル新鮮及び凍結肝細胞における薬物動態関連遺伝子発現の 経時変動並びに誘導プロファイルの比較 令和元年度(2020 年 3 月) 名古屋市立大学大学院薬学研究科 臨床薬学分野 小枝 暁子
2 一. 本論文は、2020 年 3 月名古屋市立大学大学院薬学研究科において審査され たものである。 主査 頭金 正博 教授 副査 湯浅 博昭 教授 井上 靖道 准教授 松永 民秀 教授 二. 本論文は、学術情報雑誌に収載された次の報文を基礎とするものである。 【基礎となる報文】
1. Akiko Koeda, Takahiro Iwao, Anna Nakanishi, Shota Mizuno, Misaki Yamashita, Yoko Sakai, Katsunori Nakamura, and Tamihide Matsunaga
Comparison of mRNA Expression Profiles of Drug-Metabolizing Enzymes and Transporters in Fresh and Cryopreserved Cynomolgus Monkey Hepatocytes Drug Metab Pharmacokinet. 34, 253–263 (2019).
2. Akiko Koeda, Takahiro Iwao, Anna Nakanishi, Shota Mizuno, Misaki Yamashita, Yoko Sakai, Katsunori Nakamura, and Tamihide Matsunaga
Comparison of the Inducibility of CYP mRNA Exposed to Typical Inducers in Fresh and Cryopreserved Cynomolgus Monkey Hepatocytes
Submitted.
三. 本論文の基礎となる研究は、松永 民秀 教授の指導の下に名古屋市立大学 大学院薬学研究科において行われた。
3
略語一覧
AAALAC Association for Assessment and Accreditation of Laboratory
Animal Care International
AHR Aryl hydrocarbon receptor
ALT Alanine aminotransferase
ARNT AHR nuclear translocator
AST Aspartate aminotransferase
CAR Constitutive androstane receptor
cDNA complementary DNA
CYP Cytochrome P450
DMEM Dulbecco’s Modified Eagle’s Medium
DMSO Dimethyl sulfoxide
EGTA Ethylene glycolbis (2-aminoethylether)
N, N, N’, N’-tetraacetic acid
FBS Fetal bovine serum
FDA The U.S. Food and Drug Administration
γ-GTP γ-glutamyl transpeptidase
GAPDH Glyceraldehyde-3-phosphate dehydrogenase
HPRT Hypoxanthine-guanine phosphoribosyltransferase
LDH Lactate dehydrogenase
MDR Multidrug resistance
MHC Major histocompatibility complex
MRP Multidrug resistance-associated protein
OATP Organic-anion-transporting polypeptide
OCT1 Organic cation transporter 1
OECD Organization for Economic Co-operation and Development
OME Omeprazole
PB Phenobarbital
PCR Polymerase chain reaction
PVP Polyvinylpyrrolidone
4 RXR Retinoid X receptor RIF Rifampicin RT Reverse Transcription SULT Sulfotransferase UGT UDP-glucuronosyltransferase
5 目次 第一章 序論 ... 8 第二章 カニクイザル新鮮肝細胞及び凍結肝細胞における薬物代謝酵素関連 遺伝子及びトランスポーター遺伝子のmRNA 発現プロファイルの比較 ... 11 2.1 緒言 ... 11 2.2 実験方法 ... 13 2.2.1 試薬及び材料 ... 13 2.2.2 動物 ... 14 2.2.3 カニクイザル肝細胞の調製 ... 14 2.2.4 肝細胞の凍結方法 ... 14 2.2.5 フィーダー細胞の培養 ... 15 2.2.6 新鮮肝細胞の培養 ... 15 2.2.7 凍結肝細胞の培養 ... 15 2.2.8 Total RNA の調製 ... 15 2.2.9 リアルタイムPCR 解析 ... 16 2.2.10 逸脱酵素の測定 ... 18 2.3 結果 ... 18 2.3.1 初代カニクイザル肝細胞の調製及び細胞生存率 ... 18 2.3.2 長期培養期間中におけるカニクイザル肝細胞の形態 ... 19 2.3.3 リファレンス遺伝子とアルブミンのmRNA 発現の経時変動 ... 21 2.3.4 シトクロムP450(CYP)mRNA 発現の経時変動 ... 22 2.3.5 UDP グルクロン酸抱合酵素 mRNA 発現の経時変動 ... 24 2.3.6 薬物排出トランスポーターmRNA 発現の経時変動 ... 26
6 2.3.7 薬物取り込みトランスポーターmRNA 発現の経時変動 ... 27 2.3.8 AST,ALT,LDH,γ-GTP の漏出 ... 29 2.4 考察 ... 31 2.5 小括 ... 35 第三章 カニクイザル新鮮肝細胞及び凍結肝細胞における典型的な酵素誘導 剤によるmRNA 発現レベルの比較 ... 37 3.1 緒言 ... 37 3.2 実験方法 ... 38 3.2.1 材料 ... 38 3.2.2 動物 ... 39 3.2.3 カニクイザル肝細胞の調製及び凍結保存 ... 39 3.2.4 フィーダー細胞の培養 ... 39 3.2.5 新鮮及び凍結肝細胞の培養 ... 40 3.2.6 酵素誘導剤の曝露 ... 40 3.2.7 Total RNA の調製及びリアルタイム PCR 解析 ... 40 3.3 結果 ... 42 3.3.1 肝細胞の調製及び培養 ... 42 3.3.2 カニクイザル肝細胞に典型的な酵素誘導剤を曝露した際のシトクロ ムP450 mRNA 発現における影響 ... 42 3.3.3 カニクイザル肝細胞の核内レセプターAHR,CAR,PXR の mRNA 発現レベル ... 49 3.4 考察 ... 53 3.5 小括 ... 59 第四章 総括 ... 61
7
謝辞 ... 62 引用文献 ... 63
8 第一章 序論 医薬品開発は,新規化合物の創製に始まり,候補化合物のスクリーニングな どの基礎研究を経て,非臨床試験において有効性及び安全性を確認した後に, 初めてヒトを対象とした臨床治験へと移る.少数の健常人を対象とした第I 相, 少数の患者を対象とした第II 相,多くの患者を対象とした第 III 相の治験により 安全性と有効性が確認できた医薬品は厚生労働省へ承認申請され,独立行政法 人医薬品医療機器総合機構(PMDA)により審査される.申請が認められれば市 場での販売となり,さらに発売後の安全性や使用法についても調査される. 非臨床安全性試験には単回・反復投与毒性試験,遺伝毒性試験,がん原性試 験,生殖発生毒性試験,局所刺激性試験,依存性試験等があり,PMDA によっ て調査・認定された医薬品GLP 適合施設において実験動物を用いた試験が実施 される.厚生労働省が定めた医薬品毒性試験ガイドライン(平成元年9 月 11 日 薬審1 第 24 号,一部改正:平成 5 年 8 月 10 日薬新薬第 88 号)において,単回 及び反復毒性試験においては 2 種以上の動物を使用することとし,一種はマウ スやラットなどのげっ歯類,もう一種はウサギ以外の非げっ歯類を用いること としている.このため非げっ歯類の動物として,イヌ(ビーグル)やサル(カ ニクイザル,アカゲザル)が広く用いられ,背景データも豊富である. 欧米では動物試験に代わる代替法の確立が盛んであり,国内でも動物愛護の 観点から3R の原則(Replacement,Reduction,Refinement)の遵守が強く求めら れるようになってきている.新規医薬品の安全性を確認するのに動物実験は有 効ではあるが,動物を用いない新規代替法の開発,最小限の動物数で多くの結 果を得られる評価系の構築,苦痛の軽減や飼育環境の整備などが必要とされる. 近年,in vivo の細胞に機能を近づけた新たな細胞の作出,生体内の環境を再現
