緒 言 我が国では稀にみる超高齢社会となり、国民医 療費が40兆円を超える近年、スポーツ人口拡大を 通じた健康寿命の延伸は、持続的な社会構築にお いて必要欠くべからざるものである。それに伴い、 女性アスリートも増加する一方、女性特有の問題 点も多く指摘されている。例えば、思春期におけ る激しい運動や極端なダイエットによる体重減少 などは、初経発来の遅延や無月経、骨密度の低下 などを引き起こす可能性が高いだけでなく、女性 アスリートの身体の正常な発育、発達を妨げ、引 退後の健康にも問題をきたすことがある。女性で 最も深刻な問題は、無月経や閉経で、卵巣から分 泌されるエストロゲン量が低下することである。 エストロゲンは、ステロイドの一種で女性ホルモ ンと呼ばれている。血中のエストロゲンは、細胞 内にあるエストロゲン受容体と結合すると、核内 移行し、ゲノム DNA に結合することで転写因子 として遺伝子発現を誘導する。エストロゲン受容 体はほぼすべての細胞に存在し、また組織特異的 な生理作用をもっている。したがって、エストロ ゲン低下は、全身性の代謝異常などさまざまな病 態発症に関連する5)。 骨格筋は体重の約40%を占め、全身の代謝調節 に貢献する組織である2,9)。骨格筋は極めて可塑性 に富んだユニークな臓器であり、寝たきりなどに よって活動量が低下すると急速に萎縮し、対照的 に筋力トレーニングなどによって活動量が増加す ると肥大する。また、激しい運動により筋損傷が 起きても、速やかに再生・修復することができる。 近年、共同研究者の北島博士は動物モデルを用 いて、エストロゲンの欠乏が筋肥大効率の低下、 筋力低下、萎縮や速筋化を誘導し、更には骨格筋 幹細胞のサテライト細胞の機能不全を引き起こす ことを明らかにした4)。つまり、骨格筋特異的な
* 熊本大学発生医学研究所器官構築部門 Department of Muscle Development and Regeneration, Institute of Molecular Embryology and Genetics (IMEG) Kumamoto University, Kumamoto, Japan.
筋発生再生分野
** 長崎大学大学院医歯薬学総合研究科 Graduate School of Biomedical Sciences, Nagasaki University, Nagasaki, Japan.
*** 長崎大学医学部産婦人科 Department of Obstetrics & Gynecology, Nagasaki University School of Medicine, Nagasaki, Japan.
**** 愛媛大学プロテオサイエンスセンター Division of Integrative Pathophysiology, Proteo-Science Center, Graduate School of Medicine, Ehime University, Ehime, Japan. 病態生理解析部門
性特異的な骨格筋量制御メカニズムの解明
―
筋力維持に重要なエストロゲンに着目して―
瀬 古 大 暉
*,**北 島 百合子
***今 井 祐 記
****小 野 悠 介
*ELUCIDATION OF THE MECHANISM OF SEX-SPECIFIC SKELETAL
MUSCLE MASS REGULATION: FOCUSING ON THE IMPORTANT
ROLE OF ESTROGEN IN MAINTAINING MUSCLE STRENGTH
Daiki Seko, Yuriko Kitajima, Yuuki Imai, and Yusuke Ono
エストロゲン作用の包括的理解は、エストロゲン 欠乏による複雑化した病態解明の重要な知見とな る。 骨格筋には、エストロゲン受容体 α(ERα)と β(ERβ)の両方が発現していることが知られて いる。ERα は、骨格筋のミトコンドリア機能や 代謝恒常性維持にとって重要である8)。一方、 ERβ は骨格筋の成長や再生の調節に関与すること が報告された10)。しかしながら、骨格筋における ERβ の詳細な機能は十分に理解されていない。そ こで本研究では、骨格筋特異的 ERβ 欠損マウス を用いることで、エストロゲンを介した骨格筋量 制御メカニズムを明らかにすることを目的とした。 方 法 A.実験動物 骨格筋特異的 ERβ 欠損マウスは、ERβ-floxed マウス1)とドキシサイクリン依存的かつ筋線維特 異的に Cre リコンビナーゼを発現制御できる HSA-rtTA/TRE-Cre マウスの交配により ACTA1-rtTA;tetO-Cre; ERβf/f(mKO)マウスを作出した。
