虚血性脳障害時の
LRP1 の病態生理学的変化と furin
阻害薬による神経保護の可能性
Pathophysiological changes in LRP1 during ischemic brain injury and
a possibility of neuroprotection by furin inhibitor
略語表
本文中では以下の略語を用いる。
ALS amyotrophic lateral sclerosis
AMPA α-amino-3-hydroxy-5-methyl-4-isoxazolepropionic acid ANOVA analysis of variance
Apo apolipoprotein
APS ammonium peroxodisulfate BBB blood-brain barrier
BCA bicinchoninic acid
BDNF brain derived neurotrophic factor BSA bovine serum albumin
DIV days in vitro
DMEM Dulbecco’s Modified Eagle Medium EGF epidermal growth factor
ERK extracellular signal-regulated kinase FBS fetal bovine serum
GFAP glial fibrillary acidic protein
GOMED Golgi membrane associated degradation GSK glycogen synthase kinase
HBSS Hank’s balanced salt solution ICD intracellular domain
IFN interferon
LDL low density lipoprotein
LP glia-derived apoE-containing lipoprotein LRP1 LDL receptor-related protein 1
MAPK mitogen-activated protein kinase
MCAO/R middle cerebral artery occlusion/reperfusion MMP matrix metalloproteinase
NMDA N-methyl-D-aspartate
PAGE poly-acrylamide gel electrophoresis PBS phosphate buffered saline
PC proprotein convertase
PC-LP apo E-containing lipoprotein with phosphatidylcholine PCSK9 proprotein convertase subtilisin kexin type 9
PLC phospholipase
PMS 5-methylphenazinium methyl sulfate PSD postsynaptic density
PVDF polyvinylidene difluoride QOL quality of life
RGC retinal ganglion cell SD Sprague-Dawley SDS sodium dodecyl sulfate SEM standard error of the mean TBS tris buffered saline
TEMED tetramethylethylenediamine TGN trans-Golgi network
tPA tissue plasminogen activator Trk tropomyosin receptor kinase TTC 2,3,5-triphenyl tetrazolium chloride VLDL very low density lipoprotein
... 26 1-2-11 脳梗塞時の LDL 受容体ファミリーの受容体の変化 ... 27 1-2-12 脳梗塞時の LRP1 の変化 ... 28 1-3 考察 ... 29 第 2 章 虚血性神経障害時の LRP1 の変化に対する furin の役割 ... 32 2-1 実験材料・方法 ... 33 2-1-1 実験動物 ... 33 2-1-2 培養液の調製および 培養プレートのコーティング ... 33 2-1-3 ラット大脳皮質神経細胞の初代培養 ... 33 2-1-4 大脳皮質神経細胞の障害の誘導 ... 33 2-1-5 大脳皮質神経細胞の蛍光免疫染色 ... 33 2-1-6 大脳皮質神経細胞の western blot 用サンプル調製 ... 33 2-1-7 ラット脳梗塞モデルの作製 ... 34 2-1-8 ラット脳梗塞モデルの western blot 用サンプルの調製 ... 34 2-1-9 脳梗塞モデルの脳組織凍結ブロックおよび薄切切片の作製... 34 2-1-10 脳梗塞モデルの蛍光免疫染色 ... 34 2-1-11 Western blot ... 35 2-1-12 統計 ... 35 2-2 実験結果 ... 36 2-2-1 NMDA 処置後 LDL 受容体ファミリーの受容体の変化に対する calpain 阻害薬 の影響 ... 36 2-2-2 NMDA 処置後 LRP1 の変化に対する calpain 阻害薬の影響 ... 37 2-2-3 NMDA 処置後 LRP1 の変化に対する γ-secretase 阻害薬の影響 ... 38 2-2-4 NMDA 処置後 LRP1 の変化に対する MMP 阻害薬の影響 ... 39 2-2-5 NMDA 処置後 LRP1 の変化に対する furin 阻害薬の検討 ... 40 2-2-6 NMDA 処置後の LRP1 の局在変化に対する furin 阻害薬の影響 ... 41 2-2-7 NMDA 処置後の LRP1 および TGN46 の局在とそれに対する furin 阻害薬の影 響 ... 42
3-1-4 大脳皮質神経細胞の障害の誘導 ... 49 3-1-5 大脳皮質神経細胞の細胞内 Ca2+ 濃度変化の解析 ... 49 3-1-6 大脳皮質神経細胞の生存率の解析 ... 49 3-1-7 大脳皮質神経細胞の western blot 用サンプル調製 ... 49 3-1-8 大脳皮質神経細胞の calpain 活性の解析 ... 49 3-1-9 ラット脳梗塞モデルの作製 ... 50 3-1-10 Furin 阻害薬の脳室内投与 ... 50 3-1-11 脳梗塞モデルの虚血領域の解析 ... 50 3-1-12 脳梗塞モデルの western blot 用サンプルの調製 ... 50 3-1-13 Western blot 法 ... 50 3-1-14 統計 ... 50 3-2 実験結果 ... 51 3-2-1 NMDA 処置による生存率低下に対する γ-secretase、MMP および PCSK9 阻害薬 の影響 ... 51 3-2-2 NMDA 処置による生存率低下に対する furin 阻害薬の影響 ... 52
3-2-3 NMDA 処置による calpain 活性化に対する calpain 阻害薬の影響 ... 53
