長野大学紀要 第14巻 第1号 100-112頁 1992
染色体 と複屈折
Chromosome and Birefringence
は じめ に
生物の世界は、通常痛過性病原体 (Virus)、菌 類(Fungus)、原核生物(Prokaryotes:Bacteria) および真核生物 (Eukaryotes)の4界に分け られ るが、何れ も遺伝子記号の基本構造 としてデオキ シ リボ核酸 (DNA)を共有す る。すなわち、一 見似て もつかぬ迄に分化 したこれ等の生物群 は、 43憶年前に海水 中に初めて出現 した同一の生命起 源体か ら出発 し、Watson-Crickが提示 した長 い 組状のDNAの2重 ラセ ン構造の基礎構造である アデニン とチ ミン
(
A-T)
、およびグアニンとシ トシン (G-C) を 2つの文字 として様々な順序 に組合せ、総ての遺伝子記号 を綴 っているのであ るO 言い換 えると、生物は水 とい う地球特有の物質 をたん 白質 を主体 とす る生体構成物質の溶媒 とし、 ヒモ DNAとい う安定 な紐の上 に刻 まれた遺伝子記号 に従 って形態 と機能 を維持 し、かつ次代へ伝承す るとい う方法で生 き長 らえて きた。勿論、長い時 間の経過 の うちには、方言がで きた り、新 しい遺 伝子が付け加 えられた り、消 された りす る。 それ が進化であ り、現代の生物のDNAの上には、43 億年の生物進化の歴史が刻 まれていると考 えてよ いであろ う。 DNA上の各遺伝子は、生命活性維持のために、 アデニ ン、 グアニン、 シ トシン、ウラシルの4
文 字の うち3
文字の組合 わせ、即 ち4
3-6
4
通 りに転 写 され、 メッセー ジは細胞質- と輸送 され、そこ ホンヤク で翻訳 されて特定のア ミノ酸配列 を持つペプチ ド 合成が行 はれ る。それが重合 して多種 多様のたん 白質になってゆ く。生体の必須ア ミノ酸は2
0
又は2
1
種類であるか ら、6
4
文字で充分 なのである。 ビールス以外の生命体 の基本構造 を細胞イ(Cell) と言い、DNAは核 とい う多孔性の袋に容れ られ佐
藤
英
美
Hidemi Sato
ているが、細胞は条件 さえ整 えば常に一定の速度 でDNAを複製 してお り、複製が終ればDNAを 等価配分す る為 に分裂す る。それが細胞集合体 で ある真核生物に進化 し、機能分化 と対応 した細胞 分化 をお こす と分裂 をやめ、代 りに構造の複雑化 と、それに引続 く老化が始 まる。所謂組織分化 ・ 器官分化 と呼ばれ る現象であるが、その ような組 織細胞で も分化以前の状態に戻せば分裂 を再開す る。 これが突発的に生体 内でお これば癌にな り、 人間の管理下 で試験管内で行 えば組織培養細胞 と なる。但 し分裂 を再開 して も、組織培養細胞は突 然変異 をおこさぬ限 り、単一細胞起源であるクロ ー ン細胞群は死滅す る。分裂 は細胞の重要 な属性 であるが、一旦分化 した細胞の先祖返 り系では、 老化は避け られない。 染色体 とは 細胞が増殖す るときにDNAの複製は必ずおこ るが、DNA分配の機構は生物群 によって異なる。 最 も進化 した真核生物では、有糸分裂 とい う独特 の形式が とられ る。即 ち細胞周期 の合成期 (S一 期) を通 じて複製 されたDNAは、G-2期 と有 糸分裂期 (M一期)の初期 にかけて、丹念に2重 ・3重、時には4重の コイルに巻かれ、 ヒス トンヒモ で代表 され る塩基性たん 白質で保護 された紐様の 塊 りになる。人間の細胞の大 きさは、平均 して直 径10/〃n。DNAの容れ ものであ る核は、たかだか 直径3-4/Jmにす ぎないのに、その中に封 じこ め られたDNAの全長は数 メー トルに も及ぶ。 こ ツナ れ を-繋 ぎの遺伝子群 (linkage)毎に、生物の種 によって定 まった数 と形のパ ッケー ジにまとめ る のである。 これが所謂染色体 である。 この染色体 の形成過程に少 しの誤 りもあってはいけない。部 分的な切断、修復の些細 な ミス、部分の欠失、DNA破 片の取 り込み等は、何 れ も突然変異につ な が る。 生の分裂細胞内に形成 され る染色体 は、通常の 光学顕微鏡 では殆 ん ど視 認で きない。 それは染色 体 と、 それ を取囲む細胞質の屈折率が殆 ん ど等 し いために、 コン トラス トを作 るのに充分 な光路程 差 がで きないか らであ る。可視光 は電磁波の一部 分 であ り、光 の透過 は誘電体理論 に従 うが、透過 す る細胞構造の構成 たん 白質の濃度差が極め て小 さいために波の遅 れの差が僅か で、構造 を識別 で きないのである。 そのため、19世紀 中葉か らは細胞や組織 を固定 し、染色 して内部構造 をみ るとい う方法が とられ た
。E.
R.
