有用乳酸菌の分離と水産養殖への応用に関する基礎的研究
平成18年度
三重大学大学院 生物資源学研究科 博士前期課程
生物圏生命科学専攻 海洋微生物学研究分野
家畠 俊平
目次
緒言
1章 環境中からの乳酸菌の分離・同定および魚病原因菌V)'bm'o属に対する抗菌活性測定 によるスクリーニング
1.1目的 1.2方法
1.2.1新規乳酸菌の分離および簡易同定 1.2.2魚病原因菌V)'bn'o属抗菌活性測定試験
1.2.3 16S
rRNA塩基配列相同性および系統解析を用いた種同定1.3結果
1.3.1新規乳酸菌の分離および簡易同定 1.3.2魚病原困菌V)'brlo属抗菌活性測定試験
1.3.3 16S
rRNA塩基配列相同性および系統解析をもちいた種同定1.4考察
2章 新規乳酸菌分離株の実用化に向けた評価試験 2.1目的
2.2方法
2.2.1人工胃腸液耐性試験 2.2.2塩濃度に対する評価試験 2.2.3熱に対する評価試験 2.3結果
2.3.1工胃腸液耐性試験 2.3.2塩濃度に対する評価試験
緒言
世界的魚資源の需要増大による水産養殖の増加が推測され、それに伴い魚病 の激増が懸念されている。これまで魚病対策として、主に抗生物質を使用して いるが、これらに対する耐性菌の出現や魚体内への残留性などの問題点が指摘 されている。このためワクチンや非病原性のプロパイオテイクスの利用は抗生
物質の利用低減と耐性菌発生の回避につながると考えられる(ト3)。プロパイオ テイクスとはFullerら(4)により「腸内フローラの改善により宿主動物に有益 な効果をもたらす生菌漆加物」と定義されてきた。その後この定義は改変され、
現在では「宿主に生理学的に有益な効果をもたらす生菌および死菌とそれらの 代謝産物からなる微生物調剤」の事をいう。そして、さらに「経口投与でき、
粘膜表面に付着し粘膜表面の微生物または酵素活性の改善を行い、特異的また は非特異的に免疫機能の促進させることを目的とするもの」と定義されている
(5).現在プロパイオテイクスとして研究されている微生物は主に Vl'bn'o 属,PseudomoDaS属,Bacillus属,乳酸菌( Lactobac)'llus, StzlePtOCOCCuS,
Camobacten'um)
,酵母,微細藻がある(1,5,6,7).
乳酸菌は分類学的に定義すると、グラム陽性、運動性無、胞子無形性、従属 栄養性の梓菌または球菌で炭水化物、糖、脂肪酸、アミノ酸、ペプチド、ビタ
ミンなどの栄養要求性をもっている。また、カタラーゼ産生反応陰性(または シュウドカタラーゼ反応陽性)の通性嫌気性菌で、ブドウ糖を発酵しその50%
以上を乳酸に転換する細菌のことを言い、動物の腸内や海産物、乳製品、植物
表面など自然界に広く存荏している(8,9)。特に発酵分野において風味、香り付 け、病原菌や腐敗菌に対して抗菌効果を持つといった理由から食品の製造や保
存に用いられ、古くから人間と深く関わっている微生物である(10)。乳酸菌の
もつ主な抗菌作用として乳酸や酢酸の生成によるpHの低下である。さらに
H202やCO2、 Reuterin、 Diacetylなどの低分子化合物(LMM)やバクテリオ シンのような高分子化合物(HMM)'がある(10‑17).
このような抗菌作用から乳酸菌は有用なプロパイオテイクスとして利用され ている。しかし、乳酸菌の消化器宮内での役割についての研究は人や家畜など では広くされている一方で魚での役割に関する研究は少なく、また免疫機能調 節に関しても同様のことが言える(18,19)0 invivoで病原菌に感染した魚へプ
ロパイオテイクスを投与している研究は多くないが、 Gildberg ら(20)は
Camobacten'um d)'vergeDSがVl'bn'o aDgu)'11ammのin vitroでの成長阻害は見 られなかったが、 V)'brlo aDgul'11ammによるAtlantic cod(Gadus morhua)稚魚 の死亡率を減少させたと幸艮告した。
Villamilら(2)はLactococcus lact)'sの死菌がヒラメのHKマクロファージを 増加させ、 Lactococcuslact)'sの死菌、生菌ともに血清中の一酸化窒素の値を上 昇させたと報告した。
以上の背景から本研究では抗生物質の代替品として分離した乳酸菌を飼料中 に漆加することで健康で安全な魚類の養殖を行うことが目的である。
1章 環境中からの乳酸菌の分離・同定および魚病原困菌T%m'o属に対する 抗菌活性測定によるスクリーニング
1.1目的
魚病はグラム陽性、グラム陰性菌の双方に存在するが種類数、被害の大きさ といった点でもグラム陰性菌の重要度は高く、現在魚病対策として主に抗生物 質を用いているが、耐性菌の発生や魚体内への残存性などの問題がある。その 代替品としてプロパイオテイクスやワクチンの投与などが有望であり、本研究 ではプロパイオテイクスの1つである乳酸菌の水産養殖への利用を目的とした。
本章では環境中から乳酸菌(LacticAcid Bacteria LAB)の分離を行い、グラム 陰性菌であり、ニジマスやウナギなどのビブリオ症原因菌Ⅵbrioanguillarum、腹 部膨満症原因菌Ⅵあrio harveyiなどのV)'bn'o属に対する抗菌活性能試験による菌 株のスクリーニングを行った。
そして抗菌活性の強かった株について16S rRNA塩基配列相同性および系統解 析を用いた種の同定を行った。
1.2方法
1.2.1乳酸菌の分離および簡易同定
CaC13がpH低下により白色から透明になる性質を利用して、家畜糞・乳酸菌 製品および環境から乳酸菌の分離を行った。