最 大 四 肢 筋 力 は 握 力 計 測 器(Columbus Instru-ments, Columbus, OH, USA)を用いて計測した。 トレッドミル走行能力は MK-680トレッドミル (Muromachi Kikai, Tokyo, Japan)を用いて計測した。
すべての実験動物は、温度、湿度が通年一定に保 たれ、12時間ごとの照明管理のもとで、水分、栄 養を十分に与え飼育した。本実験は長崎大学遺伝 子組み換え実験等安全委員会、動物実験委員会で 承認を受け実施した(承認番号:1203190970)。 B.糖負荷試験(GTT) マウスは16時間絶食後、体重10 g 当たり0.1 ml の10%グルコース溶液を腹腔内注射し、30分、60 分、そして120分で空腹時血糖値を測定した。 C.初代培養 サテライト細胞はマウスの長趾伸筋から採取し た。 サ テ ラ イ ト 細 胞 は、growth medium(GM: DMEM+30% FBS, 1 % CEE, 10 ng/ml bFGF, 1 % 抗生物質)で増殖させた。筋分化は、differentia-tion medium(DM: DMEM+ 5 % HS および 1 % 抗 生物質)で誘導した。 D.免疫組織化学染色 単離した前脛骨筋は凍結組織包埋剤を用いて、 液体窒素により冷却したイソペンタンにて凍結し た。凍結した骨格筋は、クライオスタットによっ て−20℃下で 8 μm の厚さにスライスし、シリコ ンコーティングスライドガラスに接着させた。 4 % Paraform Aldehyde(PFA)/PBS により固定後、 5 % goat serum/ 0.3% Triton X お よ び MOM kit (Vector Laboratories, Burlingame, CA, USA)で室温 20分間のブロッキングを行い PBST で洗浄後、一 次抗体で一晩インキュベートした。翌日 PBST で 洗浄後、適切な二次抗体を用いて室温で 1 時間イ ン キ ュ ベ ー ト し た。PBST で 洗 浄 後、VECTA-SHIELD(Vector Laboratories) により封入した。 染色した切片は、蛍光顕微鏡(Olympus, Tokyo, Japan)で観察した。デジタル画像は DP80カメラ と cellSens software(Olympus)で定量化した。画 像 は Photoshop(Adobe, San Diego, CA, USA) で 最適化した。
E.抗体
抗体および試薬は以下のソースから得た。 Mouse anti-type IIa myosin heavy chain(SC71)、 mouse anti-type IIb myosin heavy chain(BF-F3) (Deutsche Sammlung von Mikroorganismen)。 rat
anti-laminin α2 antibody(Santa Cruz Biotechnology, Santa Cruz, CA, USA)。Mounting Medium with DAPI と MOM kit(Vector Laboratories)。
F.RNA 抽出および定量 PCR
前脛骨筋は ISOGEN を用いて、RNA を抽出し、 抽 出 し た RNA は ReverTraAce®qPCR RT Master
Mix with gDNA Remover(TOYOBO, Tokyo, Japan)により逆転写を行い cDNA を合成した。 それらの cDNA をテンプレートして THUNDER-BIRD SYBR qPCR Mix(TOYOBO)と下記のプラ イマーを加え、定量 PCR 反応を行った。 TBP: Forward 5 -CAGATGTGCGTCAGGCGTTC-3 Reverse 5 -TAGTGATGCTGGGCACTGCG-3 AR: Forward 5 -GGTCTTCTTCAAAAGAGCCGCTG-3 Reverse 5 -TTACGAGCTCCCAGAGTCATCCCT-3 PGC1α:
