持久的な運動トレーニングが若年及び中年ラット骨格筋の脂肪代謝能力におよぼす影響
12
0
0
全文
(2) 冬季スポーツ研究 第8巻1号 9-19, 2005 (北海道教育大学冬季スポーツ教育研究センター紀要). 持久的な運動トレーニングが若年及び中年ラット骨格筋の脂肪代謝能力にお よぼす影響 鈴木淳一1), 平塚勇介2), 田口晴也3) 1)北海道教育大学冬季スポーツ教育研究センター, 2)北海道教育大学教育学研究科保健体育専修, 3)北海道教育大学教育学部札幌校スポーツ科学専攻 Effects of endurance training on skeletal muscle fat metabolism in young and middle-aged rats Junichi SUZUKI1), Yusuke HIRATSUKA2) and Seiya TAGUCHI3) Research and Education Center for Winter Sports1), Graduate School of Education2) and School of Education3), Hokkaido University of Education, 5-3, Ainosato, Kita-ku, Sapporo, Hokkaido 002-8502, Japan Abstract This study was designed to examine the effects of exercise training on skeletal muscle fat metabolism in young (12-week-old) and middle-aged (12-month-old) rats. Exercise training by running lasted for 6 weeks at 25 m/min for young rats, at 20m/min for middle-aged rats, 60 min/day. In the plantaris muscle (PL), aging significantly decreased 3-hydroxyacyl-CoA dehydrogenase (HAD) activity by 23% (p<0.05), whereas exercise training significantly increased the activity to the level equivalent to young sedentary group (p<0.05). In the soleus muscle (SOL), aging did not affect the HAD activity and the training significantly increased the activity by 21% in middle-aged rats (p<0.05). The activity of HAD in both muscles was negatively correlated with the increase in body weight during experimental period in middle-aged rats (p<0.05). These results indicate that although the HAD activity of skeletal muscle decreased by aging, endurance exercise training restore the activity to a level equivalent to young sedentary rats, and that greater HAD activity in skeletal muscles induced by exercise training may prevent body fat accumulation. [Keywords] aging; endurance training, HAD activity; skeletal muscle. 緒言 近年,肥満つまり脂肪組織の過剰蓄積が生活. 満者の割合は若年期から中年期にかけて急増す. 習慣病といわれる糖尿病,高脂血症,高血圧. ることや[7],30歳代以上の肥満者における糖. 症,さらには動脈硬化性疾患の発症基盤として. 尿病の発症率は肥満度が高い者ほど,また高齢. 大きな位置を占めることは周知のとおりである. になるほど増加する傾向が確認されている. [1,2]。特に,腹部型の肥満は中年期以降の肥. [8]。このような統計データから,肥満に関連. 満に伴うことが多く[3,4],生活習慣病の発症. する疾病や機能障害のリスクを軽減させるため. に深くかかわっている[5]。また,腹腔内の内. には,内臓脂肪の蓄積が促進される中年期を中. 臓脂肪の割合は中年期から高齢期にかけて増加. 心に予防対策を行う必要がある。肥満予防の原. するとの報告もある[6]。我が国では,男性肥. 則は正しい食生活と食事内容の実践を主体とす. 9.
