The author declares no con‰ict of interest.
神戸学院大学薬学部臨床薬学研究室(〒6508586 神戸 市中央区港島一丁目13)
e-mail: harada@pharm.kobegakuin.ac.jp
本総説は,平成25年度日本薬学会近畿支部奨励賞(生 物系薬学)の受賞を記念して記述したものである.
―Review―
脳虚血性耐糖能異常に起因する神経障害発現に対する orexin-A の中枢末梢臓器間連関機構を 介した役割
原 田 慎 一
Role of Orexin-A-mediated Communication System between Brain and Peripheral Tissues on the Development of Post-ischemic Glucose Intolerance-induced Neuronal Damage
Shinichi Harada
Department of Clinical Pharmacy, School of Pharmaceutucal Sciences, Kobe Gakuin University; 113 Minatojima, Chuo-ku, Kobe 6508586, Japan.
(Received May 30, 2014)
I recently found that cerebral ischemic stressper secauses hyperglycemia(i.e., post-ischemic glucose intolerance) and suppression of post-ischemic glucose intolerance might be important to improve prognosis. Here, I analyzed the e‹cacy of suppression of post-ischemic glucose intolerance using orexin-A(OXA)endogenous neuropeptide as a novel therapeutic strategy against cerebral ischemic neuronal damage. OXA in hypothalamus plays a role in many physiologi- cal functions including regulation of glucose metabolism. I previously found that the development of post-ischemic glu- cose intolerance is suppressed by OXA. Other reports have shown that the communication system between brain and peripheral tissues through the autonomic nervous system is important for maintaining glucose and energy metabolism.
The aim of this study was to determine the involvement of the hepatic vagus nerve on hypothalamic OXA-mediated sup- pression of post-ischemic glucose intolerance and neuronal damage. Intrahypothalamic administration of OXA sig- niˆcantly suppressed the development of post-ischemic glucose intolerance on day 1 and of neuronal damage on day 3 after middle cerebral artery occlusion(MCAO). In the liver, MCAO-induced decrease in insulin receptors and increase in gluconeogenic enzymes on day 1 was recovered to control levels by OXA; these eŠects were reversed by hepatic vagotomy. In the medulla oblongata, OXA induced co-localization of the cholinergic neuronal marker choline acetyltransferase with orexin-1 receptor and c-Fos. These results suggest that the vagus nerve projecting from the medul- la oblongata plays an important role in the recovery of post-ischemic glucose intolerance and mediates neuroprotection by hypothalamic OXA.
Key words―cerebral ischemia; vagus nerve; glucose intolerance; insulin sensitivity
1. はじめに
脳梗塞を始めとする脳血管疾患は,悪性腫瘍・心 疾患とともに主要な死亡要因の1つである.脳卒中 に対する治療薬に関して,動物実験レベルでの基礎 報告は多いものの,臨床試験においては望ましい治 療成績が得られないため,脳卒中に対する治療薬の 開発は,困難な状況である.13)
以前から,脳卒中の危険因子として「高血糖」は 重要視されてきた.すなわち,糖尿病の既往歴のあ
る患者では脳卒中の発症リスクが23倍に増加する とされている.4)発症リスクだけでなく,高血糖状 態では,生じる神経障害が著しく増大することが明 らかとされ,脳卒中発症の予防において,血糖値制 御の重要性は以前より唱えられてきた.その機序と しては,高血糖状態の動物に虚血ストレスを負荷す ることにより血管性の炎症,血液脳関門の透過性の 促進,細胞膜の障害並びに組織のアシドーシスが促 進され,脳虚血による神経障害が悪化するという知 見などが得られている.5,6)さらに最近では,糖尿病 の既往歴のない場合においても,脳卒中発症後に高 血糖を呈し,それをinsulinにより厳格に制御する ことにより死亡率が抑制されるという報告がなされ ている.7)これに関し,筆者は,動物モデルを用い た研究により,脳虚血ストレス負荷時において,肝
1056
原田慎一
神戸学院大学薬学部 レクチャラー(講 師),博士(薬学).2010年神戸学院大 学大学院食品薬品総合科学研究科博士 後期課程修了.20102011年日本学術 振興会特別研究員(DC2, PD).2012 年より現職.研究テーマ:脳卒中にお ける臓器間連関の変容の解明と新規治 療戦略開発のための基盤研究,中枢性 疼痛の発症機序の解明及び新規治療法 の開発,健康食品・サプリメントから の新規脳保護薬の探索.