Forward 5 -CCATACACAACCGCAGTCGC-3 Reverse 5 -ACCCTTTGGGGTCATTTGGTGA-3 Igf1: Forward 5 -CATGCCCAAGACTCAGAAGTCCC-3 Reverse 5 -AGGTCTTGTTTCCTGCACTTCCTC-3 Atrogin-1: Forward 5 -GACAAAGGGCAGCTGGATTGG-3 Reverse 5 -TCAGTGCCCTTCCAGGAGAGA -3 Murf-1: Forward 5 -TGATTCCTGATGGAAACGCTATGG-3 Reverse 5 -ATTCGCAGCCTGGAAGATGTC-3 Myh1: Forward 5 -CCCTAAAGGCAGGCTCTCTCA-3 Reverse 5 -TCCCCGAAAACGGCCATCTC-3 Myh2: Forward 5 -AAGAGTCCCGAACGAGGCTG-3 Reverse 5 -ACTCACAGACCCTTACTGGCA-3 Myh4: Forward 5 -CTTCATCTGGTAACACAAGAGGTGC-3 Reverse 5 -TTCTGGGCCTCGATTCGCTC-3 Myh7: Forward 5 -ACCAACCTGTCCAAGTTCCG-3 Reverse 5 -CTACTCCTCATTCAGGCCCTTG-3 G.統計解析 データはすべて平均値±標準偏差で表した。 2 群間の平均値の比較には、Student の t 検定を用 いた。統計学的有意は 5 %未満とした。 結 果 A.雄および雌の骨格筋特異的 ERβ 欠損マウ スにおける表現型の評価 本研究で使用する ACTA1-rtTA;tetO-Cre; ERβf/f (mKO)マウスは、ドキシサイクリン誘導性に ERβ を欠損できるモデルマウスである。雄および 雌の若齢 mKO マウスにドキシサイクリンを 3 週 間飲水投与後、Cre 誘導性のノックアウトマウス を作製した(図 1 A)。また ERβf/fをコントロー ル群(CON)として用いた。ドキシサイクリン 飲水投与前後における体重増加率を検証したとこ ろ、CON 群と比較して雌の mKO 群で有意に体 重増加率が低下した(図 1 B)。ERβ 欠損による 運動機能への影響を検証するために、握力テスト および走行テストを行った。CON 群と比較して mKO 群は雌のみ顕著な筋力低下を呈した(図 1 C)。一方、走行能力は両群に変化はなかった(図 1 D)。更に、全身の代謝異常の有無を検証する 図 1 .雌の骨格筋特異的 ERβ 欠損マウスは筋力が低下する
Fig.1.Muscle-specific ERβ-deficient mice display decreased muscle force generation. A.Schedule of DOX treatment and sacrifice of ERβf/f(CON)and ACTA1-rtTA;tetO-Cre; ERβf/f
(mKO)mice.
B.Body weight gain was decreased in female mKO mice(n = 4-9, each group). C.Treadmill running test to measure endurance exercise abilities(n = 5, each group). D.Grip test to measure muscle-force generation was evaluated(n = 7-9, each group). E.GTT was performed in CON and mKO mice fed with normal diet(n = 5-6, each group).
0 2 4 6 ♂ ♀ CON mKO
A
B
C
E
D
0 100 200 300 0 30 60 120 CON mKO (min) ♀GTT (Basal) Blood glucose ( mg/dl ) Weight gain (DOX Pre-Post) ♂GTT (Basal) ERβf/f(CON)ACTA1-rtTA;tetO-Cre; ERβf/f(mKO) Doxycycline
(DOX)
0 6 9 12 weeks Sacrifice
♂&♀ Max grip strength (n)
2 2.2 2.4 2.6 2.83 3.2 CON mKO ♂ 2.4 2.6 2.8 3 * CON mKO ♀ 0 100 200 300 0 30 60 120 CON mKO * Running time (min) 0 50 100 150 200 250 ♂ ♀ CON mKO
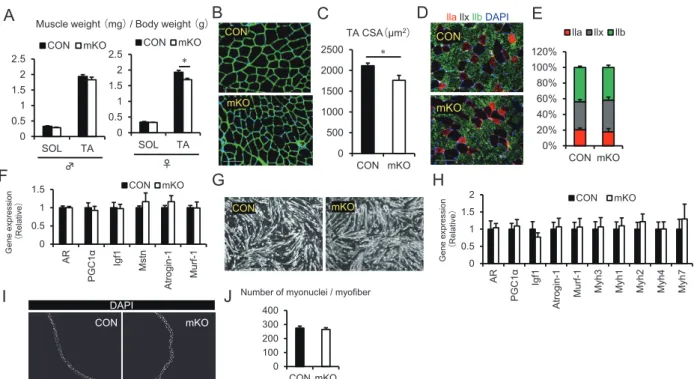
図 2 .雌の ERβ 欠損マウスは速筋特異的に筋量低下する
Fig.2.Loss of ERβ results in a reduction of fast-type muscle mass in female mice.
