博 士 学 位 論 文
エビ養殖場におけるウイルス・細菌の感染防御
に関する研究
近畿大学大学院
農学研究科水産学専攻
赤澤典昭
博 士 学 位 論 文
エビ養殖場におけるウイルス・細菌の感染防御
に関する研究
平成 30 年 3 月
近畿大学大学院
農学研究科水産学専攻
赤澤典昭
目 次
第1章 序論・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・ 1 第2章 エビ養殖場におけるウイルスの複合感染・・・・・・・・・・・・・・・ 5
第3章 エビ養殖場におけるウイルスに対するバイオセキュリティー・・・・・・14 第4章
Penaeus vannamei
養殖における Puddle Wheel の微細藻類に対する影響・・・・・・・・・・・・・・・・・・・・・・・25 第5章
Penaeus vannamei
養殖におけるEMS/AHPND 発症原因の考察・・・・35第6章 養殖池のSludge と pH の上昇がもたらす EMS/ AHPND の発症
(投稿論文)・・・・・・・・・・・・・・・・・・・・・・・・・・・・53 第7章 総合考察・・・・・・・・・・・・・・・・・・・・・・・・・・・・・62 要旨・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・66 Summary ・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・70 参考文献・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・74 謝辞
- 1 -
第1章 序論
2016 年中国・Guangzhou で行われた GOAL ミーティングの James L.Anderson によ
るプレゼンテーションは、FAO 等のデータをもとにしたものであるが、2016 年の世界の養 殖エビ生産量は、4,000,000 (Mt)に達するとしている。そのうち 1,500,000(Mt)が東南ア ジアで、さらに1,400,000 (Mt)が中国での生産としている。つまり、世界の養殖エビ生産 の73%がアジアで生産されている。さらに、東南アジアでは、38%を生産していることにな る。また、2005 年(2,700,000 Mt)から 2011 年(4,000,000 Mt)にかけての生産量の増加は 特に顕著であり、ブラックタイガーからバナメイへのシフトのインパクトがいかに大きか ったかを示している。一方2011 年から 2015 年にかけての落ち込みも大きく、EMS/AHPND
(Early Mortality Syndrome/ Acute Hepatopancreatic Necrosis Disease) による被害の大 きさを物語っている。 東南アジアの生産量の拡大は、各国の発展とともにインフラの整備が進んできたことも 大きな要因であるとともに、エビ養殖が熱帯~亜熱帯という気候適応もあり、主要な外貨獲 得産業となっている。一方、エビ養殖の急激な拡大は、マングローブ帯をはじめとする環境 破壊を想起させ、負のイメージに繋がっている側面もある。 筆者は1999 年以来、マレーシア・マレー半島東海岸のパハン州にてエビ養殖に携わって きた。その間に経験したエビの病気、現在においても未だに甚大な被害をもたらしている WSSV (White Spot Virus)をはじめとするウイルス病、2008 年頃から東南アジアに中国か
ら拡大した EMS/AHPND の原因と特定された Vibrio parahaemolyticus などの細菌病、
Sritunyalucksana et al.,(2015)による NACA の緊急報告でも取り上げられている、EHP
(Enterocytozoon hepatopenaei) などの寄生虫症など甚大な被害とも対峙してきた。
筆者は、主として大型の養殖場のマネジメントを通して上述の変化を経験してきた。 Agrobest 社(Pahang 州)と BlueArchipelago 社(BAB 社/iKerpan 養殖場・Kedah 州)で
あるが、それぞれ450 池・200 池を有する規模のマレーシアでは有数の養殖場である。
これらの養殖場を運営するにあたって、持続可能なビジネスモデルにするためには何が必 要なのかを常に考えてきた。先述の負のイメージに対していかにそれを変えられるか、また 養殖場立地地域との共存についても大きな課題として取り組んできた。そんな中で、安定し た養殖事業の運営には、いかに病気の侵入を防ぎ、拡大を防ぐかというウイルス・細菌に対
- 2 - するバイオセキュリティーの問題について考えることが欠かせないと理解してきた。 本研究は、特に大型の養殖場を持続可能にするために、何が必要なのかを養殖場での実践 を通して研究してきたものである。 第2章では、ブラックタイガー養殖におけるウイルスの複合感染に関する研究を行った。 2000 年代中盤まで、ブラックタイガーの養殖は天然の親エビに依存していた。その天然の 親エビ資源が複合的なウイルス罹患によってブラックタイガー養殖を困難なものとしてい った。本章では、複合感染がいかに成長を阻害しているかをエビの成長に沿って段階的に PCR を使ってウイルスの有無を検査しながら追跡した。 第3章では、養殖場におけるバイオセキュリティーに関して考察した。本章は、養殖場に おけるウイルスキャリアーの検査を通して養殖場のウイルス感染の評価が可能であること を示した。これは、ウイルスの痕跡を探ることによって、ウイルスの侵入を予測することに もつながるものとして考察した。また、養殖場の履歴において、ウイルスキャリアー、宿主 の存在が簡単に排除できるものではないことも示唆するものであった。養殖場の管理にお いて、ウイルス感染の防御とともに感染の拡大をいかに防ぐかの指針にもなりえると考え る。この考察からは、ウイルスの感染が起こった場合、何が感染を拡大させる要因になるの かを指摘した。 第4章は、養殖池において必要なPuddle Wheel の効果に関して、設置数と溶存酸素の関 係および微細藻類にどのように影響しているかを検証した。第5章・第6章において述べる EMS/AHPND の問題に対しても重要な対策要素として考えられる。これは、養殖池の中央
部に集積するSludge が嫌気性細菌の巣窟となることで、Puddle Wheel の重要性を認識で
きる。 先述のとおり、2006 年頃からバナメイの生産が東南アジア各国で拡大していった。バナ メイの特性は、不安定化したブラックタイガーの親エビに対して、すでに陸上での親エビ生 産が可能になっていたことが挙げられる。この変化によって、安定的に稚エビの確保ができ るようになった。また、当初のバナメイの養殖結果は、ブラックタイガーに親しんだ養殖企 業にとって驚くような結果をもたらした。まず、池入密度がブラックタイガーの 3-5 倍に
- 3 - なり、生存率も高く、生産性は著しく向上した。一方、飼料効率もブラックタイガーより 30-40%良好で、コストも安価な結果となった。これによって、東南アジアではエビ養殖域が増 えたことも相まって、東南アジアのエビ生産量は一気に増加した。この状況は、2010 年頃 から原因不明の病気として拡がったEMS/AHPND が発生するまでは継続していた。 第5章では、EMS/AHPND が起こった時からその原因の究明に取り組んだ経緯をまとめ ている。当初はウイルスなのか、細菌なのかも理解できない状況から研究を開始した。特に、 罹患したエビ個体の特徴や、発症した池の細菌にターゲットを絞り検討をした。その後、養 殖データを詳細に調べ、同時期・同稚エビの池で、罹患した池と罹患しなかった池のデータ を比較した。罹患池と非罹患池の比較から、pH の上昇と微細藻類の組成に違いを見出した。 この環境要因、pH の上昇との関連性を確認するために水槽実験を行い、バナメイへの感染 を確認するとともに初期感染(神経障害症状の発症)エビから組織検査や細菌の同定を目指 した。また、初期感染期にエビの共食いも観察し、EMS/AHPND の拡大が起こる経緯につ いても考察した。水槽実験による一連のバナメイの動向は、神経毒の可能性と使用した汚泥 の関係から細菌症であることを強く示唆するとともに、組織観察からの破壊の状態も確認 できた。さらに、細菌の同定においても、永田ら ( 2013)の検証によってVibrio属の可能 性を強く抱かせるものとなっていた。
この後 Tran et al. (2013) によってバナメイに対する 感染実験が行われ、Vibrio
parahaemolyticusが原因菌として特定された。また、Hirono et al.(2014)は、PCR の開 発を行い、EMS/AHPND の原因菌・毒素に対するアプローチは進化していった。しかし、 養殖現場におけるEMS/AHPND が完全に抑え込まれているという状況には至っていない。 第6章では、第5章での研究をもとに、高pH がバナメイに与える影響を実験レベルで観 察し、EMS/AHPND の発症を再現したものとしてまとめた。この研究から、環境ストレス と病原の関係によって病気の発症に至る過程を明らかにした。この実験によって、養殖環境 下でのEMS/AHPND 発症のメカニズムが理解され、病気を食い止め、損失を食い止めるこ とに繋がると考える。 第7章の総括では、養殖場のウイルスおよび細菌に対するバイオセキュリティーの観点 から養殖場全般のマネジメントにも触れた。本研究を通して、安定した養殖場の運営を目指
- 4 -
す指針となることを目指した。さらに、将来に向けて、エビ養殖が持続可能なものとなって いくことを希望する。
- 5 -
第2章 エビ養殖場におけるウイルスの複合感染
背景と目的