Vi
r
c
ho
w
は "細胞は細胞か ら生ず る''と 喝破 し、病理学者は必ず顕微鏡 を使用す るよ うに 説 いたが、今世紀半 ば迄、見難 い細胞構造 を見易 くす るために敢 て時間軸 を放棄 し、 その代 りにで きる限 り生 きた状態に近 く細胞 を殺 して保有 し、 構成 たん 白質のpHの差 を利用 して酸性や塩基性 の色素 で、或 いは重金属 イオン との親和力 を利用 して構造 を染め分 け る工夫 に全力が注がれた。染 色体(
Ch
r
o
mo
s
o
me
)
とい う名称 は、 こ うして生 れた。DNAとい う核酸 を基礎成分 に持 ち、有糸 分裂期 に コンパ ク トに まとめ られ るこの構造は、 酸性 であ るために塩基礎性色素 と反応 し、良 く染 ま っ た か らで あ る。 因 み にCh
r
o
mo
は 色 を、So
me
は物の塊 を表す。 しか し固定 ・染色法 に頼 っていては、細胞の動 態は見 えて こない。時々刻々 と変化す る細胞の生 理状態 を追跡す るこ とはで きない。 これ を打破 し たのがF.Ze
r
n
i
c
ke
(
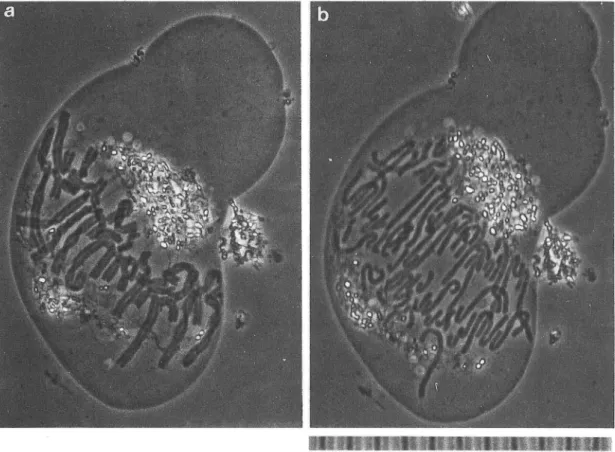
27)の結像理論 に基 く位相差顕 微鏡 の開発 であ り、実用化 であった。屈折率 の差 恥 幻 怪 ∃騨 隆 軸 蜜図1 アマ ))))ス科のHaemanthuskatherinaeの内肱乳細胞内に、形成された有糸分裂期の染色体 を示す。 aは中期染色体で核板 を形成 している。bは20分後に後期に入 り、対合 していた娘染色体が分離 して両 極へ移動 している像 を示す。た ゞし、紡錘体は観察できない。位相差顕微鏡像.右下段のスケールの1
目盛は10FLm0
-102 長野大学紀要 第14巻 第1号 1992 が僅少 なために観察 し難か った生体細胞内構造 を、 小 さな屈折率差 を増幅 して コン トラス トに変 える こ とに成功 したこの装置は革命的であった。 その 例 を図
1
に示す。 さらに偏光 を効果的に利用す るこ とで、 これ迄 見 ることので きなか った細胞内超微細構造の配列 や生体高分子の配向の動 態 を、複屈折性 または偏 光 のベ ク トル を利用 した光干渉 を強調 して コン ト ラ ス トへ 変 換 し、定 量 す る方 法 が 確 立 さ れ た(9・20・21)。顕微鏡の改良が、これ迄 の形態学の泣 き 所 であった時間軸の導入 を許 し、細胞に直接問い かけ、生体 内反応 を追跡す るこ とを可能に した。1
9
5
0
年の頃である。 この時か ら、形態記載に終始 していた細胞学 は、学際的な細胞生物学へ と大 き く変貌 してゆ く。 複屈 折 につ い て 例 を方解石 に とる。方解石 は立方体 と異 り歪ん だ結晶、即 ち光学的異方性 を持つ結晶の代表であ るが、方解石が作 る二重像 は子供達に とっては驚 異の的であ り、それは子供 に とって宝 であった。 この二重像 は、入射光がSn
e
llの法則 に従 う正常 光 と、従 わない異常光の2
つのベ ク トルに分け ら れ る為 におこる。 この現象 を、互 に直交す る光 の 二成分 が夫々異 る屈折率n。とn。に分 かれ るこ と によって結像 された と考 え、複屈折(
Bi
r
e
f
r
i
n
g
e
n-ce)と一呼ぶ(9・20・21)。 コラー ゲンやDNA
分子の よう に、分子 内原子配列 自体 に光学的異方性があ るた めに起 こる複屈折 を固有複屈折 とい う(図3
)。 こ れに対 して もともと複屈折性 を持 たないたん 白質 などの ミセルが、分子配向 して規則正 しい微細構 造 を作 った時、分子配向度や ミセル相互間の距経、 ミセル と媒体 または溶媒 との屈折率 の差等か ら生 起す る複屈折性 を構造複屈折 とい う。生体 内微細 構造 では、横紋筋や微小管 で構築 された分裂紡錘 体 (図2)が この例 であ る。 この場合、構造 と溶 媒の屈折率差が無 くなる と構造複屈折 は失われ る が 22)、DNA