試料原液および希釈液1.Omlを0.1% CaCO3加DeMan, Rogosa, and Sharpe (MRS)培地に混釈法で接種した。 30oCで2‑3日間培養後、クリアゾーン形成し たコロニーを異なった形毎にランダムに釣菌し、同液体培地に接種後、再度MRS 寒天培地に画線した。そしてそれを数回繰り返し乳酸菌の単離を行った。
単離した菌株を形態観察、グラム染色、運動性試験、カタラーゼ試験、硝酸 還元試験、リトマス試験に基づく簡易性状試験を行った。
分離株の培養はMRS培地を基本培地として30oCで1‑2日間静置培養した。
また、単離した菌株の保存は15%グリセロ‑ル加MRS液体培地で‑30oCで凍 結保存した。
1.2.2.魚病原困菌V)'bm'o属抗菌活性測定試験 1.2.2.1乳酸菌およびV)'btT'0属の培養.
1.2.1で分離した乳酸菌をMRS液体培地、 30oC、 1‑2日間静置培養した。 Vl'bn'o 属の培養にはZobel12216 (Zobell)寒天平板培地(人工海水11、ポリペプトン5
g、酵母エキス1g pH7.6‑7.8)を用いた。また、培養条件として魚類消化器宮 内が微好気条件であることを考慮し、スクリューキャップ試験管を用いて25oC で2‑3日間静置で行った。
1.2.2.2乳酸菌上清液および魚病原困菌の細胞懸濁液の調整
Vl'bplo属の培養液を10000rpm、 10min、 4oCで遠心分離後、ペレットを滅菌済 み生理食塩水(0.9%NaCl)で2匝I洗浄し、 OD600=0.1となるように生理食塩水 に懸濁したものを細胞懸濁液とした。
乳酸菌上清液0.7ml (コントロールにはZobell液体培地0.7ml)、 Vl'brlo属細胞 懸濁液0.1mlを入れ、 25oC、 2日間静置培養を行った。その後分光光度計を用 いOD600で魚病原困菌の生育阻害率の測定を行った。
1.2.3
16S rDNA塩基配列相同性および系統解析を用いた種同定1.2.3.1培養液からのTotal DNAの抽出
乳酸菌培養液を遠心分離(8000rpm,30min,4oC)し、デカンテ‑ションにより上 清を除去した。残ったペレットをTE溶液1mlで懸濁し、遠心分離(15000叩m,
5min,
4oC)で洗浄を行った。その後フェノール・クロロホルム法でtotal DNAを抽出した。
抽出したDNAを100plに溶解後lO/oアガロ‑スゲル電気泳動(AGE)を行い、
約21kbp付近に単一バンドの出現を確認した(Fig.1.A)0
1.2.3.2
PCRによる16Sr
DNAの増幅およびサイクルシークエンスによる種同定
バンドの確認後抽出した DNA をテンプレートにしてプライマー
16S・8F (5'AGAGTTTGATCCTGGCTCAG3') 、 16S・ 1492R(5'GGCTACCTTGTTACGAC TT3')を用いて1 6SrDNA遺伝子(1500bp)の増幅を行った。 PCRサイクリ
ングはサーマルサイクラー(TaKaRaPCRThermalCyclerMP)を用いて、反
応液を94oC, 1分保った後に、変性94oC, 1分、アニーリングを56oC, 30秒、
伸長72oC, 2分の反応を25サイクル行い、その後予備伸長に72oC, 2分行っ た。
増幅させたDNAを1%アガロ‑スゲル電気泳動(AGE)を行い約1.6kbpの単一 バンドの出現を確認した(Fig. 1.B)0
16S
rDNA領域の増幅の確認後、Big‑Dye terminater &t
v.3.1(AppliedBiosystems) を用いたサイクルシークェンスによるラベリングを行った。プライマーは16S,8F, 16S1356F(5'ACTCCTACGGGAGGCAGCA3') , 16S・704F(5'GTAGCGGTGAAATGCGTA GA3'), 16S・1114F(5'GCAACGAGCGCAACCC3'), 16S・338R(5'CTGCTGCCTCCCGTAG GAGT3り, 16S・517R(5'GTAGTTACCGCGGCTGCTGGC3り, 16S・1100R(5'AGGGTTGCG CTCGTTG3り, 16S・1492Rを用いて行った。
その後ABI PRISM 3100‑Avant Genetic Analyzer (ABI)を用いて塩基配列を 決定した。
1.2.3.3
1后SrDNA塩基配列に基づく系統解析シークエンサにより得られた塩基配列を遺伝子解析ソフトchromasで編集し、
16S rDNA遺伝子の塩基配列を決定した。
その配列を元にNational Center for Biotechnology lnformation(NCBI)のホ ームページ(bttp://www.ncbi.nlm.nib.gov/)のBLASTで塩基配列相同性検索
を行い、近縁株の塩基配列を取得した。その後マルチプルアライメントソフト clustalXを使いマルチプルアライメントを行い、両端を揃えて余分な塩基を削
除した。そしてclustalXによるブートストラップ検定を行い、 NJplotにより系 統樹を作成した。
1.3結果
1.3.1乳酸菌の分離および簡易同定
CaCO3加MRS培地での混釈法による分離の結果、 10サンプルから33株の 乳酸菌を分離した。そして、全ての分離株がグラム陽性、運動性なし、カタラ
ーゼ陰性、酸性化陽性、硝酸還元能陰性だった。
1.3.2.魚病原因菌V)'bElo属抗菌活性測定試験
分離株33株の魚病( V aDgu111amm, V Aan(eyl', V)'bn'o.