近年、アジアの新興国は目覚ましい発展を遂げてきており、国内総生産(gross domestic product: GDP)の上昇は世帯当たりの平均収入を持ち上げ、食生活も大きく変化している。 エビの消費においても大きな変化が出てきている。需要の増大に対して供給も増加傾向で あり、特にアジア各国ではエビ養殖が盛んになってきている。一方、エビ養殖の拡大に伴 いウイルス病の報告も増えてきており、安定した養殖生産・供給に不安を呈している。養 殖エビの安定供給に対する需要は今後とも増大すると考えられる。東南アジアのエビ養殖 は、2000 年代初頭に南米からPenaeus vannamei種(バナメイ)が導入され、現在では養 殖および消費現場での主流となっているが、現在においても養殖を取り巻く環境はウイル ス・細菌類をはじめとした病気とどのように向き合うのかが重要な問題と言える。 本章は、2000 年代中盤のエビ養殖事情の中で、養殖の最大阻害要因となっていたウイル スの感染に関して、Penaeus monodon(ブラックタイガー)の実際の養殖場での事例から ウイルス感染がエビ養殖に及ぼす影響について考察した。筆者は、1999 年よりマレーシア・ パハン州の Agrobest 社においてブラックタイガーの養殖を行ってきたが、ウイルス特に WSSV(White Spot Syndrome Virus)の問題は、当初より大きな問題であった。早期に PCR 分析装置を設置し、養殖場において活用してきた。親エビから養殖中のエビ、さらに養殖 場周辺の環境を知る意味でも大きな役割を果たしていた。 その中で、ブラックタイガーの養殖が成長不良や生存率の低下といった困難な状態にな った。これが、養殖種がバナメイへと移行した一因でもあるが、それは、PCR による検査 によって、ウイルスの感染が単一の感染だけでない事例が多く見受けられるようになった ことからも裏付けられる。ウイルス感染とブラックタイガーの成長・生存率の関係を把握 する目的で研究を行った。材料と方法
本章では、Agrobest 社で行われてきた実際の事業養殖データをもとに研究を進めた。 Agrobest 社は、マレーシア・マレー半島東海岸中央部に位置し、南シナ海に面して立地し ており、1984 年に設立され 1999 年より本格的にエビ養殖に取り組んできた。- 6 -
本章は、ウイルス防御に対する取り組みの一環として行ってきたウイルス検査から得ら
れた知見を集約し、分析したものである。ウイルス検査についてはPCR 検査キットとして
すべて、IQ2000 Detection & Prevention System (GeneReach, Taiwan)を使用した。
手順1.親エビの検査
Agrobest 社で採卵に利用していたブラックタイガーの親エビは、Fig. 2-1.の2海域すな わちマレーシア・タイ・インドネシア国境地帯のアンダマン海漁場とマレーシア・サバ州 のラブアン島周辺漁場から漁獲されたものである。
Fig. 2-1. Penaeus monodonの親エビ漁獲海域
漁獲された親エビは、海水に酸素を封入し、個体別に梱包された後Agrobest 社に送られ
た。搬入時に親エビを一時保管し、4種のウイルスWSSV (White Spot Syndrome Virus)、
IHHNV (Infectious hypodermal and hematopoietic necrosis virus)、MBV (Monodon baculovirus)、HPV(Hepatopancreatic parvovirus)に関して PCR 検査を行い WSSV の 検出された個体や、重篤なウイルス感染が見られた個体に関しては採卵に適さないものと して除外した。
サバ・ラブアン漁場 アンダマン漁場
- 7 - 手順2.PCR 検査によるウイルス感染池の確認 親エビを選別した後、自社ハチェリーにて採卵を行い池入れするPost Larva (PL)の養成 を行った。PL に至る前の段階、さらに池入前には再度 PCR 検査によってウイルスの確認 を行った。PL は 10-12 尾をランダムに取り出し検査し、Agrobest 社の基準に基づき PL を 評価し、池入れ基準に達したPL を養殖池に池入し、池入後1ヵ月でのウイルス保有状況を 確認した。池入後 1 カ月での検査は、3±1g のエビの個体の5尾を取り出し、末節(尾前) 部分をPCR 検査試料とした。なお、検査には、池中での遊泳反応が弱いと判断した個体を 選択した。ウイルスの確認を行い池揚まで養殖を継続し、ウイルスによる成長阻害、生存 率を調べた。 評価 手順2.のPCR 検査によって選定した養殖池をウイルス感染の感染数によって分類し、A 評価の対象とした。感染したウイルスの違いにより成長阻害、致死率の比較を行った。な お、Fig.2-3、Fig.2-4、Fig.2-5 に示したグラフで、No detect は PCR 検査でウイルスが検 出できなかったもので、Table 2-2.にあるとおり、77 池がこれに該当した。また、標準とし たものは、健常時の経験的なエビの成長をもとにしたものである。 ①生存率の算出 1日4回の飼料投下量から池中のバイオマスを算定する。日々の飼料投下量は、ブラッ クタイガーの1尾重量に対する標準飼料要求量(重量%)を給餌した。この標準に対して 4回の給餌での配分を決め、給餌毎にブラックタイガーの飼料摂餌量を基準時間内に消費 したかどうかで判断し、飼料投下量を決定した。1週間毎のサンプリングに際し得られた 平均重量から飼料摂取量が理解できるので、実際の投下量との比較からバイオマスを想定 し、平均重量で割ると推定尾数が算出される。この時、合わせて日々の摂餌状況を 1m×1m の正方形の網カゴを観察することで生存率を想定する a) 総給餌量 × 平均重量 ×池入尾数×生存率=Biomass (生物量) b) Biomass ÷ 総試料給餌量 = FCR (Feed Conversion Rate: 飼料効率) c) Biomass ÷ 平均重量 = 生存尾数
上記の a)において、総給餌量を週間給餌量に変え、平均重量を週間増加重量に変えると1 週間での Biomass の変化が理解できる。この時、b) の週間 FCR が異常であれば、給餌を
- 8 - 疑い、エビの状態により注意する。これを繰り返し、生存率をより確実に判断する。 ②平均体重 すべての養殖池では、1週間毎にサンプリングを行っていた。サンプリングは、1か所 で投げ網にてブラックタイガーを捕獲し、100 尾あたりの体重から平均体重を算出した。 1か所のサンプリングで数値に誤差があると判断した場合、さらに1か所追加し、サンプ リングを行った。
結果
手順1で行ったアンダマン漁場およびサバ州・ラブアン沖漁場にて捕獲された親エビを PCR 検査すると、Table 2-1.の結果となった。2007-2009 年の 3 年間について、Andaman 漁場では、1種保有は34%、36%、25%となっていたのに対して、2008-2009 年の 2 年間 のSaba 漁場では 53%、45%になっていた。特に Saba 漁場で捕獲した親エビは、2009 年 において、2種・3 種のウイルスを保有する個体の割合が、それぞれ 28%、5%になってお り、ウイルスが検出されなかった個体は、23%に過ぎなかった。Table 2-1. Andaman 漁場・Saba 州ラブアン沖漁場捕獲の親エビのウイルス感染状況
ウイルス数 尾数 比率 尾数 比率 尾数 比率 非感染 69 63% 368 60% 87 74% 1種感染 37 34% 221 36% 29 25% 2種感染 3 3% 22 4% 2 2% 3種感染 0 0% 2 0% 0 0% 合計 109 100% 613 100% 118 100% ウイルス数 尾数 比率 尾数 比率 尾数 比率 非感染 136 36% 9 23% 1種感染 198 53% 18 45% 2種感染 37 10% 11 28% 3種感染 4 1% 2 5% 合計 375 100% 40 100% Andaman Saba 2007 2008 2009 手順2では、Fig. 2-2.のとおり、PCR 検査によって3種のウイルス IHHNV、MBV、HPV
- 9 -
それぞれの感染が認められた事例を写真で示した。9 番目のスポットの S7 試料は、HPV、 IHHNV、MBV いずれも陽性となっていた。
IHHNV MBV HPV
Sample Result Sample Result Sample Result
1-Maker - 333/630/848bp 1-Maker - 333/630/848bp 1-Maker - 333/630/848bp 2-Positive Control 2000 copies 286/560/848bp 2-Positive Control 20000 copies 225/444/665bp 2-Positive Control 20000 copies 339/553/730bp 3-S1 Negative 3-S1 Negative 3-S1 Negative
4-S2 Positive Light 286bp 4-S2 Negative 4-S2 Positive Light 339bp
5-S3 Negative 5-S3 Negative 5-S3 Negative 6-S4 Negative 6-S4 Negative 6-S4 Negative
7-S5 Positive Light 286bp 7-S5 Negative 7-S5 Negative
8-S6 Negative 8-S6 Positive Light 225bp 8-S6 Positive Light 339bp
9-S7 Positive Light 286bp 9-S7 Positive Light 225bp 9-S7 Positive Light 339bp
10-S8 Positive Light 286bp 10-S8 Positive Light 339bp
1 2 3 4 5 6 7 8 9 10 1 2 3 4 5 6 7 8 9 1 2 3 4 5 6 7 8 9 10