(3) 鈴木淳一,平塚勇介,田口晴也. るが,これに運動を併用すると効果的である. 酸化能がどの程度向上するか検討した研究はこ. [2]。. れまで報告されていない。. 運動による生理効果は,大きく分けて二つ考. そこで本研究では,骨格筋の脂肪酸β酸化能. えることができる。単一の運動中の効果と運動. が若年期から中年期にかけて低下するか明らか. を繰り返すことによって生じる効果である。筋. にすると共に,中年期に行う持久的な運動ト. は安静時,脂肪組織から放出される血中の遊離. レーニングが骨格筋の脂肪酸β酸化能に及ぼす. 脂肪酸(FFA)を主たるエネルギー源としてい. 効果を調べ,それが肥満予防に貢献するか,. る[9]。しかし,ここで軽度から中等度の運動. ラットを用いて検討することを目的とした。. が開始されると一般に初期(5∼10分)には, 筋中に貯蔵されているグリコーゲンが消費さ. 実験方法. れ,続いて肝臓からのブドウ糖産生に由来する 血中グルコースが動員される。そして,運動が. 実験動物. 長時間(30分以上)に及ぶと筋活動に伴うエ. ト16匹と12月齢のWistar系雄ラット16匹を用. ネルギー源の大部分(40∼60 %)は血中FFA. いた。若年期と中年期で性が異なるラットを用. の利用へと変化し,体脂肪量の減少を助長する. いているが,先行研究において[15,16],若年. といわれている。また,長時間の持久的な運動. 期のラットでは,性が異なっていても骨格筋の. を長期間行った後の効果に関しては,基礎代謝. 代謝特性に性差はないことが報告されている。. の増加や脂肪合成の抑制,インスリン感受性の. トレーニング開始1週間前に全てのラットに. 向上,心肺機能の増強などのほか[1,2],脂肪. 対しトレッドミルに慣らすための走運動(10. 酸β酸化能つまり脂肪酸分解・利用能力の向上. m/min,10分,0 %勾配)を2日間負荷した.. が骨格筋で認められている[10]。骨格筋のミト. その後,若年期安静対照群(YC;n=8),若. コンドリアで脂肪酸のβ酸化が亢進されれば,. 年期トレーニング群(YT;n=8),中年期安. 安静時でも血中FFAの利用が促進され,内臓脂. 静対照群(MC;n=8),中年期トレーニング. 肪の蓄積が抑制されることが報告されている. 群(MT;n=8)の4グループに任意に分け. [11-13]。. た.全てのラットは室温24 1 ℃,相対湿度約. 逆に,内臓脂肪の蓄積が促進される中年期に. 50 %,12時間の明暗周期の環境下で飼育し,. おいて,骨格筋の脂肪酸β酸化能が低下してい. 飼料(CE-2,日本クレア)と水道水を自由摂. る可能性は高いが,加齢により骨格筋の脂肪酸. 取させた。. 実験には12週齢のWistar系雌ラッ. β酸化能が低下したという報告は,若年期の ラットと老年期のラットを比較した研究で報告. 運動負荷. されている[14]。この研究において,老年期で. ラット用トレッドミル(KN-73,夏目製作所). 低下した骨格筋の脂肪酸β酸化能は,持久的な. による持久的走運動を負荷した。トレーニング. 運動トレーニングを長期間実施すれば若年期と. は週5日の頻度で6週間行った。YT群では, 速. ほぼ同等の水準まで回復できることが確認され. 度25 m/min,勾配20 %に設定した。これは. ている。しかし,中年期を対象にして,持久的. 最大酸素摂取量の約80 %に相当する運動負荷. な運動トレーニングによって骨格筋の脂肪酸β. であった[17]。MT群では,最大酸素摂取量に. 10. トレーニング群のラットには,.
(4) 持久的トレーニングと骨格筋の脂質代謝能. 関する先行研究がないため,ミトコンドリア酸. ・クエン酸合成酵素(CS, EC 4.1.3.7)活性. 化酵素の活性が有意に増加した運動負荷を用い. CS活性の測定にはSrereの方法を用いた. た(速度20 m/min,勾配0 %)[18]。運動時. [20]。酵素溶液に1 mM DTNB,10 mM. 間はトレーニング初日の10分間から開始. acetyl-CoAを含む0.1 M Tris-HCl buffer(pH. し,2日目以降は1日あたり3分ずつ延長さ. 8.1)を加え,25 ℃の温水中に10分間静置し. せ,4週目に60分間に達した後はトレーニング. 