9 する培養環境の開発などが盛んに行われている. 薬物は体内に取り込まれた後,主に肝臓で代謝が行われる.肝臓から実質細 胞を分離した初代肝細胞は,様々な動物種で市販されており,生存率や接着効 率の高い良質なヒト初代肝細胞も入手可能である.肝臓中の薬物代謝酵素には 種差があり,ヒト初代肝細胞がin vitro 薬物代謝試験ではゴールドスタンダード であるが,ヒトの個人差は大きく,ロットの選定によって結果が大きく異なる 可能性がある. カニクイザルは動物実験として背景データを豊富に有しているのに加え,近 年,生息地によって遺伝的背景が異なることが分かってきた.東南アジアに生 息するカニクイザルは,大陸産(中国,カンボジア,ベトナム),島国産(イン ドネシア,フィリピン)で系統が異なることがミトコンドリアDNA の D ループ 領域の解析により明らかにされている1).また免疫にかかわるMHC 分子に着目 すると,フィリピン産のカニクイザルは MHC 遺伝子領域の多型の頻度が低く, 遺伝的に均質であることが報告されている2).このため,MHC の型がマッチし たカニクイザル間における iPS 細胞由来の心筋細胞の移植実験にも応用されて いる3). 本研究では,カニクイザルの初代肝細胞を試験材料として選択した.初代肝 細胞は 1 頭分の肝臓あるいは 1 葉程度の肝臓組織から十分量を調製することが 可能で,最低限の動物数で複数の試験が実施できる利点がある.同一個体の肝 細胞を用いた複数の化合物のスクリーニングや,また凍結肝細胞での評価が可 能であれば,個体差を考慮せずに複数回の試験に用いることができる利点があ る.本実験では遺伝的に均質な背景を有するフィリピン産のカニクイザルを選 択し,12 頭から初代肝細胞を調製し,6 頭分の新鮮肝細胞と 6 頭分の凍結肝細 胞を用いて,薬物代謝酵素関連遺伝子のmRNA 発現レベルを指標に,肝細胞の
10
機能について評価した.初めに培養中のmRNA 発現レベルの推移を追うことで
細胞の状態をモニタリングし,次に酵素誘導剤によるmRNA 発現レベルの変動
を検証し,カニクイザル肝細胞の非臨床試験におけるin vitro 薬物代謝試験への
11 第二章 カニクイザル新鮮肝細胞及び凍結肝細胞における薬物代謝酵素関連遺 伝子及びトランスポーター遺伝子のmRNA 発現プロファイルの比較 2.1 緒言 初代肝細胞の調製方法はラットにおいて40 年以上前に開発され4),近年では 様々な動物種の初代肝細胞が同様の方法で調製され,多くの実験に用いられて いる.ヒト肝臓からの初代肝細胞の調製方法も確立され,詳細に報告されてい る5, 6).それにも関わらず,初代肝細胞の調製技術は容易ではなく,高い生存率 と収量,代謝機能を有する肝細胞を得るには十分な経験を必要とする. 近年高い生存率と接着効率及び代謝機能を有する凍結肝細胞が複数のサプラ イヤーから入手が可能となった.高品質のヒト凍結肝細胞も購入可能であるが, ドナーの選択において人種や年齢の偏りや,大きな個人差等の問題が存在する. FDA ドラフトガイダンスにおいては,ヒト初代肝細胞は少なくとも 3 ドナーを 使用するよう推奨している7).このため,臨床試験で大きなバイアスを引き起こ さないためにもドナーの選択が非常に重要となっている. ヒト初代肝細胞を用いたin vitro 薬物代謝試験において,比較対照あるいは代 替細胞として,ヒト肝がん細胞株である HepaRG® 細胞や HepG2 細胞が広く用 いられている.HepaRG® 細胞は,ヒト肝細胞や HepG2 細胞と比較して,候補薬 物の酵素誘導試験や肝細胞毒性試験などのスクリーニング試験に有用との報告 がある 8-10). HepaRG® 細胞はヒト肝細胞に匹敵することが保証されているが, ドナーの表現型を反映したヒト初代肝細胞とは異なり,ロット差が無く万能で は無い. 一般的に,初代肝細胞は肝臓から単離後から肝機能を失うとされ,肝特異的 なマーカー(アルブミン分泌など)や細胞生存率,接着効率の維持などから, 従来法の単層培養は約1~2 週間が限度としている11-13).近年,肝細胞の肝特異
12 的機能の改善のため,様々な三次元(3D)培養方法が開発されている14).各々 の方法には利点があるが,3D 培養方法は未だ標準化はされていない.欧米では, 代替法として皮膚及び角膜のin vitro 3D モデルが OECD テストガイドラインに 採択されている 15-17).新規候補化合物の肝細胞毒性や薬効を評価できる肝臓モ デルが強く望まれる. 医薬品開発における非臨床試験では,マウスやラットなどのげっ歯類やイヌ などに加え,カニクイザルやアカゲザルも,霊長類であり遺伝的にヒト近いこ とからモデル動物として使われる.カニクイザルは通常東南アジアに生息し, フィリピン産のカニクイザルは MHC ハプロタイプに着目すると他のカニクイ ザル(インドネシア産及びベトナム産)と比較して遺伝的に均質であることが 報告されている1).フィリピンは,個体の移動が非常に制限されている島国であ るため,個体差が比較的大きくないと予想される.さらにカニクイザルに関す る背景データは豊富であり,ヒトへの外挿を見据えたサル試験は,ヒト臨床試 験と比較して有利であると考えられる. サルからの初代肝細胞の調製方法は,一般的に標準化されていないが,サル 初代肝細胞を用いた研究の報告は複数ある 18-22).我々の研究室では,ヒト肝細 胞の調製方法に若干の修正を加え,高品質のサル肝細胞の調製を行っている. これらの初代肝細胞は,短期培養における薬物代謝酵素誘導試験及びmRNA 発 現レベルの変動の検証に用いている23-25). 本実験では,スフェロイドを形成させる長期3D 培養法に着目し,医薬品開発 においてカニクイザル肝細胞が有用なツールとなる可能性について検証した. カニクイザル肝細胞機能の維持は,薬物代謝関連遺伝子のmRNA 発現レベルに 基づいて評価した.また3D スフェロイド培養法として Cell-able® 培養プレート を選択した.このプレートは,細胞非接着面をプレート底面にパターニングす
13 ることで,直径100 μm の肝細胞接着面に細胞スフェロイドを形成させるもので ある.肝細胞播種の前にマウス線維芽細胞をフィーダー細胞として事前に播種 することにより,フィーダー細胞非存在下と比較して,肝細胞の接着効率が改 善することが報告されている11). さらに我々は,新鮮肝細胞と凍結肝細胞のmRNA 発現レベルの違いについて も比較した.これまでにヒト新鮮肝細胞と凍結肝細胞における薬物取り込み能 について比較した報告はあったが 26),長期培養における取り込み能のモニタリ ングは行われていなかった.このため,我々はカニクイザルの新鮮及び凍結肝 細胞を調製し,28 日間まで培養し,培養期間中の mRNA 発現レベルについて検 証した 2.2 実験方法 2.2.1 試薬及び材料
Collagenase type IV 及び Dulbecco’s Modified Eagle’s Medium(DMEM)-high glucose は Sigma-Aldrich(St. Louis,MO,USA)から購入した.Williams’ medium E(WME),fetal bovine serum(FBS),penicillin(10,000 units/mL)-streptomycin (10,000 µg/mL),High-Capacity RNA-to-cDNATM Kit,TaqMan® Gene Expression
Master Mix,TaqMan® Gene Expression Assay,SYBR™ Select Master Mix は Thermo
Fisher Scientific Inc.(Waltham,MA,USA)から購入した.Dimethyl sulfoxide(DMSO) は富士フイルム和光純薬株式会社(Osaka,Japan)から購入した.RM101 medium
及びCell-able® 24-well plates は東洋合成工業株式会社(Tokyo,Japan)から購入
した.Collagen I-coated 24-well plates は AGC テクノグラス株式会社(Shizuoka, Japan)から購入した.RNeasy Mini Kit 及び QIAshredder は QIAGEN(Hilden, Germany)から購入した.3T3 Swiss albino(3T3)cells は Japan Collection of Research
14
Bioresources Cell Bank(JCRB9019,Osaka,Japan)から購入した.その他の試薬 は全て特級品を使用した. 2.2.2 動物 雄5 頭及び雌 1 頭のフィリピン産カニクイザル(2~6 歳)を使用した.本実 験は,AAALAC(国際実験動物管理公認協会)完全認証を取得している株式会 社イナリサーチにおける動物実験審査のもとで行われた. 2.2.3 カニクイザル肝細胞の調製 カニクイザル肝細胞からの調製方法は23) に従って実施した.カニクイザルに 0.33 mg/㎏のヘパリンナトリウムを静脈内投与してからチオペンタールナトリ ウム投与による麻酔下で放血致死させた.その後速やかに肝臓を摘出し,一時 的に氷上のWME 培地中に保存した.肝臓は葉にそって 4 分割し(約 10~30 g), 1 片ずつ処理した.カルシウム不含 0.6 mM EGTA 含有ハンクス溶液で脱血し, 0.05 %コラゲナーゼ含有ハンクス溶液を灌流した.肝臓は Krebs-HEPES 溶液中 で肝細胞を分散しガーゼでろ過した.肝細胞を冷 Krebs-HEPES 中で分散させ, 凍結保存または培養に用いた. 2.2.4 肝細胞の凍結方法 洗浄した肝細胞は,滅菌した冷凍結培地(2.5%PVP,2% BSA,20% FBS 含有 WME,NaHCO3でpH7.4 に調整)に再懸濁し21),細胞生存率はトリパンブルー 染色法により算出した.50×g,4°C,1 分間の遠心分離後,細胞を同じ培地に再 懸濁し,DMSO(最終濃度 10%)を加え,細胞濃度が 1×107 cells/mL となるよう 調整した.細胞は凍結バイアルに移し,-30°C で 40 分,-90°C で 15 分凍結後, 使用時まで液体窒素中で保存した22).