Fig. 2-2. 3種ウイルス保有の個体の PCR 検査写真 Table 2-2. 養殖1ヵ月時点でのウイルス保有状況
Virus 保有ウイルス Total Reject Analysis
No Detect 0 210 133 77 HPV 1 136 77 59 IHHNV 1 43 30 13 MBV 1 6 2 4 HPV&IHHNV 2 36 16 20 MBV&HPV 2 20 7 13 MBV&IHHNV 2 2 0 2 IHHNV+HPV+MBV 3 3 0 3 手順2では、池入れ後1ヵ月の時点で、養殖池のエビをランダムに取り出し、PCR 検査し た。これを Table 2-2.のように保有するウイルスの数とウイルスの種類によって養殖池を分 類した。分類した中から、ウイルスの感染が認められなかったNo Detect については 77 池、 HPV に関しては 59 池というように生存率、平均体重の推移というような評価に使用した。 ①生存率 Fig. 2-3. は、池揚時点での保有するウイルスの種類とブラックタイガーの生存率の関 係を示した。HPV および IHHNV は生存率の面からみると最終段階まで養殖を継続し、生 存率も 80%前後と比較的高く保つことが可能であったが、MBV は単独でも生存率が低かっ
- 10 -
た。また、単独感染に比較し複合感染が見られた養殖池では、生存が低くなった。MBV に 関しては、単独の感染でも低い生存率となったが、MBV との複合感染の池はさらに生存率 が低下する傾向となった。
Fig. 2-3. 各種ウイルスとPenaeus monodonの生存率の関係
②平均体重 HPV、IHHNV、MBV の単一感染のブラックタイガーの成長に対する影響では、HPV と IHHNV の感染したエビでは、ウイルス非感染のエビと大きな違いがなかったが、標準の成 長と比較すると成長阻害は認められた(Fig.2-4.)。一方、MBV の感染池では顕著な成長阻害 が認められた。複合感染池では、さらに成長阻害が大ききなっていたが、とくに MBV が介 在している複合感染が成長の阻害がより大きくなっており、3種の複合感染が見られた池 ではさらに成長が阻害されていた(Fig.2-5)。本研究では、ウイルス感染が少なかった過去 の成長曲線を標準とし、本研究期間に PCR 検査によってウイルスを検出しなかったエビを 非感染(No detect)として区分した。 MBV HPV+MBV HPV HPV+IHHNV+MBV IHHNV IHHNV+MBV HPV+IHHNV 非検出 標準
- 11 -
Fig. 2-4. 各種ウイルス感染のPenaeus monodonの成長に与える影響
Fig. 2-5. ウイルスの感染種類による生存率と平均体重の関係 生存率(%) 標準 非検出 HPV IHHNV MBV HPV+IHHNV HPV+IHHNV+MBV HPV+MBV IHHNV+MBV 標準 非検出 IHHNV HPV MBV HPV+MBV IHHNV+MBV HPV+IHHNV+MBV HPV+IHHNV
- 12 - HPV、IHHNV、MBV の各種ウイルスの感染がブラックタイガーの生存率と成長に与える 影響を Fig. 2-5.に示した。HPV、IHHNV に関しては生存率の上では大きく影響を与えな いが、ウイルスの複合感染がブラックタイガーの生存率・成長に与える影響が大きいこと が理解できた。
考察
エビ養殖において報告されているウイルス病は多々あるが、親エビ段階で PCR にて検出 される場合も多く見受けられた。海洋で捕獲される親エビの感染率が高いということは、 養殖場からの排水などによって広がったとも考えられる。そして、親エビから再び養殖場 にウイルスが持ち込まれるという連鎖が起こっていると考えられる。 本章で取り上げているウイルスは、HPV、IHHNV、MBV であり、それぞれ単独の感染 と複合感染に関して比較を行いながら、ブラックタイガーの養殖成績が低迷した理由を探ってきた。 IHHNV は、Lu et al.(1991)によって報告され、MBV は、
Lightner et al. (1981) によって、台湾の養殖ブラックタイガーから検出され報告された。 いずれも甚大な被害をもたらす病原として取り上げられてきた。 本章の調査は、事業養殖場にて実際の養殖期間に行ったものである。したがって、完全 な管理下で比較条件以外の与件を管理している状況とは違うため、自然環境による変化を 考慮に入れた比較とは異なる。しかしながら、養殖条件をできるだけ同一にして研究を進 めた結果としてのデータは、事業養殖の実態に即した研究となっているとも言える。養殖 環境下で、ブラックタイガーに対する感染したウイルスの違いによる生存率・成長に与え る影響について考察を進めるとともに、単一の感染と複合の感染の違いに関して調べてき た。この結果、単一では影響が少ないと考えられるウイルスも複合感染を起こすと成長阻 害がより大きくなる傾向があった。このことは、単一の軽微なウイルス感染であれば問題 ないということではない。昨今のバナメイ養殖の拡大においても周辺環境の悪化から来る ウイルスの複合感染についてより注意深く養殖を行わなければならないことを示唆してい る。タイでは 2001~2002 年ころブラックタイガーの成長が著しく遅い傾向が見られ、
Kanokporn et.al (2006)は、MSGS(Monodon Slow Growth Syndrome)と命名し、HPV、
MBV、IHHNV の複合感染についても論じている。このことからも、本章で提唱したウイ ルスの複合感染が実際の養殖場で起こっていたことは容易に理解され、特定のウイルスが 原因ではないと考えられる。
- 13 -
第3章では、エビの養殖場におけるウイルスに対するバイオセキュリティ-について水 平感染の観点から研究しているが、本章で述べたウイルスの垂直感染と合わせてウイルス を養殖場に入れないための方策について研究した。
- 14 -
第3章 エビ養殖場における
ウイルスに対するバイオセキュリティー
背景と目的
筆者がマレーシアの養殖場において、エビ養殖に取り組んだのは1999 年からであるが、 2000 年代中盤にかけて、通常の集約的養殖場ではPenaeus monodon(ブラックタイガー) の親エビを海洋にて捕獲し、稚エビを生産し養殖する形態が一般的であった。種苗業者も 多数存在し、生産を行っていた。養殖場の安定的な経営には種苗の安定供給が欠かせず、 その品質も大きな養殖成立要因と考えられていた。これは現在においても同じ認識である。 筆者の経験からして養殖場へのウイルスの発現は、稚エビからのものと周辺環境からの ものに大別できると考える。昨今、東南アジアではPenaeus vannamei (バナメイ)の養殖 が圧倒的になっている。これはブラックタイガーの種苗生産が海洋からの天然親エビに依 存し、第2章で述べたとおり、ウイルスの保有リスクが増大した。一方、バナメイは陸上でSPF 種苗(Specific Pathogen Free) が生産されるようになり、養殖場にウイルスの持ち
込みリスクが減少したこともバナメイの生産が増大したことに繋がっている。バナメイは、 高密度での養殖にも適し、養殖日数もブラックタイガーに比較して短く、生産コストも大 きく下げられる効果があり、養殖業者にとって養殖しやすい養殖品種と言える。市場にも 大量に供給され、安価な値段とともに一定の支持を受け購買層を広げたものと考えられる。 もちろんこの間、東南アジア諸国の飛躍的経済成長が購買拡大要因の大きなものでもあっ た。 一方、SPF 種苗が生産される昨今でも、ウイルス病は根絶されるに至っていない。それ は、養殖場において、ウイルスの発現が種苗由来のものだけではないことを示唆している。 本章では、ブラックタイガーの養殖を通して、養殖場におけるウイルスに対するバイオセ キュリティーに関して考察した。エビ養殖場の運営には大きな投資も必要なため、ウイル ス病の防御は重要であるとともに、感染の拡大は周辺の自然に対するインパクトも与える。 持続可能な養殖を行う上で道義的責任にもかかわる。このため、ウイルスの存在を理解し、 対処する方策を考えるための指針となることを目指した。
- 15 -
材料と方法
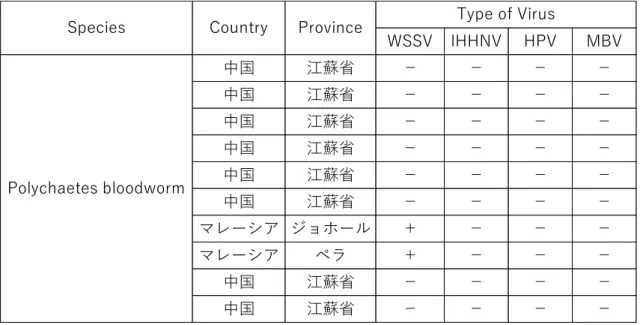
(1)ウイルスの垂直感染に関して A, 親エビのウイルス感染の変遷 ブラックタイガー養殖における天然捕獲親エビのウイルス感染状況は、Agrobest 社が導 入しているブラックタイガーの主な親エビ漁獲域のFig. 3-1.に赤丸で示す2地域から採取 された親エビを使用し調査した。1か所はマレー半島西海岸のアンダマン海、マラッカ海 峡にあるタイ、インドネシア、マレーシアにまたがる地域である。もう1か所は東マレー シア(ボルネオ島)サバ州ラブアン島沖である。なお、両地域とも沿岸部では、エビ養殖 が盛んに行われている状況にあった (Fig. 2-1)。 B, 親エビの餌料に関する検討Polychaetes bloodworm (Pic. 3-1.)の高度不飽和脂肪酸は、親エビの産卵促進に役割を果 たしている(Middledich et al., 1979,1980; Lytle et al., 1980)。また、Gomez and Arellano, (1987) は、親エビの産卵再生産にも Polychaetes bloodworm が良いとしている。これら
の情報から、親エビの餌料としてPolychaetes bloodworm を利用している。通常導入して
いる中国産のものに加えて、マレー半島南部のJohor 州および西海岸北部の Perak 州から
購入した。継続的に外部から購入する親エビの餌料についてもPCR 検査を実施した。
Fig. 3-1. Polychaetes bloodworm (Nereis sp.)