3-2-4 NMDA 処置による生存率低下に対する calpain 阻害薬の影響 ... 54
3-2-5 NMDA 処置による calpain 活性化に対する furin 阻害薬 の影響 ... 55
3-2-6 NMDA 処置による calpain 活性化に対する furin および calpain 阻害薬 の影響 ... 56
3-2-7 NMDA 処置による細胞内 Ca2+ 濃度変化に対する furin 阻害薬 の影響... 57
1
緒言
脳血管疾患は日本の死因別死亡者数第 4 位の疾患である 1)。脳血管疾患には脳梗塞、脳内出血、
くも膜下出血などが含まれ、なかでも脳梗塞は要介護認定を受ける原因の中で最も多くの患者数
を有する疾患である 2)。このように脳梗塞は、その後遺症による患者の quality of life (QOL) の低
下だけでなく、患者家族の負担増大も大きな問題となる疾患である。 脳梗塞は血栓や塞栓により脳動脈が閉塞することで発症し、脳血管疾患による死亡者数の約 6 割を占める 1)。血管閉塞により、その支配領域が虚血状態に陥ると酸化ストレスや栄養因子欠乏、 興奮性神経伝達物質であるグルタミン酸の大量遊離が誘発される。虚血によるグルタミン酸遊離 は神経細胞内への過剰な Ca2+ 流入を誘導し、Ca2+ 依存性のプロテインキナーゼやプロテアーゼ の活性化に起因する神経細胞死を惹起する 3)。中枢神経系の機能は、主に神経細胞やグリア細胞 によって発揮される。そのうち、グリア細胞は神経細胞を物理的・生理的に支える役割だけでな く、シナプス形成や神経伝達に関わるなど、神経機能に積極的に働きかけている 4)。 脳は生体内で最も豊富にコレステロールを含む器官であり、神経細胞とグリア細胞の間で緊密 なコレステロール代謝の調節が行われている 5-9)。この中枢神経系における脂質代謝、特にコレス テロール代謝のバランスの崩壊は、アルツハイマー病やニーマンピック病C 型、ハンチントン病 などの神経変性疾患と密接に関わっている 10-12)。神経細胞は脂質の供給をグリア細胞からのリポ タンパク質による輸送に依存しており、中枢神経系ではアポリポタンパク質 E を含むリポタンパ ク質 (LP) がその中心的役割を担っている。その LP を細胞内に取り込むのが low density lipoprotein (LDL) 受容体ファミリーに属する受容体である。これらの受容体は中枢神経系に豊富 に発現しており、細胞内に脂質を取り込むだけでなくシグナル受容体としても働くことが明らか となっている。また、発生時の細胞遊走や増殖にも重要な働きを担っている 13)。これまでの研究 で、このLDL 受容体ファミリーのひとつである LDL receptor-related protein 1 (LRP1) は初代培養 網膜神経節細胞 (retinal ganglion cell : RGC) で細胞内シグナル伝達を誘導し、栄養因子欠乏および
グルタミン酸誘導性神経細胞死を抑制することが明らかとなっている14)。この神経保護効果は、
緑内障モデル動物に対しても観察されている。しかしながら、脳における LRP1 の病態生理学的
役割は明らかになっていない。
プロタンパク質転換酵素 (proprotein convertase : PC) は Ca2+ 依存性のセリンプロテアーゼで、
不活性なタンパク質前駆体を活性体へと変化させる 15)。PC は PC1、PC2、furin (proprotein
convertase subtilisin kexin 3 : PCSK3)、PC4、PC5、paired basic amino acid cleaving enzyme 4 (PACE4)、 PC7、subtilisin kexin isozyme 1 (SKI-1) および PCSK9 を含む 9 種類で構成されており、ホルモン
や神経ペプチド、成長因子など、様々な因子の成熟化に関与している 15,16)。PC の中でも furin は
主にトランスゴルジネットワーク (trans Golgi network : TGN) に存在し、特定のアミノ酸配列 (Arg-X-X-Arg) を標的として切断する。特に、中枢神経系では脳由来神経栄養因子 (brain derived neurotrophic factor : BDNF) や LRP1 の成熟化に関与する 17-20)。また furin は、がんや心血管疾患、
神経変性疾患などの様々な病態に関与することが報告されているが 21,22)、脳血管疾患での役割は
3
第
1 章 大脳皮質神経細胞の虚血性神経障害による LRP1 の変化
グルタミン酸は記憶の形成などに関与する内因性神経伝達物質であるが、虚血などの刺激によ り大量に放出されると興奮性の神経障害を誘導する。イオンチャネル共役型グルタミン酸受容体 はリガンドの違いにより -amino-3-hydroxy-5-methyl-4-isoxazolepropionic acid (AMPA) 受容体、カ イニン酸受容体および N-methyl-D-aspartate (NMDA) 受容体に分類され、特に NMDA 受容体は脳 虚血などの急性神経疾患のみならず、てんかん、アルツハイマー病、筋萎縮性側索硬化症 (amyotrophic lateral sclerosis : ALS) またはハンチントン病などの慢性神経疾患の細胞死にも寄与し
ていると考えられている 23)。
リポタンパク質の受容体として働く LDL 受容体ファミリーは中枢神経系に広く分布しており、
共通の構造を持つ 7 種のコアメンバー(LDLR、VLDLR、ApoER2/LRP8、LRP4/MEGF7、LRP1、
LRP1b および megalin/LRP2)といくつかの共通構造を欠く 3 種のサブメンバー(LRP5、LRP6 お
よび SorLA/LR11) で構成されている。これらの LDL 受容体ファミリーに属するコアメンバー
の受容体は、リガンド結合ドメイン、epidermal growth factor (EGF) 前駆体相同ドメイン、細胞膜
貫通ドメインおよびシグナル伝達やエンドサイトーシス誘起を司る NPxY モチーフを有してい
る (Figure 1)。
4
当研究室は、LP の結合による LRP1 の活性化が、網膜神経節細胞の細胞死に対し保護効果を
発揮することを報告している。この神経保護機構のひとつとして、LRP1 を介した、ホスホリパー
ゼC1、プロテインキナーゼ C および glycogen synthase kinase 3 (GSK3) の順次的な活性化が
5 1-1 実験材料・方法 実験動物 本研究では胎生 16 日目および生後 3 日齢の Sprague-Dawley (SD) 系ラットおよび 7 週齢の SD 系雄性ラット (日本エスエルシー株式会社) を使用した。ラットには飼料と水を自由に摂取さ せ、恒温 (23±1℃)、恒湿 (55±5 %)、定時照明 [12 時間明所 (7:00~19:00)、12 時間暗所 (19:00~7:00)]の人工環境下で飼育した。動物の飼育および実験操作は、東京薬科大学動物実験規定 およびガイドライン “大学などにおける動物実験” (昭和 62 年第 141 号) を遵守して行われた。 本実験計画は、東京薬科大学動物実験委員会で審査および承認を受けた後に実施された。 培養液の調製および培養プレートのコーティング