な どの固有複屈折 は媒体 の屈折率 の 変動 とは無関係 であ り、媒体 の屈折率 を変 えるこ とで両者 を識別 で きる (図4)
(9・21)。 このほかに流動複屈折、構造の慎みや 曲げモー メン トに よって起 る歪み複屈折 、流晶等が知 られ てい る。複屈折性 は生命現象 と深 く関わってお り、 細胞骨格 の構築、細胞運動、細胞流動等には複屈 折性が しば しば単独 に、 あるいは複数の要 因 とな って開通 し、発現 してい る場合が 多い。DNA
は分子 自体 強い固有複屈折性 を持 ち、偏 光係数 (n。-n。)は10 2、負の複屈折性 を示す。 それに も拘 らず、真核生物の染 色体 は全 く複屈折 性 を示 さない。 これは保護物質 であ る塩基性 たん 白質の ヒス トンが、DNA
分子 を染 色体 内でば ら ば らにほ ぐしたためにお こる分子配向の乱れが複 屈折性 を相殺 した結果 であろ うと推測 されて きた。 しか し、分馳 した染色体 を誘電率 の低 いアル コー ル中で引張 ると、強い、 国有複屈折が発現す る。 これはDNA
分子が並 んだ、或いは無理 に配向さ れた為 と説明 されて きた。 しか し、複屈折性 を示 さぬ真核細胞の染色体 は長い間謎の ま ,放置 され ていた。 細 胞分 裂 と核 分 裂 装 置 図2
に示す ように、核分裂装置 は真核 細胞が有 糸分裂 を行 う時に構築 され る一過性 の細胞骨格で あって、複製 されたDNA
の束である染色体 を赤 道面に配列 し、染色体分 経、染色体輸送 に直接関 与す る細胞器官であ る。 中心体 と星糸か らなる両 極 の星状体 と、中央の染色体 を含 む紡錘体が主な 構造 であ り、一般に光学的異方性 を示すか ら(18,22) 高感度高解像偏光顕微鏡 で容易 に識別 で きる(9・25)0 これは観察光 の波長領域 よ りず っ と小 さな次元で の分子配向や微小管の配列 によって生ず る構造複 屈折に よる(22)。ウニ受精卵の核分裂装置は、骨組 み となっている微小管たん 白質、チュー ブ リンの 誘電率 を下 げ るか、 または疎水性 を高め るなどの 操作 で安定化 し、細胞外-取だす こ とが で きる。 この分離紡錘体の横 断面 を電子顕微鏡で観察す る と、紡錘体 は両極間のに配列 された約5,000本の微 小管で構成 されてい るこ とが明 らかになる。従 っ て紡錘体が示す構造複屈折は、微小管の配列 と微 小管数 を正確 に反影 してお り、 その動態は一見安 定 にみえなが ら、実 は細胞の生理条件の僅か な変 動 に鋭敏 に反応す る動的平衡系であ ることが明か になった(12・13・23)。 紡錘体微小管は明かに染色体輸送に直接関係す る重要 な骨組みであ り、核毒や分裂抑止剤 と反応 して脱重合す ると染 色体運動 は停 止す る(24)。微小図2 コシデ カウニ (Mespiliaglobulus)受精 卵の第1分裂期 におけ る紡錘体 の構築 と脱構築の過程 と、そ れに対応 した複屈折性の消長 を示す。配向微小管数 を反影す る紡錘体 の構造複屈折 は明かに認め られ る が、染色体 は全 く複屈折性 を示 さない.連続写真 の各右下段の数字 は媒精後C7)発生時間 を示す。偏光顕 微鏡像。右下段の スケールの1目盛 は10/Jm。
103-104 長野大学紀要 第14巻第1号 1992 図3 典型的なバ ッタの成熟精子。精子頭部は精子染色体 内に束ね られたDNA分子の量 と比例 した強い負 の固有複屈折 を示す。 これに対 して9+2のパ ター ンをとる微小管 を基本構造 とす る精子鞭毛は、正の 構造複屈折 を示す。偏光顕微鏡像。 (井上 ・佐藤、'66よ り)0 管の再構 築 を誘 導す る と、染 色体 輸送 は再 開 され 明か に変化 を受 け る筈 なの で あ るが、 それが複 屈 る。 折性 の変化 として表 われて こない。真核 細 胞 の染 一 見安 定 にみ え る染 色体 も、核毒 投 与 に よ り、 色体 は、 多量 のDNAを含 む に も拘 らず 、 ど うし