sp.
17A, V)'bn'osp.
18C)に対する抗菌活性試験を行った。その結果、 33株中13株が特に強い抗菌 活性を示したのでpH6.8調整前後で比較を行い、その結果をFig. 2.に示す。
分離株上清のpH未調整のものでは分離13株全てが魚病原困菌5種の増殖を
阻害した.また、上清をpH6.8に調整したものでは全てのV aDgu)'naz・umに 対してb15が約55%と最も高い阻害率を示し、それ以外の12株は約20‑40%
の阻害率を示した. Vhaz・veyl'に対してs6, s7が約80%と最も高い阻害率を示 し、それ以外の11株は約60‑70%と高い阻害率を示した. V)'brjo 17Aに対し
てc5, z4が約90%と最も高い阻害率を示し、それ以外の11株は40‑80%の阻害 率を示した. Vl'bn'o
sp.
18Cに対してc5が90%と最も高くそれ以外の12株は 35‑85%の阻害率を示した。1.3.3
16S rDNA塩基配列相同性および系統解析をもちいた種同定16S rDNA塩基配列相同性および系統解析を行った結果をTable
l、
Fig. 3.に 示す.塩基配列相同性結果からa3がLactobacillus paLlaPlaDtaZ・um ・C8,
c9が Lb. plaDtamm ・ b2, b4,z4,
z5がLb.paracasel'
・ b6, c5がLactobacilluscasel'
・ b15がLb. (ermeDtum ・ 02がLeucoDOStOCmeSeDteL101'des ・s6,
s7がEDterOCOCCuS mZmdtli'、 UDCultuzled bacten'umと99%以上の相同性を示した.
系統解析結果から、 a3がLactobacl'nus pamplaDtamm ・
C8,
c9がLb.plaDtamm ・ b2, b4,
c5, z4,
z5がLb.pamcase)'・ b6がLactobacillussp.
・ b15 がLb. fermeDtum ・02がLeucoDOStOC meSeDteZ101'des・s6,
s7がEnteL10COCCuSmuDdtli'、 UDCultwedbacten'zLmと同クラスターを形成した.
またa3は米ぬか由来、b2,b4,b6,b15は乳酸菌水由来、c5,
c8,
c9は廃水由来、o2は牛糞由来、 s6,s7は馬糞由来、 z4,z5はEM菌由来であった。
1.4考察
乳酸菌はグラム陽性菌門、バチルス綱、ラクトバチルス目に属する細菌で、
食品や動物の消化器宮内のほかに土壌や廃水などの環境中にも生息している。
プロパイオテイクスとして用いられている乳酸菌には Camobacten'um属、
StTePtOCOCCuS属、 EDteL10COCCuS属、 LeucoDOStOC属、 Lactobacl'11us属がある.
本研究で分離した株はEDteZ10COCCuS属、 LeucoDOStOC属、 Lactobacillus属と近 縁関係を示していた。このことから本研究で分離した13株はプロパイオテイク
スとして利用できる可能性があることを示している。
多くの魚類は消化器官や免疫機能が未発達な幼生の状態で生まれてくるため、
体表やえらからだけでなく飲む海水からも微生物が侵入し、蓄積していく。こ のことは常在菌の形成と病原菌感染の原因の1つと考えられ、継続した養殖を
行うためには病原菌の増殖の抑制が必要であると考えられる(21)。本研究では、
分離した乳酸菌の培養液上清を用いて魚病原困菌に対する抗菌活性試験を行っ た。その結果、 pH未調整の培養液上清では魚病原困菌に対し、高い抗菌活性を
示した.一方pHを中性に調整した培養液上清の抗革活性結果では、 V
aDgu)'11ammに対する阻害率が他の3種に対する阻害率40‑90%であるのに対 し20‑55%と低かった。この事からVaDgul'11arumに対する主な抗菌活性効果
はpH低下によるものであると考えられる.一方 V harvey)', Vl'bn'o
sp.