Neurobasal medium (Invitrogen, 21103-049) 49 mL に 対 し 、 B27 serum-free supplement (Gibco,17504044) 1 mL、100 mM L-glutamine (SIGMA-Aldrich, G8540) 250 L、penicillin-streptomycin (Wako, 168-23191) 500 L を添加し、大脳皮質神経細胞用の培養液 (以下 culture medium) とした。 2.4 mM CaCl2 (Sigma-Aldrich, C5670)、20 mM 4-(2-hydroxyethyl)-l-piperazineethane sulfonic acid
(Sigma-Aldrich, H4034)/Hank’s Balanced Salt Solution (Gibco, 14175-095) を pH 8.0 に調製し、サン プル調製用の緩衝液 (以下 HBSS buffer) とした。
24-well plate (Falcon, 353043) を poly-D-lysine (Wako, 168-19041) でコーティングした。1%
poly-D-lysine となるように滅菌水で調製しプレートに添加後、1 時間インキュベートした。その後、滅
6 ラット大脳皮質神経細胞の初代培養
胎生 16 日目のラット胎児から Dulbecco’s-phosphate buffered saline (PBS) (Wako, 045-29795) 中
で大脳皮質を摘出した。その後、外科用メスを用いてガラスシャーレ上で約 300 回細切し、PBS
5 mL で回収後、静置した。5 分間静置した後に上清を除去し、2.5% trypsin (Invitrogen, 15090-046) 500 L、0.4% DNase Ⅰ (Worthington, 2139) 400 L / PBS 2.5 mL を加え、37 ℃ で 20~25 分間 イ ンキュベートした。その後、上清を取り除き 20% fetal bovine serum (FBS) (Gibco, 26140) / Neurobasal medium 3 mL を加え、パスツールピペットで 5 回ピペッティングし、組織を懸濁した (必要に応 じて DNase を 100 L 加えた)。懸濁液を 2~3 分間 静置し上清を回収する操作を 3 回繰り返し た後、回収した上清を 120 x g、 5 分間 遠心した。得られたペレットを culture medium 5 mL で 懸濁し、血球計算盤を用いて細胞数を計測した。コーティングしたプレートに 2 x 105 cells / well となるように細胞を播種した。37 ℃、 5 % CO2 条件下で培養し、4~6 時間後に culture medium を全量入れ替えた。その後 3~4 日ごとに culture medium を半量交換し、培養した。 大脳皮質神経細胞の障害の誘導
初代培養後 10 日目に NMDA および グルタミン酸処置を行った。Culture medium を全量除き、 HBSS buffer で 37 ℃、 5% CO2 条件下にて 15 分間 2 回洗浄を行った。続いて HBSS buffer で
各濃度に調製した NMDA (Sigma-Aldrich, M3262) およびグルタミン酸 (Sigma-Aldrich, G5889) 15
分間 インキュベートし NMDA 処置とした。その後、culture medium に交換した。洗浄から処置
後までの操作を treatment とした。また、treatment による細胞への障害を考慮するため 10 日目
まで同様に培養し treatment を行わなかったものを untreated control とした。
大脳皮質神経細胞の細胞内 Ca2+ 濃度変化の解析
大脳皮質神経細胞の培養液を Fluo-8 AM (AAT Bioquest) 3 L/culture medium 2 mL に調製した培
養液と入れ替え、37 ℃、30 分間インキュベートベートした。その後 treatment 時の蛍光輝度の変
7 大脳皮質神経細胞の生存率の解析
ミトコンドリア活性を指標とする XTT 法により細胞生存率を解析した。1 mg/mL 3’-[1(-phenyl
amino carbonyl-3,4-tetrazolium]-bis (4-methoxy-6-nitro) benzene sulfonic acid hydrate (XTT) sodium salt (Fluka, X4251)/PBS : 1.5 mg/mL 5-methylphenazimium methyl sulfate (PMS) (Wako,166-09211) / PBS = 200 : 1 で混合し、XTT-PMS 溶液とした。この XTT-PMS 溶液を culture medium と 1 : 3 で混合 し反応液とした。Treatment から 24 時間後に culture medium を除き、XTT 反応液を入れ 37 ℃、 5% CO2 条件下にて 1 時間インキュベートした。この反応液の 450 nm の吸光度を 96-well plate
(アズワン, 1-6776-03)で、EMax Plus プレートリーダー (Molecular Devices) を用いて測定した。未
反応の反応液の吸光度を blank として用いた。 大脳皮質神経細胞の蛍光免疫染色 Poly-D-lysine コーティングしたカバーガラス上で 10 日間培養した大脳皮質神経細胞に treatment を行い、処置 0.5、1 および 4 時間後に蛍光免疫染色を行った。 細胞を PBS で 5 分間 2 回洗浄し、4% paraformaldehyde (Wako, 163-20145) で 10 分間 室温で 固定した。その後、PBS で 5 分間 2 回洗浄し、0.2% Triton-X-100 (MP Biomedicals,194854) / PBS にて 10 分間 室温で浸透処理を行った。次に、 10% goat serum (Merck Millipore, S26) または donkey serum 、1% bovine serum albumin (Sigma-Aldrich , A 7030)、0.2% Triton X-100 / PBS (blocking buffer) 中で 30 分間ブロッキングした。その後、1 次抗体を添加して 4 ℃ で一晩、または室温 1 時間インキュベートした。
インキュベート後、PBS で 5 分間 3 回洗浄し、2 次抗体を加え室温 1 時間インキュベートし
た。その後、0.05 % Hoechst 33342 (DOJINDO, H342) / PBS で 5 分間 3 回洗浄し、Fluoromount / Plus (Diagnostic BioSystems, K048) を用いてスライドガラス上に封入した。