15 2.2.5 フィーダー細胞の培養 フィーダー細胞(Swiss 3T3 細胞)はカニクイザル肝細胞を播種する 3 日前に 培養を開始した.3T3 細胞は 10%FBS とペニシリン・ストレプトマイシン含有 DMEM 培地で培養し,24 ウェルコラーゲン I コートプレート及び Cell-able®プレ ートに4×104 cells/well になるよう播種し,5%CO 2濃度,37°C 条件で培養した. 2.2.6 新鮮肝細胞の培養 洗浄したカニクイザル肝細胞はRM101 培地に懸濁し,細胞生存率を計測した. 50×g,4°C,1 分間の遠心分離後細胞を同じ培地に再懸濁し,細胞濃度を調整し た.単層培養用には2.5×105 生細胞/ウェルとなるよう 24 ウェルコラーゲン I コ ートプレートに播種した.共培養及び3D スフェロイド培養用にはフィーダー細 胞上に1×105 生細胞/ウェルとなるよう 24 ウェルコラーゲン I コートプレートま たはCell-able® プレートにそれぞれ播種した.これらの細胞を 5%CO 2濃度,37°C 条件で培養し,播種後 3 時間,及び,以降は週 2 回培地交換を行った.単層培 養は14 日間,共培養及び 3D スフェロイド培養は 28 日間培養を行った. 2.2.7 凍結肝細胞の培養 カニクイザル凍結肝細胞は 37°C の恒温水槽中で解凍し,冷 RM101 培地に再懸 濁した.細胞は50×g,4°C,3 分間の遠心分離後,同じ培地に再懸濁した.細胞 生存率を計測し,生細胞は2.2.6 項と同様の手順で播種及び培養を行った. 2.2.8 Total RNA の調製 培養3,7,14 及び 28 日目に n=3 で細胞を回収し,total RNA の調製に使用し た.新鮮肝細胞及び凍結肝細胞はさらに培養 0 日目にも細胞を回収した.Total
16
RNA は QIAshredder 及び RNeasy Mini Kit を用いて調製した.Total RNA 濃度は NanoDrop(ND-1000,Thermo Fisher Scientific Inc.)で測定し,純度を確認した.
2.2.9 リアルタイム PCR 解析
High-Capacity RNA-to-cDNATM Kit を用いて,Total RNA から一本鎖 cDNA を合
成した.各cDNA は Nuclease-free water で 25 倍希釈した.
リアルタイムPCR は反応溶液 10 μL とし,384 ウェルプレートにて n=3 で行
った.TaqMan® Gene Expression Master Mix,300 nM フォワードプライマー及び
リバースプライマー,200 nM TaqMan®プローブの混合液,またはSYBR™ Select
Master Mix,300 nM フォワードプライマー及びリバースプライマーの混合液に, cDNA テンプレートを加え,ViiA7 リアルタイム PCR 機を用い測定を行った(プ ライマー及びプローブの塩基配列はTable1-4 に示した). データ解析は ViiA7 ソフトウェアを用い,検量線法により行った.リファレ ンス遺伝子としてHPRT を選択し,データの補正に使用し,数値は平均±S.D.(n=3) で表した.平均とS.D.は 3 頭のカニクイザルから算出した.
17
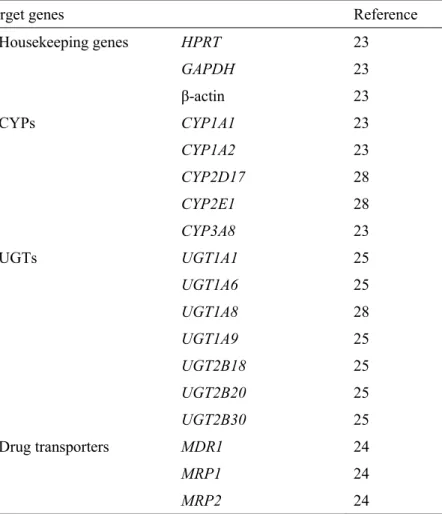
Table 1. List of target genes, primers, and probes for real-time PCR analysis.
Target genes Reference Housekeeping genes HPRT 23 GAPDH 23 β-actin 23 CYPs CYP1A1 23 CYP1A2 23 CYP2D17 28 CYP2E1 28 CYP3A8 23 UGTs UGT1A1 25 UGT1A6 25 UGT1A8 28 UGT1A9 25 UGT2B18 25 UGT2B20 25 UGT2B30 25 Drug transporters MDR1 24 MRP1 24 MRP2 24
Table 2. Primers and probe sequences of albumin for real-time PCR analysis.
Primers and probe sequences (5'→3') Albumin
Forward primer CAAGGCATACGAAGCCACTC Reverse primer TTCTGAGGCTCGTCCACAAGA TaqMan probe CTGTGCCGCTGCAAATCCTCATGAA
18
Table 3. Primers and probe of CYPs for real-time PCR analysis.
Primers and probe Assay ID* CYP2B6 Mf03043065_m1 CYP2C43 Mf04363679_m1 CYP2C75 Mf04363854_m1 *: TaqMan® Gene Expression Assay (Thermo Fisher Scientific Inc.)
Table 4. Primers of drug transporters for real-time PCR analysis.
Primer and probe sequences (5'→3') OATP1B1
Forward primer GAAGGAATCTGGGTCATACATGTG Reverse primer CCCCTATTCCCCGAAGCAT
OATP1B3
Forward primer GCTTGGTGGCTTGGTTTCC
Reverse primer GGCAAGAAAAAAAATGGTATGGA OATP2B1
Forward primer CTGCGCCTTTATGTGGACATT Reverse primer TCCTTTATGGTCAGGCTGATACC OCT1
Forward primer CGCGGGAGTGACGCTACT Reverse primer CTTCATGGTCTCTGGCAAAGC
2.2.10 逸脱酵素の測定 培養上清中のAST,ALT,LDH,γ-GTP は自動分析装置 7180(株式会社日立 ハイテクノロジーズ)を用いて測定した. 2.3 結果 2.3.1 初代カニクイザル肝細胞の調製及び細胞生存率 カニクイザル肝臓からの初代肝細胞調製方法はヒト肝臓からの肝細胞調製方 法27)を参考に,カニクイザル肝臓に適した条件に修正して行った.
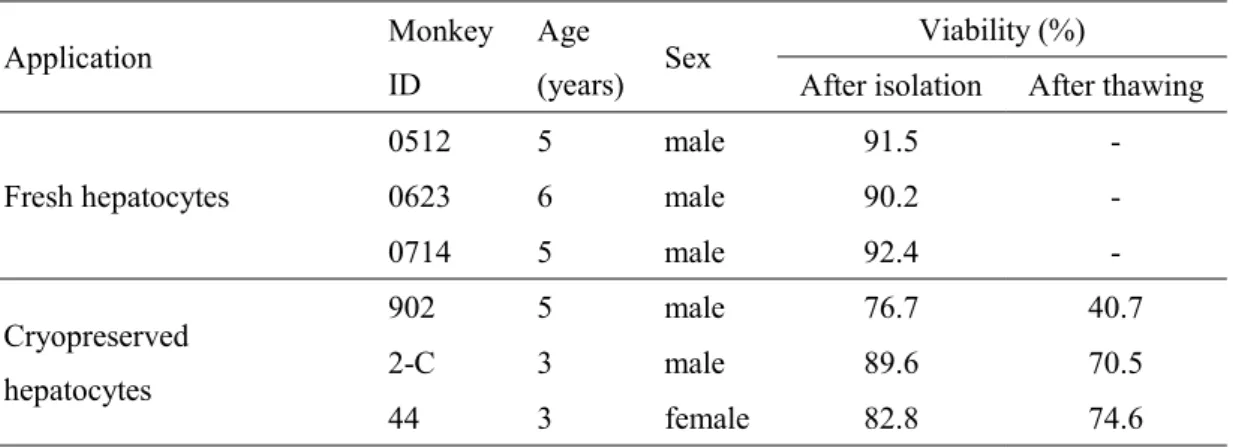
19 肝臓摘出後,灌流まで WME 培地中に肝臓を保存した.我々は,冷蔵下で 1 日または 2 日間 WME 中に肝臓を保存しても,細胞生存率及び収量に明らかな 違いがみられないことを確認している(未発表データ).このため,本実験では 肝臓を採取した当日または翌日に灌流を行った. 本実験に用いたカニクイザルのリストをTable 5 に示した.新鮮肝細胞及び凍 結肝細胞用に 3 頭のカニクイザルから初代肝細胞を調製した.調製直後の新鮮 肝細胞は高い生存率を示し(> 90%),高い効率で接着性を示した.解凍後の凍 結肝細胞の生存率は高くはなかったが(40.7–74.6%),培養期間中は長期におい て細胞の接着と維持がみとめられた.
Table 5. Characteristics of cynomolgus monkey hepatocytes used in this study.
Application Monkey ID
Age
(years) Sex
Viability (%)
After isolation After thawing Fresh hepatocytes 0512 5 male 91.5 - 0623 6 male 90.2 - 0714 5 male 92.4 - Cryopreserved hepatocytes 902 5 male 76.7 40.7 2-C 3 male 89.6 70.5 44 3 female 82.8 74.6 2.3.2 長期培養期間中におけるカニクイザル肝細胞の形態 本実験では,異なる3 つの培養方法である単層培養,フィーダー細胞との 3D スフェロイド培養,新規のフィーダー細胞との単層培養である共培養で評価し た. 新鮮肝細胞は,単層培養においては高い効率で肝細胞がプレートへ接着し, 培養3 日目でコンフルエントに達した.3D スフェロイド培養では典型的な細胞
20 の凝集がみとめられた.共培養においても肝細胞は高い効率でフィーダー細胞 へ接着した.共培養における肝細胞はより立体的に接着したが,単層培養と比 較してコンフルエントには達しなかった.培養 7 日目で小型肝細胞の増殖が認 められ,コロニー化しているように見えた(Fig. 1).培養 14 日目及び 28 日目で は小型肝細胞の数は明らかに増加し,細胞の増殖は成熟肝細胞と比較して速か った. 凍結肝細胞の細胞生存率は高くなかったが,新鮮肝細胞と同じ数の生細胞を 各培養方法で播種した.凍結肝細胞の 3 ロットはいずれの培養方法においても 接着能を示したが,新鮮肝細胞と比較して凍結肝細胞では浮遊した細胞がみと められ,細胞密度は低かった.しかしながら培養 7 日目で小型肝細胞が認めら れ,新鮮肝細胞の時と同様に増殖がみとめられた.