17AV)'bn'osp. 18Cの3株に対する主な抗菌活性効果はpH低下によるものだけでは
ないことが明らかになった。
乳酸菌の抗菌活性には、乳酸生成によるpH低下や栄養塩の競合以外にも過酸
化水素やバクテリオシンなどの抗菌性物質を生成するものが多く(9,12,14)、こ れらの結果から、過酸化水素やバクテリオシンなどの抗菌性物質の生成による
ものであることが示唆された。
これをうけて次章では更なる評価試験を行い、プロパイオテイクスとしての 利用に有効な可能性の株のスクリーニングを行った。
(A)
M 1 2 3 4 6 6 7 8 9 10 11 12 13
t]い
M 1 2 3 4 5 6 7 8 9 10 11 12 13
Fig. 1.
(A)Agarose Gel
Electrophore8is仏GE)of extracted totalI)NA
(B)
Agarose gel Electrophoresis of 16S rRNA gene by PCR Lane
M:入作IindⅢ ・EcoR I double digest Markez・
Lane 1:a3 Lane2:b2 Lane3:b4 Lane4:b6 Lane5:b15
Lane6:c5 Lane7:c8 Lane8:c9 Lane9:o2 LanelO:s6
Lane ll:s7 Lane 12:z4 Lane 13:z5
o
0.8
:司 く○
【⊃
O o.6
cent a3 b2
⊂】帽
b4 b6 b1 5 (;5
四Iめ
c8 c9
□17A
o2 s6 $7 z4 z5
■18C
Table 1. Results of the BLAST Homology oftbe sequences of 16S rRNA gene
isolates CIosest relative
homology(㌔)LaotobacI'uus parap/antarum AJ 306 29 7 LaotobacI'uus par;aCaSe/'subsp. Par;acase/' LactobacJ'uus case/'ATC C334
Lactobacmus casei ATCC334
Lactobac/'uus fe′mentum strain SFCB216c Laotobao/'uus caseI'ATCC334
Laotobacmus p/antarlum AB 104855 LactobacI'uus p/antanJm AB 1 04855 Leuoonostoo 177eSente/りI'des ATCC 8293 Enter10COCCuS mund/I'ABO6 626 6 Enter10COCCuS mundI'I'ABO6 6266 LaotobacI'uus paracaseI'subsp. ParacaseI' LactobacI'uus paracase/'subsp. Paraoase/'
99 99 99 一oo
99
99
99
99
99
99
99
99
99
(.7・t751l
L
(1l.
lltP.t・PII(mlt'(1(.1・..ATK:C 82鍋Lb・fmllPllhlltt・
SFCE)2‑&a)5
Ai'b'ppt(1(
(1
( (1LS.
〟l'iAolltilll'1Jl
lI・†
2章 新規分離株の実用化に向けた評価試験
2.1目的
養殖魚へのプロパイオテイクス利用には消化器宮内への到達および生存性が 重要であり、消化器官内での生存性には消化器宮内の細胞への付着能と胃腸内 での胃液pHおよび胆汁酸に対する耐性がある。また人工飼料作成の評価試験と して、海水魚への投与を考慮したNaClへの耐性と養殖飼料形成時の加熱処理へ の耐性が必要である。このため本章では分離株の人工胃腸液および耐塩・耐熱 性試験を行った。
2.2方法
2.2.1人工胃腸液耐性試験 2.2.1.1人工胆汁塩耐性試験
Walker
andGillilandら(22)の方法に従って実験を行った。 OD600=0.8‑1.0
となるまで培養した分離株100plを試験区としてpH
8.0,
0.3% oxgall入りMRS液体培地(コントロールに0.3% oxgallの入っていないMRS液体培地を 用いた) 10mlに接種し、 30oCで12時間培養を行い、 3時間ごとにOD600で吸 光度測定し試験区とコントロールでの胆汁酸耐性の比較を行った。そして6時 間および12時間培養時での阻害率を下式で求めた。
阻害率‑ (1‑ABS/AcT)
×100
(%)BS: Bile Salt CT
:
control2.2.1.2胃腸液耐性試験
Rodrigo(23)らの方法に従った。 1.5mlエツベンドルフチューブにpH
1.5‑5.0
と調整した人工胃液900plずつ入れ、 OD600=0.8‑1.0となるまで培養した分離株を100pl接種し、ウオーターバスで37oC,3時間培養を行った。そして100pl
を各希釈濃度に希釈し、 MRS平板培地に接種し、 2日間30oCで培養し、その後 コロニー計数した。
なお人工胃液の代わりにMRS液体培地を入れたものをコントロールとして、阻 害率を下式で求めた。
阻害率‑ (1‑logcfuAGJ/logcfu cT) ×100 (0/.)
AGJ
:
Arti丘cial Gastric Juice CT:
Control2.2.2
NaClに対する評価試験2.2.2.1.細胞懸濁液の調整
分離株培養液を10000rpm、 10min、 4oCで遠心後沈殿を生理食塩水(0.9%
NaCl)で2匝】洗浄し、 OD600=0.1となるように生理食塩水に懸濁した.