て も固有複屈折 を示 さないのである。 精 子染色体 ところが例外があった。精子染色体である。精 子形態の記載 で浩翰 な業績 を残 した
Re
t
z
i
us
(16)は、 精子 を 4つの基本型に分け、その うちの下等生物 に共通な精子 を "原始型"精子 とよんだ。 しか し 精子のかたちの複雑化 と進化 とは必ず しも対応せ ず、非複屈折性の原始型は水 中に卵 と精子 を放 出 して体外受精 を行 う種頬に広 く分布す る。一方、 体 内受精や特殊 な生殖様式 を持つ動物では、複雑 な形態 をもつ精子が多 く認め られるか ら、多様 な 精子形態は生殖様式に深 く依存 し、 またその変化 に伴 う卵保護屑の分化や組織の違 い等 も原因 とな っているのであろう(I,4)。但 し精子の多様性につい ての十分 な説明は未だなされていない。 生殖細胞の原細胞は、発生のかな り早い時期 に 体細胞か ら分化 して くる。 そ して何 回かの分裂 と 成熟 を繰返 した後に、雄性体 に形成 され る精母細 胞は、減数分裂 とよばれる連続2
回の分裂 を経て、 染色体数が半減 した4個 の精細胞 となる。染色体 の複製は、例外 を除いて第1
減数分裂の前期 で終 り、第2
減数分裂お よびそれに引続 く精子形成過 程 を通 じてDNA
の複製は行 われない。但 し部分 的なターン・オーバーはあるとい う(7)。従 って個々 の精細胞に配分 されたDNA
基幹の遺伝情報群は、 増減す ることな くその ま 、精子頭部へ凝縮 され、 頑 丈 に 保 護 さ れ て ゆ く。Co
ur
ot
(4)やFawc
e
t
t
等(6,7)は、精子の形態変化 を、1) 球状の精細胞核 の形成、2) 精核の伸長又は横方向への凝縮、3) 精子の形成の3つに分 けて考 える。特に図3に示 す ように、バ ッタ等の直辺頬の針状精子頭部形成 を取上げ、精子形成過程 を通 じて精子1個当 りのDNA
量は全 く変化 しない という顕微測光の事実 と、核質の規則的凝縮 を最大のイベ ン トとして考 えている。 精子頭部の形成は、精子染 色体の顕著 な凝縮 と、 代謝か らみて も完全に不活性 で酵素消化 を受つけ ない高度に安定な構造の構築 とい う点で際立って いる。例 えばイカやバ ッタの精子では、頭部の乾 燥重量の殆ん ど4
0%
がDNA
で占め られるか ら、 頭部の光学的密度、即 ち屈折率は高 くな り、微細 構造の解像は不可能になる。 しか も、成熟につれ て負の固有複屈折が急速に増す。紫外線吸収二色セツ 性やⅩ線回折像の解析か ら、 イカやバ ッタの精子 頭部の複屈折性は、DNA
(染色体)がほ ゞ長軸 方向に規則正 しく配列 し、半結 晶状構造 となって いる事 が強 く示唆 され る(10・11)。但 し、扇平又は球状 の非複屈折性の精子で も、やは り規則正 しく凝縮 された クロマチンの存在が予想 される(7)0 カマ ドウマの仲 間 であ るc
avec
r
i
c
ke
t
の針状 精子 (図4)の頭部には、 ジクザ ク状に配列 した 多数の固有複屈折性のマイクロ ドメインが認め ら れ る。 この微細構造の中にDNA
が どの ような結 晶体 となっているかにつ いて、井上 と佐藤(10・19)は 高感度 ・高解像偏光顕微鏡 と紫外偏光顕微照射法 とを組合せ、理論 と実験による偏光解析か ら、次 のような結果 をえた。1
) Ca
vec
r
i
c
ke
t
の精頭部 には約2
5
0
個 の負の複 屈折性マイクロ ドメインがあ り、 これが精子核 の 主構造 を形成 している。2
) DNA
ゲル と等 しい強複屈折性のマ イクロ ド メイン群は、精子長軸に対 して±1
0
9の方位角 を持 って配列 されている。 3) 紫外線スペ ク トルの吸収の方向性の解析結果 か ら、各マイクロ ドメインは主軸に対 して±1