温度平衡に達した後,10 mM oxaloacetateを. 最終日までこの条件を維持した。. 添加して反応を開始させた。反応開始後5分間 にわたり分光光度計を用いて412nmにて吸光. 筋標本. 最終トレーニングの48時間後にα-. 度の変化を測定した。. クロラロース(0.06 g/kg)+ウレタン麻酔下. ・ヘキソキナーゼ(HK, EC 2.7.1.1)活性. (0.7 g/kg)のラットから心臓,足底筋,ヒラ. HK活性の測定にはSrereらの方法を用いた. メ筋を摘出し,湿重量を秤量した。筋のサンプ. [20]。酵素溶液に,5 mM MgCl 2 ,10 mM. ルは液体窒素で急速冷凍し,生化学的分析を行. NADP,5. うまで-80 ℃で冷凍保存した。. glucose-6-phosphate dehydrogenaseを含む. mM. ATP,1. U/mL. 100 mM Triethanolamine-HCl buffer(pH 酵素溶液の調製. 筋サンプルは−25 ℃下で. 7.6)を加え,25 ℃の温水中に10分間静置し. 微粉状にし,5 mM β-mercaptoethanol,0.5. 温度平衡に達した後,4 mM D(+)-glucoseを添. mM EDTAを含む20 mM phosphate. 加して反応を開始させた。反応開始後5分間に. buffer(pH7.4)を用い,氷冷下にてポリトロ. わたり分光光度計を用いて340 nmにて吸光度. ン型ホモジナイザーでホモジネート(15,000. の変化を測定した。. rpm,15 sec. ・乳酸脱水素酵素(LDH, EC 1.1.1.27)活性. 3)した。そして,0. ℃,13,000 gで15分間遠心分離した後,その. LDH活性の測定にはBassらの方法を用いた. 上澄み液を酵素溶液として酵素活性の測定まで. [19]。酵素溶液に,5 mM EDTA,0.3 mM. −80 ℃で冷凍保存した。. NADHを含む100 mM triethanolamine-HCl buffer(pH 7.6)を加え,25 ℃の温水中に10. 酵素活性の測定. 分間静置し温度平衡に達した後,0.24 mM. ・3-ヒドロキシアシル-CoA脱水素酵素(HAD,. sodium pyruvateを添加して反応を開始させ. EC 1.1.1.35)活性. た。反応開始後5分間にわたり分光光度計を用. HAD活性の測定にはBassらの方法を用いた. いて340 nmにて吸光度の変化を測定した。. [19]。酵素溶液に,5 mM EDTA,0.45 mM NADHを含む100 mM triethanolamine-HCl. 統計処理. 全てのデータは平均値 標準誤差. buffer(pH 7.0)を加え,25 ℃の温水中に10. で 示 し た 。 各 群 の デ ー タ は ま. 分間静置し温度平衡に達した後,0.1 mM. ず,Kormogorov-Smirnov検定によって正規. acetoacetyl-CoAを添加して反応を開始させ. 性の検定を行い,正規分布していることを確認. た。反応開始後5分間にわたり分光光度計を用. した。有意差検定は二元配置分散分析を行い,. いて340 nmにて吸光度の変化を測定した。. 有意差がみられた場合には一元配置分散分析及. 11.
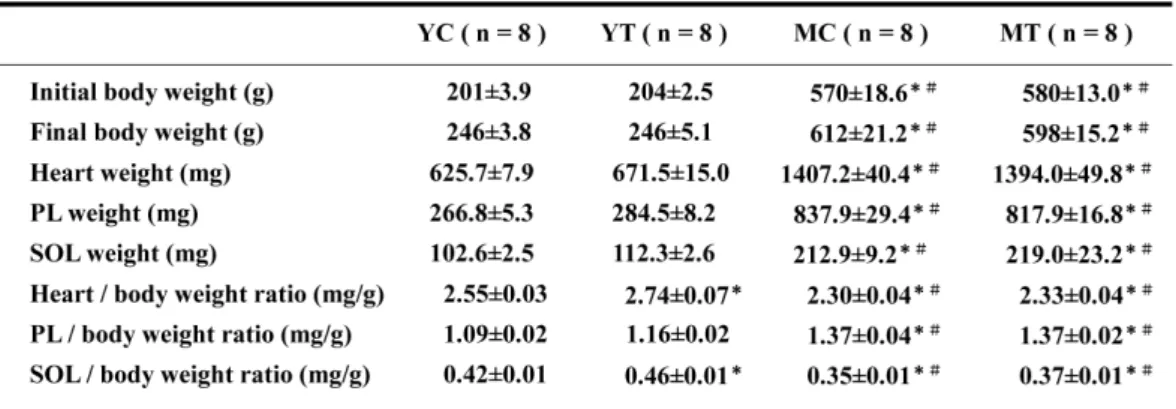
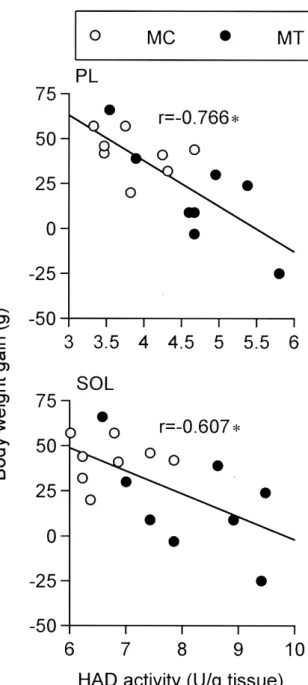
(5) 鈴木淳一,平塚勇介,田口晴也. びFisherのpost-hocテストを行い,4群間の比. 高い値を示した(p<0.05)。ヒラメ筋におい. 較を行った.検定結果は危険率5 %未満を有意. てもYT群,MT群共にそれぞれの安静対照群よ. 水準とした。. りも有意に高い値を示した(p<0.05)。HK活 性(Fig. 2)は,足底筋においてMT群がMC群. 実験結果. よ り も 有 意 に 高 い 値 を 示 し た が (p<0.05),YC群と比較すると約64 %低い値. 1. 体重および筋重量の変化. であった(p<0.05)。CS活性(Fig. 3)は,. Table 1に各群の体重,筋組織重量,体重あ. 足底筋とヒラメ筋においてYT群及びMT群共に. たりの筋組織重量を示した。