1056 Vol. 134 (2014)
臓におけるinsulin抵抗性を介した一過性の耐糖能 異常が発症し,この血糖値上昇が,学習・記憶障害 などの予後の悪化に関与することを明らかとしてき た.8,9)
近年,糖代謝制御は,肝臓を含む個々の末梢臓器 における制御のみではなく,視床下部を介した中枢 末梢臓器間連関による神経支配を受けることが注目 されている.10,11)中枢末梢臓器間連関は,最近にな って明らかにされた概念であり,10,11)末梢臓器から のエネルギー代謝情報に対する中枢での受容並びに 応答様式については,いまだ不明な点が多い.実際 に,末梢で産生されるレプチンやインクレチンなど いくつかの生理活性物質が中枢領域へ(特に,視床 下部)へシグナルを伝達し,それが全身の代謝調節 に重要な役割を果たすことはよく知られている.12) さらに,こうした中枢神経系から末梢臓器への神経 入力,あるいは末梢臓器から中枢神経系への神経入 力は各種臓器の機能調節だけでなく,中枢神経の機 能調節に影響を及ぼしていると考えられる.加え て,最新の創薬研究においては,安全性及び治療後 の高いQOLを可能とする治療戦略の開発を目的 に,内因性生理活性ペプチドを用いたトランスレー ショナルリサーチと画期的な治療法の確立が注目さ れている.
そこで筆者は,脳虚血ストレス負荷後の血糖値変 化を制御できる内因性生理活性ペプチドを探索する ことを目的として,近年中枢末梢臓器間連関によっ て,視床下部を介して末梢組織での糖代謝を制御す ることが報告された神経ペプチドorexin-Aの関与 に着目した.13)
本総説では,筆者が最近明らかにした一過性脳虚 血ストレス負荷による耐糖能異常の発現と神経障害 との関係について紹介するとともに,orexin-Aに よる脳保護作用について,中枢末梢臓器間連関を介 した血糖値制御という観点から概説する.
2. 脳虚血性耐糖能異常及び神経障害発現に対す るOrexin-Aの役割
Orexin-Aは,1998年に同定されたエネルギー代
謝調節関連神経ペプチドであり,摂食中枢である視 床下部外側野に散在する神経細胞に存在する.14)視 床下部外側野で産生されたorexin-Aは,中枢にお いてnorepinephrine, serotonin, dopamineなどの起 始核に興奮性の入力を送り,食欲や報酬系に係わる
ほか,睡眠や覚醒を制御すること,さらにはエネル ギー 恒 常性 の 維持 に関 与 する こと が 知ら れて い
る.15,16)加えて,脳虚血ストレス負荷により,梗塞
部位におけるorexin-1受容体の発現上昇,並びに パーキンソン病やナルコレプシーなどの神経変性疾 患に罹患している患者の脳脊髄内で,orexin-Aの 量が減少していることが報告されている.1719)さら に,orexin-Aは脳室内投与によって,脳血流量の 改善を介した脳保護作用を持つことも報告されてい る.20)以上のことから,orexin-Aは中枢神経活動を 適正に制御する統合的な機能を有する可能性が高い.
近年,このorexin-Aは視床下部腹内側核に作用 することにより交感神経及び骨格筋のb2受容体の 活性化によりinsulinの感受性を亢進させるとの報 告がなされた.13)さらに,orexin欠損マウスにおい て,insulin抵抗性や耐糖能異常が加齢に伴い生じ ることも知られている.21)これらの報告からも,糖 代謝制御は,個々の臓器だけではなく,視床下部を 介した中枢末梢臓器間連関が関与し,orexin-Aは それに関与する因子の1つであると考えられる.
筆者はこれまでに,局所脳虚血モデルである中大 脳 動 脈 閉 塞 モ デ ル (middle cerebral artery occlu- sion; MCAO,梗塞時間:2 h)を用いて,MCAO 1日後の空腹時血糖値が増加すること,及び経口糖 負荷試験における糖負荷後血糖値の回復が遅延する ことを見い出した.8)さらにinsulin投与後の血糖値 の経時的な変化を観察したところ,偽手術群では,
insulinの処置により速やかに血糖値が低下するの
に対し,MCAO群では血糖値の低下が遅延した.