2.2.2.2.透過光濁度法による耐塩性試験
NaCl濃度(0, 513,
1280, 1710,
2140mM)を調製した1%グルコース加MRS 液体培地に細胞懸濁液100plを接種し、 (この時間を0時間とする) 3日間24 時間毎に分光光度計を用いて、吸光度(OD600)を測定した。2.2.3.2.コロニーカウント法による耐熱性試験
1.5mlエツペンチュ‑ブに1mlずつ細胞懸濁液を分注し,ヒートブロック上 で各温度(50,60,70,80,90,95oC)、 10分間熱ショックを与えた。
その後、滅菌済みミリQ水で希釈し、 1%グルコース加MRS平板培地に100pl ずつ塗抹した。 30oCでト2日間静置培養を行い、形成したコロニー数の計数を 行った。
なお熱ショックを行わなかったものをコントロールとし、コロニー数の減少で 耐熱性の有無を検討した。
2.3結果
2.3.1人工胃腸液耐性試験
胆汁塩の分離株への阻害率をTable 2.に示す。6時間培養での阻害率はb15,
z4
株を除く11株で低かった。また12時間培養ではz4株を除く12株で阻害を受
けなかった。特にa3株は6時間、 12時間培養の双方で0.3% bile saltsの有無 に関係なく高い増殖能を示した。
人工胃液の分離株への阻害率をTable 3.に示す。 13菌株全てでpH2.0‑5.0ま で高い増殖能を示したが、 b6,
s6,
s7株を除く10株がpHl.5で増殖不可能になった。またpHl.0では全ての株が増殖不可能になった。
2.3.2
NaClに対する評価試験72時間培養でのNaClに対する評価試験結果をFig. 4.に示す。 13株全てで NaC13%漆加MRS培地中において0%と比較しても増殖に差は見られなかった。
またa3,
c9,
o2の3株が7.5%漆加でも高い増殖を示した。しかし、 10%以上の 添加では増殖は見られなかった。2.3.3熱に対する評価試験
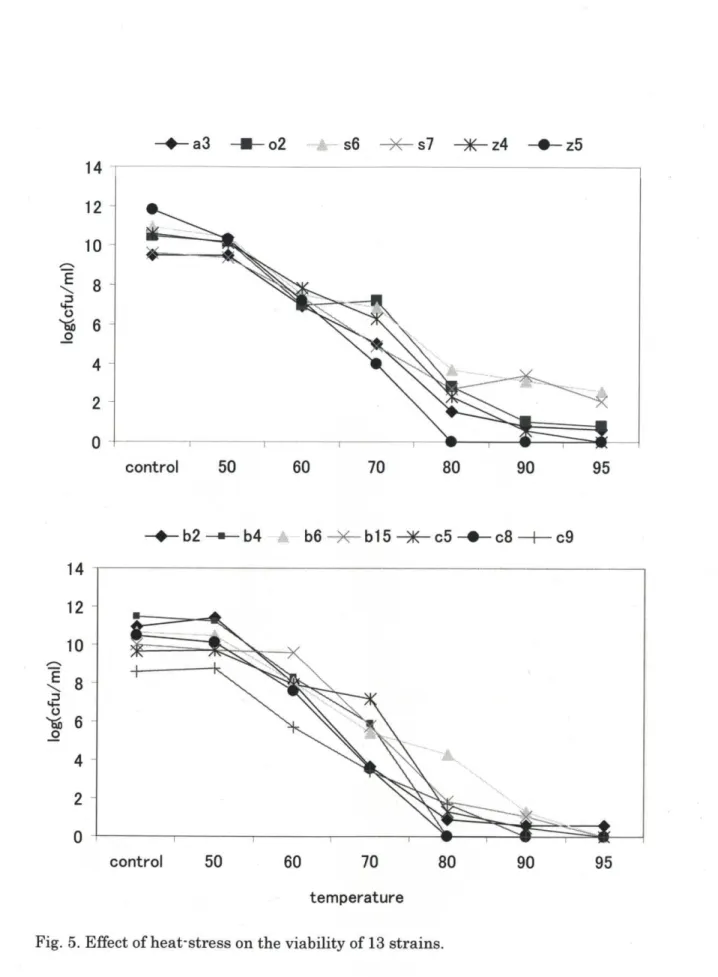
熱に対する評価試験結果をFig. 5.に示す。 50oCでは13株全てが高い生残数
2.4考察
魚類の消化器官は大きく分けて食道、胃、腸にわけられる。多くの魚類では 食道部には粘液分泌腺が存荏しているため、ただ単に餌を円滑に飲み込むため の器官ではなく、特にコイなどの無胃魚では食道のアミラーゼなどの酵素も消 化に役立っているといわれている。
胃内のpHは2.0‑4.0で、胃で最も強く作用する消化酵素であるペプシンの至適 pH2.0‑4.0とほぼ一致することが知られている。そして胃液pHの変動として 摂餌後胃液や塩酸の分泌が盛んになり、胃液pHは急激に変化し摂餌直後には pH3.0以下となる。しかし、その後急速にアルカリに傾いた後元の酸性値に戻 り、これは消化速度に関係していることがわかっている。腸内腔のpH値は中性 から弱アルカリを示し、胃におけるpH同様に摂餌後変動するが範囲は狭い。し かし、腸内にはトリプシンやキモトリプシンなどの様々な消化酵素が存在し、
摂餌後それらの活性が増加することが報告されている(24)0
現在の水産養殖分野においてプロパイオテイクスは飼料への漆加または飼料 水への添加の経口投与により魚体内へ摂取される。そのためプロパイオテイク スは上に記したような厳しい環境で生存しなければならない。 Huangら(25)は pH2.0、pH3.0での生存性はプロパイオテイクスの選択する際の判定基準にでき
ると報告している。またNeumannら(26)は死菌が宿主の免疫機能を促進するの で十分量を経口投与した時、プロパイオテイクスの生存性は必要条件ではない と報告している。一方でRodrigoら(23)は生物的療法が栄養塩の直接的な競 合や抗菌物質の分泌などの代謝活性に依存しており、効果的な経口ワクチンに
は細菌の生存性が重要であると報告している。これらのことからプロパイオテ イクスの魚類への利用には消化器宮内への進入時の胃液pHや胆汁塩といった 局所的なストレスへの耐性が必要不可欠である。そのため本研究では、作成し
た人工胃腸液を用いて分離した13株に対する増殖阻害率を試験した。人工胃液 実験においてpH2.0‑5.0まですべての株が増殖阻害されなかったが、 pHl.5で はb6,
s6,
s7株のみが耐性を示し、pHl.0では13株全てが増殖を示さなかった。そして人工腸液実験ではb15, z4株を除く11株が胆汁塩の有無に関係なく高い 増殖を示した。またプロパイオテイクスは上記の厳しい環境を通過した後消化 器宮内で一定期間生存し宿主に有益な作用をもたらす必要があるので、消化器 官内細胞に付着しコロニー形成をする必要性がある。この事からプロパイオテ イクスには胃液pHや胆汁に対する耐性のほかに消化器宮内での付着能も必要 不可欠であると考えられ、今後消化器宮内への付着能実験を行う必要が考えら れる。