A.Change of muscle weight(mg)/body weight(g) of ERβf/f(CON)and ACTA1-rtTA;tetO-Cre; ERβf/f(mKO)mice. (n = 5-9, each group). SOL; Soleus, TA; tibialis anterior.
B.Immunostaining of Laminin-α2(green)on TA muscle cryosections of female mice.
C.The cross sectional area(CSA)in TA muscles was decreased in mKO mice compared with those of CON mice(n = 7-8, each group).
D.Immunohistochemistry to visualize fiber types in TA muscle of female mice. E.The proportion of fiber-type composition(n = 4-5, each group).
F.Q-PCR analysis revealed the expression of muscle mass-related genes(AR; Androgen receptor, PGC1α, Igf1; insulin-like growth factor 1, Mstn; Myostatin, Atrogin-1, and Murf-1; Muscle Ring Finger 1)in TA muscle(n = 5-6, each group).
G.Representative bright-field images of primary cultured satellite cells maintained in DM for 3 days.
H.Q-PCR analysis revealed the expression of muscle mass-related genes(AR, PGC1α, Igf1, Atrogin-1, and Murf-1)and MHC isoform genes(Myh3, Myh1, Myh2, Myh4, and Myh7)in satellite cell derived myotube(n = 5-6, each group).
I.Immunohistochemistry of DAPI in EDL-derived myofiber.
J.The number of myonuclei in EDL-derived myofiber(n = 3-4, each group). Data represent means ± SEM, *P < 0.05, significant difference vs CON mice.
A
B
C
D
E
CON mKO 0 500 1000 1500 2000 2500 CON mKO * TA CSA(μm2)F
0% 20% 40% 60% 80% 100% 120% CON mKO lla llx llb CON mKO llallxllbDAPI CON mKOMuscle weight (mg) / Body weight (g)
0 0.5 1 1.5 2 2.5 SOL TA CON mKO ♂ 0 0.5 1 1.5 2 2.5 SOL TA CON mKO * ♀ 0 0.5 1 1.5 AR PGC 1α Igf1 Ms tn Atro gin-1 Murf-1 CON mKO G ene expressio n ( Rela tiv e) 0 0.5 1 1.5 2 AR PGC 1α Igf1 Atro gin-1 Murf-1 Myh 3 Myh 1 Myh 2 Myh 4 Myh 7 CON mKO G ene expressio n ( Rela tiv e)
H
G
I
CON mKO DAPI 0 100 200 300 400 CON mKO Number of myonuclei / myofiberJ
図 3 .性成熟後における ERβ 欠損は筋力および筋量に影響しない
Fig.3.ERβ deficiency after sexual maturation does not affect muscle strength and muscle mass. A.Schedule of DOX treatment and sacrifice of ERβf/f(CON)and ACTA1-rtTA;tetO-Cre; ERβf/f(mKO)mice. B.Body weight gain was decreased in female mKO mice(n = 5-7, each group).
C.Treadmill running test to measure endurance exercise abilities(n = 4-7, each group).
D.Grip test to measure muscle-force generation was evaluated in mKO mice(n = 5-7, each group). E.Change of muscle weight(mg)/body weight(g)(n = 5-9, each group)SOL; Soleus, TA; tibialis anterior. F.The cross sectional area(CSA)in TA muscles was not changed in mKO mice(n = 6-7, each group).