5
0の 傾 き角 を持つDNA
の束で構成 されてい る。従 っ て全マ イクロ ドメインは、+2
5
0
、+50
、-5
0、-2
5
0の4
成分 で作 られているはずである。Cavec
r
i
c
ke
t
の精子の巾は0.
8
〟m
、計算 されたDNA
の束の外径は1
5
0
nm
、ピッチ角は±1
00、複 屈折量の測定値か ら、 2つのマイ クロ ドメは対 を 作 っていると考 えられ る。 これか ら、実 際の精子 核構造 は、 2
本の2
重 コイルの重ね合せ になって ハズ いる筈 である。唯 しこれ丈では充分ではない。二 重 コイルの重ね合わせ を少 し押 しつぶ し、断面が 楕円になるようにす ると、すべての測定値が満足 され る。図4B
は、偏光解析か ら理詰めで作図さ れた"
Cavec
vi
c
ke
t
の精子頭部内におけ るDNA
分子配列模型"である。 このモデルは理論計算値 とも良 く-致 したく10,ll,19)。 では、実際の精子染色体の配列パ ター ンは どう なっているのだろうか ? 精子形成過程 を通 じて 凝縮 され、箱詰めにされた精子染色体 は、マ イク ロ ドメインの整然たる配列か ら考 える と、当然規 則正 しい配列 をとる可能性が大 きい。従 って染色- 1
0
5-106 長野大学紀要 第14巻 第1号 1992
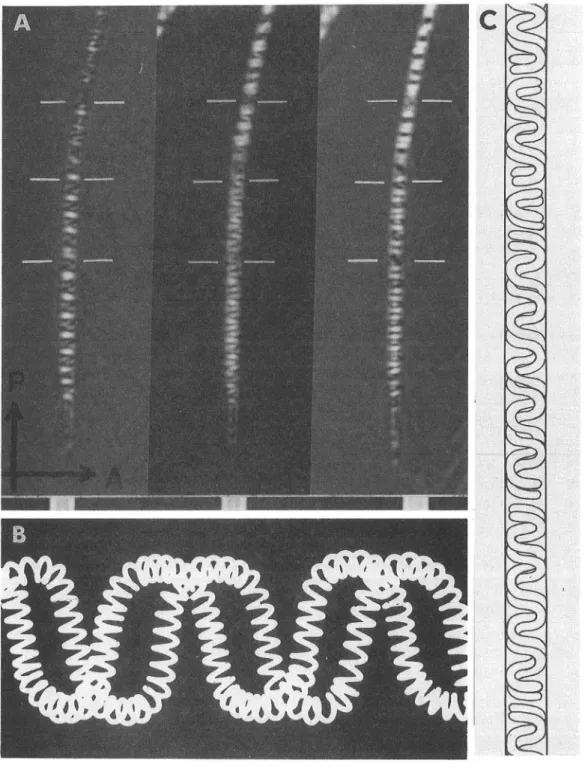
図4 直遡頬 カマ ドウマの仲間であ るCaveCricket(Ceuthophilusnigricans)の成熟精 子の頭部 をダイメヒモ チルサルフオキサ イ ド(DM SO)で液浸 し、膜、微小管等に由来す る構造複屈折 を消去す ると、紐状 となった精子染色体 の基幹構造 であ るラセ ンに巻 いたDNAの分 子配列が明瞭に示 され る。Aの3枚 の 写真 は同一精子の頭部 を補償板 の補償角 を少 しづつ変 えることで精子染色体 の切 れ 目を明かに した組写 真。 Bは偏光解析か らえ られた精子染色体 の2本撚 りになったDNAの2重 ラセン構 造のモデル。 断面 は楕 円 となるよ うに少 し押 しつぶ してある。Cは、偏光解析 の結果か ら明かにされ た精子染色体 の タン デム配列の模様。精子染色体 は決 して雑然 とつめ こまれた構造ではない。一定の順序 に従 って、整然 と 配列 されていると考 えられ る。
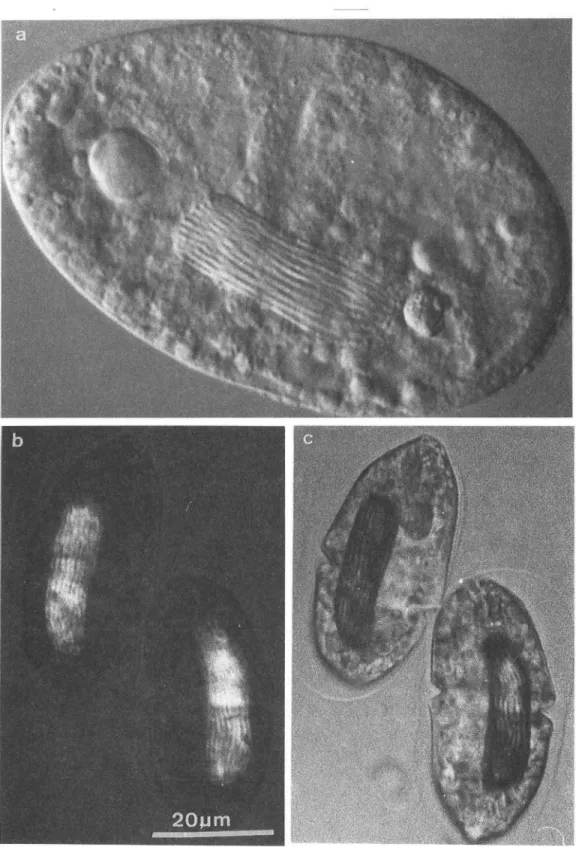
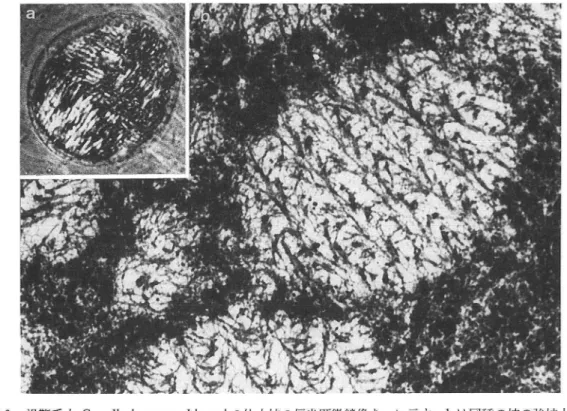
図5 分裂 中の渦鞭毛 虫Gyrodinium sugashimaniの棒状に並 んだ染色体群。 この類 は典 型的 な核 内核分裂 を行 い、決 して中期赤道板 を形成す るこ とはないが、複製 された娘染色体群 は紡錘体微小管によって両 極 に分 配 され る。aは微分干渉位相差像 bとCは偏光顕微鏡像 であって、補償板 を回転 し、染色体 の 強い正 の複屈折 を白お よび黒の コン トラス トで示す。胴体 中央の くびれは、分裂割構面 であ る。
7-108 長野大学紀要 第14巻 第1号 1992 体 の一部分 であ るマ イ クロ ドメインをつ な ぎ合せ た画像構築 か ら、常 に精 子頭部 の同 じ位 置に切 れ 目が で き、定 まった長 さの撚糸構造が定位 置に再 現 され るこ とが明 らかになった。つ ま り
、 2
本撚 りの クロマ チ ンか らな る精 子染色体 は、精 子頭部 に タンデムに並べ られてお り、順序 も定 まってい る と結論 で きる。 それ を図4C