すわなち,MCAOによってinsulin抵抗性(耐糖能 異常)が発現した.8)また,その発現と連動して,
肝臓におけるinsulin受容体の活性化の低下と,糖 新生関連酵素の発現上昇が生じることを明らかにし てきた.9)また,脳虚血ストレス負荷により,肝臓
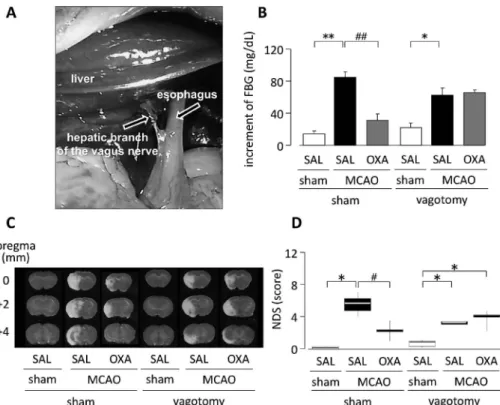
Fig. 1. EŠect of Hepatic Branch Vagotomy on Hypothalamic Orexin-A-mediated Suppression of Elevated FBG Levels and Deve- loped Neuronal Damage on Day 1 and 3, Respectively after MCAO
Intrahypothalamic administration of orexin-A immediately following MCAO on day 7 after hepatic branch vagotomy.(A)Operative ˆeld for selective hepatic nerve of the vagus nerve interruption.(B)FBG levels on day 1 after MCAO.p<0.01,##p<0.01,p<0.05.(C)Representative photomicrographs of TTC stain- ing on day 3 after MCAO.(D)Results of the neurological deˆcit score(NDS)on day 3 after MCAO.p<0.05,#p<0.05. MCAO/saline-treated group:n=6, other group:n=5. SAL: saline, OXA: orexin-A(modiˆed from Ref. 25).
内insulin受容体量自身が減少していたことから,
これがinsulin受容体活性化の減少の引き金になっ
たと考えられる.9)脳虚血ストレス負荷後早期にお いて,orexin-A産生部位である視床下部外側野に
おけるorexin-Aの発現量が偽手術群に比較して有
意に減少していた.22)一方で,orexin-Aの受容体で
あるorexin-1受容体の発現にはなんら影響してい
なかった.この条件下において,orexin-Aを脳室 内又は視床下部内に局所投与することによって,
MCAO 1日 後 の 空 腹 時 血 糖 値 の 上 昇 , 並 び に
MCAO 3日後の神経障害発現(梗塞巣形成,行動
障害並びに学習・記憶障害)は,偽手術群に比較し て有意に抑制された.22)同時に,MCAO 1日後の 肝臓におけるinsulin受容体並びにリン酸化insulin 受 容 体 の 減 少 , 糖 新 生 関 連 酵 素 で あ る phos- phoenolpyruvate carboxykinase(PEPCK) 並 び に glucose-6-phosphatase(G6Pase)の発現上昇のいず れにおいても偽手術群レベルへの有意な回復が認め られた.22)すなわち,視床下部におけるorexin-A は末梢組織におけるinsulin感受性の改善に重要な
役割を担っていると考えられる.