試料は蛍光顕微鏡 (OLYMPUS, IX71) または共焦点顕微鏡 (OLYMPUS, FV1000) で観察し、画 像解析ソフトウェア MetaMorph (Molecular Devices)を用いて解析した。
8 Table 1. Antibodies for immunofluorescence staining
Primary antibody Dilution Secondary antibody (1:200) Microscope Mouse anti-NeuN (Abcam,
ab104224) 1:1000
Alexa Fluor 488 goat anti-mouse
(Invitrogen, A11029) Fluorescence microscope Rabbit anti-GFAP (Abcam,
ab7260) 1:5000
Alexa Fluor 594 goat anti-rabbit (Invitrogen, A11037)
Rabbit anti-LRP1
(Epitomics, 2703-1) 1:2000
Alexa Fluor 488 donkey anti-rabbit
(Life technologies, A21206) Confocal laser scanning microscope Goat anti-LRP1 (CⅡ)
(R&D systems, AF2368) 1:100
9
大脳皮質神経細胞の western blot 用サンプル調製
Treatment 後に 細胞を PBS で 2 回洗浄し、sample buffer [5% 2-mercaptoethanol (Wako, 137-06862), 0.125M Tris-HCl (pH 6.8), 4% sodium dodecyl sulfate (SDS), 20% glycerol, 0.06% bromophenol blue] で細胞を回収した。24-well plate の 1 well に対して 50 L の sample buffer で 2 度回収し た (total 100 L)。回収した細胞は 95 ℃ で 5 分間タンパク質を可溶化し western blot 用サンプ ルとした。 ラット大脳皮質グリア細胞の初代培養とリポタンパク質を含む培養上清の作製 グリア細胞の初代培養は Hayashi ら (2012) の方法に従って行った。生後 3 日のラットを断頭 し、実体顕微鏡下で大脳皮質を摘出した。その後、外科用メスを用いてガラスシャーレ上で約 300 回細切し、PBS 5 mL で回収後、静置した。5 分間静置した後に上清を除去し、2.5% trypsin 500 L、4% DNase 400 L / PBS 2.5 mL を加え、37 ℃で 20〜25 分間インキュベーションした。上清 を除き、2 ml のグリア細胞培養液 [(10% heat-inactivated FBS / Dulbecco’s Modified Eagle Medium (DMEM; Gibco 11995-065)] を加えパスツールピペットで丁寧に懸濁した。懸濁液を 2~3 分間静置
し、上清を回収する操作を 3 回繰り返したのち、回収した細胞懸濁液を 120 x g、5 分間した。上
清を除き、得られたペレットにグリア細胞培養液を 5 mL 加え懸濁した。その後 40 m セルスト
レイナー (Falcon) で濾過し、作製した細胞懸濁液を 1 脳 / flask に調製後、75 cm2 flask (Falcon)
10 人工再構成アポリポタンパク質 E 含有リポタンパク質(PC-LP)の作製 PC-LP の作製は Hayashi ら (2012) の方法に従って行った。1-パルミトイル-2-オレイル-グリセ ルホスホコリン (SIGMA-Aldrich, P3017)、ヒトリコンビナントアポリポタンパク質 E (モル比 100:1) を含むリポタンパク質を作製した。2.17 mg 1-パルミトイル-2-オレイル-グリセルホスホコ リンをクロロホルムに溶解し、窒素ガス下で乾燥した。400 l の 10 mM TBS (10 mM Tris、0.9% NaCl) を加え、15 分ごとに攪拌しながら氷上で 2 時間インキュベーションした。その後 100 l の 15 mg/ml コール酸ナトリウムを加え、さらに 2 時間インキュベーションした。この混合液に 1 mg ヒトリコンビナントアポ E を加え氷上で 1 時間インキュベーションした。次に 100 mg バ イオビーズ (Bio-Rad, 152-3920) を加え、攪拌しながら 3 時間 4 ℃でインキュベーションした。 インキュベーション後、0.45 m フィルターを通し、ビーズを除いたリポタンパク質混合液を回収 し、4 ℃で保存した。 LP および PC-LP の単離 上記で作製したグリア細胞由来リポタンパク質、PC-LP を含む培養上清またはリポタンパク質 混合液を、1.3 g/ml、1.2 g/ml、1.1 g/ml、1.006 g/ml スクロース溶液に積層し、160,000×g、48 時 間、4 ℃で超遠心した。遠心後、上層より 1 ml ずつ回収し、アポリポタンパク質 E を含む 4~ 6 番目の画分を採取した。アポ E の分布は western blot 法で確認した。画分 4~6 を混合し、50 kDa molecular weight cut-off Amicon Ultra filter (Millipore, UFC905008) を用いて濃縮後、BCA 法に
てタンパク質定量を行った。単離・濃縮したアポリポタンパク質 E 含有リポタンパク質 (LP) お
11 ラット網膜神経節細胞の初代培養
生後 3 日目のラット新生児から PBS 中で眼球を取り出し、網膜を摘出した。20 眼分の網膜を
10 mL PBS に回収後、165 unit papain を加え、37 ℃で 30 分間インキュベートした。上清を除去 し 、0.15% trypsin inhibitor、0.15% bovine serum albumin (BSA) および rabbit anti-macrophage antiserum / minimum essential medium (Gibco, 12360-038) を加え、パスツールピペットで組織を懸濁 した。懸濁液を 120 x g、7 分間遠心後、上清を除去し、0.02% BSA / minimum essential medium に
懸濁した。その後、goat anti-rabbit IgG をコーティングしたディッシュ上で、室温で 40 分間イン
キュベーションした。ディッシュ上の細胞懸濁液をNitex® membrane (SEFAR, NY20-HC) で濾過
後、goat anti-mouse IgM および mouse-anti-Thy1.1 antibody をコーティングしたディッシュ上で、
室温で 35 分間インキュベーションした。ディッシュを HBSS buffer で 8~10 回洗浄し、ディッ