実験期間中の体重. それぞれの安静対照群よりも有意に高い値を示. は , Y C 群 , Y T 群 で 有 意 な 増 加 が. した(p<0.05)。LDH活性(Fig. 4)は,足底. (p<0.05),MC群で増加傾向が認められた. 筋においてMT群がMC群よりも有意に低い値. が,MT群では変化が認められなかった。体重. を示した(p<0.05)。. あたりの筋組織重量は,心臓及びヒラメ筋にお いてYT群がYC群よりも有意に高い値が認めら. 4.骨格筋のHAD活性と実験期間中の体重増. れた(p<0.05)。足底筋においてはYT群がYC. 加量との関係. 群より高い値を示す傾向が観察されたが,有意. 中年ラット骨格筋のHAD活性と実験期間中. 差は認められなかった。. の体重増加量との関係をFig. 5に示した。骨格 筋のHAD活性は,体重増加量との間に足底筋. 2. 加齢に伴う骨格筋の代謝酵素活性の変化. で強い負の相関が(p<0.05,r=-0.766),ヒ. HAD活性(Fig. 1)は,足底筋でYC群より. ラメ筋で有意な負の相関が(p<0.05,r=-. もMC群の方が約23 %有意に低下したが. 0.607)認められた。. (p<0.05),ヒラメ筋では僅かな低下しか観 察されなかった.HK活性(Fig. 2)は,加齢. 考察. に伴い足底筋で顕著に低下した(約71 %,p<0.05)。ヒラメ筋でも約11 %低下した. 実験期間中の形態的変化. が,有意な差は認められなかった。CS活性 (Fig. 3)は,加齢に伴い足底筋及びヒラメ筋. 本研究において,中年期の安静対照群実験期. 共に有意に増加した(p<0.05)。LDH活性. 間中に体重が約7 %増加した。中年期以降に筋. (Fig. 4)は,足底筋及びヒラメ筋共に加齢に. 組織が発達・肥大するとは考えにくいことから. 伴う変化は観察されなかった。. [21],これはおそらく脂肪量増加に起因すると 推測できる。これに対し,トレーニングを行っ. 3.持久的な運動トレーニングによる骨格筋の. たラットでは,実験期間中に体重がほとんど変. 代謝酵素活性の変化. 化しなかった。また,トレーニングによって筋. HAD活性は,足底筋においてYT群がYC群. 組織重量が変化しなかったことから,中年期で. よりも有意に高い値を示した(Fig. 1;. は本研究のトレーニングによって筋組織の発. p<0.05)。また,MT群もMC群よりも有意に. 12.
(6) 持久的トレーニングと骨格筋の脂質代謝能. 達・肥大は起こらないが,脂肪の蓄積は顕著に. 性を比較した。先行研究[14 16]と本研究の骨. 抑制されたと考えられる。. 格筋の各種代謝酵素活性を比較したところ,若 年期のラットでは,足底筋及びヒラメ筋の代謝. 加齢に伴う骨格筋の脂質代謝特性の変化. 特性に性差がないことが確認された。 脂肪酸β酸化系のカギ酵素(Key enzyme). 本研究では,若年期から中年期にかけて骨格. として重要な役割を果たすHADの活性は,若. 筋の脂質代謝特性が変化するか検討するため,. 年期(YC群)から中年期(MC群)にかけて. 若年ラットと中年ラットの骨格筋の代謝酵素活. 足底筋で約23 %有意に低下した(Fig. 1)。こ. Figure 1 Effects of exercise training on 3-hydroxyacyl-CoA dehydrogenase (HAD) activity in plantaris (PL) and soleus (SOL) muscles. All values are expressed as means±SE. *, # and $, Significantly diffferent from YC, YT and MC, respectively, at P<0.05.. Figure 2 Effects of exercise training on hexokinase (HK) activity in plantaris (PL) and soleus (SOL) muscles. All values are expressed as means±SE. *, # and $, Significantly diffferent from YC, YT and MC, respectively, at P<0.05.. 13.