B
A
C
D
E
0 0.5 1 1.5 2 2.5 CON mKO Body weight gain (DOX Pre-Post) 0 0.5 1 1.5 2 SOL TA CON mKO 0 500 1000 1500 2000 2500 CONmKO TA CSA (μm2)F
2 2.2 2.4 2.6 2.83 3.2 Max grip strength (n) CON mKO 0 40 80 120 160 200 CON mKO Running time (min) ERβf/f(CON)ACTA1-rtTA;tetO-Cre; ERβf/f(mKO) Doxycycline (DOX) 0 20 23 26 weeks Sacrifice ♀ Muscle w eights ( mg ) / Bod y w eight ( g)
ために糖負荷試験(GTT)を行った。GTT によ る血糖値の高低差は雄雌ともに変化がみられな かった(図 1 E)。以上の結果より、骨格筋特異 的 ERβ 欠損マウスは雌特異的に筋力低下を呈す ることが明らかとなった(図 1 )。 B.雌の ERβ 欠損は速筋特異的に筋量が低下 する 雌の mKO マウスは筋力低下がみられたため、 ERβ が骨格筋にどのような影響を及ぼしているの かを検証した。ノックアウト誘導後、筋重量を定 量したところ、雄マウスは遅筋優位の Soleus お よび速筋優位の TA の筋重量に変化がみられな かったが、雌マウスの TA で有意な筋重量低下が みられた(図 2 A)。雌の mKO マウスの TA にお いて筋重量の低下が確認されたため、免疫組織化 学染色による筋組織像の観察を行った。ノックア ウト誘導後、TA を採取し Laminin による筋横断 面の染色を行ったところ、mKO 群において筋横 断面積の顕著な低下がみられた(図 2 B, C)。次 に骨格筋特性の変化を検証するために、筋線維タ イプ(Type lla, Type llb, DAPI)の免疫組織化学染 色を行った。筋線維タイプそれぞれの割合は、 CON 群と比較して mKO 群に変化がみられなかっ た(図 2 D, E)。ERβ 欠損筋における前脛骨筋の 筋量および筋横断面積の低下メカニズムを明らか にするために、骨格筋の成長、肥大、および萎縮 に関連する遺伝子(Androgen receptor; AR, PGC1α, insulin-like growth factor 1 ; Igf 1 , Myostatin; Mstn,
Atrogin-1, and Muscle Ring Finger 1 ; Murf-1)発現
を検証した。CON 群に比べて mKO 群でいずれ の遺伝子も顕著な変化はなかった(図 2 F)。更に、 骨格筋特異的 ERβ 欠損マウスの長趾伸筋から単 一筋線維を単離し、骨格筋幹細胞(サテライト細 胞)を増殖させ、分化誘導させた。筋管の形状お よび分化度合いは ERβ 欠損した筋管は CON 筋管 と比べて、見た目に大きな変化はみられなかった (図 2 G)。またこれらの筋管細胞における筋成長、 肥大、萎縮および筋線維タイプの遺伝子発現を検 証したが、CON 群に比べて mKO 群で大きな変 化はみられなかった(図 2 H)。骨格筋の萎縮・ 肥大時には筋核数がそれぞれ減少・増加すること が知られている。長趾伸筋から単離してきた単一 筋線維の核を DAPI で染色し(図 2 I)、筋核をカ ウントしたが、両群に変化はみられなかった(図 2 J)。以上により、生体内における骨格筋 ERβ 欠損雌マウスは速筋特異的に筋量および筋横断面 積が小さくなることが示された(図 2 )。 C.性成熟後における ERβ 欠損による骨格筋 への影響 若齢期における ERβ 欠損は顕著な筋力低下お よび筋量低下を示したため、この表現型が成長期 を終えた性成熟後のマウスにもみられるかどうか を検証した。性成熟後20週齢の雌の mKO マウス にドキシサイクリンを 3 週間飲水投与し、Cre 誘 導性にノックアウトを誘導した(図 3 A)。ドキ シサイクリン飲水投与前後における体重増加率を 検証したところ、CON 群と比較して mKO 群で 変化はみられなかった(図 3 B)。性成熟後にお ける ERβ 欠損による運動機能への影響を検証す るために、若齢期に行った同様のプロトコルを用 いて、握力テストおよび走行テストを行った。 CON 群と比較して mKO 群は握力および走行能 に変化はみられなかった(図 3 C, D)。更にノッ クアウト誘導後、筋重量および筋横断面積を定量 したところ、両群に変化は確認できなかった(図 3 E, F)。