シュに結合した細胞を0.125% trypsin で 37 ℃、10 分間インキュベートした。次に、20% FBS / Neurobasal medium を加え、細胞を回収した。細胞懸濁液を 120 x g、12 分間遠心後、上清を除去 し、RGC culture medium [1 mM glutamine, 5 µg/mL insulin, 60 µg/mL N-acetylcysteine, 62 ng/mL progesterone, 16 µg/mL putrescine, 40 ng/mL sodium selenite, 0.1 mg/mL BSA, 40 ng/mL triiodothyronine, 0.1 mg/mL transferrin, 1 mM sodium pyruvate, 2 % B27 supplement (Invitrogen), 10 µM forskolin, 50 ng/mL brain-derived neurotrophic factor (BDNF; PeproTech), 50 ng/mL ciliary neurotrophic factor (CNTF; PeproTech) and 50 ng/mL basic fibroblast growth factor (bFGF; PeproTech) / Neurobasal medium] に懸濁 した。細胞数を計測後、poly-D-lysine および laminin (SIGMA-Aldrich, L7264) でコーティングした 96-well plate に播種した。
網膜神経節細胞の障害の誘導
初代培養 14~21 日目にグルタミン酸処理を行った。RGC culture medium を全量除き、HBSS buffer で 37 ℃、5%CO2 条件下で 15 分間 2 回洗浄を行った。続いて HBSS buffer で調製した
300 µM グルタミン酸 (SIGMA-Aldrich, G5889) で 2 時間インキュベートし、グルタミン酸処理 とした。その後、RGC culture medium から B27 supplement、forskolin、BDNF、CNTF、bFGF を除
12 網膜神経節細胞の生存率の解析 Treatment 後の細胞を PBS で 2 回洗浄し、4% paraformaldehyde で 15 分間室温で固定した。 その後、PBS で 1 回洗浄し、0.05% Hoechst 33342 / PBS で 15 分間室温で核染色を行った。PBS で 1 回洗浄後、蛍光顕微鏡下で画像解析ソフト MetaMorph を用いて自動解析を行った。細胞生 存率の算出は、全細胞数のうち、核の凝集または断片化像を示した細胞(死細胞)数を除いた細 胞の割合で算出した。 網膜神経節細胞の western blot 用サンプル調製
Treatment 後に細胞を PBS で 2 回洗浄し、sample buffer で回収した。96-well plate の1well に 対して、20 µL の sample buffer で回収した。回収した細胞は、95 ℃ で 5 分間タンパク質を可溶 化し western blot 用サンプルとした。
ラット脳梗塞モデルの作製
ラット右中大脳動脈閉塞再灌流 (middle cerebral artery occlusion reperfusion : MCAO/R) モデルを
作製した。モデル の作製は Longa ら (1989) の方法を一部改良して行った。5% イソフルラン吸 入麻酔薬 (Pfizer) で麻酔導入し、その後 2.5% イソフルランで術中の麻酔を維持した。頸部正中 線を切開し、右総頸動脈、右外頸動脈および右翼突口蓋動脈を周囲結合組織から剥離した。続い て外頸動脈を切開し栓子 (先端部約φ200 µm) を内頸動脈に向けて約 20 mm 挿入し、中大脳動脈 起始部を閉塞した。絹糸で外頸動脈を固定し、麻酔を解除し覚醒させた。中大脳動脈閉塞 90 分 後に再度麻酔し、栓子を除去し血液を再灌流させた。再灌流 24 時間後にサンプルとして回収し、 以下の実験を行った。
13 脳梗塞モデルの虚血領域の解析
ミトコンドリア活性を指標とする TTC 法により梗塞領域の割合を解析した。MCAO/R 24 時
間後に断頭し、脳を摘出した。摘出した脳を嗅球から 2 mm 厚で冠状切片を作製し、計 7 切片を
得た。得られた切片を 0.2 % 2,3,5-triphenyl tetrazolium chloride (TTC) (Wako, 207-05832)/生理食塩水
で 10 分間反応させた。TTC 中のテトラゾリウム塩がミトコンドリアのコハク酸脱水素酵素など
で還元され赤色のテトラゾリウム塩を生じるため、非虚血 (生存) 領域は赤く、虚血 (非生存) 領
域は白く観察される。Image J を用いて梗塞領域の割合を測定した。
Figure 3. Photograph of TTC-stained sections from MCAO/R-operated rats
脳梗塞モデルの western blot 用サンプルの調製
MCAO/R 24 時間後に 1-1-17 のように脳組織を摘出し、脳切片のうち 3 切片目を western blot
用サンプルとした。この切片を下記に示すように、虚血領域 (Ischemic area : I) および 非虚血領
域 (Non-ischemic area : N) に分画し、左右の計 4 ヶ所をサンプルとして回収した。組織を 10% protease inhibitor (Sigma Aldrich, 30496700) / Lysis buffer [1% Triton X-100, 0.1% sodium deoxycholate (Wako, 190-08313)、1% ethylenediaminetetraacetic acid (DOJINDO, 345-01865) 37.2 mg / 50 mM TBS (pH 7.4)] 中で超音波処理し、タンパク質定量を行った。サンプルを Lysis buffer で希釈し、等量 の 10 % 2-mercaptoethanol / sample buffer を加え、最終濃度 1 mg / mL となるよう調製した。これ を 95 ℃、5 min 加熱処理し、-20 ℃で凍結保存した。
14 Western blot 法
検出するタンパク質の分子量に適したゲルを作製し、サンプルをアプライ後 SDS-PAGE buffer
(25mM Tris, 190 mM glycine, 3.5 mM SDS) 中で電気泳動を行った。使用した試薬の組成を Table 2 に、gel 濃度およびその組成を Table 3 および 4 に示す。電気泳動後、blotting buffer [25 mM Tris, 192 mM glycine, 20% methanol (Wako, 25183-80)] 中 で 、 polyvinylidenedifluoride (PVDF) 膜 (Immobilon-P, IPVH00010) に転写した。PVDF 膜は使用前に methanol で活性化させた。
泳動および転写の条件を以下に示す。
電気泳動 : stacking gel : 20 mA/gel、separating gel : 30mA/gel、RT 転写 : 80 V、4℃、90 分間
LRP1 α-chain の検出は以下の条件で行った。 電気泳動 : 10mA / gel、4℃、5 時間