(7) 鈴木淳一,平塚勇介,田口晴也. Figure 3 Effects of exercise training on citrate synthase (CS) activity in plantaris (PL) and soleus (SOL) muscles. All values are expressed as means±SE. *, # and $, Significantly diffferent from YC, YT and MC, respectively, at P<0.05.. Figure 4 Effects of exercise training on lactate dehydrogenase (LDH) activity in plantaris (PL) and soleus (SOL) muscles. All values are expressed as means±SE. * and $, Significantly diffferent from YC and MC, respectively, at P<0.05. れは骨格筋のβ酸化能は加齢に伴い低下するこ とを示している。若年ラットと老年ラットを比 較した研究でも[14,15],加齢により骨格筋の HAD活性の低下が観察されている。加齢に伴 い骨格筋のミトコンドリアで脂肪酸のβ酸化が 抑制されれば,骨格筋での脂肪酸消費が減少し て血中に遊離脂肪酸(FFA)が増加し,これが 肝臓や脂肪細胞での中性脂肪合成を亢進させる. 14. Figure 5 Relationship between body weight gain and HAD activity in plantaris (PL) and soleus (SOL) muscles in middle-aged rats. *, Correlation was statistically significant at p<0.05. と考えられている[2]。中性脂肪の合成及び蓄 積が過剰に進行すると,体脂肪のさらなる増加 に対する防御機構として脂肪細胞から血中への.
(8) 持久的トレーニングと骨格筋の脂質代謝能. FFA放出が促進されるが[22,23],β酸化能が. れがβ酸化能の低下を軽減させているのかもし. 低下した骨格筋においてFFA消費は亢進され. れない。ただヒトの場合,立位時にのみ下肢に. ず,結局は中性脂肪合成の方向に向かう。この. 負荷が掛かるということは十分に考慮する必要. ような悪循環が成立してしまうと,少ない摂取. がある。また,中年期にβ酸化系が低下する機. エネルギーでも太りやすい体質に移行し,体脂. 序に関しては,骨格筋などの末梢組織へのFFA. 肪の蓄積がさらに加速すると考えられている. 運搬に深くかかわる微小循環系と[18],骨格筋. [2]。現に肥満者(Body Mass Index;. 及びミトコンドリア内へのFFA輸送系の加齢変. BMI 41.3 2.0. kg/m2)では,骨格筋の1時間. 化を含めて今後さらなる検討が必要である。. あたりの脂肪酸酸化(消費)量が非肥満者 (BMI 22.6 0.6 kg/m 2 )よりも約45∼60. 持久的な運動トレーニングによる骨格筋の脂質. %低下しているという報告がある[25]。このよ. 代謝特性の変化. うに,若年期から中年期にかけて骨格筋の脂肪 酸β酸化能が低下することと,中年期に肥満者. 本研究において,中年ラット骨格筋のHAD. が急増する統計的事実との間には,何かしら因. 活性は,足底筋及びヒラメ筋共にトレーニング. 果関係があると思われる。. によって約21 %有意に増加し,若年期の非ト. また,β酸化の低下に起因する血中FFAの上. レーニングラットと同等か(足底筋),あるい. 昇は,インスリンによる骨格筋へのグルコース. はそれ以上の水準に(ヒラメ筋)達していた. 輸送を阻害するため[25],その結果グルコース. (Fig. 1)。これは加齢に伴い骨格筋の脂肪酸. を基質とするHK活性が抑制される可能性は高. β酸化能は低下するが,持久的な運動トレーニ. い。現に本研究では,加齢に伴い足底筋のHK. ングを長期間実施すれば,若年期とほぼ同等か. 活性が約71 %も低下した(Fig. 2)。このこと. それ以上の水準まで回復できることを示してい. から,中年期ではβ酸化系の低下に伴い解糖. る。骨格筋のミトコンドリアで脂肪酸β酸化能. 系,特に血中グルコースの有酸素的代謝能力が. が回復すれば,骨格筋で多くの血中FFAを代謝. 顕著に低下すると考えられる。インスリン刺激. できるようになり,これが上昇していた血中の. による血中グルコース取り込みの75 %以上は. FFA濃度を減少させ,やがて正常値に戻ると考. 