C, PCR 検査
Agrobest 社におけるPCR 検査は、簡易法としての検査キットを利用する場合と、場合
に応じては、詳細確定のためにPCR 装置による場合と、当初から PCR 装置で検査を行う
- 16 -
(Infectious hypodermal and hematopoietic necrosis virus)、MBV (Monodon baculovirus)、 HPV(Hepatopancreatic parvovirus)の4種とした。ウイルス検査については PCR 検査
キットとしてIQ2000 Detection & Prevention System (GeneReach 社)を使用した。また、
インドネシアでのフィールド調査では、現地の会社のPCR 装置を利用し判定を行った(現 地のPCR 装置は確認できず)。 サンプルの調整は、下記のとおり行い、PCR 検査した。 a) 対象のサンプルから、確認部位を滅菌したハサミとピンセットを使用し細かく刻 み、10g ペトリ皿に入れた。 b) PCR 室に送り、対象に応じてDNA・RNA の抽出を行った。 c) 抽出したDNA・RNA サンプルを電気泳動にかけて、PCR 検査を行った (2)ウイルスの水平感染に関して A, 稚エビ孵化施設におけるウイルス感染確認 Agrobest 社ハチェリーにおいて親エビの孵化を行う過程で、PCR 検査によってウイル スの出現の変遷を調べた。ウイルスについては、以下の8 種について PCR 検査を行った
(Table 4/5/6) 。 ①WSSV ・ ② TSV(Taura Syndrome Virus) ・ ③ IMNV(Infectious
Myonecrosis Virus)・④IHHNV・➄YHV(Yellow Head Virus)・⑥GAV(Gill Associated Virus)・⑦HPV・⑧MBV なお、PCR 検査・キットについては、(1)-③と同様にした。 B, インドネシア・ベトナムにおけるエビ養殖場周辺のサンプリング B-1: インドネシア・東ジャワにおけるサンプリング 2010 年 8 月末から 9 月はじめにかけて、インドネシア・ジャワ島東部に視察にサンプ リングに出掛ける機会を得た。ウイルスによって大きな被害を受けている地域の養殖場を 運営する会社からの調査依頼に応えてのものであった。 ジャワ島東部Banyuwangi 地区(Fig. 3-2)でも最盛期 9,000t の生産量が、2010 年で は5,000t を下回るのではないかとの見通しと聞いたが、実際はその数値よりはるかに低い のではないかと思わせる養殖池の稼働率であった。ジャワ島東部のバリ島の対岸に位置す る沿岸部に 30-40Km にわたって養殖場が拡がっている養殖過密地帯であった。採取した サンプルは13 サンプルで、結果は Table 3-7 に示した。
- 17 - Fig. 3-2. ジャワ島東部のサンプリング地点の衛星画像
B-2: ベトナム南部におけるサンプリング 2010 年ベトナム南部・メコンデルタ流域の養殖場においてもサンプリングを行い、イン ドネシアと同様に、PCR 検査を行い評価した。
結果
(1) ウイルスの垂直感染に関して A, 親エビのウイルス感染の変遷 Table 3-1. ブラックタイガーの親エビから検出されたウイルス感染の変遷 YearVirus Tails WSSV IHHNV MBV HPV Tails WSSV IHHNV MBV HPV Tails WSSV IHHNV MBV HPV Tails WSSV IHHNV MBV HPV
Jan 49 3 Feb 54 0 54 0 1 Mar 140 0 19 24 4 58 2 15 1 1 Apr 119 0 20 10 6 May 64 0 30 5 0 Jun 128 0 46 8 4 Jul 87 1 39 9 17 Aug 68 2 3 10 9 Sep 63 0 0 0 0 Oct 64 0 0 0 0 Nov 60 0 30 4 0 Dec 49 0 8 0 1 Total 49 3 236 0 38 4 1 660 3 211 66 41 58 2 15 1 1 Percent 6.1% 16.1% 1.7% 0.4% 0.5% 32.0% 10.0% 6.2% 3.4% 25.9% 1.7% 1.7% Jan 40 5 20 3 18 Feb Mar Apr May Jun Jul 87 39 9 17 Aug Sep 74 9 7 51 Oct 68 26 6 23 Nov 73 1 13 5 25 Dec 73 1 19 4 27 Total 375 2 106 31 143 40 5 20 3 18 Percent 0.5% 28.3% 8.3% 38.1% 12.5% 50.0% 7.5% 45.0% S ab ah Location A nd am an 2006 2007 2008 2009 Jawa Island Area B Area A・C Bali Island
- 18 - Table 3-1. のデータは、年々親エビのウイルス保有が高くなってきたことを示している とともにウイルスを複数有している親エビの出現を示している。保有するウイルスの保有 率が高くなるとともにウイルスの多様性も認められた。とくにIHHNV の保有率が非常に 高くなっていた。また、養殖場に大きな被害を与えるWSSV についても親エビ段階で検出 された。 B, 親エビの餌料に関する検査
Table 3-2. 産卵前に与える親エビの餌料(Polychaetes Bloodworm)の PCR 検査結果
WSSV IHHNV HPV MBV 中国 江蘇省 - - - - 中国 江蘇省 - - - - 中国 江蘇省 - - - - 中国 江蘇省 - - - - 中国 江蘇省 - - - - 中国 江蘇省 - - - - マレーシア ジョホール + - - - マレーシア ペラ + - - - 中国 江蘇省 - - - - 中国 江蘇省 - - - - Type of Virus Polychaetes bloodworm Country Species Province Table 3-2.の PCR 検査データから、親エビの産卵前の餌料として 使用している Polychaetes bloodworm から WSSV のウイルスが検出された。 (2) ウイルスの水平感染に関して A, 親エビとノープリウス期のウイルス検出 Agrobest 社の孵化施設での稚エビ生産のために親エビを購入した。その時のウイルス検 査データをTable 3-3 に示した。この検査から MBV 感染個体は親エビから排除したが、 IHHNV 感染個体は IHHNV 自体の感染被害が軽微ということで親エビとして使用してい た。 Table 3-3. 親エビ到着時のウイルス検査結果
Sex
Female
Male
Total
30
31
WSSV HPV MBV IHHNV
0
0
0
0
3
1
0
3
- 19 -
Table 3-3. の親エビを産卵用水槽(Maturation Tank: MT-21)に移し、親エビの片眼の Eyestalk ablation(眼柄除去)を行い、産卵を誘発させた。Table 3-4.にはメス親エビにタ グ番号を付け、親エビ毎、産卵回数毎に水槽を分けて卵を確保し、孵化したノープリウス と糞(feces)をPCR 検査にかけた結果を示している。11 尾の親から採卵した中で、1 回目 の採卵分では、2 水槽のノープリウスから IHHNV 陽性が得られ、3 水槽は糞から IHHNV 陽性が得られた。2回目の採卵ができたのは、8 尾の親であったが、このうち 3 水槽のノ ープリウスが IHHNV 陽性となった。また、YHV 陽性となった水槽も1水槽あった。3 回 目の採卵ができたのは、2 尾のみとなったが、この 2 水槽とも IHHNV 陽性となった。
Table 3-4. 1st Eyestalk ablation; 親エビ毎、産卵回別のウイルス検査結果
MT-21(Maturation Tank -21)の結果 ① ② ③ ④ ⑤ ⑥ ⑦ ⑧ Tank Tag No. No. ① ② ③ ④ ⑤ ⑥ ⑦ ⑧ ① ② ③ ④ ⑤ ⑥ ⑦ ⑧ ① ② ③ ④ ⑤ ⑥ ⑦ ⑧ ① ② ③ ④ ⑤ ⑥ ⑦ ⑧ ① ② ③ ④ ⑤ ⑥ ⑦ ⑧ MT-21 nauplius - - - - - - - - - - - - - - - - feces - - - - - - - - nauplius - - - - - - - - - - - +++ + - - - - - - +++ - - - - feces - - - - - +++ - - - +++ - - nauplius - - - - - - - - feces - - - - nauplius - - - - - - - - feces - - - - - - - - nauplius - - - +++ - - - - - - - - - - - - feces - - - - - - - - nauplius - - - - - - - - - - - +++ - - - - - - - +++ - - - - feces - - - - - +++ - - - +++ - - - +++ - - - +++ - - nauplius - - - +++ - - - - feces - - - - nauplius - - - +++ - - - - feces - +++ - - - +++ - - nauplius - - - - - - - - feces - +++ - - - - - - nauplius - - - - - - - - feces - + - - nauplius feces - - - - - +++ - -
1st Spawning 2nd Spawning 3rd Spawning 4th Spawning 5th Spawning P477 P487 P500 P490 P479 P495 P498 P475 P485 G416 P483 HPV MBV TSV
WSSV IMNV IHHNV YHV GAV
同様にMT-23 についても調べたが、Table 3-5.のとおり IHHNV 陽性となる確率が極めて高か
った。
Table 3-5. 1st Eyestalk ablation; 親エビ毎、産卵回別のウイルス検査結果