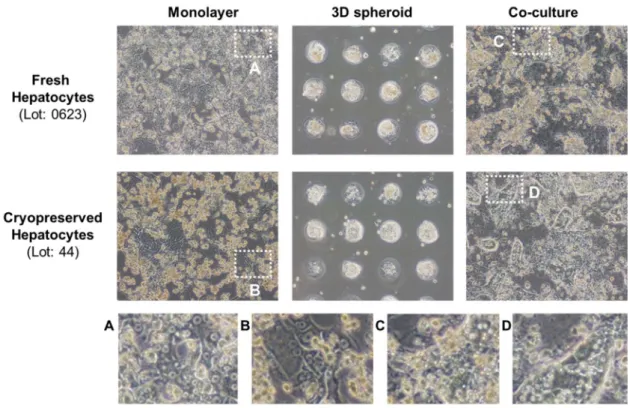
Figure 1. Morphology of cynomolgus monkey hepatocytes on day 7.
Microscopic images of monolayer culture, 3D spheroid culture, and co-culture of cynomolgus monkey hepatocytes. Freshly isolated hepatocytes (Lot 0623) and cryopreserved hepatocytes (Lot 44) were cultured with each culture method.
21 2.3.3 リファレンス遺伝子とアルブミンの mRNA 発現の経時変動 HPRT,GAPDH 及び β-actin は一般的に広く用いられているリファレンス遺伝 子であるが,各培養条件におけるmRNA 発現が安定しているリファレンス遺伝 子を選択するため,本実験ではそれぞれのmRNA 発現量を別のリファレンス遺 伝子で補正し,一定であるか否かについて検証した.また肝機能維持を検証す るため,肝特異的なマーカーであるアルブミンも選択した. 新鮮肝細胞における培養3 日目の HPRT で補正した GAPDH の mRNA 発現レ ベルは,単層培養で14 日間,3D スフェロイド培養及び共培養で 28 日間まで安 定していた(Fig. 2).凍結肝細胞においては,培養初日の mRNA 発現レベルは 新鮮肝細胞よりも低かったものの,培養 3 日目以降は維持されていた.新鮮及 び凍結肝細胞のいずれも,HPRT で補正後の GAPDH mRNA 発現レベルは培養 3 日以降いずれの培養条件においてもほぼ一致し,本実験における培養期間中は どちらのmRNA 発現レベルも同じような変動を示していたことが示唆された. 我々の過去の報告23)と同様に,HPRT を本実験のリファレンス遺伝子として用い ることとした.一方,HPRT で補正後の β-actin mRNA 発現レベルは培養条件に よって特に凍結肝細胞において差がみとめられ,このためβ-actin はリファレン ス遺伝子としては使用しなかった. アルブミンmRNA 発現レベルは新鮮肝細胞及び凍結肝細胞のいずれの培養条 件においても,培養初日から3 日目にかけて減少した.しかし 3D スフェロイド 培養及び共培養においては,その後培養初日のレベルまで発現量が回復し,培 養28 日目まで維持された.単層培養では,mRNA 発現レベルは新鮮肝細胞にお いて培養14 日目まで徐々に減少したのに対し,凍結肝細胞では中程度の抑制の みとなった.
22
Figure 2. Time courses of mRNA expression of housekeeping genes and albumin.
mRNA expression of housekeeping genes (GAPDH and β-actin) and albumin was determined by RT-qPCR during the culture period of 14 (monolayer culture) and 28 days (3D spheroid culture and co-culture). The data were normalized to HPRT (reference gene) and are represented as the mean ± S.D. (n = 3) of freshly isolated hepatocytes (A) and cryopreserved hepatocytes (B).
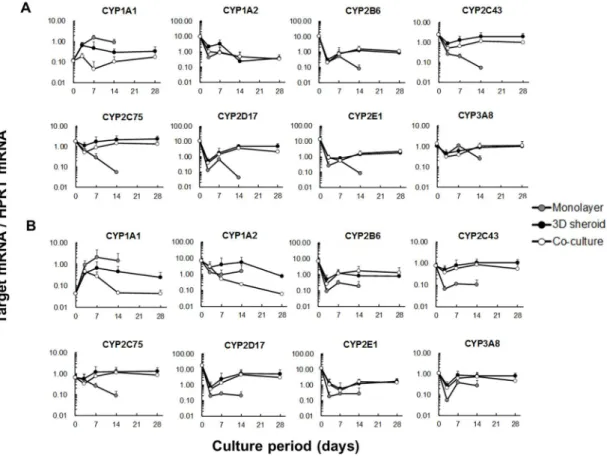
2.3.4 シトクロム P450(CYP)mRNA 発現の経時変動 CYP1A1 mRNA 発現レベルは新鮮肝細胞と凍結肝細胞でほぼ同じ挙動を示し たが,培養方法によって明らかに発現レベルが異なった(Fig 3).全てのグルー プにおいてmRNA 発現レベルは培養初日から 3 日目にかけて増加し,単層培養 では培養 7 日目までさらに増加して 3 種の培養条件において最も高い発現レベ ルを示す結果となった.3D スフェロイド培養では,mRNA 発現レベルは新鮮肝 細胞では培養 3 日目,凍結肝細胞では培養 7 日目でピークとなった.共培養で は,mRNA 発現レベルは培養 7 日目以降で新鮮と凍結肝細胞でばらついた結果 となったが,培養14 日目以降は,培養初日とほぼ同じレベルであった. CYP1A2 mRNA 発現レベルは,カニクイザル肝細胞では低いことが報告され
23 ている28).CYP1A2 の検量線において,cDNA テンプレートを 5 濃度調製したが, 高いほうから 3 濃度しか検出できなかった.本実験では,複数のサンプルで定 量下限以下となり,mRNA 発現レベルがサンプル間で大きく異なっていたこと から,検出されたサンプルの測定値が高くみなされることとなった.新鮮及び 凍結肝細胞において,CYP1A2 の mRNA 発現レベルは,培養初日と比較して培 養期間中に減少していく結果となった. CYP2C43,CYP2C75,CYP3A8 mRNA 発現レベルは新鮮及び凍結肝細胞のい ずれも似通った変動を示した.これらCYPs の mRNA 発現レベルは培養初日か ら3 日目までに減少し,3D スフェロイド培養及び共培養においては,培養 7 日 目または 14 日目までに回復後,培養 28 日目まで維持されていた.一方で,単 層培養においては培養14 日目までに劇的に mRNA 発現量が減少し,3 種の培養 条件でも最も低いレベルを示す結果となった. CYP2B6,CYP2D17,CYP2E1 の mRNA 発現レベルは,新鮮及び凍結肝細胞 で似通った変動を示したが,3D スフェロイド培養及び共培養においては培養初 日のレベルまでは回復しなかった.
CYP1A1 及び CYP1A2 を除く CYPs の mRNA 発現レベルは 3D スフェロイド
培養と共培養で非常に似通った結果となり,培養 14 日目と 28 日目でほぼ変化
24
Figure 3. Time courses of mRNA expression of CYPs.
mRNA expression of eight CYPs (CYP1A1, CYP1A2, CYP2B6, CYP2C43, CYP2C76, CYP2D17, CYP2E1, and CYP3A8) was determined by RT-qPCR during the culture period of 14 (monolayer culture) and 28 days (3D spheroid culture and co-culture). The data were normalized to HPRT (reference gene) and are represented as the mean ± S.D. (n = 3) of freshly isolated hepatocytes (A) and cryopreserved hepatocytes (B). The following data are represented as the mean (n = 2) because one of three values was below the quantitation limit: at day 14 and 28 in the 3D spheroid culture of fresh hepatocytes, at day 28 in the 3D spheroid culture of cryopreserved hepatocytes, and at day 7, 14, and 28 in the co-culture of cryopreserved hepatocytes.
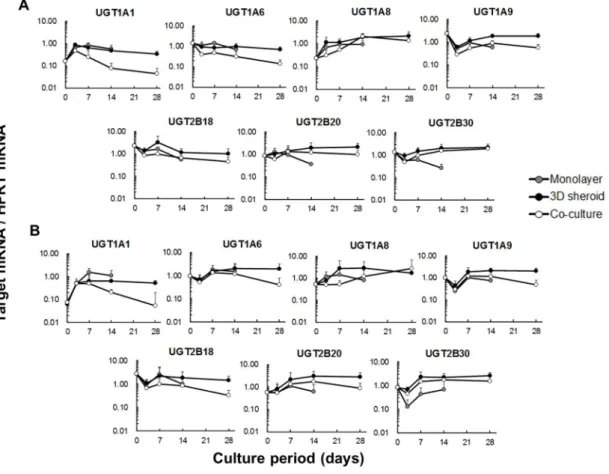
2.3.5 UDP グルクロン酸抱合酵素 mRNA 発現の経時変動
UGT1A1 mRNA 発現レベルは,新鮮及び凍結肝細胞のいずれも培養初日から 3 日目にかけて増加した(Fig 4).mRNA 発現レベルは単層培養で最も高く,共培 養が最も低かった.