骨格筋によるものであり[22],骨格筋への糖輸. えられる。安静時の生体内では,血中FFA濃度. 送を含めたグルコース代謝系の低下が,血糖上. が正常な状態であれば,骨格筋でのFFA消費量. 昇さらには糖尿病発症の誘因となる可能性は高. が増加すると,これに伴い脂肪組織からのFFA. い。. 放出量も増加する[28]。骨格筋の脂肪酸β酸化. 一方,ヒラメ筋では加齢に伴いHAD活. 能が回復し,このような代謝状態を獲得できれ. 性,HK活性共に僅かな低下しか観察されな. ば,太りにくい体質に移行し,体脂肪の蓄積が. かった(Figs 1&3)。ラットの後肢にはゲー. 抑制されると考えられる。現に肥満モデルラッ. ジ内の限られた日常活動においても常に体重が. トでは,脂肪酸異性体の一種である共役リノー. 掛かっており,特に姿勢維持筋としての主働筋. ル酸を添加した食事によって骨格筋のβ酸化系. であるヒラメ筋(遅筋)において,代謝的な負. を亢進させた場合,1日あたりのエネルギー消. 担(FFA代謝)が増加していると考えられ,こ. 15.
(9) 鈴木淳一,平塚勇介,田口晴也. 費量が増加して,内臓脂肪の蓄積量が低下した. るが,若年レベルに戻すことは非常に困難であ. という報告がある[13]。. ることを示している.. 長期間の持久的な運動トレーニングによって 骨格筋の脂肪酸代謝経路の進行がより加速すれ. 骨格筋の脂肪酸β酸化能と体脂肪蓄積の抑制効. ば,運動時の血中グルコースの利用量が減少す. 果. ると考えられている[9,29,30]。例えば,日々 持久的な運動トレーニングを行っている者で. 本研究において,中年ラット骨格筋のHAD. は,自覚的運動強度がややきついとされる程度. 活性と体重増加量との間に有意な負の相関が認. の運動(最大酸素摂取量の約65 %)を120分. められた(Fig. 5)。これは,骨格筋の脂肪酸. 間実施した場合,血中グルコースのエネルギー. β酸化能が優れているほど,体重増加がより軽. 貢献率は常に10 %未満である(非トレーニン. 減されたことを示している。つまり,運動に. グ者では約23 % [30])という報告がある. よって骨格筋の脂肪酸β酸化能が回復すれば,. [31]。同様に,HAD活性が高く脂肪酸代謝に. 骨格筋での脂肪酸消費が亢進して,体脂肪の蓄. 優れる若年ラットの骨格筋においても,運動時. 積が大きく抑制されると考えられる。運動時で. に血中グルコースの代謝が促進される可能性は. は,エネルギー需要増大に伴い骨格筋のβ酸化. 低い。本研究のトレーニングでグルコースを基. 系がほぼ最大限に進行していると考えられる.. 質とするHKの活性が若年ラットの骨格筋でほ. このときβ酸化系を進行させるHADの活性も. とんど変化しなかったのも(Fig. 2),おそら. ほぼ最大限に亢進されており,このため運動時. く運動時に血中グルコースの代謝が促進されな. ではHADの最大活性能力の差がそのまま骨格. かったためであろう。一方,中年ラット骨格筋. 筋での脂肪酸代謝量に反映されると推測でき. のHK活性はトレーニングによって有意に増加. る。例えば,トレーニングによって骨格筋の. した(Fig. 2)。これは,中年ラットでは本研. HAD活性が約2倍に増加した場合,中等度運動. 究のトレーニングによって,骨格筋のHK活性. 時(最大酸素摂取量の約65 %)の骨格筋での. を高めるために十分な代謝的刺激が負荷された. 脂肪酸代謝量も約2倍に増加したとの報告があ. ことを示している。つまり,脂肪酸代謝能力が. る[30]。. 低下している中年ラットの骨格筋では,筋収縮. 以上,本研究では加齢に伴い低下した中年. に必要なエネルギーを確保するために血中グル. ラット骨格筋のHAD活性は,長期間の持久的. コースの代謝を促進させる必要があったと考え. な運動トレーニングによって若年ラットと同等. られる。ただ,中年ラットの足底筋ではトレー. か,それ以上の水準まで回復できることが確認. ニングによってHK活性が約23 %有意に増加し. された.また,骨格筋のHAD活性が優れてい. たが,若年期の非トレーニングラットと比較す. ることは,特に運動時の骨格筋での脂肪酸代謝. ると約64 %も低い値を示していた(Fig. 2)。. 量をより増加させると考えられた。これらのこ. これは,グルコース代謝系は,加齢により一旦. とから,肥満者が急増する中年期において,体. 顕著に低下してしまうと,持久的な運動トレー. 脂肪の蓄積を予防する手段として,骨格筋の. ニングによってある程度回復させることはでき. HAD活性をより高める運動を行うことの有効 性が示唆された。. 16.