以上の結果より、若齢期にみられた ERβ 欠損マウスの筋力低下や速筋特異的な筋重量 低下は、性成熟したマウスではみられず、ERβ は 若齢期における筋力および速筋重量の維持に重要 であることが示唆された(図 3 )。 考 察 本研究では、エストロゲンを介した骨格筋量制 御メカニズムを明らかにするために、ERβ 欠損に よる骨格筋への影響を性差に着目して検証した。 骨格筋特異的 ERβ 欠損マウスは、成長期の雌に おける筋力および速筋の筋量の維持に重要である ことが示唆された。 まず本研究では、骨格筋特異的 ERβ 欠損マウ スの性特異的な表現型の有無を検証した。ERβ を 欠損させた若齢雌マウスは、ノックアウト誘導前 後の体重増加率が減少し、筋力低下を呈した。骨 格筋以外の主要な組織(肝臓,脂肪,心臓,膵臓, および子宮)の重量計測を行った(データ掲示な
し)が、どの組織においても両群で変化はなかっ たため、この体重増加率の変化は骨格筋量低下に よるものと考えられる。共同研究者の北島博士は、 卵巣摘出によるエストロゲン欠乏は、雌マウスの 筋力低下と速筋線維の萎縮を引き起こし、その萎 縮はエストロゲンを補充することにより正常マウ スまで回復されることを報告している3,4)。つまり、 エストロゲンには筋力や筋量を維持する作用があ ることを示唆しており、本実験における雌マウス での筋力および筋量低下は、エストロゲンの作用 を一部、介したものであることが考えられる。 エストロゲンの細胞への作用は、エストロゲン 受容体(ERα および ERβ)を介したものと、受 容体非依存的なシグナルがある。ERα および ERβ の機能に共通点はあるものの、それぞれ拮抗作用 があると報告されている6)。雌マウスの骨格筋特 異的 ERα 欠損マウスは、ミトコンドリア機能不 全や代謝恒常性が破綻する8)一方で、骨格筋特異 的 ERβ 欠損マウスは代謝異常を確認できなかっ た。すなわち、エストロゲンを介した代謝調節機 構は、ERα を介したものであることが考えられる。 また、エストロゲンを欠乏させたマウスの骨格筋 は、遅筋から速筋への線維タイプが変換(速筋化) することがわかっている4)。ERβ を欠損させた骨 格筋の線維タイプ特性は、野生型マウスと変化が みられないため、エストロゲンによる筋線維タイ プの変化は ERα もしくは、受容体非依存的な作 用が制御している可能性が考えられる。骨格筋特 異的 ERβ 欠損マウスの下肢筋から単一筋線維を 単離し、骨格筋幹細胞(サテライト細胞)を増殖 させ、分化誘導した。野生型マウス由来の筋管細 胞と比較しても、筋分化度合いや形状に大きな変 化はなかった。これは生体内におけるダイレクト な筋線維へのエストロゲン曝露を模倣できていな いためだと考えられる。 過度なトレーニングをした10代の女性アスリー トの無月経によるエストロゲンの減少は、筋骨格 系の傷害リスクを高めることが報告されている7)。 ERβ が、生涯を通じて筋力および筋量維持に寄与 しているかどうかを検証したところ、興味深いこ とに、性成熟後に ERβ を欠損すると若齢期でみ られていた筋力低下や筋量低下がみられなくなっ た。このことは、若齢期の雌マウスでのエストロ ゲンを介した筋力および筋量維持機構には、少な くとも ERβ を介していることが示唆できる。 ERαKO マウスは ERβKO マウスよりも重度な エストロゲン標的組織異常を示すため、ERα は エストロゲンの主要な生理作用を介する主要な受 容体であると考えられている1)。本研究において も、ERβ 欠損は雌の筋力、筋量低下を引き起こす ことは示されたが、若齢期から長期間 ERβ 欠損 を継続しても、萎縮が顕著に大きくなることはな かった(データ掲示なし)。したがって、骨格筋 における ERβ 欠損は萎縮によるものではなく、 成長期における筋発達や発育に必要であることが 示唆された。ERβ 欠損マウスの骨格筋量低下メカ ニズムを検証するにあたり、骨格筋量調節に関連 すると報告されている既知の遺伝子発現を検証し た。しかしながら、いずれの遺伝子発現にも顕著 な変化はみられなかった。これらの結果を踏まえ、 今後は萎縮モデルなどを組み合わせた ERβ 欠損 マウスの検証、ならびに網羅的な遺伝子発現解析 により骨格筋量調節メカニズムを解明していく必 要があると考えている。 近年、骨格筋幹細胞(サテライト細胞)と性ホ ルモンに関連する知見が報告されている。骨格筋 の可塑性を議論するうえで、筋肥大や再生に重要 な役割を担うサテライト細胞を含めた、包括的な 視点は必要不可欠である。ERβ は、分化筋管細胞 よりもサテライト細胞で強く発現していることを 確認している(データ掲示なし)。以上のことから、 今後はサテライト細胞における ERβ の機能も含 めた包括的な検証も行い、エストロゲンを介した 骨格筋制御機構を明らかにしていきたい。 総 括 本研究では、骨格筋特異的 ERβ 欠損マウスを 用いて、性特異的な骨格筋量制御メカニズムを明 らかにすること目的とした。若齢期の骨格筋特異 的 ERβ 雌マウスでは、筋力低下と速筋特異的な 筋量低下を示したため、成長期における筋発達、 成長に ERβ が重要な役割を担うことが示唆され た。今後は、ERβ がどのように骨格筋量を制御し ているのか機序解明を進めるために、骨格筋幹細