25 日目までは全ての培養条件で増減のパターンがほぼ一致し,以降,共培養では 減少したが 3D スフェロイド培養では変化はなかった.新鮮肝細胞においては, 培養3 日目以降,3D スフェロイド培養と共培養で違いはみられなかった. UGT1A8 mRNA 発現レベルは,新鮮及び凍結肝細胞のいずれも RT-qPCR 測定 時にCt 値が 35 前後になるなど他の UGTs と比較して低い発現量を示し,特に凍 結肝細胞において個体差も大きかった.他の UGT1As と異なり,共培養におい てmRNA 発現レベルの増加がみとめられた. UGT2B18 mRNA 発現レベルは培養 3 日目で減少,培養 7 日目で増加し,その
後徐々に減少した.UGT2B20 及び UGT2B30 mRNA 発現レベルは,単層培養を
除き培養7 日目から徐々に増加する傾向となった.
殆どのUGT 分子種において,mRNA 発現レベルは 3D スフェロイド培養で高
く維持される傾向となった.また凍結肝細胞において個体差が大きくなる傾向 がみられた.
26
Figure 4. Time courses of mRNA expression of UGTs.
mRNA expression of seven UGTs (UGT1A1, UGT1A6, UGT1A8, UGT1A9, UGT2B18, UGT2B20, and UGT2B30) was determined by RT-qPCR during the culture period of 14 (monolayer culture) and 28 days (3D spheroid culture and co-culture). The data were normalized to HPRT (reference gene) and are represented as the mean ± S.D. (n = 3) of freshly isolated hepatocytes (A) and cryopreserved hepatocytes (B). 2.3.6 薬物排出トランスポーターmRNA 発現の経時変動 排出トランスポーターである MRP1,MRP2 及び MDR1 はカニクイザル肝細 胞で発現していることが知られているが,MRP1 mRNA 発現レベルは他の排泄 トランスポーターよりも低い24, 28). MRP1 及び MDR1 mRNA 発現レベルは全ての培養条件において,新鮮及び凍 結肝細胞のいずれも非常に似通った変動を示した(Fig 5).どちらも mRNA レ
27 ベルも培養初日から 3 日目あるいは 7 日目にかけて増加し,その後変化はみら れなかった.さらに単層培養で最も高く,共培養で最も低い発現レベルを示し た. 一方で,MRP2 mRNA 発現レベルは新鮮及び凍結肝細胞のいずれにおいても 個体差が大きくなる傾向がみられ,3 個体のうち 1 個体は,他の 2 個体とは異な る発現プロファイルを示した.しかしながら培養初日と比較して,培養28 日目 のmRNA 発現レベルに明らかな違いは認められなかった.
Figure 5. Time courses of mRNA expression of drug efflux transporters.
mRNA expression of three efflux transporters (MDR1, MRP1, and MRP2) was determined by RT-qPCR during the culture period of 14 (monolayer culture) and 28 days (3D spheroid culture and co-culture). The data were normalized to HPRT (reference gene) and are represented as the mean ± S.D. (n = 3) of freshly isolated hepatocytes (A) and cryopreserved hepatocytes (B).
2.3.7 薬物取り込みトランスポーターmRNA 発現の経時変動
28
OCT1 は,新鮮及び凍結肝細胞で似通った変動を示した(Fig 6).OATP1B1, OATP1B3 及び OCT1 mRNA 発現レベルにおいて,特に OATP1B3 は単層培養で
培養14 日目までに減少した.OATP2B1 mRNA レベルは新鮮肝細胞で 3 種の培
養条件で似たような変動を示したが,凍結肝細胞では単層培養の培養 3 日目が
非常に低く,共培養の培養14 日目以降で増加したため,培養形態により大きく
異なる結果となった.
Figure 6. Time courses of mRNA expression of drug uptake transporters
mRNA expression of four drug uptake transporters (OATP1B1, OATP1B3, OATP2B1 and OCT1) was determined by RT-qPCR during the culture period of 14 (monolayer culture) and 28 days (3D spheroid culture and co-culture). The data were normalized to HPRT (reference gene) and are represented as the mean ± S.D. (n = 3) of freshly isolated hepatocytes (A) and cryopreserved hepatocytes (B).
29
Table 6. mRNA expression levels of fresh and cryopreserved hepatocytes on day 0.
Target genes Fresh hepatocytes Cryopreserved hepatocytes GAPDH 0.540 ± 0.419 0.351 ± 0.026 β-actin 0.138 ± 0.036 0.072 ± 0.029 Albumin 3.91 ± 0.26 3.90 ± 3.33 CYP1A1 0.123 ± 0.035 0.0434 ± 0.0292 CYP1A2 9.71 ± 4.70 7.02 ± 6.09 CYP2B6 11.0 ± 9.1 7.09 ± 4.46 CYP2C43 2.46 ± 1.96 0.795 ± 0.240 CYP2C75 1.83 ± 1.34 0.663 ± 0.179 CYP2D17 11.1 ± 5.9 17.6 ± 10.5 CYP2E1 13.5 ± 8.4 11.9 ± 3.4 CYP3A8 1.03 ± 0.34 1.06 ± 0.13 UGT1A1 0.163 ± 0.140 0.0753 ± 0.0375 UGT1A6 1.42 ± 0.89 0.927 ± 0.201 UGT1A8 0.226 ± 0.105 0.527 ± 0.466 UGT1A9 2.38 ± 0.78 0.993 ± 0.324 UGT2B18 2.27 ± 1.15 2.78 ± 0.98 UGT2B20 0.874 ± 0.748 0.563 ± 0.170 UGT2B30 1.36 ± 1.47 0.828 ± 0.320 MRP1 0.105 ± 0.065 0.0831 ± 0.0649 MRP2 0.122 ± 0.118 0.0993 ± 0.0847 MDR1 0.906 ± 0.717 0.503 ± 0.184 OATP1B1 2.21 ± 0.42 1.84 ± 0.41 OATB1B3 8.25 ± 1.84 8.86 ± 1.84 OATP2B1 1.33 ± 0.30 0.877 ± 0.049 OCT1 0.866 ± 0.193 1.06 ± 0.31 2.3.8 AST,ALT,LDH,γ-GTP の漏出 培養期間中の細胞ダメージを検証するため,培養上清中の AST,ALT,LDH 及びγ-GTP の漏出を測定し,1 日あたりの漏出量(mU/well)で表した.AST,
30 ALT 及び LDH は細胞膜ダメージや細胞死により細胞から漏出する酵素であり, 細胞毒性マーカーとして広く使われている.γ-GTP は胆汁うっ滞または胆管細胞 の損傷が起きると細胞や胆管から漏出し,肝胆道系の特異的な酵素である. γ-GTP の数値は 3D スフェロイド及び共培養でのフィーダー細胞による影響はな い. AST 及び ALT の数値は新鮮及び凍結肝細胞のいずれも培養 3 日目が最も高か った(Fig 7).その後,培養 7 日目または 14 日目までに減少し,培養 28 日目ま で変化はなかった.LDH の数値はほぼすべての細胞において定量限界以下とな り,LDH の漏出は極めて低いことが分かった.γ-GTP の数値は 3D スフェロイ ド培養でもほぼ定量限界以下となった.新鮮及び凍結肝細胞のいずれも,γ-GTP の漏出は共培養で徐々に減少したが,一方で単層培養では徐々に増加した.フ ィーダー細胞からのこれらの漏出酵素はほぼ定量限界以下であった. 以上から,AST,ALT,LDH 及び γ-GTP の検出濃度は非常に低く,単層培養 では14 日間,共培養及び 3D スフェロイド培養では 28 日間細胞が生存していた ことが裏付けられた.
31
Figure 7. Leakage of AST, ALT, and γ-GTP in culture medium.