転写 : 0.05% SDS / blotting buffer、30 V、4 ℃、16 時間
Table 2. Composition of buffer for western blot
30% Acrylamide 29.2 g Acrylamide (Wako, 011-08015), 0.8% bis-acrylamide (Wako, 138-08173), in 100 mL MilliQ water
Separating gel buffer 90.8 g Tris, 2 g SDS in 500 mL MilliQ water, pH 8.8 Stacking gel buffer 6.06 g Tris, 0.4 g SDS in 100 mL MilliQ water, pH 6.8
Table 3. Composition of stacking gel for western blot
3% 5%
MilliQ water (mL) 2.24 2
30% Acrylamide (L) 354 600
Stacking gel buffer (L) 888
10% Ammonium peroxodisulfate (APS) (Wako, 012-08023) (L) 56 N', N', N', N',-Tetramethylethylenediamide (TEMED)
15 Table 4. Composition of separating gel for western blot
5% 7.5% 10% 12.5% 15% MilliQ water (mL) 4..18 3.62 2.99 2.37 1.74 30% Acrylamide (mL) 1.32 1.88 2.51 3.13 3.76 Separating gel buffer (mL) 1.9
10% APS (L) 112
TEMED (L) 5
転写終了後、PVDF 膜を 10 mM Tris buffered saline with 0.1% Tween20 (TBST) (Wako, 166-21115) (TBST) で洗浄し、5% skim milk (Wako, 190-12865)/TBST で 1 時間 ブロッキングした。ブロッキ ング後、TBST で 5 分間 3 回洗浄し、1 次抗体を 4 ℃ で一晩反応させた。
1 次抗体を反応させた後、PVDF 膜を TBST で 5 分間 3 回洗浄し、対応する 2 次抗体 (1:5000)/5% skim milk/TBST を 1 時間室温で反応させた。その後 TBST で 5 分間 3 回洗浄し、 LuminoGraph Ⅱ (ATTO) でタンパク質を検出した。検出試薬は、ImmunoStar Basic (Wako, 291-75103)、ImmunoStar Zeta (Wako, 295-72404) または Super SignalTM West Femto (Thermo, 34095) を使
用した。得られた画像を画像解析用ソフトフェア CSAnalyser4 (ATTO) を用いて解析した。
同一の PVDF 膜を用いて他のタンパク質を解析するために、PVDF 膜をストリッピングバッフ
ァー for Western (ARPO SCIENCE, BW-6010) 中に 20 分間浸透し、1 次抗体および 2 次抗体を除
去した。その後 TBST で 5 分間 3 回洗浄し、ブロッキングを行った後、同様の工程でタンパク
質を検出した。
16 Table 5. Antibodies for western blot
Primary antibody Dilution Secondary antibody (1:5000) Mouse anti-β-actin (SIGMA-Aldrich, A5441) 1:10000 Goat anti mouse IgG
(Thermo, 31430) Mouse anti-GluN1 (BD Pharmingen, 556308) 1:500 Goat anti-mouse IgG
Rabbit anti-GluN2A (Merck Millipore, AB1555) 1:1000 Goat anti-rabbit IgG (Thermo, 31460)
Mouse anti-GluN2B (Invitrogen, 32-0700) 1:300 Goat anti-mouse IgG
Goat anti-LRP1 (CⅡ) (R&Dsystems, AF2368) 1:2000 Mouse anti-goat IgG (Thermo, 31400)
Rabbit anti-LRP1 (Epitomics, 2703-1) 1:5000 Goat anti-rabbit IgG Mouse anti-LR11 (BD, 611861) 1:1000 Mouse anti-goat IgG Rabbit anti-LDLR (Epitomics, 1956-1) 1:5000 Goat anti-rabbit IgG Goat anti-ApoER2 (Abcam, ab 51945) 1:1000 Goat anti-mouse IgG
統計
実験結果は全て平均値 ± 標準誤差 (standard error of the mean, SEM) で示した。
17 1-2 実験結果 1-2-1 初代培養大脳皮質神経細胞の細胞構成 初代培養後 10 日目の培養細胞の免疫染色像および陽性細胞の割合を Figure 5 に示す。 NeuN (神経細胞マーカー) 陽性細胞は 93.9 ± 2.1% 、GFAP (アストロサイトマーカー) 陽性細 胞は0.6 ± 2.1% であり、神経細胞の比率の高い培養条件であると考えられた。
Figure 5. (a) Cortical cells were fixed at 10 days in vitro (DIV) and stained for NeuN (green) and GFAP (red) and with Hoechst (blue). The scale bar represents 100 µm. (b) The number of NeuN- and GFAP-positive cells was counted. Results were expressed as the percentage of these cells among the total number of Hoechst-positive cells and as the means ± SE in 4 independent experiments.
18
1-2-2 初代培養大脳皮質神経細胞の NMDA 受容体発現量の経日的変化
培養細胞の NMDA 受容体サブユニット発現を western blot 法で解析した結果を Figure 6 に示 す。
NMDA 受容体を構成するサブユニットである GluN1、GluN2A、GluN2B はそれぞれ経日的に
増加し、10 日目、あるいは 12 日目に最大となった。この結果より、本研究では培養 10 日目の
細胞を用いて以降の実験を行った。
19
1-2-3 初代培養大脳皮質神経細胞の NMDA 処置による障害誘導