現在水産養殖の大半は主に海上で行われており、プロパイオテイクスが飼料 中で生きたまま魚体内に吸収されるためにはNaClに対する耐性が必要と考え られ、このような理由から現在研究されているプロパイオテイクスの多くは海 洋環境由来のものがほとんどである。 Raoら(27)はLD.plaDtamm541および A6をNaCl濃度0‑30%添加MRS培地で24時間培養後OD600を用いて測定し
たところ2株とも5%で高い増殖能を示し、また10%においても増殖可能であ
る事を報告した。本研究において、分離した13株全てがNaCl濃度3.0 %でも 高い増殖能を示した。特にa3,c9,o2,の3株が7.5%添加でも高い増殖を示した。
いる。またDong・Fang(29)らはwhite sturgeonの幼生に加水分解したデンプン またはグルコースを含む飼料を加熱処理したものを与える事で摂餌率を増加さ せたと報告している。またDong・Fangらはグルコースを含む飼料の高温長時間 加熱が飼料内のリジンなどのアミノ酸とメイラード反応による栄養価の損失を 引き起こし魚の生育を減少させるとも報告している。これらのことから飼料の 加熱をする事で消化率や摂餌率を向上させると考えられ、プロパイオテイクス を飼料に漆加する際の耐熱性が必要であると考えられた。そのため本研究では
70oC、 10分加熱処理までは全ての株が高い生残率を示したが、 80oCではb2,b4,
c8,z5の4株が検出限界以下となった。また本研究の加熱時間10分に対して塩 崎(30)らは分離した乳酸菌が80oC、60分でも高い生残率を示した事を報告した。
これらのことから今後加熱温度を70oCまたは80oCに固定し、処理時間の延長実 験を試みる必要があることが考えられた。
本研究で分離した13株について1章の抗菌活性測定結果と本章の結果及び Huangらの報告するプロパイオテイクス利用にはpH2.0での生存性が必要であ
るということから判断するとa3,b6,c9,s6の4株が有効ではないかと思われた。
しかし、摂餌直後に胃液pHが2.0以下になる魚種も存在し、必ずしもpH2.0 での生存性がプロパイオテイクス利用に万全であるとは言い切れない。そのた め、本研究ではpHl.5で生存性を示した分離株b6,s6の2株をプロパイオテイ
クス利用に有効であると考え選定した。今後は培養液上清中のどの抗菌性物質 が魚病原困菌の増殖抑制効果を示すのかについての更なる研究が必要であると 考えられる。
Table 2. Growth inhibition of 13 strains by bile salts
Isofates Bi一e salts inhibition
(㌔)aBi一e salts inhibition
(%)ba3 b2 b4 b6 b15
¢5 c8 c9 o2 s6
$7 z4 z5
0 27.4
14.2 45.6 99.3 30.7 12.9 9.59 15.I 38.2 38.0 58.0 40.2
0 20.4 21.7 39.8 5.05 45.0 4.27 I.22 34.9 8.33 0.00 64.3 43.6
Percentage of optical density reduction a洗er 6 h
(a)and 12 h
(b)growing in MRS with
Table 3. Growth inhibition of 13 strains by simulated gastric
juiceswith different pH
isolates Gastric
juicesinhibition
(%)pH 5.0 pH 4.0 pH 3.0 pH 2.5 pH 2.0 pH 1.5 pH 1.0
0 6.67 5.52 4.43
3.77 2.97 4.21 1 3.3
0 0
0 0
1.29 7.36
0 0.53
0 0
0 0
5.06 4.46
1 3.3 1 5.0
2.88 3.36
0 0
0 0
3.46 25.0
0 10.9
10.4 16.I
2.37 0
0 0
1.63 1.36
ll.0 18.9
20.4 20.6
6.46 8.43
0 5.74
0 7.01
4.84 1 00
18.2 100
6.03 1 00
I 6.4 72.9
19.5 100
6.35 100
6.32 100
2.64 100
24.9 100
30.2 69.7
8.51 59.3
19.2 100
12.2 100
i
8コ
l●‑
く)
、滋 6 .望
controJ 50 60 70 80 90 95
‑●‑b2
‑I‑‑b4ゼ′こ btトー×‑‑‑b15うlトc5 ‑●‑c8+c9
!8
.I? O
漕6
contro1 50 80 70 80 90 95
temperature
Fig. 5. Effect of heat‑stress 0ntheviability of 13 Strains.