MT-23 の結果 ① ② ③ ④ ⑤ ⑥ ⑦ ⑧ Tank Tag No. No. ① ② ③ ④ ⑤ ⑥ ⑦ ⑧ ① ② ③ ④ ⑤ ⑥ ⑦ ⑧ ① ② ③ ④ ⑤ ⑥ ⑦ ⑧ ① ② ③ ④ ⑤ ⑥ ⑦ ⑧ MT-23 nauplius - - - + - - - - - - - + + - - - feces - + - - - + - - - - - +++ - - - - nauplius - - - + - - - - feces - - - - - +++ - -
1st Spawning 2nd Spawning 3rd Spawning 4th Spawning Y259
Y261
- 20 -
さらに、2 回目の Eyestalk ablation(眼柄除去)を行った 6 尾のメスの親エビについて調べると (Table 3-6.)、1 回目の産卵では、6 水槽のうち 2 水槽のノープリウスが IHHNV 陽性となり、1水
槽は糞がIHHNV 陽性であった。IHHNV 陽性となった水槽のうち1水槽は YHV も陽性であっ
た。2回目の産卵に至ったものは4 尾であったが、3 水槽のノープリウスが IHHNV 陽性となり、他
の 1 水槽も糞がIHHNV 陽性となった。このうち 2 水槽が YHV 陽性となり、1水槽は MBV 陽性
となった。親エビ番号W416 は、1 回目陰性であったにも関わらず、2 回目は YHV と MBV が陽
性となっていた。
Table 3-6. 2nd Eyestalk ablation; 親エビ毎、産卵回別のウイルス検査結果
MT-20 の結果 ① ② ③ ④ ⑤ ⑥ ⑦ ⑧ Tank Tag No. No. ① ② ③ ④ ⑤ ⑥ ⑦ ⑧ ① ② ③ ④ ⑤ ⑥ ⑦ ⑧ ① ② ③ ④ ⑤ ⑥ ⑦ ⑧ ① ② ③ ④ ⑤ ⑥ ⑦ ⑧ MT-20 nauplius feces - - - - - + - - nauplius - - - - + - - - - - - - - - - - feces - - - - - - - + nauplius - - - +++ + - - - feces - - - - - - - +++ - - - - nauplius - - - +++ + - - - feces - - - - nauplius - - - +++ - - - - feces - +++ - - nauplius - - - - - - - - feces - +++ - - - - - +++ - - - - W485 W416 W444 W499 W491 W493
1st Spawning 2nd Spawning 3rd Spawning 4th Spawning WSSV TSV IMNV IHHNV YHV GAV HPV MBV
① インドネシア・ベトナムの養殖場におけるサンプリング
Table 3-7. ジャワ島東部および北東部における採取サンプルとウイルス検査結果
No. Sample Name Place Location Scientific Name TSV WSSV IMNV IHHNV
1Shrimp Culturing Pond A Litopenaeus vannamei - - - -
2Helmite Crab Seaside B Pagurus sp. - - - -
3Ghost Crab Seaside A Ocypode sp. - - + -
4Remis Seaside A Spondylus sp. - - - -
5Crab Seaside B Brachyura sp. - - - -
6Snail Seaside B Nassarius sp. - - - -
7Redworm Seaside B Eisenia sp. - - - -
8Cycle Snail Seaside B Nerita sp. - + - -
9Remis Seaside B Spondylus sp. - - - -
10Shrimp Culturing Pond B Litopenaeus vannamei - - - -
11Crab Drain B Brachyura sp. - - - -
12Crab Drain B Cardisoma sp. - - - -
13PL Hachery C Litopenaeus vannamei - - - -
14Negative Control 15Positive Control 16Marker
* Location A: Banyuwangi area
B: North East Jawa Island C: Banyuwangi area
- 21 - この当時、インドネシアでは、IMNV が流行してた。その当時、当該地域ではエビの生 産量が著しく減少していた。3か所でのサンプリングを行ったが、養殖場では、給水・排 水域からを採取し、養殖池からもエビを採取した。今回の結果から、甲殻類からIMNV を 検出し、貝類からWSSV を検出した。 Fig. 3-3. Nerita sp. ベトナム南部の養殖場におけるサンプリング Table 3-8. ベトナム南部の養殖場で実施したサンプリングの PCR 結果
WSSV IHHNV TSV IMNV YHV GAV HPV MBV
2 Crab + ve - ve - ve - ve - ve - ve - ve - ve 3 Snail - ve - ve - ve - ve + ve - ve - ve - ve 4 Abalone - ve - ve - ve - ve - ve - ve - ve - ve 5 Black Tiger(Dead) + ve + ve - ve + ve - ve - ve - ve + ve 6 Black Tiger(Live) - ve +++ ve - ve - ve - ve - ve - ve - ve 7 Red Worm - ve ++ ve - ve - ve - ve - ve - ve + ve 8 Vannamei (WSSV Pond) +++ ve - ve - ve - ve 9 Crab 1 + ve - ve - ve - ve - ve - ve - ve - ve 10 Crab 2 + ve - ve - ve - ve - ve - ve - ve - ve 11 Crab(Big) - ve - ve - ve - ve - ve - ve - ve - ve 12 Snail 1(Big) - ve - ve - ve - ve - ve - ve - ve - ve 13 Snail 2(Small) - ve - ve - ve - ve - ve - ve - ve - ve METHOD
Nested PCR Single Step PCR
Location 1 IQ2000 Detection & Prevention Systems Location 2 Location 3 NO 検査試料 採取場所 TYPE OF VIRUS Table 3-8. は、2010 年にベトナム南部で行ったサンプリングの PCR 検査結果である。 この地域は、養殖池が密集したメコンデルタの養殖場であったが、複数のウイルスの存在 が小動物から確認されるとともに、へい死したエビ、養殖池で養殖中のエビからもウイル スが検出された。特に甲殻類は、やはり高い確率でWSSV を持っていた。Fig. 3-4、Fig. 3-5、 にそれぞれWSSV と IHHNV の PCR 検査写真を示した。
- 22 -
Fig. 3-4. PCR 写真(WSSV) Fig. 3-5. PCR 写真(IHHNV)
考察
養殖場におけるウイルスをターゲットとしたバイオセキュリティーは、垂直感染と水平 感染の両面から考えていかなければならない。 ブラックタイガーの養殖においては、天然の親エビに依存する養殖体系であるため、親 エビに対するウイルスの感染の広がりは、重大な問題を抱えている。この結果は、ブラッ クタイガーの養殖が困難なものとなった要因の1つと考えられる。特にTable 3-1 の結果 から親エビの IHHNV 感染が非常に高い確率となっていた。しかしこの当時、Lightner (1999) の IHHNV による被害は軽微であるとの報告もあり、また親エビの供給量も考慮し、 IHHNV 感染は確認されていたが、排除することはしなかった。 また、親エビの餌料として利用していたPolychaetes bloodworm からも WSSV が検出 された(Table 3-2)。このことから養殖の拡大とウイルスの拡大、天然資源への影響という シナリオが想起された。これは、Polychaetes bloodworm が沿岸部の砂の中から採取され ていることから考察される。Santiago (2005) は、Polychaete bloodworm が WSSV のベ クターとなっていることを示唆しているが、筆者の経験もそれを裏付けている。親エビへ のWSSV 感染は、このように産卵前の沿岸部での摂餌によっておこると考えれば、垂直感 染というより水平感染として扱った方が良いかもしれないが、基本的に親の時点でウイル ス感染が確認された場合は、親からの感染ということで、垂直感染として定義する。 水平感染としては、陸上・海水からのキャリアーの混入ということが考えられるが、 Table 3-4、3-5、3-6 を通して親エビからの孵化を行う際に稚エビや糞からウイルスが検出 されたことは、水槽内での感染があったと考えられる。この段階でウイルスの混合感染が- 23 - あったことは、第2章のエビ養殖場におけるウイルスの混合感染でも示したとおり、著し い成長阻害が起こる要因となる。 インドネシアの養殖場からサンプリングした小動物のPCR 検査(Table 3-7)で示されて いるとおり、周辺域のウイルス感染の状況を調べる場合、これら甲殻類や小動物も指標と して活用できる。また、ベトナムのサンプリングにおいても同様の結果が得られた。筆者 の経験では、かなりの長期にわたって使用していなかった養殖池においてもウイルスの検 出は可能であった。養殖場周辺の小動物はベクターとしてウイルスを保有している状態に ある。また、動物性プランクトンのロティファー(ワムシ)からもWSSV が検出されてお り、養殖周辺域の生物がベクターやキャリアーとなっている可能性を述べている(Yan et al., 2004; Liu, 2004)。