3. Orexin-Aによる視床下部延髄迷走神経系 を介した脳虚血性耐糖能異常に起因する神経障害の 抑制作用
Orexin-Aの受容体は,脳全体に広く存在し,特
に視床下部においては,腹内側核,室傍核,弓状核 及び外側野において確認されている.23,24)これら各 部位は,肝臓や骨格筋の機能制御を担う交感神経並 びに副交感神経の起始核でもある.18,25)さらに,視 床下部の腹内側核や外側野神経の活性化が,交感神 経や副交感神経を介して,肝臓や骨格筋における
insulin受容体基質の活性化や糖産生の制御を担っ
ていることも知られている.13)すなわち,視床下部
のorexin-Aの作用発現に自律神経系が関与してい
る可能性が考えられた.そこで筆者は,脳虚血スト レス負荷後の血糖値変化に対するorexin-Aの視床 下部を介した制御機序を解明するために,肝機能を 制御する神経の1つである迷走神経に着目し,その 起始核が存在する延髄を介した視床下部における
orexin-Aの脳虚血後の糖代謝異常及び神経障害発
1058
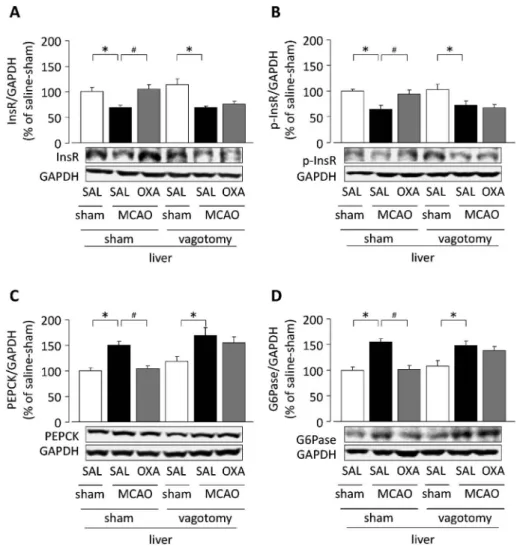
Fig. 2. EŠect of Hepatic Branch Vagotomy on Hypothalamic Orexin-A-mediated EŠects on Hepatic Expression of Insulin Receptor (InsR), Phosphorylated(p)-InsR, PEPCK and G6Pase on Day 1 after MCAO
Intrahypothalamic administration of orexin-A immediately following MCAO at 7 d after hepatic branch vagotomy. Representative western immunoblots and quantiˆcation(ratio to GAPDH)as%of sham-SAL of hepatic: InsR(A), p-InsR(B), PEPCK(C), G6Pase(D).p<0.05,#p<0.05.n=12. SAL: saline, OXA:
orexin-A(modiˆed from Ref. 25).
1058 Vol. 134 (2014)
現への関与について検討を行った.
まず,orexin-Aによる脳虚血誘導性耐糖能異常 発現の抑制機序における延髄orexin-1受容体の関 与 に つ い て 検 討 し た と こ ろ ,orexin-A に よ る
MCAO 3日後の梗塞巣形成,行動障害,学習・記
憶障害発現及びMCAO 1日後のFBGの上昇の抑 制は,延髄へのSB334867(orexin-1受容体のアン タゴニスト)処置によって有意に拮抗された.26)同 時に,orexin-Aによる肝臓でのinsulin,リン酸化 insulin 受 容 体 の 発 現 減 少 並 び に 糖 新 生 関 連 酵 素
(PEPCK, G6Pase)の発現上昇の抑制も,延髄への
SB334867処置によって有意に拮抗された.26)また
orexin-Aは,迷走神経の起始核である延髄弧束核
及び迷走神経背側核において,神経活性マーカーで あるc-Fosの発現を上昇させた.26)さらに,このc-
Fosはorexin-1受容体又は迷走神経のマーカーであ るcholine acetyltransferase との共局在,及び同部 位 に お い てorexin-1 受 容 体 と choline acetyltrans- feraseとの共局在が認められた.26)すなわち,視床
下部のorexin-Aは,延髄から投射している迷走神
経上に発現しているorexin-1受容体を介して,末 梢組織である肝臓になんらかのシグナルを伝達し,
それが肝臓におけるinsulinシグナル系の賦活化を 促進している可能性が考えられた.さらに,肝臓枝 迷走神経を特異的に切除することによって[Fig. 1 (A)],上述したMCAO後の空腹時血糖値の増加
[Fig. 1(B)],梗塞巣形成[Fig. 1(C)],行動障害
[Fig. 1(D)],学習・記憶障害の発現及び肝臓にお けるinsulin,リン酸化insulin受容体[Figs. 2(A) and(B)]の発現減少並びに糖新生関連酵素[Figs.