に示 した。 渦 鞭 毛 虫類 の 染 色 体 若 い精 細胞の染色糸や 、真核細胞の有糸分裂 時 に形成 され る染 色体 が複屈折性 を示 さないこ とは、 既 に指摘 しておいた。 そ して、染色体 内ではDN A分 子 は配 向せ ずにほ ぐれ た状態 になってお り、 その隙間 をヒス トン等の塩 基性 たん 白質 が埋め て い るため に、DNAの 固有複屈折が相殺 されてい るのだ と説明 しておいた(17)。 しか しDNAが染色 体 の基幹構 造 であ る以上、 どうして も納得 がゆか ない。 これが永 い間、頭 に こび りつ いて離 れぬ疑 問であ った。 ところが、複屈折性 の染色体 を持つ生物 が見近 かにいたのであ る。 まさに燈 台 もと暗 Lであった。 それ は 海 産 又 は 汽 水 域 産 の 渦鞭 毛 虫 類 (Di no-flagellate)の休 止核 お よび分裂核 の染色体 であ っ た(3)。 図5に名古屋大学理 学部 附属 臨海実験所付 近 で採集 したGy710dinium sugashimani(故Jean Cachon教 授 に よって新種 記載) の強複屈 折性 の 染色体 を示す。 図5aは休 止核染色体 を微分干渉 顕微鏡像 で、図5b、 Cは分裂期 の染 色体 を偏光 顕微鏡像 で 白又 は黒の コン トラス トで示す。染色 体 の 複 屈 折 は、染 色 体 の 長 軸 に 対 して 正 で あ る。(2
・
3)
。
これは大変重要 なこ とであ り、DNA分 子 の固有複屈折 の符号 は分 子長軸 に対 して負 であ るか ら、渦鞭毛 虫類 の各染 色体 は、長軸方 向へ し っか りとコイルに巻 いたDNAで構 成 されてい る と考 えなければ な らないO偏光係数 (n。- n。)は+
2×10 2と計算 され、純粋 のDNAゲルの値 と 等 しい。 また、顕微測光 のデー タ も、 また干渉法 や 溶媒の屈折率 を変 え る方法 で も偏光係数 は変 ら ないか ら、 ヒス トンや塩基性 たん 白質 は殆 ん ど存 在 しない と考 え られ る。Herzog等(8)やRizzo等(17) も、細胞化学的 に渦鞭毛 虫類 の始原型種 の核 には 塩 基性 たん 白質 は無 いか ら、Gyrodinium等 の原 始 的渦鞭毛 虫類 の染色体 は、 原核生物 であ るバ ク テ リアのnucleoidに比肩 すべ き構 造 であ り、 その図 6 渦鞭毛虫Gy710dimiumsugashimαniの休止核の偏光顕微鏡像 をaに示すo bは同種の核の強拡大電子
意味 で渦鞭毛 虫類 は中間生物 として取扱 われば な らぬ と主張す る。
斑 6
は、G.
sugtzshimaniの分離核 と電子顕微鏡 像 で あ るが、染色体 はDNAの単一 又は2重 コイ ル では ないか とい う従 来 の考 え(5・15)、又 は解 釈 と 全 く一致 しない像 であ り、や は りDNAの束が染 色体 の長軸 に対 して コイル として巻 かれてい る と み るべ きであろ う。 そ して、塩 基性 たん 白質以外 のマ トリックスの存在 を予測 させ る像 で もあ る。 現在世 界 で飼育 ・培養 されて い る渦鞭毛 虫類 は 数 十種 あ るが、 その うちの数種 類 は フランスで培 養 されてい る。 その一種 の電子顕微 鏡像 と正 の複 屈折性 を示 す核 の偏光像 か ら、
Bo
ul
i
ga
n
d
ら(2,14・26) は、 この よ うな正 の固有複屈折 を示 す染 色体 は、 r T ■■■E,I. t t 図7 渦鞭毛虫Ceratiumfurcaの半ば脱穀 した休止核 と染色体 を示す。 bおよび Cは脱穀 した休止核の押 しつぶ し標本 を微分干渉顕微鏡像 (b)、偏光顕微鏡像 で示す。 この頬の染色体は コイル様 に巻 いてお り、コイルの方向によって正又は負の複屈折性が示 されている。下段右の図は屈折率楕円であってNeは 異常光 を長軸 を表わす。下端のスケールの 1目盛は10〝m。 - 109-110 長野大学紀要 第14巻第1号 1992
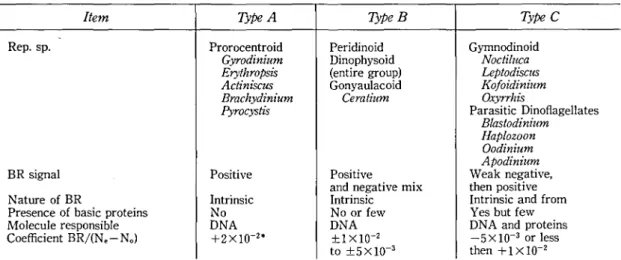
TABLE 1.l Comparisonbetweenthe3typesofchromosomesindinoflageHates.