これは、養殖環境下において多様なキャリアー、ベクターの存在を 示唆している。Liu et al. (2007) は、Juvenile (幼生エビ) において WSSV は伝搬すると 報告している。これらのことから、エビ養殖地域で、いくつかの小動物のサンプリングを 行えばその地域のウイルス汚染の状況が理解できると判断された。 養殖場において、ウイルスに対するバイオセキュリティーを考える場合、親エビからの 垂直感染を防ぐことが重要であるが、輸送時のストレス状態によっては、隠れたウイルス 感染が発現する可能性がある。親エビ到着時の検査で検出できなかったウイルスがその後 の継続的な検査で検出される事例もあった(Table 3-4,3-5,3-6)。これは、検査精度の問題か その後のストレスが発現させたのか、二次感染なのかを疑わなければならない。経過観察 においては、親エビ到着時では見られなかった感染が、その後のストレスで発現すること が伺われた。また、親エビ捕獲に際して、漁業者や仲介業者がバイオセキュリティーの概 念をもっているとは考えられず、その時点でコンタミが起こるということも否定できない。 今回の検査結果は、漁獲、集積、運送とかなりのストレスがかかっていると判断される親 エビを使用して得られたものである。港からの輸送だけでも 10 時間程度を要しているに も関わらず、到着時にウイルスが検出されず、その後の産卵の後のノープリウス段階でウ イルス発現に至ったことは、産卵のストレスの方がそれ以前のストレスより大きかったと も考えられる。精度を上げるためには、検査サンプル量を増やしたり、検査回数を増やす ことが求められる。厳格で継続的な検査実施が稚エビの管理上重要であり、養殖場のバイ オセキュリティーにとっても重要であり、周辺域のサンプリングも水平感染を予知する上 で必要と考えられる。
- 24 - 昨今、Penaeus monodonに関しても種苗のSPF 化が試みられている。また、ウイルス 汚染の少ないアフリカ種の導入なども進められているが、経験上、稚エビ品質は養殖の主 要な成功要因である。垂直感染・水平感染の防御は、各段階のリスクを知った上での管理 が必要であり、ウイルスに対する知見も重要な問題となる。 Penaeus vannamei養殖では、種苗生産は厳格に管理されたハチェリーによって行われ、 SPF 化されて垂直感染の可能性は減ってきたが、依然として品質の悪いものもある。 し かしながら、通常の養殖池では、水平感染の可能性も排除できないので、周辺環境のモニ タリングや、他の養殖場の情報、人・車両・器具・自社検査施設などもつねに把握し管理 することが肝要である。周辺の小動物のサンプリング検査は、特に養殖過密地帯の周辺養 殖業他社のウイルス感染を知る上での指標としても有効であり、今後さらに調査を進め、 感染確認指標として確立していきたい。 養殖場運営において、立地条件を考え周辺事情を理解することは非常に意義があると思 う。持続可能な養殖を行う上で、自社のバイオセキュリティーは最も重要であるが、これ を周囲地域とも協調したバイオセキュリティーにしていくことも地域社会として必要なこ とと考える。 今後とも養殖場のバイオセキュリティーについて研究を進め、持続可能なエビ養殖場の 実現を目指した提言を継続していきたい。
- 25 -
第4章
Penaeus vannmei
養殖における Puddle Wheel の
微細藻類組成に与える影響
背景と目的
Penaeus vannmei(バナメイ)は、近年のエビ養殖において主たる養殖品種になってい る。以前に主たる養殖品種であったPenaeus monodon (ブラックタイガー)に比較し高密度 の養殖が可能である。エビの体長・体重は小さめではあるが、養殖期間も短くできること で生産性も高く、東南アジア各国にバナメイの養殖が拡がった。 Agrobest 社でも 2008 年初頭からブラックタイガー養殖と並行し、バナメイ養殖に本格 的に取り組んできた。バナメイは、短期間で出荷サイズになり生産性も高いため、短期間 で養殖品種がブラックタイガーからバナメイへと変化した。Agrobest 社でも養殖関連要因 をひとつひとつ見直すことによってバナメイ養殖に適した養殖方法を検討してきた。本章 では、撹拌パドルの設定の違いによる養殖用水のAlgae 組成の違いについて研究を行った。Puddle Wheel については、2011 年から大きな被害を出した、EMS(early mortality syndrome)/AHPND(acute hepatopancreatic necrosis disease)の対策にも有効なものとな っていった。
材料と方法
① 比較区の設定 養殖池は、Agrobest 社の養殖池を使用した。 Agrobest社の養殖池は、平均 0.7ha で465 池を有し、
底面・側面ともに全面 HDPE (High Density
Polyethlene)のシートで覆われ、中央排水システム の池となっている。海水は南シナ海から直接採取し、 塩素ガスでウイルスキャリアーの存在を考慮
- 26 - に入れた消毒を行う。(殺菌工程:この工程は、2002 年から継続して行っている方法である。) 塩素の消滅を確認して養殖用水を作り始める。(水作り工程) 水作りは、一般に池揚後の養 殖池のコンディションを整え、天然餌料を作ることで、稚エビの成長を助ける重要なステ ップである。第1段階は、主としてLime (消石灰)を使って、池揚後の池底を強アルカリ 化し、酸性土壌を中和し、不要な細菌・生物の除去などを行ったり、pH を整えたり、天然 の餌料を作り出すことを目指したものである。肥料として米ぬかなどを与えることによっ て、プランクトンを発生させ養殖適正用水にする工程であり、通常 1-2 週間で池入可能とな る。また、比較を行うために、下記の養殖条件を揃えることとした。 ・稚エビ:ハチェリー・稚エビ・池入日をそれぞれ同一とした。(池入尾数は、80 尾/㎡) ・養殖池:2池を1セットとして隣接する池を同一管理者が管理した。 ・飼料・化学品は同一のものを使用するが、池の状態によって使用量は変化させた。 ② Puddle Wheel (養殖用水車)
Puddle Wheel とは、Pic. 4-2.のとお り、水産養殖で用いる水面に浮かせて 水車で外気養殖池の水を撹拌する装 置で、溶存酸素を上げる効果や養殖池 の底に溜まる汚泥を集積させる効果
を期待して設置している。
Fig. 4-2. Puddle Wheel が設置された養殖池 ③ 比較養殖池の Puddle Wheel 配置
比較区1; 養殖池 H1-7 と H2-7 (各池の大きさは、50m×140m 程度)
- 27 - Fig. 4-4. 池番号 H2-7 の Puddle Wheel 配置
青色は、Puddle Wheel の設置場所を示し、黄色で示した部分は水車によるエアーの吹き出し方向
を示している。これにらによる流れによって、養殖池の水は大きな渦を巻き中央部に集積する。
H1-7 と H2-7 はともに 0.75ha の養殖池で、水深は 1.5m(中心部)である。双方の池では、
池底堆積汚泥(Sludge)を池の中心に集めるように Puddle Wheel の設置を行った。H1-7 に
は1hp(馬力)の Puddle Wheel を 18 基設置、H2-7 は 12 基を図のとおり設置した。図中
の赤文字は、池底のサンプリングを行ったポイントを示している。 比較区 2 養殖池 J2-11 と J2-12
Fig. 4-5. 池番号 J2-11 の Puddle Wheel の配置
Fig. 4-6. 池番号 J2-12 の Puddle Wheel の配置
- 28 - 置し、J2-12 には 12 基の Puddle Wheel を設置した。 実験 4-1. 養殖日数と Puddle Wheel 数の違いによる溶存酸素に与える影響 各比較区の池に設定した定点観測点(Fig. 4-1~4.4;赤丸数字)の池底部分の水の溶存酸素 を図った。溶存酸素の測定は、ポータブルの溶存酸素計を使用した。 実験 4-2. 各養殖日数における Puddle Wheel の微細藻類組成に与える影響 各比較区の池から養殖用水を採取する。採水は、各池1地点とした。微細藻類の確認は以 下の手順で設定した養殖日数毎に行い、Table 4-1.・4-2.のとおり 1900 年代前半に提唱され たPascher の分類に準じて微細藻類を分類し比率を算出した。微細藻類の観察方法は下記 のフローのとおりとした。 ① 採水した養殖用水10ml を遠心分離機用のチューブに入れた。 ② 7 分間遠心分離機にかけた。 ③ 底部の1ml を取り出し、スライドグラスに乗せた。(カバーはしない。) ④ 40 倍の顕微鏡で観察した。 ➄ 細胞数と微細藻類の種を確認した。 Table 4-1. 植物性微細藻類の分類(Agrobest での分類細目) 分類
通称 Yellow-Green Algae Green Algae Blue-Green Algae Brown Algae Dinoflagellate Unicellular flagellates
Crysophyta Chlorophyta Cyanophyta Phaeophyta Pyrrophyta Euglenophyta
黄金色植物門 緑藻植物門 藍色植物門 褐色植物門 炎色植物門 ミドリムシ植物門
1 Ochromonas sp. Actinastrum sp. Agmenellum sp. Amphora sp. Actiniscus sp. Euglena sp. 2 Prymnesium sp. Ankistrodesmus sp. Anabaena sp. Asterionella sp. Alexandrium sp. Paramecium sp. 3 Astasia sp. Aphanocapsa sp. Biddulphia sp. Amphidinium sp. Phacus sp. 4 Chaetomorpha sp. Chroococcus sp. Chaetocerus sp. Amyloodinium sp. Prymnesium sp. 5 Charachium sp. Coelosphaerium sp. Coscinodiscus sp. Ceratium sp. Trachelomonas sp. 6 Chlamydomonas sp. Cylindrospermopsis sp. Cyclotella sp. Cochlodinium sp.