胞を含めた検証をしていく必要がある。本研究を 通して、女性特有の骨格筋量制御メカニズムの一 端が明らかになれば、無月経や加齢によってエス トロゲンが低下する女性の筋力・筋量維持につな がり、生涯を通じた女性の健康増進に、有益な知 見をもたらす基礎医学研究として位置付けられる と考えている。 謝 辞 本研究を遂行するにあたり、助成を賜りました公益財 団法人明治安田厚生事業団および関係者の皆様に厚く御 礼申し上げます。また本研究を遂行するにあたり、実験 にご協力いただいた長崎大学医学部産婦人科 竹本由美子 氏、および多大なるご指導ご助言を賜りました、熊本大 学発生医学研究所器官構築部門筋発生再生分野の皆様に 御礼申し上げます。 参 考 文 献
1) Antal MC, et al.(2008): Sterility and absence of histopath-ological defects in nonreproductive organs of a mouse ERbeta-null mutant. Proc Natl Acad Sci USA, 105, 2433-2438.
2) Fujita R, et al.(2018): Zmynd17 controls muscle mitochondrial quality and whole-body metabolism. FASEB J, 32, 5012-5025.
3) Kitajima Y, et al.(2017): Soymilk improves muscle weak-ness in young ovariectomized female mice. Nutrients, 9, pii: E834.
4) Kitajima Y, et al.(2016): Estrogens maintain skeletal muscle and satellite cell functions. J Endocrinol, 229, 267-275.
5) Mauvais-Jarvis F, et al.(2013): The role of estrogens in control of energy balance and glucose homeostasis. Endocr Rev, 34, 309-338.
6) Ogawa M, et al.(2015): Female-specific regulation of skeletal muscle mass by USP19 in young mice. J Endocrinol,
225, 135-145.
7) Rauh MJ, et al.(2010): Relationships among injury and disordered eating, menstrual dysfunction, and low bone mineral density in high school athletes: a prospective study. J Athl Train, 45, 243-252.
8) Ribas V, et al.(2016): Skeletal muscle action of estrogen receptor alpha is critical for the maintenance of mitochon-drial function and metabolic homeostasis in females. Sci Transl Med, 8, 334ra54.
9) Seko D, et al.(2016): mu-Crystallin controls muscle function through thyroid hormone action. FASEB J, 30, 1733-1740.
10) Velders M, et al.(2012): Selective estrogen receptor-beta activation stimulates skeletal muscle growth and regeneration. FASEB J, 26, 1909-1920.