3章.総括
現在魚病対策として抗生物質を用いているが、抗生物質の利用は耐性菌の発 生や魚体内への残存などの懸念がある。その代替品として乳酸菌などのプロパ イオテイクスやワクチン投与がある。そこで分離した乳酸菌を飼料中に漆加す ることで健康で安全な魚類の養殖を行うことを目的とした。プロパイオテイク ス利用には以下の3つの条件1)魚病原困菌に対する抗菌活性能2)消化器宮内 での生存性(胃液pH、胆汁塩に対する耐性) 3)消化器宮内への付着能が必要 であると考えられる。これらの条件を考慮して以下の実験を行った。
本研究では環境中から乳酸菌を分離し、条件1, 2と養殖魚飼料作成時の評価 試験として耐塩性および耐熱性実験を行った。魚病原因菌のうちV)'bn'o属細菌 4株( VaDgul'llamm, VharveyI, V)'b)'osp. 17A, V)'bn'osp. 18C)に対する培養 上清液を用いた抗菌活性試験において、分離した33株中13株が抗菌活性能を
示した。これら抗菌活性を示した13株についてpHを中性に調整した培養液上 清で同様の実験を行った。その結果、 V aDgul'11ammに対する阻害率が若干低 下したものの他の3株(Vhanleyl, V)'bl'o
sp.
17A, Vl'bn'osp. 18C)に対する阻害 率は維持された。また人工胃腸液を作成して耐性試験を行った結果、 b15,z4株 を除く11株で胆汁塩に対する耐性を示し、胃液pHが2.0では13株全てが阻それら2株はそれぞれ乳酸菌水、馬糞を分離源とした。それら2株の塩基配列
相同性及び系統解析結果からb6株がLactobacinus sp.と、s6株がEDtemCOCCuS muDdtl')'と近縁関係があることが明らかになった。
謝辞
本論文を作成するにあたり多くのご指導をいただきました三重大学 前田広 人教授、三重大学 田中礼二助教授に深く感謝の意を表します。本研究で使用
した魚病微生物を提供してくださいました鹿児島大学水産学部山本淳教授、宮 崎大学田岡洋介氏に深く感謝の意を表します。また多くの助言を頂き、惜しみ ないご協力を下さいました海洋微生物学研究室の中野みよ氏ならびに当研究室 の皆様に深く御礼申し上げます。最後に6年間もの長きにわたり私を支えてく れた両親に感謝の意を表します。
参考文献
1. Gram, L., a. Melchiorsen, B. Spanggaard, Ⅰ. Huber, and T. F. Nie18en. 1999.
Inhibition of V)'bn'o aDgu)'llamm by PseudomoDaS nUOreSCeDS AH2, a Possible Probiotic Treatment of Fish. Appl. Environ. Microbiol. 65: 969‑973.
2. Villamil, L., C. Tafalla, A.Figuera8, and B. Novoa. 2002. Evaluation of lmmunomodulatory Effects of Lactic Acid Bacteria in Turbot
(Scophthalmusmaxl'mus).
Clin. Diag. Lab. Immunol. 9: 1318・1323.
3. Niko8kelainen, S., S. Salminen, G. Bylund, and A. C. Ouwehand. 2001.
Characterization of the Properties of Human‑ and Dairy・Derived Probiotics for Prevention of Infectious Diseases in Fish. Appl. Environ. Microbiol. 67: 2430・2435.
4. Verschuere, L., G. Rombaut, P. Sorgeloo8, and W. Ver8traete. 2000. Probiotic Bacteria as Biological Control Agents in Aquaculture. Microbiol. Mol. Biol. Rev. 64:
655・671.
5. Jansen, G. a.
,and D. V. D.
Waaji.1995. Prospects of use in opportunistic infections:review of the internal discussion. In probiotics: Prospects of use on opportunistic infections.
(Fuller.R.
ed.),Institute for microbiology and biochemistry,Old Herbon University, Herbon Dill. 173・ 184.
6. Maurilio, L. F., A. 0. N. Miguel, E. G. M. Beatriz, and L. M. Wilberth. 2003. Use of the bacteria StzlePtOCOCCuS (aeclum and Lactobacillus ac)'doph)'1us, and the yeast Saccharomyces ceLlelr7'sIae as growth promoters in Nile tilapia
(OTeOChrom)'s D)Plot)'cus
).Aquaculture. 216.: 1931201.