AST, ALT, and γ-GTP levels in the culture medium were measured during the culture period of 14 (monolayer culture) and 28 days (3D spheroid culture and co-culture). The data were calculated as mU/well/day and are represented as the mean ± S.D. (n = 3) of freshly isolated hepatocytes (A) and cryopreserved hepatocytes (B). 2.4 考察 本研究では,カニクイザルから調製した新鮮及び凍結肝細胞を用いて,薬物 代謝酵素遺伝子のmRNA 発現レベルの違いについて比較した.培養初日では, 25 遺伝子のうち 19 遺伝子において,凍結肝細胞よりも新鮮肝細胞で mRNA レ ベルが高く発現していた(Table 6).しかし,ほとんどの遺伝子は新鮮及び凍結 肝細胞で培養期間中ほぼ同等に発現し,明らかな違いはみとめられなかった. 本実験では,同じ個体から得られた新鮮及び凍結肝細胞のmRNA 発現レベル について検証しようと試みたが,凍結肝細胞はおいて解凍後の細胞生存率及び 接着効率が低く,長期培養には不向きであった.そのため,細胞生存率と接着 率が確認できており,長期培養も問題ないと思われた別個体の凍結肝細胞を使 用した.培養28 日間の mRNA 発現レベルの経時変動を評価したところ,細胞源
32 として異なる 3 個体を使用したにもかかわらず,それぞれの肝細胞で明らかな 違いはみとめられなかった.UGT1A8 及び MRP2 mRNA レベルにおいて,3 個 体のうち 1 個体のみが培養期間中で異なる変動パターンを示すなどの個体差が みられたが,UGT1A8 は mRNA 発現レベルが極めて低いことにより細胞の状態 による影響を受けやすいことが原因として考えられた.また MRP2 は胆管側の 細胞膜上に局在する排出トランスポーターであることから,小型肝細胞の増殖 と成熟化が個体間で不均一に進行し,毛細胆管の形成に伴いMRP2 の mRNA 発 現量に個体差が生じた可能性も考えられた.他の遺伝子については個体差によ る明らかな違いはみられなかった.よってカニクイザル肝細胞は,異なる個体 を由来としても同じ培養条件においてはmRNA レベルは似たような挙動を示す ことが示唆された.しかし,短期間の培養におけるmRNA レベルの変動につい ては,さらなる検証が必要と考える. 本実験では雌のカニクイザルも使用しており,培養初日における CYP1A1 及 びCYP1A2 の mRNA 発現レベルは雄の方が高いレベルを示していた.しかしこ れまでの報告によると29-32),薬物代謝酵素のmRNA 発現レベル及びミクロソー ム酵素活性は,カニクイザルにおいて性差がないことが示されている.培養期 間中における雄と雌のmRNA 変動パターンにおいて明確な違いがなかったこと から,性差については考慮しなかった. さらに,単層培養,マウス線維芽細胞と共存させた3D スフェロイド培養及び 共培養法について,mRNA レベルの経時変動についても検証した.従来法であ る単層培養は医薬品開発段階においてはスタンダードな手法とされている.近 年,単層培養は新たに開発された3D 培養システムの有用性を検証するための比 較コントロールとして用いられることが多い.実際に,今回の3D スフェロイド 培養法は,単層培養と比較して,アルブミンや CYPs の mRNA 発現レベルや
33 γ-GTP の漏出などを含めて,高い肝機能の維持を可能としている.本実験では, 単層培養とは異なり,3D スフェロイド培養と似通った結果を示す,肝細胞とフ ィーダー細胞による共培養法とも比較した.この結果は,フィーダー細胞の存 在により改善がみられた肝細胞の接着率にも寄与し,3D スフェロイド構造を形 成するよりも,肝細胞とフィーダー細胞との相互作用がより重要であることを 示唆している.肝臓は,主に肝実質細胞,星細胞,類洞内皮細胞,クッパー細 胞から構成されているが,このうち星細胞はコラーゲンなどの細胞外マトリッ クスや,サイトカイン,増殖因子などを分泌し細胞増殖に関わっている.フィ ーダー細胞である繊維芽細胞からも,サイトカイン等の液性因子が分泌され, 肝細胞の接着や機能維持に大きく関与していると考えられ,フィーダー細胞の 存在が非常に有用であることが示唆された.例外として,CYP1A1 及び UGT1A1 のmRNA 発現レベルはいずれも共培養法において速やかに減少した.この結果 の理由についてはさらに検証が必要である. カニクイザル肝細胞を単層培養で培養中に,培養 7 日目で小型肝細胞が確認 され,コロニー様に増殖が見られた.この現象はフィーダー細胞と共存させた 共培養法においても観察された.3D スフェロイド培養においては,成熟肝細胞 と小型肝細胞を見分けるのは困難であった.おそらく高密度の細胞の凝集は小 型肝細胞の増殖によるものと考えられた. これまでの報告で,ラット肝細胞においては小型肝細胞の増殖がみとめられ ている33, 34).肝細胞を線維芽細胞と30 日間共培養すると,アルブミン生成量が 増加し小型肝細胞が増殖する.さらに小型肝細胞のCYP 蛋白発現についても検 証され,テストステロンの代謝プロファイルは成熟肝細胞と似ていることが示 されている35). カニクイザル由来の小型肝細胞は強い増殖能を示し,ラット小型肝細胞と同
34 様に36),数回の継代が可能であった.さらにラット小型肝細胞はCYP 及びトラ ンスポーターの遺伝子発現レベルが減少し,継代回数が増えると細胞分裂がゆ っくりになることも報告されている.しかし,本実験においては小型肝細胞の mRNA レベルや機能については検証しなかった.このためカニクイザルの小型 肝細胞の機能について検証が今後必要になるかもしれない.
カニクイザルCYPs の mRNA 発現レベルは,CYP1A1 及び CYP1A2 を除き培
養14 日目と培養 28 日目でほぼ同等であったことから,これらの mRNA 発現量 が維持されている期間で実験を行うと,安定したデータが得られることが予想 され,また CYP1A1,CYP2C43,CYP2C75,CYP3A8 は培養初日と同等の実験 結果を得られる可能性が示唆された.なお培養14 日目以降では,肝細胞中に発 現するCYP mRNA 発現量の比率は培養初日と大きく変化していると考えられる が,mRNA 発現量に依存した CYP 遺伝子発現の誘導,あるいは代謝酵素の発現 量に依存した代謝物濃度の正確な定量は難しいものの,酵素誘導スクリーニン グ試験や,代謝物の探索などの定性には有用であると考える. 我々は以前に,カニクイザルのCYP の mRNA 誘導プロファイルはヒトと似て いることを示している23, 24).多くの薬物はCYP1A または CYP3A によって代謝 されるため,医薬品開発段階においてカニクイザル肝細胞の使用は酵素誘導試 験やスクリーニング試験に有用である.カニクイザルを用いる利点の 1 つとし て,新鮮初代肝細胞を使用できることが挙げられる.例えば,考えられるオプ ションとして,医薬品毒性試験や薬理試験などのin vivo 試験から得られた肝臓 の使用,肝臓組織中と初代肝細胞中のmRNA 発現レベルの比較,代謝プロファ イルのバリデーション等がある.医薬品開発において酵素活性とともに mRNA 発現レベルの評価は重要であることがFDA ガイダンスに示されている7).開発 段階においては,新規化合物が遺伝子発現への影響を与える可能性について検
35 証することは有用である.以前に報告されているように,ヒトにおけるCYP1A1, CYP1A2,CYP3A4,CYP2D6 の mRNA 発現レベルは代謝活性と高い相関性がみ とめられている37, 38).一方,CYP2E1,CYP2C9,CYP2A6 の mRNA 発現レベル は相関性が低く,転写効率の違いやmRNA 及び酵素の安定性,遺伝子多型等が 理由として考えられている.mRNA 発現解析のみでは完全な評価系ではないこ とは理解しているが,候補医薬品の影響を検証する強いバイオマーカーである と考えている. mRNA 発現レベルの検証に加え,肝細胞傷害の程度についても検証を行った が,一般的な肝細胞傷害マーカーであるAST,ALT は,培養 3 日目が最も高く, 肝細胞分離あるいは凍結融解の工程の影響を受けて細胞膜ダメージを受けてい たと考えられる.このため培養 3 日目頃までは肝細胞が新しい環境に適応する ために,細胞の修復などが行われている期間なのかもしれない.一方で γ-GTP が単層培養において増加し,明らかに共培養と3D スフェロイド培養と異なる挙 動を示したが,これはわずかに混在していた胆管細胞が単層培養において増殖 し,γ-GTP の細胞外分泌が増加した可能性も考えられた. 本実験では,共培養方法は低い接着能の細胞においても接着率を改善できる 有用な方法であることを示した.さらに共培養法は長期培養も可能とした.よ って Cell-able® プレートを使用する 3D スフェロイド培養法よりも費用対効果 が高く,また酵素誘導や毒性試験のような様々な評価システムに使用可能と考 えられる.酵素活性や蛋白発現レベルへの影響評価は本研究の範囲を超えたも のであるが,今後の研究には必要であると考える. 2.5 小括 カニクイザル新鮮及び凍結肝細胞を 3 種類の方法で培養し,長期培養期間中
36
のmRNA 発現レベルの違いについて比較した.新鮮肝細胞と凍結肝細胞の間で
明確な違いはみとめられなかったが,遺伝子発現レベルの維持には,3D 構造の 形成よりも,フィーダー細胞の存在がより重要であることが示唆された.小型 肝細胞の増殖及び影響についてはさらなる検証が必要である.