NMDA 処置後の細胞生存率を XTT assay により測定した結果を Figure 7 に示す。
NMDA 処置による細胞死誘導の検討を行った結果、濃度依存的に細胞生存率は低下し、NMDA 未処置と比較して 10 μM では 76.2 ± 4.7%、30 μM では 52.9 ± 6.8%、100 μM では 39 ± 5.2% で
あった (Fig.7a)。以降の実験は、約 50% の細胞死を誘導する 30 μM NMDA を使用した。
NMDA 処置 (30 μM) による生存率の低下 (44 ± 1.5%) は、NMDA 受容体アンタゴニストであ る MK801 (10 μM) によって抑制された (97.6 ± 9. 0%) (Figure 7b)。
Figure 7. (a) Effect of NMDA treatment on cell viability. Cortical neurons were incubated with NMDA at the indicated concentrations for 15 minutes. After 24 h of NMDA treatment, cell viability was determined by performing the XTT dye-reduction assay. The absorbance at 450 nm was measured, and the relative cell viability was expressed as the percentage of the absorbance at 450 nm of each treatment group against that of the untreated control group. Results are the means ± SE (n = 5 independent experiments). *Indicates a significant difference from the NMDA-untreated group (p < 0.05). (b) Effect of NMDA receptor antagonist on NMDA induced cell injury. Cell viability in cultures of 0 µM and 30 µM NMDA-treated cells without (white bars) or with (black bars) 10 µM MK801. The relative cell viability was expressed as the percentage of the absorbance at 450 nm of each treatment group against that of the untreated control group. Results are the means ± SE (n = 3 independent experiments). *Indicates a significant difference from the NMDA-untreated group (p < 0.05) and # indicates a significant difference from the NMDA-treated and MK801-untreated group (p < 0.05).
20 1-2-4 NMDA 処置による細胞内 Ca2+ 濃度変化 NMDA 処置後の細胞内 Ca2+ 濃度変化をカルシウム指示薬 Fluo-8AM を用いて測定した結果 を Figure 8 に示す。 細胞内 Ca2+ 濃度は NMDA 添加直後に上昇し、その上昇は NMDA 受容体アンタゴニストで ある MK801 によって抑制された。
21 1-2-5 NMDA 処置による生存率低下に対する LP の影響 LP 添加後の細胞生存率の変化について XTT assay で評価した結果を、Figure 9 に示す。 網膜神経節細胞 (RGC) ではグルタミン酸により低下した細胞生存率が、人工再構成 LP (PC-LP) および LP (0.1 μg/ml) によって回復した。しかしながら、初代培養大脳皮質神経細胞 (cortical neuron) では、0.01-1 μg/ml のいずれの濃度でも、LP は保護効果を示さなかった。
Figure 9. (a) Effect of glutamate treatment on cell viability of RGCs. RGCs were incubated with 300 µM glutamate for 2 hours. After 24 h of glutamate treatment, cell viability was determined by nuclear staining with Hoechst 33342. Shrunken or fragmented nuclei were counted as dead cells, and round/smooth nuclei were counted as healthy cells. Reconstituted ApoE-containing lipoproteins with phosphatidylcholine (PC-LP) or glia-derived ApoE-containing lipoproteins ((PC-LP) at 0.1 µg protein/ml were treated with glutamate. The relative cell viability was expressed as the percentage of healthy cells in the total number of cells stained by Hoechst 33342. Results are the means ± SE (n = 5 independent experiments). (b) Effect of NMDA treatment on cell viability of cortical neurons. Cortical neurons were incubated with 30 µM NMDA for 15 min. The relative cell viability was expressed as the percentage of the absorbance at 450 nm of each treatment group against that of the untreated control group. Results are the means ± SE (n = 4 independent experiments). *Indicates a significant difference from the control group (Con) (p < 0.05) and # indicates a significant difference from the glutamate-treated and LP untreated group (Glu-0) (p < 0.05).