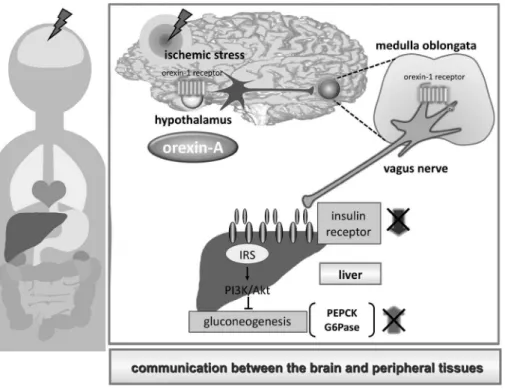
Fig. 3. Schematic Drawing of Activation of Hepatic Insulin Signaling Mediated Hypothalamic Orexin-A through Activation of Hypothalamus-Medulla Oblongata-Vagus Nerve Axis
It is possible that hypothalamas orexin-A activates hypothalamus-medulla oblongata-vagus nerve axis, resulting regulates for hepatic glucose metabolism.
2(C) and(D)]の発現上昇に対する視床下部の
orexin-Aの作用が,消失した.26)これらの結果から,
orexin-Aによる中枢領域(視床下部)から末梢組
織へのシグナル伝達に延髄から投射している迷走神 経が関与している可能性が示唆された(Fig. 3).
以上の知見から,orexin-Aによる肝臓のinsulin シグナルの賦活化を介した脳虚血性耐糖能異常及び 神経障害の発現の抑制作用には,視床下部延髄迷 走神経間の活性化が関与する可能性が示唆された.
このように,中枢領域における内因性生理活性ペプ チドの活性調節や全身性の耐糖能異常の改善が脳梗 塞に対する新たな治療戦略の1つとして有用である 可能性が本研究により示唆された.
4. おわりに
本研究は,これまでその実行分子としては重要視 されてこなかった「糖」に着目し,その作用点を解 明する点に大きな特色がある.本研究結果は,上位 中枢における内因性生理活性ペプチドの活性調節 が,脳卒中治療に対して有用であることを示唆して いると同時に,脳虚血ストレス負荷後の全身性の耐 糖能亢進による脳保護という新たな概念を脳卒中治 療に導入するものと考えられる.一方で,血糖値そ のものが脳梗塞の予後の直接的な原因か否かについ
ては,糖尿病患者において,多臓器の動脈硬化の進 行,あるいは高血糖が重症脳卒中で多くみられるこ となどから,依然として議論すべき重要な課題であ る.しかしながら,その発現機序や治療効果に対す る科学的知見をエビデンスに基づいて考察するため には,本研究のような研究を遂行することが,その 第一段階として必要であると考えられる.今後は,
さらに詳細な機序解明に取り組み,動物における基 礎研究の成果をヒトにトランスレートできるよう に,更なるエビデンスの構築を行う必要がある.
謝辞 本研究は神戸学院大学薬学部臨床薬学研 究室において行われたものであり,研究遂行に際 し,本研究の機会を与えて頂き,終始懇切な御指導 と御鞭撻を賜りました神戸学院大学薬学部 徳山尚 吾教授に深甚たる謝意を表します.さらに,本研究 遂行に際し,御助言を賜りました神戸学院大学薬学 部 中本賀寿夫講師に心から御礼申し上げます.ま た,肝臓枝迷走神経切除モデルの作製方法を御教授 頂きましたアスビオファーマ株 幸田修一博士に深 く感謝致します.末筆ながら,本研究を遂行するに あたり多大な御協力を頂きました研究室の諸氏に心 より感謝の意を表します.
1060
1060 Vol. 134 (2014)
REFERENCES
1) Shuaib A., Lees K. R., Lyden P., Grotta J., Davalos A., Davis S. M., Diener H. C., Ash- wood T., Wasiewski W. W., Emeribe U.,N.
Engl. J. Med.,357, 562571 (2007).
2) Watanabe T., Tanaka M., Watanabe K., Takamatsu Y., Tobe A., Yakugaku Zasshi, 124, 99111(2004).
3) Yamaguchi T., Mori E., Minematsu K., Nakagawara J., Hashi K., Saito I., Shinohara Y.,Stroke,37, 18101815(2006).
4) Doi Y., Ninomiya T., Hata J., Fukuhara M., Yonemoto K., Iwase M., Iida M., Kiyohara Y.,Stroke,41, 203209(2010).
5) Widmer H., Abiko H., Faden A. I., James T.
L., Weinstein P. R., J. Cereb. Blood Flow Metab.,12, 456468(1992).
6) Dietrich W. D., Alonso O., Busto R.,Stroke, 24, 111116(1993).