Item
T
jPeA TyPeBT
y
J
vC
Rep.sp. BRsignal Prorocentroid Peridinoid Gymnodinoid Gyrodinium Dinophysoid Noctiluca ErythroPsis (entiregroup) LeptodisclLS Actinなcus Gonyaulacoid KofoidinilEm Brachydz'nium Jyo叩 ぬPositive IPoanntCednsriintriesatvgieciaumtiVemix WetPaⅠnhetApoHBkOxOorrnpoaaianknJstpldisrrsidittiniooicanscDihizedtniioontgaum-siniuVでdftenum1Voe爪ar,omgellates NatureofBR Ⅰntrinsic
Presenceofbasicproteins No Noorfew Yesbutfew Mo】ecuⅠeresponsible DNA DNA DNAandproteins
表1 染色体の複屈折を指標にすると、渦鞭毛虫矧 ま3群に大別されるのであるが、形態学的に組立てられ た系統樹 とは一致 しない, しか し私達は複屈折性からみて、渦鞭毛虫類はA型- C型 というパターン をとりながら進化 したのではないかと考えている タタ 長軸方向に折 り畳 まれたDNAの液 晶であろ うと 推論 した。面 白い仮設であ り、彼は頑 固に液晶説 を信奉 しているのだが、仮 りにそ うだ とす ると、 短 い細胞周期 で急速 に増殖す るこの生物の染色体 DNAの複製は期待 で きない。Cachonと私 は、 Gyrodiniumの個 々の染色体 は、や は りバ クテ リ アの nucleoidと対 比 され るべ き構 造 ではな いか と考 えている。 そ うであ る とす るな ら、てDNA の複製は個 々の染色体毎に独 自に行 われ、複製が 一区切 りつけば中期核板 を形成す ることな く、ず るず ると両極-移動す る核 内核分裂 とい う独特の 分裂様式が説明で きるのである川。 渦 鞭 毛 虫類 の 系 譜 染 色体 の複屈折性、換言すれば裸 のDNAで作 られ た染色体 の発見は、私達 を採集可能 な総 ての プ ラ ン ク トン型の dinaflagellatesのサーベ イ-と駆 りたてた。火がついたのであ る。 そ して渦鞭 毛 虫類は、染色体の複屈折性 を指標 に とる と、大 別 して3群 に分 け られ ることが わか った。即 ち、
A)
正 の強い固有複屈折 を持 ち、塩基性 たん 白質 を欠 くグループ、 B) 正 ・負の固有複屈折 が混在 し、かつ少量の塩 基性 たん 白質 を持つ グループ (図 7)、C)
通常 の後生生物の よ うに染色体 は殆ん ど複屈 折性 を示 さず、かつ ヒス トン系の塩基性 たん 白質(Cachon,Sato,Cachon,Sato,'89)
を相 当量含む グルーフで ある。
表 1に示す TypeA が始原型であ り、 この類の あるものが液晶型の染色体 を持つ であろ う可能性 は否定 で きない。Rep.sp.は各 グループ に属す る 代 表 種 を、BR signalは 複 屈 折 性 の 符 号 を、 NatureofBRは複屈折性の起源、即 ちDNA 自 身によるのか、 またはたん 白質の混在 に よる構造 複屈折が附加 されてい るか否かにつ いて を、そ し て Coefficient
BR/
(
Ne
-
No
)
は偏 光係数、す なわ ちDNAや塩基性 たん 白質の量 と配向の度合 を指 す。 この類別 に よる渦鞭毛 虫類の系譜は、 これ迄 の 形態に重点 を置いた進化の系統樹 か ら大 き くはず れ、最 も進化 している と考 え られ て きたグループ がA
グループ とC
グループであ り、進化 していな い と考 えられた、系統樹の下段の グループが実 はC
グループであった りす る。 この研究は今 も継続 中であ り、 日本 とフランスの協力研究 とい う形 を とり続け るこ とになろ う。 おわ リに 裸 のDNAか ら成 る染色体 の発 見は、私達 に大 きなインパ ク トを与 えた。30年間抱 き続けた疑問 に解答が与 え られたか らであ る。 同時に この発見 は新 たな疑問 を生む。を持つ に もか ゝわ らず、その核 内核分裂 は有糸分 裂型である。電子顕微鏡像 も、蛍光抗体染色法 も、 ともに染色体 の輸送 は微小管に よって行 われてい るこ とを示 している。 これはチュプ リン分子が核 内に存在す るこ と、有糸分裂 が行 われ る事 を意味 す る。 もともと、運動器官 として
2
本 の鞭毛 を持 ち、 それを巧みに操 りなが ら運動す る非細胞性 と も言 える生物 であ り、 中には レッキ とした レンズ と光感覚色素 を備 えた桂頬 もある。 しか るに、他 の重要 な細胞骨格 たん 白質 であ るア ク トミオシン や 中間径フィラメン トは存在 しない。代 りにア ク チ ンよ り細 い微細繊維が、細胞運動 に深 く関係 し てい る。ア クチ ンや 中間径 フィラメン トの遺伝子 は初めか ら無か ったのか、それ とも失 われたのか、 また、進化 の途 中で一体何がお こったのか、甚だ 興 味 をそ 、る。 何 よ りも、染色体が裸 のDNAでで きてい ると ナゾ い う事実は、見逃すわけにはいかない大 きな謎 で あ る。染色体 を保護 してい ると考 えられ るヒス ト ンは、渦鞭毛 虫類の進化の後期 になって獲得 され たのか、それ ともAグループでは一旦存在 してい た ヒス トン遺伝 子が不活性化 したのか、疑問が次 々 と湧 き出て くる。Gyrodinium とその仲 間 は、実 は赤 潮 (Ocean bloom)の原因になる原生生物 である。裸 のDNA 群 の複製は速 く、増殖率 は極 めて高い。か って第 3紀 に Gyrodinium の ような渦鞭毛 虫類が大発生 し、100万年 のオー ダー で巨大 な死骸の堆積 を海底 に作 り、それが石油へ と変質 したのではなか ろ う か ? 石油の無機生成説には無理があ り、石油 は や は り化石燃料 とすべ きであろ う。 とも角夢 を持 たせ る可能性 を秘めた生物群 であ り、将来裸 のD NAか らな る染色体がその謎 を解 く鍵 とな るであ ろ うと考 えている。 (さ とう ひでみ 教授) 1992. 4.14受理) 参考 文献
1.Baccetti,B.a Afzelius,B_A.Thebiologyof spermcell.MomographsinDev.Biol.,S.Kager,
Basel(1976)
2.Bouligand.Y.,Soyer,M.0.&Puiseux-Dao,S. Lastructurefibrillaireetl'orientationdeschr
o-mosomeschezlesdinoflagelles.Chromosoma,24; 25ト287.(1968)