37 第三章 カニクイザル新鮮肝細胞及び凍結肝細胞における典型的な酵素誘導剤 によるmRNA 発現レベルの比較 3.1 緒言 カニクイザルは医薬品開発段階において,毒性試験や薬効試験などの非臨床 試験に広く用いられている.カニクイザルはマウス,ラット,イヌなどの一般 的な実験動物と比べて遺伝的にヒトと近いという利点がある.実際に,薬物代 謝関連酵素においてはヒトとの相同性が報告されている 39-41).実験動物として カニクイザルは飼育環境(年齢,産地,給餌など)をコントロールすることが 可能で,ヒトの個人差(食事,服薬,生活習慣に起因するもの)をカバーする ことができると考えられる.さらにカニクイザルの背景データは豊富で,非臨 床試験において信頼性の高いデータを得ることが可能である. 薬物の多くは肝臓で代謝されるため,医薬品開発の初期において新規化合物 の肝細胞毒性評価や,代謝物プロファイルの予測など,初代肝細胞は非常に有 用なツールとなっている.市販で入手可能はヒト初代肝細胞の使用はFDA ガイ ダンス 7) にも記載されているように,新規医薬品の酵素誘導試験,薬物相互作 用評価への使用を推奨している.ヒト凍結肝細胞は複数の会社から購入可能で あるが,人種の偏りに加え,多くのドナーに薬歴があり,高齢ドナーが多い. よって肝細胞を選択するのにドナーの背景を考慮しなければならない.医薬品 開発の初期段階におけるスクリーニングでは,個人差が評価に与える影響は小 さい方が望ましい. FDA ガイダンスでは,CYP1A2,CYP2B6,CYP2C8,CYP2C9,CYP2C19 及 びCYP3A4/5 について新規薬物の CYP 誘導能について評価するよう定めており, オメプラゾール(CYP1A2),フェノバルビタール(CYP2B6)及びリファンピシ ン(CYP2C8,CYP2C9,CYP2C19,CYP3A4/5)を陽性対照として推奨している 42).これまでの報告から,カニクイザル肝細胞はヒト肝細胞と同じ調製方法で,
38 肝全葉あるいは肝切片から調製できることを示した.3 個体のカニクイザル肝細 胞を用いた短期培養での酵素誘導試験を実施した結果から,CYP1A1(CYP1A2 ではない)及びCYP3A(ヒト:CYP3A4,カニクイザル:CYP3A8)mRNA 発現 は,ヒトとサルで似たメカニズムで制御されることを示した23). 酵素誘導は遺伝子の転写を活性化させる核内レセプターによって制御されて
いる.AHR はヒト CYP1A1 及び CYP1A2,PXR は CYP2B6,CYP2C9,CYP2C19,
CYP3A4 の酵素誘導に,CAR は CYP2B6 及び CYP3A4 の誘導に関わる43-46).複
数の化合物が核内レセプターを活性化するが,リファンピシンはPXR の活性化 物質としてよく知られている.典型的なAHR 活性化物質としてオメプラゾール, CAR 活性化物質としてフェノバルビタールがある.核内レセプターを活性化す る化合物には種差がある 47, 48).例えばオメプラゾールとリファンピシンは,ラ ットのCYP1A 及び CYP3A を誘導しない23). 本実験では,カニクイザル肝臓から新鮮及び凍結肝細胞を調製し,3 種類の異 なる培養条件で培養した際の主なCYPs の酵素誘導能を比較し,CYPs 及び核内 レセプターのmRNA 発現レベルについて検証した. 3.2 実験方法 3.2.1 材料
FBS,penicillin(10,000 units/mL)-streptomycin(10,000 µg/mL),High-Capacity RNA-to-cDNATM Kit , TaqMan® Gene Expression Master Mix , TaqMan® Gene
Expression Assay,SYBR™ Select Master Mix は Thermo Fisher Scientific Inc. (Waltham,MA,USA)から購入した.DMSO は富士フイルム和光純薬株式会
社(Osaka,Japan)から購入した.RM101 medium 及び Cell-able® 24-well plates
39
plates は AGC テクノグラス株式会社(Shizuoka,Japan)から購入した.RNeasy Mini Kit 及び QIAshredder は QIAGEN(Hilden,Germany)から購入した.3T3 Swiss albino (3T3)cells は Japan Collection of Research Bioresources Cell Bank(JCRB9019, Osaka,Japan)から購入した.その他の試薬は全て特級品を使用した. 3.2.2 動物 雄10 頭及び雌 2 頭のフィリピン産カニクイザル(2~6 歳)を使用した.本実 験は,AAALAC 完全認証を取得している株式会社イナリサーチにおける動物実 験審査のもとで行われた. 3.2.3 カニクイザル肝細胞の調製及び凍結保存 カニクイザル肝細胞の調製方法及び凍結方法は 2.2.3 項及び 2.2.4 項に従い実 施した.6 頭のカニクイザル(3 頭を新鮮肝細胞用,3 頭を凍結肝細胞用)をオ メプラゾール及びリファンピシン曝露試験に使用した.さらに別の 6 頭のカニ クイザル(3 頭を新鮮肝細胞用,3 頭を凍結肝細胞用)をフェノバルビタール曝 露試験に使用した. 3.2.4 フィーダー細胞の培養 フィーダー細胞(Swiss 3T3 細胞)は 10%FBS とペニシリン・ストレプトマイ シン含有 DMEM 培地で培養し,24 ウェルコラーゲン I コートプレート及び Cell-able®プレートに4×104 cells/well になるよう播種し,カニクイザル肝細胞を 播種するまでの3 日間,5%CO2濃度,37°C 条件で培養した.
40 3.2.5 新鮮及び凍結肝細胞の培養 新鮮及び凍結肝細胞の培養については 2.2.6 項及び 2.2.7 項に記載した方法で 行った. 新鮮肝細胞及び凍結肝細胞は,単層培養用には 2.5×105 生細胞/ウェルとなる よう24 ウェルコラーゲン I コートプレートに播種した.共培養及び 3D スフェ ロイド培養用にはフィーダー細胞上に 1×105 生細胞/ウェルとなるよう 24 ウェ ルコラーゲンI コートプレートまたは Cell-able® プレートにそれぞれ播種した. RM101 を培養培地とし,5%CO2濃度,37°C 条件で培養した.培養期間は,一般 的な短期培養として3 日間,長期培養として 28 日間を選択し,推移を検証する ために7 日間と 14 日間の培養も行った. 3.2.6 酵素誘導剤の曝露 リファンピシン,オメプラゾール及びフェノバルビタールはDMSO に溶解し, それぞれ50 mM,50 mM,1000 mM のストック溶液を調製した.使用時に,ス トック溶液をRM101 培地で 1000 倍希釈し,それぞれ 50 μM,50 μM,1000 μM (最終DMSO 濃度は 0.1%)とした.0.1%DMSO 含有 RM101 培地を媒体コント ロールとした. 培養2 日目,4 日目,11 日目,25 日目に誘導剤を含む培地に交換し,短期培 養では24 時間,それ以外は 3 日間 n=3 で曝露した. 3.2.7 Total RNA の調製及びリアルタイム PCR 解析 培養3 日目,7 日目,14 日目,28 日目に細胞を回収し,Total RNA 調製に使用
した.Total RNA は QIAshredder 及び RNeasy Mini Kit を用いて調製した.Total
41
High-Capacity RNA-to-cDNATM Kit を用いて,Total RNA から一本鎖 cDNA を合
成した.各cDNA は Nuclease-free water で 25 倍希釈しリアルタイム PCR 測定に
使用した.HPRT をリファレンス遺伝子として補正に用いた.CYP1A1,CYP3A8
及びHPRT の発現解析には TaqMan® Gene Expression Master Mix,300 nM フォワ
ードプライマー及びリバースプライマー,200 nM TaqMan®プローブの混合液に
cDNA テンプレートを加えた.PXR,AHR 及び CAR の発現解析には SYBR™ Select Master Mix,300 nM フォワードプライマー及びリバースプライマーの混合
液にcDNA テンプレートを加えた(プライマーの塩基配列は Table 7 に示した).
CYP2B6,CYP2C43 及び CYP2C75 の発現解析には TaqMan® Gene Expression
Master Mix 及 び TaqMan® Gene Expression Assay ( そ れ ぞ れ Assay ID:
Mf03043065_m1,MF04363679_m1,Mf04363854_m1)の混合液に cDNA テンプ
レートを加えた.測定はViiA7 を用いて行い,データ解析は ViiA7 ソフトウェア
を用いた検量線法により行った.平均と S.D.は 3 頭のカニクイザルから算出し
た.
Table 7. Primers of the nuclear receptor for real-time PCR analysis.
Target genes Primer sequences (5'→3') AHR
Forward primer AAAATGAAGCATATGCAAGTTAATGG Reverse primer TCTTGCCGTGGACAACTGAA
CAR
Forward primer GCAAGTCATCAAGTTTACCAAGGA Reverse primer TGAGAAGGGAGATCTGGTCTTCA
PXR
Forward primer ACGCTCAGATGAAAACCTTTGAC Reverse primer GGCATCTCACAGCCACTGCTA
42 3.3 結果 3.3.1 肝細胞の調製及び培養 2 章で使用した 6 頭のカニクイザルに加え,追加で 6 頭のカニクイザルから肝 細胞を調製した.調製直後及び解凍後の細胞生存率はTable 8 に示した.これら の肝細胞の生存率はいずれも先の6 頭と同等または高い値を示した. 新鮮及び凍結肝細胞のいずれも,生細胞は培養3 日目までに効率よく接着し, コンフルエントに達した.凍結肝細胞については,浮遊細胞もみられたが,培 養 7 日目以降,新鮮肝細胞と比較して明確な違いはみとめられなかった.細胞 の凝集と小型肝細胞の増殖は,単層培養及び共培養法にて確認された.3D スフ ェロイド培養では,顕微鏡下では肝細胞,小型肝細胞,フィーダー細胞の見分 けはできなかった.
Table 8. Characteristics of the hepatocytes from cynomolgus monkey.
Application Monkey ID
Age
(years) Sex
Viability (%)
After isolation After thawing Fresh hepatocytes F4 4 male 97.1 - F5 3 female 91.2 - F6 5 male 92.9 - Cryopreserved hepatocytes C4 4 male 93.4 79.2 C5 4 male 92.5 73.9 C6 4 male 93.0 86.6 3.3.2 カニクイザル肝細胞に典型的な酵素誘導剤を曝露した際のシトクロム P450 mRNA 発現における影響
オメプラゾールは核内レセプターAHR を活性化させ,CYP1A mRNA 発現レベ
ルを制御することが知られている50).CYP1A1 mRNA レベルの誘導率は,培養
43 は,培養4 日目から 7 日目の誘導率は低かったが,培養 11 日目から 14 日目に はわずかに回復した.一方で共培養においては,新鮮肝細胞の培養4 日目から 7 日目と11 日目から 14 日目で 20 倍以上,凍結肝細胞の 11 日目から 14 日目と 25 日目から28 日目で,30 倍以上の誘導率がみとめられた.3D スフェロイド培養 においては,培養期間中で誘導率は4~12 倍の間で一定であった.