(10) 持久的トレーニングと骨格筋の脂質代謝能. 8. Tsugane S, Sasaki S, Tsubono Y. 2002. Under- and over-weight impact on. 謝辞 本研究の一部は,平成16年度北海道教育大学. mortality among middle-aged Japanese. 学長裁量経費(若手教員研究支援経費)によっ. men and women:a 10-y follow-up of. て行われた。. JPHC study cohort I. Int J Obes Relat Metab Disord 26, 529-537. 9. Lorraine P Turcotte, Erik A Richter,. 参考文献. Bente Kiens. 1992. Increased plasma 1. 池田義雄, 井上修二, 井口利樹, 大村裕, 大. FFA uptake and oxidation during. 野誠, 奥田拓道, 片岡邦三, 河上征治, 川村. prolonged exercise in trained vs.. 功, 衣笠昭彦, 佐藤祐造, 徳永勝人, 丸浜喜. untrained humans. Endocrinol Metab. 亮, 吉松博信. 1997. 肥満・肥満症の指導マ. 25, 791-799.. ニュアル. 医歯薬 出版株式会社. 2. 井 上 修 二 , 池 田 義 雄 , 大 野 誠 , 宗 像 伸 子 .. 10.鈴木淳一, 平塚勇介, 東浦拓郎. 2004. 持久 的走行トレーニング初期段階における骨格 筋の脂質代謝関連酵素活性の変化過程. 北. 1994. 肥満症テキスト. 南江堂. 3. Vansant Gm, Den Besten C, Weststrate JA, et al. 1988. Body fat distribution and. 海道教育大学冬季スポーツ教育研究セン ター紀要7, 9-14.. weight. 11.Rahman SM, Wang YM, Yotsumoto H,. reduction:preliminary results. Int J. et al. 2001. Effects of conjugated linoleic. Obesity 12, 133-140.. acid on serum leptin concentration,. the. prognosis. for. 4. Den Besten C, Vansant G, Weststrate. body-fat accumulation, and. JA, et al. 1988. Resting metabolic rate. beta-oxidation of fatty acid in OLETF. and diet-induced thermogenesis in. rats. Nutrition 17, 385-390.. abdominal and gluteofemoral obese. 12.Wang YM, Rahman SM, Nagao K, et al.. women before and after weight. 2003. Comparison of the effects of. reduction. Am J Clin Nutr 47, 840-847.. triacylglycerol-CLA on hepatic lipid. 5. 下方浩史. 1993. 体脂肪分布−腹部型肥満 の基礎と臨床−. 杏林書院.. metabolism in OLETF obese rats. J Oleo Sci 52, 121-128.. 6. B o r k a n G A , H u l t s D E , G e r z o f S G ,. 13.柳田晃良, 永尾晃治. 2003. 共役リノール酸. Robbins AH, Silbert CK. 1983. Age. の抗肥満・抗高脂血症作用とその機序. 肥. changes in body composition revealed. 満研究 9, 194-196.. by computed tomography. J Gerontol 38, 673-677. 7. 厚生労働省. 2003. 平成14年の国民栄養調 査.. 14.J o r g e S a n c h e z , C a t h e r i n e B a s t i e n , Hugues Monod. 1983. Enzymatic あ daptations to treadmill training in skeletal muscle of young and old Rats. Eur J Appl Physiol 52, 69-74.. 17.