7 Chlorella sp. Gloeotrichia sp. Cymbella sp. Dinophysis sp. 8 Chlorococcum sp. Gomphosphaeria sp. Frustulia sp. Gonyaulax sp. 9 Chodatella sp. Hormothamnium sp. Gyrosigma sp. Gymnodinium sp. 10 Closterium sp. Lyngbya sp. Isochrysis sp. Gyrodinium sp. 11 Coccomyxa sp. Merismopodia sp. Melosira sp. Katodinium sp.
Phyto Plankton(植物性プランクトン)
DIVISON
Table 4-2. 動物性微細藻類の分類
分類
通称 Protozoan Rotifer Arthropod Protozoa Rotifera Arthropoda
原生動物 輪形動物 節足動物
1 Actinophry sp. Asplanchna sp. Ceriodaphnia sp.
2 Difflugia sp. Brachionus sp. Cyclops sp.
3 Epistylis sp. Filinia sp. Daphnia sp.
4 Euplotes sp. Hexarthra sp. Diaptomus sp.
5 Favella sp. Keratella sp. Moina sp.
6 Stentor sp. Naplius sp. Ostracoda sp.
7 Tintinopsis sp. Polyarthra sp. Streblocerus sp.
8 Vorticella sp. Trichocerca sp.
9 Zoothamnium sp.
Zoo Plankton (動物性プランクトン) DIVISON
- 29 -
結果
実験 4-1. 各養殖日数における撹拌 Puddle Wheel 数の違いによる溶存酸素の違い 比較区 1; 池番号 H1-7 1Hp*18 Puddle H2-7 1Hp*12 Puddle 養殖日数 1 20 40 50 Length0 Length1 Length2 Length3 Length4Width 0Width 1 Width 2 Width 3 Width 4 Width 5Width 6
Length0 Length1 Length2 Length3 Length4
Width 0Width 1 Width 2 Width 3 Width 4 Width 5Width 6
Length0 Length1 Length2 Length3 Length4
Width 0Width 1 Width 2 Width 3 Width 4 Width 5Width 6
Length0 Length1 Length2 Length3 Length4
Width 0Width 1 Width 2 Width 3 Width 4 Width 5Width 6
Length0 Length1 Length2 Length3 Length4
Width 0Width 1 Width 2 Width 3 Width 4 Width 5Width 6
Length0 Length1 Length2 Length3 Length4
Width 0Width 1 Width 2 Width 3 Width 4 Width 5Width 6 Length0 Length1 Length2 Length3 Length4
Width 0Width 1 Width 2 Width 3 Width 4 Width 5Width 6
4.5-5 5-5.5 5.5-6 6-6.5 6.5-7 7-7.5 Length0 Length1 Length2 Length3 Length4
Width 0Width 1 Width 2 Width 3 Width 4 Width 5Width 6
4.5-5 5-5.5 5.5-6 6-6.5 6.5-7 7-7.5 溶存酸素(mg/L) Fig. 4-7. 比較区1の養殖期間 (1/20/40/50 日目) の推移と池底溶存酸素の変化。(溶存酸素 の単位は mg/L)。表示の色の違いで溶存酸素量(DO)を示した。 Puddle Wheel が 18 基の H1-7 の方が、12 基の H2-7 よりも池全体酸素溶存濃度が高 い部分が多く広がるとともに、中央部の低溶存酸素領域でも違いがみられた。
- 30 - 比較区2; 養殖開始からの池底の溶存酸素量の変化をマッピングしてい。養殖開始50 日では、18 基のパドルを配置したH1-7 池の方が 12 基の H2-7 に比較して平均 1.0mg/L 溶存酸素が高 かった。また、中央部のスラッジが集積している溶存酸素が低いエリアも小さいエリアに とどまっていた。 池番号 J2-12 1hp*12基 J2-11 1hp*24基 養殖日数 34 48 61 Length0 Length1 Length2 Length3 Length4
Width 0 Width 1 Width 2 Width 3 Width 4 Width 5 Width 6 7-7.5 6.5-7 6-6.5 5.5-6 5-5.5 4.5-5 4-4.5 3.5-4 3-3.5 2.5-3 Length0 Length1 Length2 Length3 Length4 Width 0 Width 1 Width 2 Width 3 Width 4 Width 5 Width 6 7-7.5 6.5-7 6-6.5 5.5-6 5-5.5 4.5-5 4-4.5 3.5-4 3-3.5 2.5-3 Length0 Length1 Length2 Length3 Length4 Width 0 Width 1 Width 2 Width 3 Width 4 Width 5 Width 6 7-7.5 6.5-7 6-6.5 5.5-6 5-5.5 4.5-5 4-4.5 3.5-4 3-3.5 2.5-3 Length0 Length1 Length2 Length3 Length4 Width 0 Width 1 Width 2 Width 3 Width 4 Width 5 Width 6 7-7.5 6.5-7 6-6.5 5.5-6 5-5.5 4.5-5 4-4.5 3.5-4 3-3.5 2.5-3 Length0 Length1 Length2 Length3 Length4 Width 0 Width 1 Width 2 Width 3 Width 4 Width 5 Width 6 7-7.5 6.5-7 6-6.5 5.5-6 5-5.5 4.5-5 4-4.5 3.5-4 3-3.5 2.5-3 Length0 Length1 Length2 Length3 Length4 Width 0 Width 1 Width 2 Width 3 Width 4 Width 5 Width 6 7-7.5 6.5-7 6-6.5 5.5-6 5-5.5 4.5-5 4-4.5 3.5-4 3-3.5 2.5-3 溶存酸素濃度(mg/L) Fig. 4-8. 比較区2の養殖期間 (34/48/61 日目) の推移と池底溶存酸素の変化 養殖期間61 日目では、Puddle Wheel が 12 基の J2-12 が池底全体の溶存酸素が 4.0mg/L 以下となり、中央部は3.5mg/L 以下の領域が大きく拡がっていた。一方、24 基の Puddle Wheel を配した J2-11 池は、池全体として 4.0mg/L 以上であり、溶存酸素の低い中央部も 3.5-4.0mg/L にとどまるとともに、溶存酸素の低い領域も小さかった。
- 31 -
実験 4-2. 各養殖日数における Puddle Wheel の微細藻類組成に与える影響 比較区1 微細藻類組成の変化
Fig. 4-9. 比較区1H1-7 池の微細藻類組成の変化
Fig. 4-10. 比較区 1, H2-7 池の微細藻類組成の変化
Fig. 4-11. 比較区1の H1-7 と H2-7 の微細藻類組成比較。グラフ中の DOC は、Days of Culture (養殖期間)を示す。微細藻類の分類は、Table 4-1・4-2 に基づいて行った。
比較区1のPuddle Wheel 設置が 18 基で溶存酸素が高かった H1-7 池と Puddle 設置が
- 32 -
Algae (緑藻類)が Blue Green Algae (ラン藻類)と同じ程度の比率を占めていた。また、 H2-7 では、Blue Green Algae が多くなっていた。また、H1-7 では、養殖後期(49 日以降)
になるとBlue Green Algae が増加傾向になっていた。比較池において最終養殖期間が違う
のは、養殖途上での問題が起こり、池揚を行ったためである。
比較区2 微細藻類組成の変化
Fig. 4-12. 比較区2 J2-11 池の Algae 組成変化
Fig. 4-13. 比較区 2 J2-12 池の Algae 組成変化
- 33 -
比較区2においても養殖 50 日目前後まではPuddle Wheel の多い J2-11 の方が Puddle
Wheel の少ない J2-12 より Blue Green Algae が少なく安定していた。Table 4-3 に、各池 の観察最終日の微細藻類の組成を区分ごとに示した。
Table 4-3. 各池の主な微細藻類組成の区分と種類
Pond No Green Blue Green Diatom Red Tide Euglenophyta Protozoa Rotifer Arthropod
H2-7 Cocomyxa sp. Aphanocapsa sp. Chaetocerus sp. Euglena sp. Euplotes sp. Brachionus sp. Dictyosphaerium sp. Merismopodia sp. Cycotella sp.