Figure 8. Effects of omeprazole on CYP1A1 mRNA levels in the hepatocytes of cynomolgus monkey.
The mRNA expression level of CYP1A1 after exposure to omeprazole was determined by RT-qPCR. Data were normalized to those of HPRT (reference gene) and the induction rate relative to the control is presented as mean ± S.D. (n = 3) for freshly isolated hepatocytes (A) and cryopreserved hepatocytes (B). Statistical analysis was performed using Student’s t-tests: *p < 0.05 vs. DMSO.
リファンピシンは核内レセプターPXR を活性化させ,主に CYP3A mRNA 発
44 胞のいずれも,CYP3A8 mRNA 発現レベルは 3 種の培養方法において増加がみ とめられた(Fig. 9).誘導率は,単層培養の新鮮肝細胞における培養 11 日目か ら 14 日目と,凍結肝細胞のいずれの培養日数において 10 倍以上を示した.共 培養及び3D スフェロイド培養はいずれも 4~8 倍程度で,似かよった誘導率を 示した.
Figure 9. Effects of rifampicin on CYP3A8 mRNA levels in the hepatocytes of cynomolgus monkey.
The mRNA expression level of CYP3A8 after exposure to rifampicin was determined by RT-qPCR. Data were normalized to those of HPRT (reference gene) and the induction rate relative to the control is presented as mean ± S.D. (n = 3) for freshly isolated hepatocytes (A) and cryopreserved hepatocytes (B). Statistical analysis was performed using Student’s t-tests: *p < 0.05 and **p < 0.01 vs. DMSO.
フェノバルビタールは核内レセプターCAR を活性化させ,主に CYP2B mRNA
45 イザル新鮮及び凍結肝細胞のいずれの培養条件においても弱い増加を示した (Fig. 10).新鮮肝細胞では媒体コントロールと比較して 2 倍以内の誘導率であ ったが,凍結肝細胞では単層培養の培養 11 日目から 14 日目を除き,全てのポ イントで2 倍以上の増加を示し,3D スフェロイド培養の培養 11 日目から 14 日 目では4.6 倍であった.加えてフェノバルビタールはヒトにおいて CYP2C 及び CYP3A を誘導することが知られている52).カニクイザル新鮮及び凍結肝細胞に
おいては,CYP2C43 及び CYP2C75 mRNA レベルの弱い誘導,及び CYP3A8
mRNA レベルの誘導がみとめられた(Figs. 11-13).CYP2C43 では単層培養にお
いて新鮮及び凍結肝細胞のいずれも,培養 4 日目から 7 日目と 11 日目から 14 日目で10 倍以上の誘導率を示した.CYP2C75 は凍結肝細胞の方が若干高い誘導 率を示したが,全体で2.4~8.6 倍であった.CYP3A8 については,誘導率は新鮮 肝細胞よりも凍結肝細胞でより明確で,3D スフェロイド培養の 25 日目から 28 日目を除き,10~23 倍の誘導率を示した.新鮮肝細胞では,単層培養の 2 日目 から3 日目のみで高い誘導率を示し(18 倍),他の培養期間では 2~7 倍であっ た.
46
Figure 10. Effects of phenobarbital on CYP2B6 mRNA levels in the hepatocytes of cynomolgus monkey.
The mRNA expression level of CYP2B6 after exposure to phenobarbital was determined by RT-qPCR. Data were normalized to those of HPRT (reference gene) and the induction rate relative to the control is presented as mean ± S.D. (n = 3) for freshly isolated hepatocytes (A) and cryopreserved hepatocytes (B). Statistical analysis was performed using Student’s t-tests: *p < 0.05 vs. DMSO.
47
Figure 11. Effects of phenobarbital on CYP2C43 mRNA levels in the hepatocytes of cynomolgus monkey.
The mRNA expression level of CYP2C43 after exposure to phenobarbital was determined by RT-qPCR. Data were normalized to those of HPRT (reference gene) and the induction rate relative to the control are presented as mean ± S.D. (n = 3) for freshly isolated hepatocytes (A) and cryopreserved hepatocytes (B). Statistical analysis was performed using Student’s t-tests: *p < 0.05 vs. DMSO.
48
Figure 12. Effects of phenobarbital on CYP2C75 mRNA levels in the hepatocytes of cynomolgus monkey.
The mRNA expression level of CYP2C75 after exposure to phenobarbital was determined by RT-qPCR. Data were normalized to those of HPRT (reference gene) and the induction rate relative to the control is presented as mean ± S.D. (n = 3) for freshly isolated hepatocytes (A) and cryopreserved hepatocytes (B). Statistical analysis was performed using Student’s t-tests: *p < 0.05 and **p < 0.01 vs. DMSO.
49
Figure 13. Effects of phenobarbital on CYP3A8 mRNA levels in the hepatocytes of cynomolgus monkey.
The mRNA expression level of CYP3A8 after exposure to phenobarbital was determined by RT-qPCR. Data were normalized to those of HPRT (reference gene) and the induction rate relative to the control is presented as mean ± S.D. (n = 3) for freshly isolated hepatocytes (A) and cryopreserved hepatocytes (B). Statistical analysis was performed using Student’s t-tests: *p < 0.05 vs. DMSO.
3.3.3 カニクイザル肝細胞の核内レセプターAHR,CAR,PXR の mRNA 発現 レベル
前項で述べたように,オメプラゾール,リファンピシン,フェノバルビター
ルは主にAHR,PXR,CAR をそれぞれ活性化させる.本実験では,これら核内
レセプターのmRNA 発現レベルについて検証した.AHR については,mRNA 発
現レベルはオメプラゾール曝露によりわずかに減少する傾向がみられた(Fig.
14).PXR については,mRNA 発現レベルはリファンピシン曝露により,単層培
50 PXR mRNA 発現レベルが減少する培養期間があった.(Fig. 15).特に新鮮肝細 胞の 3D スフェロイド培養の 25 日目から 28 日目で顕著な減少がみられたが, CYP3A8 の誘導に影響は見られなかった.PXR はフェノバルビタールによって も活性化されることが知られており,フェノバルビタール曝露による PXR mRNA 発現レベルについても検証した.結果として,PXR mRNA レベルはばら ついたものの,フェノバルビタール曝露によるPXR mRNA レベルの減少はみら
れなかった(Fig. 16).CAR mRNA レベルはフェノバルビタール曝露によりいず
れの培養条件でも媒体コントロールと比較して減少がみられ,特に培養期間の
後半においてその減少は顕著であった(Fig. 17).
Figure 14. Effects of omeprazole on AHR mRNA levels in the hepatocytes of cynomolgus monkey.
The mRNA expression level of AHR after exposure to omeprazole was determined by RT-qPCR. Data were normalized to those of HPRT (reference gene) and the expression relative to the control are presented as mean ± S.D. (n = 3) for freshly isolated hepatocytes (A) and cryopreserved hepatocytes (B).
51
Figure15. Effects of rifampicin on PXR mRNA levels in the hepatocytes of cynomolgus monkey.
The mRNA expression level of PXR after exposure to rifampicin was determined by RT-qPCR. Data were normalized to those of HPRT (reference gene) and the expression relative to the control are presented as mean ± S.D. (n = 3) for freshly isolated hepatocytes (A) and cryopreserved hepatocytes (B). Statistical analysis was performed using Student’s t-tests: *p < 0.05 vs. DMSO.
52
Figure 16. Effects of phenobarbital on PXR mRNA levels in the hepatocytes of cynomolgus monkey.
The mRNA expression level of PXR after exposure to phenobarbital was determined by RT-qPCR. Data were normalized to those of HPRT (reference gene) and the expression relative to the control are presented as mean ± S.D. (n = 3) for freshly isolated hepatocytes (A) and cryopreserved hepatocytes (B).
53
Figure 17. Effects of phenobarbital on CAR mRNA levels in the hepatocytes of cynomolgus monkey.
The mRNA expression level of CAR after exposure to phenobarbital was determined by RT-qPCR. Data were normalized to those of HPRT (reference gene) and the expression relative to the control are presented as mean ± S.D. (n = 3) for freshly isolated hepatocytes (A) and cryopreserved hepatocytes (B). Statistical analysis was performed using Student’s t-tests: *p < 0.05 vs. DMSO.
3.4 考察 本実験では,カニクイザル 6 個体の新鮮肝細胞及び 6 個体の凍結肝細胞を用 いて,典型的な酵素誘導剤によるCYP mRNA 発現レベルの誘導能について比較 した.それぞれ 3 個体の肝細胞はオメプラゾールとリファンピシンの曝露に用 い,別のそれぞれ 3 個体はフェノバルビタールの曝露に使用した.前者の各 3 個体の肝細胞は,第二章の実験において 3 種類の培養条件で 14 日間または 28 日間培養した時のmRNA 発現プロファイルの検証に使用している49).前実験で