3.Cachon,∫.,Sato,H.,Cachon,M.& Sato,Y. Analysis by polarizing microscopy of chromosomalstructureamong dinoflagellates anditspbylogeneticinvolvement.Biol.ofthe Ce11.65;51-60.(1989)
4.Courot,M.,Hochereau-deRevies,M-T.& Ortavant,R.Spermatogenesis. In;Thetestis (ed.byJohnson,A.D.,Comes,W.R.& Van-demark,N.L.),Vol.Ⅰ.p.339-432. Academic Press,New York.(1970)
5.Dodge,∫.Chromosome structure in
Dyno-pbycear.I.Thespiralchromonema.Arch.Mi -krobio1.,45;46-47.(1963)
6.Fawcett,D.W.The mammalian spem at -ozoon.Dev,Bio1.,44;394-436.(1975)
7.Fawcett,D.W.& Bedford,∫.M.(eds.)The spermatozoonmaturation. Motility,surface propertiesandcomparativeaspects. Urban&
Schwarzenberg,Bergel.(1979)
8.Herzog,M.& Soyer,M.0.Aspects par -ticuliersde la chromatine desprotistes'di no-flagelles.Biol.Cell,35;64a.(1979)
9.Inouさ,S.Videomicroscopy.Plenum Press,
New York.(1986)
10.Inoue,S.& Sato,H.ArrangementofDNAin livingsperm.A byophysicalanalysis. Science, 126;1122-1124.(1962)
ll.井上信也 ・佐藤英美、精子核内のDNA分子配列 - 生体超微細構造解析のための光学的アプロー チ。生物物理学講座、(日本生物物理学会編)、vol.
ⅠIl"遺伝情報m"、p.151-22.(1966)
12.Inoue,S.
&
Sato,H.Cellmotilityoflabile associationofmolecules:Thenatureofmitotic spindlefibersandtheirrolein chromosome movement.J.Gen_Physio1.,50;2591292.(1967) 13.Itoh,T.∫.& Sato,H.Theeffectofduteriumoxide(2H20)Onthepolymerizationoftubulinin
vitro.Biochem.Biophys.Acta,800;21-27.(1984) 14.Livolant,F.Lastructurecristallineliquidede l'ADNinvivoetinvit710.TheseDoctoratEtat, Univ.ParisVI.(1984)
1-112 長野大学紀要 第14巻 第1号 1992
15.Livolant,F.
&
Bouligand,Y.Newobserv a-tionsonthetwistedarrangementofspreaddino・ flagellatechromosomes.Chromosoma,80;97-118.(1980)
16.Retzius,G.BiologischeUntersuchungen,
NeueFolge.Vol.XII,ⅩⅠV,ⅩⅤⅠ,GustavFischer Verlag,一ena.(1906-1911)
17.Rizzo,P
_
J.& Nooden,L.D.Isolationand chemicalcompositionofdinoflagellatenuclei.∫. Protozoo1.,20;666-672.(1979)18.Sato,H.Roleofspindlemicrotubulesforthe anaphasechromosomemovementsinfertilized seaurchin eggs.CellDiff"2・,345-348.(1982) 19.佐藤英美、精子の形成 と構造 :染色体 の配列、臨 床科学、20:620-631.(1984) 20.佐藤 英 美、偏光 に よる構 造 別定、生体 の科 学、 39、別刷 "細胞光学特性測定法一、530-533. (1988) 21.佐藤英美、偏光 で見たい ものだけ選 んで見 る (偏 光顕微鏡)、 日本分光学会、測定法 シ リー ズ (宝谷紘 一 ・木下一彦編)、21、31-51、学会 出版 セ ンタ-、 東京 (1991)
22.Sato,H.,Ellis,G.W.&Inoue,S.Microtubule
origin ofmitoticspindleform birefringeilCe. DemonstrationoftheapplicabilityofWiener's equation.∫.CellBiol.67;501-517.(1975) 23.Sato,H.& Bryan,∫.Thethermodynamicsof
molecularassociationinthemitoticspindlewith orwithoutheavywater(D20).Zool.Sci.,5;73
3-742.(1988)
24.Sato,H.,Kobayashi,
A.
&
Itoh,∫.T.Molec u-larbasisofphysicalandchemicalprobesfor spindleassembly(Review article).CellStruct. andFunct.,14;1-34.(1989)25.Sato,H.,Takenaka,H.& Niboshi,T.An attempttoconstructsuper-sensitivepolarizing microscopewithVideo-enhancedsystem. Acta Histochem.Cytochem.,24;3431347.(1991)
26.Stzelecka,T.E.Davidson,M.W.,&Rill,R.Ⅰ. MultipleliquidcrystalphasesofDNA athigh concentrations_Nature,331 :457-460.(1988) 27.Zemike,F.Thewavetheoryofmicroscopic
imageformation. In;ConceptsofClassical Optics(ed.Strong∫.).p.525-536.W.班.Freeman Co.,SamFrancisco.(1958)