(11) 鈴木淳一,平塚勇介,田口晴也. 15.Scott K Powers, John Lawler, David. diabetes and obesity. In Insulin. Criswell, Fu-Kong Lier, Daniel Martin.. resistance (ed. By Moller DE) pp.. 1992. Aging and respiratory muscle. 187-252, John Wiley, Chichester, 1993.. metabolic plastisity:effects of endurance training. J Appl Physiol 72, 1068-1073.. 23.Flatt JP. 1996. Substrate utilization and obesity. Diabetes Rev 4, 433-449.. 16.Abdelmalki A, Fimbel S, Mayet-Sornay,. 24.Kim Jong-Yeon, Robert C Hickner,. Sempore B, Favier R. 1996. Aerobic. Ronald L Cortright, G Lynis Dohm,. capacity and skeletal muscle properties. Joseph A Houmard. 2000. Lipid. of normoxic and hypoxic rats in. oxidation is reduced in obese human. response to training. Eur J Physiol 431,. skeletal muscle. Am J Physiol. 671-679.. Endocrinol Metab 279, 1039-1044.. 17.B r o o k s G A , W h i t e T P . 1 9 7 8 .. 25.Drensner A, Laurent D, Marcuccl M, et. Determination of metabolic and heart. al. 1999. Effects of free fatty acids on. rate responses of rats to treadmill. glucose transport and IRS-1-associated. exercise. J Appl Physiol 45(6),. phosphatidylinositol 3-kinase activity. J. 1009-1015.. Clin Invest 103, 253-259.. 18.Suzuki J, Gao M, Koyama T, Tatra S. 1997. Effects of treadmill training on the. 26.永田和宏. 1995. ストレス蛋白質とは何か. 生体の科学 46, 298-302.. arteriolar and venular portions of. 27.Mattson JP, Ross CR, Kilgore JL, Musch. capillary in soleus muscle of young and. TI. Induction of mitochondorial stress. middle-aged rats. Acta Physiol Scand. proteins following treadmill running.. 159, 113-121.. Med Sci Sports Exerc 31, 365-369.. 19.Bass A, Brdiczca D, Eyer P, Hofer S,. 28.Klein S, Peters EJ, Holland OB, Wolfe. Pette D. 1969. Metabolic differentiation. RR. 1989. Effect of short and long-term. of distinct muscle types at the level of. β-adrenergic blockade on lipolysis. enzymatic organization. Europian J. during fasting in humans. Am J Physiol. Biochem 10, 198-206.. 257 (Endocrinol Metab 20), 65-73.. 20.S r e r e P A . 1 9 6 9 . C i t r a t e s y n t h a s e . Methods in Enzymology 13, 3-11.. 29.Martin WH, Dalsky GP, Hurley BF, Matthews DE, Bier DM, Hagberg JM,. 21.山 下 勝 正 , 代 田 琢 彦 , 平 岡 亮 , 吉 岡 利 忠 .. Rogers MA, King DS, Holloszy JO. 1993.. 1994. 加齢に伴い低下した筋機能に及ぼす. Effect of endurance training on plasma. 運動トレーニングの意義と適切な負荷方法.. free fatty acid turnover and oxidation. 体力研究 85, 6-17.. during exercise. Am J Physiol 265. 22.Seely BL, Olefsky JM. Potential cellular. 18. (Endocrinol Metab 20), 65-73.. and genetic mechanisms for insulin. 30.Jansson E, Lennart K. 1987. Substrate. resistance in the common disorders of. utilization and enzymes in skeletal.
(12) 持久的トレーニングと骨格筋の脂質代謝能. muscle of extremely endurance-trained men. J Appl Physiol 62(3), 999-1005. 31.Romijin JA, Coyle EF, Sidossis LS, Gastaldelli A, Horowitz JF, Endert E, Wolfe RR. 1993. Regulation of endogenous fat and carbohydrate metabolism in relation to exercise intensity and duration. Am J Physiol 265 (Endocrinol Mtab 28), 380-391.. 19.
(13)
図
関連したドキュメント
どにより異なる値をとると思われる.ところで,かっ
その ため に脂肪 酸代 謝 に支.. Cation/Carnitine
そこでこの薬物によるラット骨格筋の速筋(長指伸筋:EDL)と遅筋(ヒラメ筋:SOL)における特異
Effects of age on functional independence measure score gain in stroke patients in kaifukuki rehabilitation ward. Multivariate analysis of improvement and outcome
タービンブレード側ファツリー部 は、運転時の熱応力及び過給機の 回転による遠心力により経年的な
いられる。ボディメカニクスとは、人間の骨格や
本学陸上競技部に所属する三段跳のM.Y選手は
敷地と火山の 距離から,溶 岩流が発電所 に影響を及ぼ す可能性はな