7.Joborm, a., a. C. OIsson, A. We8terdahl, P. L. Conway, and S.
Kjelleberg.1997.
Colonization in the fish intestinal tract and production of inhibitory substances in intestinal mucus and faecal extracts by Carnobacterium sp. strain Kl. a. Fish. Disea.
20: 383・392
8. Ringo, lL, and F. a. Gatesoupe. 1998. Lactic acid bacteria in fish: a review Aquaculture. 160: 177・203.
9.内村泰、岡田早苗1992.乳酸菌実験マニュアル朝倉書店1・19
10. Parente, E., and A. Ricciardi. 1999. Production, recovery and purification of bacteriocins from lactic acid bacteria. AppI MicrobioI Biotechnol. 52: 628・638.
ll. Ammor, S., G. Tauveron, E. Dufour, and I. Chevallier. 2006. Antibacterial activity of lactic acid bacteria agalnSt spoilage and pathogenic bacteria isolated from the same meat small・scale facility 1‑Screening and characterization of the antibacterial compounds. Food Control. 17: 454・461
12. Schimnger, U., and F. K Lucke. 1989. Antibacterial Activity of Lactobacl'llus sake
Isolated from Meat.Appl. Environ. Microbiol. 55:1901‑ 1906.
13. Song, Y. L., N. Kato, Y. Mat8umiya, C. X. Liu, H. Kato, and K. Watanabe. 1999.
Identi丘ca
tion of and Hydrogen Peroxide Production by Fecal and Vaginal Lactobacilli lsolated from Japanese Women and Newborn Infants. J. Clin. Microbiol. 37: 3062・3064.
14. Yap, P. S., and S. E. Gilliland. 2000. Comparison of Newly Isolated Strains of La ctobacl'nus delbyuechli'・
subsp. 1actl's for Hydrogen Peroxide Production at 5oC. J. Dairy Sci. 83: 628・632.
15. Mart1'n, R., M. 0livare8., M. L. Man'n., a. Xau8., L. Ferna'ndez., and a. M.
Rodn′guez. 2005. Characterization of a reuterin・producing Lactobac111us
coryn)'(orm)'s strain isolatedfrom a goat's milk cheese. Int. a. Food. Microbiol. 104:
267・277.
16. Rodn′guez, E., a. L.Anque′s., R. Rodn′guez., M. Nun‑ e2:., and M. Medina. 2003.
Reuterin production by lactobacilli isolated from pig faeces and evaluation of probiotic traits. Lett. in Appl. Microbiol. 37: 259・263.
17. James, M. a. 1982. Antimicrobial Properties of Diacetylt. Appl. Environ. Microbiol.
44: 525‑532.
18. Isolauri, E., P.
V.Kirjavainen,and S. Salminen. 2002. Probiotics: a role in the treatment of intestinal
infection and inflammation? Gut. 50: 54・59.
19. Gill, H. S. 1998 Stimulation of the Immune System by Lactic Culture. Int. Dairy Journal. 535・544.
20. Gildberg, A., H. Mikkelsen, E. Sandaker, and E. Ringo. 1997 Probiotic effect of lactic acid bacteria in the feed on growth and survival offry ofAtlantic cod
(Gadusmoz:hua).
Hydrobiologia. 352: 279・285.
21.田岡洋介2003プロパイオテイクスのヒラメとその飼育環境に関する効果 鹿児島 大学修士学位論文
tolerance of potential probiotic dairy proplOnibacteria. Int. J. Food Microbiol. 91:
253・260.
26. Neumann, E., M. A. P.
0liveirよ.,c. M. Cabral., L. N. Moura., J. R. Nicoli., E. C.
Vieira., D. C. Cara., G. Ⅰ.Podoprigora., and L.
Q.Vieira. 1998. Monoassociation with Lactobacillus ac)'doph)'1us UFV・H2b20 stimulates the immune defense mechanisms ofgermfree mice. Braz J Med BioI Res. 31: 1565・1573.
27. Rao, M. S., a. Pintado., W. F. Steven8., and a. P. Guyot. 2004.Kinetic growth parameters of different amylolytic and non・amylolytic Lactobac)'llus strains under various salt and pH conditions. Bioresource Technology. 94: 331・337.
28. Pere8a, H., C. Limb., and P. H. mesius. 2003. Nutritional value ofheat・treated soybean meal for channel catfish
(ZctalumspuDCtatuS).Aquaculture. 225: 67・82.
29. Deng, D. F., G. Ⅰ.Hemre, T. Storebakkenc, S. Y. Shiaud,and S. S. 0. Hung. 2005.
Utilization of diets with hydrolyzed potato starch, or glucose by
juvenilewhite
sturgeon
uc)beDSer tZ7aDSmODtaDuS),aS affected by Maillard reaction during feed processing. AqllaCulture. 248: 103・ 109.
30塩崎靖志前田広人田岡洋介越塩俊介吉川毅坂田泰造2006水産養殖における新規 プロパイオテイクス候補菌のスクリーニングBokinBobai.