7) Gentile N. T., Seftchick M. W., Huynh T., Kruus L. K., Gaughan J., Acad. Emerg.
Med.,13, 174180(2006).
8) Harada S., Fujita W. H., Shichi K., Tokuya- ma S.,Brain Res.,1279, 174181(2009).
9) Harada S., Fujita-Hamabe W., Tokuyama S., J. Pharmacol. Sci.,115, 155163(2011).
10) Yamada T., Katagiri H.,Endocr. J.,54, 497
505(2007).
11) Imai J., Katagiri H., Yamada T., Ishigaki Y., Suzuki T., Kudo H., Uno K., Hasegawa Y., Gao J., Kaneko K., Ishihara H., Niijima A., Nakazato M., Asano T., Minokoshi Y., Oka Y.,Science,322, 12501254(2008).
12) Friedman J. M., Halaas J. L., Nature, 395, 763770(1998).
13) Shiuchi T., Haque M. S., Okamoto S., Inoue T., Kageyama H., Lee S., Toda C., Suzuki A., Bachman E. S., Kim Y. B., Sakurai T., Yanagisawa M., Shioda S., Imoto K., Minokoshi Y., Cell Metab., 10, 466480 (2009).
14) Sakurai T., Amemiya A., Ishii M., Matsuzaki I., Chemelli R. M., Tanaka H., Williams S.
C., Richardson J. A., Kozlowski G. P., Wil- son S., Arch J. R., Buckingham R. E., Haynes A. C., Carr S. A., Annan R. S., McNulty D.
E., Liu W. S., Terrett J. A., Elshourbagy N.
A., Bergsma D. J., Yanagisawa M.,Cell,92, 573585 (1998).
15) Hagan J. J., Leslie R. A., Patel S., Evans M.
L., Wattam T. A., Holmes S., Benham C. D., Taylor S. G., Routledge C., Hemmati P., Munton R. P., Ashmeade T. E., Shah A. S., Hatcher J. P., Hatcher P. D., Jones D. N., Smith M. I., Piper D. C., Hunter A. J., Porter R. A., Upton N.,Proc. Natl. Acad. Sci. USA, 96, 1091110916(1999).
16) Yamanaka A., Tsujino N., Funahashi H., Honda K., Guan J. L., Wang Q. P., Tomina- ga M., Goto K., Shioda S., Sakurai T., Biochem. Biophys. Res. Commun.,290, 1237
1245(2002).
17) Mihara Y., Dohi K., Yofu S., Nakamachi T., Ohtaki H., Shioda S., Aruga T.,J. Mol. Neu- rosci.,43, 162168(2011).
18) Ripley B., Overeem S., Fujiki N., Nevsimalo- va S., Uchino M., Yesavage J., Di Monte D., Dohi K., Melberg A., Lammers G. J., Nishida Y., Roelandse F. W., Hungs M., Mignot E., Nishino S.,Neurology,57, 22532258(2001).
19) Nakamachi T., Endo S., Ohtaki H., Yin L., Kenji D., Kudo Y., Funahashi H., Matsuda K., Shioda S., Regul. Pept., 126, 4954 (2005).
20) Kitamura E., Hamada J., Kanazawa N., Yonekura J., Masuda R., Sakai F., Mochizuki H., Neurosci. Res.,68, 154157(2010).
21) Tsuneki H., Murata S., Anzawa Y., Soeda Y., Tokai E., Wada T., Kimura I., Yanagisawa M., Sakurai T., Sasaoka T.,Diabetologia,51, 657667 (2008).
22) Harada S., Yamazaki Y., Tokuyama S., J.
Pharmacol. Exp. Ther.,344, 276285(2013).
23) Trivedi P., Yu H., MacNeil D. J., Van der Ploeg L. H., Guan X. M., FEBS Lett., 438, 7175(1998).
24) Marcus J. N., Aschkenasi C. J., Lee C. E., Chemelli R. M., Saper C. B., Yanagisawa M., Elmquist J. K., J. Comp. Neurol., 435, 625 (2001).
25) Shimazu T., Ogasawara S., Am. J. Physiol., 228, 17871793(1975).
26) Harada S., Yamazaki Y., Koda S., Tokuyama S., PLoS One,9, e95433(2014).