Golenkinia sp. Micrasterias sp. Isochrysis sp. Scenedesmus sp. Microcystis sp. Nitzschia sp. Cosmarium sp. Oocystis sp.
Oscillatoria sp.
H1-7 Cocomyxa sp. Microcystis sp. Amphora sp. Amphidium sp. Euglena sp. Epistylis sp. Brachionus sp. Nanno sp. Oocystis sp. Chaetocerus sp. Gonyaulax sp. Euplotes sp.
Dictyosphaerium sp. Oscillatoria sp. Isochrysis sp. Gymnodinium sp. Favella sp. Golenkinia sp. Nitzschia sp. Peridinium sp.
Scenedesmus sp. Tetraselmis sp.
J2-12 Cocomyxa sp. Micrasterias sp. Chaetocerus sp. Alexandrium sp. Euplotes sp. Brachionus sp. Nanno sp. Microcystis sp. Cycotella sp. Amphidium sp.
Dictyosphaerium sp. Oocystis sp. Nitzschia sp. Gymnodinium sp. Golenkinia sp. Oscillatoria sp. Gyrodinium sp. Scenedesmus sp. Peridinium sp. Tetraselmis sp. Prorocentrum sp.
J2-11 Cocomyxa sp. Microcystis sp. Cycotella sp. Alexandrium sp. Euplotes sp. Brachionus sp. Golenkinia sp. Oocystis sp. Isochrysis sp. Gonyaulax sp.
Tetraselmis sp. Oscillatoria sp. Nitzschia sp. Gymnodinium sp. Peridinium sp.
考察
Puddle Wheel の増加よって池底の溶存酸素は増加した。しかし、期待した養殖池の微細 藻類バランスを整える効果は明確ではなかった。これは、養殖期間 50 日以降に関しては、 バナメイの成長に対する給餌量の増加が顕著となる。給餌量の変化により養殖池の富栄養 化が進むことが考えられ、養殖池の環境は大きく変化する。この場合、中央排水システム でのスラッジの排除が機能しないと富栄養化は急速に進むと考えられた。Moriarty (1999) が述べているように、富栄養化に伴う細菌の増殖を防ぐ意味でも環境プロバイオティクス の利用を考えなければならないと指摘している。そのためには、溶存酸素を増やすこととともにPuddle Wheel を利用した Sludge コントロールも重要なファクターとなる。
養殖池のマネジメントでは、その他にも水質に関係する化学的指標や生物学的指標をモ ニタリングしていかなければならない。2008 年刊行の「養殖と水」において、江口らは、
環境ストレスとしてこれらの重要性を取り上げている。単にPuddle Wheel を増やし溶存酸
- 34 - また、養殖コストの面からも、養殖密度に対する適正なパドル数量・配置を考えていかな ければならない。 本研究の結果から、Puddle Wheel の配置と溶存酸素の関係については、養殖池中の変動 要因とのバランスを考慮し運営していくことが肝要である。これは、実験を通して Puddle Wheel を増や、溶存酸素は高かったにも関わらず、養殖途上でへい死が見られた、J2-11 池 の例がある。これは、単に溶存酸素が高いだけで養殖結果が改善できるわけではないこと
を示している。Lightner et al., (1998) は、稚エビの SPF(Specific Pathogen Free)化や
SPR (Specific Pathogen Resistance) 化が不可欠になると述べている。これらは、バナメイ の種苗生産において実現されたきたが、エビの病気を抑え込むことには至っていないのが 現状である。環境とともに、池の形状や立地条件なども総合的に考えていく必要がある。 今後とも様々の養殖用水の環境バランスを考えた研究を継続していきたい。
- 35 -
第5章 バナメイ養殖における
AHPNS /EMS の発症原因の考察
背景と目的
Penaeus vannamei(バナメイ)の養殖は、2000 年頃東南アジアに導入され、2006 年以 降急速に養殖域を拡大し、それまでの東南アジアの主要な養殖品種であったPenaeus monodon (ブラックタイガー)を凌駕し、圧倒的な養殖量となってきた。バナメイの特徴は、 育成環境にもよるが、ブラックタイガーに比較し養殖日数が短く、高密度の養殖が可能で 養殖効率が高い(Fig. 5-1.)。また、ブラックタイガーが天然種苗に依存する割合が圧倒的で ウイルスを保有するなど不安定であるのに比較し、陸上での管理種苗生産が確立し、特定のウイルスに対してSPF (Specific Pathogen Free)種苗を得ることができることで病気に
も強い養殖品種であったことが養殖拡大の要因と言える。
バナメイ養殖の成功によって、東南アジア各国でもバナメイ養殖は拡大を続けてきた (Lightner, 2012)。しかし、中国南部において 2009 年頃に確認され、2010 年頃ベトナム・
マレーシアと拡大していったとされるEMS(Early Mortality Syndrome)/AHPND(Acute
Hepatopancreatic Necrosis Disease)によって、大きく考え方を変えなければならない事態 となった。EMS は、バナメイの Post Larva を養殖池に入れてわずかな日数で死亡が観察
- 36 - され、その後死亡率が高くなる症例であるが、Eduarudo, (2012)によると、中国・ベトナ ムは甚大な被害を受け、マレーシアにおいてもエビ養殖業は壊滅的な被害を受けるに至り、 さらにインドネシア、タイにも拡大している。一例を挙げると、中国のHainan・ Guangdong・Fujian・Guangxi では 2011 年の上半期で生産量が 80%減少した。ベトナム では、メコンデルタを中心に被害をもたらし、ブラックタイガーにも発症が確認されてい る。マレーシアのバナメイ生産量は2010 年 70,000t であったが、2011 年には 35,000t と
なった (NACA: Network of Aquaculture Centres in Asia-Pacific, 2012.8 Final Report)。
EMS の発症要因については、様々な検証がされてきているが、未だに原因が特定されて
いない。 すでに、病理学の側面からの報告ではウイルスが原因ではないことが明らかにな
ってきている (Lightner et al.,2012)。 本章では、生産現場のデータの分析を基に細菌をタ
ーゲットとしたEMS の発症メカニズムについて実験を通して検証した。なお、Aquaculture
Pacific Magazine( 2012)によると EMS の定義すら明確でないとの報告もあったが、2012
年 8 月 10 日、バンコクで開かれたNACA の会合において、他の原因の初期段階でのへい
死と区別を明確にするため、本症例をAHPNS(Acute Hepatopancreatic Necrosis
Syndrome)として扱うことが提案された。そして、2013 年に AHPND として呼称すること になった。
材料と方法
5-Ⅰ, 病理学および組織学的アプローチ a)ウイルスの確認 罹患検体のHepatopancreas(肝すい臓)・筋肉部位・遊泳脚を PCR にてウイルスの確認を行った。PCR プライマーは、IQ2000 (GeneReach Biotechnology Corp)を標準として
用いた。PCR 検査したウイルスおよび細菌は、9 種のウイルス WSSV(White Spot Syndrome
Virus)・IHHNV(Infectious hypodermal and hematopoietic necrosis virus)・TSV(Taura Syndrome Virus)・IMNV(Infectious Myonecrosis Virus)・YHV(Yellow Head Virus)・ GAV(Gill Associated Virus)・PvNV(Penaeus vannamei nodavirus)・
HPV(Hepatopancreatic parvovirus)・MBV(Monodon baculovirus)および病原細菌 NHPB(necrotizing hepatopancreatitis bacterium)である。顕微鏡による病理組織の観察は 国立研究開発法人水産研究・開発機構の増養殖研究所に依頼した。