22
1-2-6 NMDA 処置による LDL 受容体ファミリーの受容体の経時的変化
NMDA 処置 0、0.5、1、2 および 4 時間後の LR11、LDLR あるいは ApoER2 の発現量の経 時的変化について western blot 法で解析した結果を Figure 10 に示す。
LR11 は NMDA 処置による影響を受けなかった。LDLR および ApoER2 は NMDA 処置後、 経時的に減少する傾向を示した。
Figure 10. Effects of NMDA treatment on receptors of the LDL receptor family at 0, 0.5, 1, 2 and 4 hours after 0 µM (0) or 30 µM (30) NMDA treatment. Proteins from primary cultured cortical neurons were analyzed by western blotting with anti-LR11, anti-LDLR, anti-ApoER2 and anti-β-actin antibodies. Bands corresponding to LR11, LDLR, ApoER2 and β-actin were scanned, the scanned bands were normalized by untreated control on the same blot. β-Actin was used as a loading control. Results are the means ± SE (n = 6 independent experiments).
23 1-2-7 NMDA 処置による LRP1 の経時的変化
NMDA 処置 0、0.5、1、2 および 4 時間後の LRP1 の発現量の経時的変化について western blot 法で解析した結果を Figure 11 に示す。
LRP1 α-chain は、NMDA 処置後経時的に減少傾向を示した。-Chain から切り出される細胞内 ドメイン (intracellular domain : ICD) は NMDA 処置 30 分後から増加傾向を示し、1 時間以降に 著しく増加した。
24
1-2-8 NMDA 処置による LRP1 細胞内局在の経時的変化
NMDA 処置 0.5、1 および 4 時間後の LRP1 細胞内局在の経時的変化について蛍光免疫染色 法で観察した結果を Figure 12 に示す。
LRP1 α-chain および β-chain は細胞質および神経突起上に存在した。NMDA 処置により α-chain
は減少し、LRP1 ICD を含む β-chain は NMDA 処置により核周辺に集積する傾向が見られた。そ
の変化は 1 時間後から顕著に観察され、経時的に増加した。以降の実験は NMDA 処置 4 時間
後の変化を観察した。
25 1-2-9 NMDA 処置による LRP1 の変化
NMDA 処置 による LRP1 の発現量変化について western blot 法で解析した結果を Figure 13 に示す。
LRP1 を構成する α-chain、β-chain および β-chain から切り出される ICD の NMDA 処置によ る変化を検討した。α-Chain は NMDA 処置により減少し、β-chain は変化せず、ICD は増加した。
26
1-2-10 大脳皮質神経細胞および網膜神経節細胞のグルタミン酸処置による LRP1 の変化
大脳皮質神経細胞 および 網膜神経節細胞のグルタミン酸による LRP1 の変化について
western blot 法で解析した結果を Figure 14 に示す。
大脳皮質神経細胞は NMDA 処置後と同様に、グルタミン酸処置による ICD の増加が観察さ
れた。一方、LP により LRP1 を介した保護効果が発揮される網膜神経節細胞 (RGC) では、グル
タミン酸処置後に ICD は産生されなかった。
27
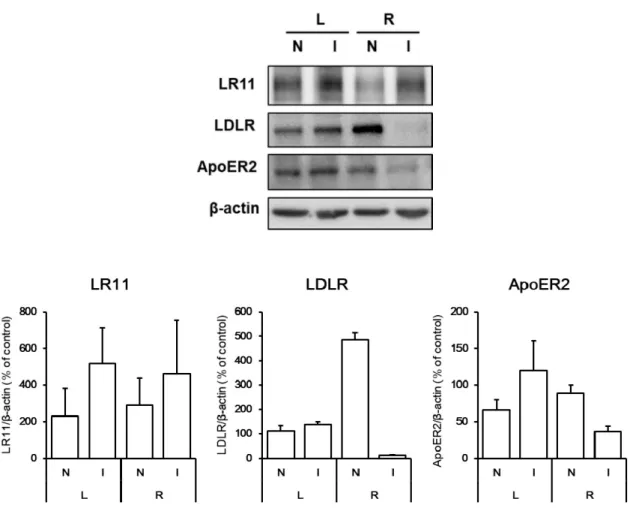
1-2-11 脳梗塞時の LDL 受容体ファミリーの受容体の変化
脳梗塞 4 時間後の LR11、LDLR および ApoER2 の発現量の変化について western blot 法で 解析した結果を Figure 15 に示す。
今回作製した右中大脳動脈閉塞・再灌流モデル (MCAO/R) では、再灌流 24 時間後に右脳大脳
皮質および線条体が梗塞領域となることが示されている。
再灌流 24 時間後の両側の大脳皮質の梗塞領域 (I) 、非梗塞領域 (N) を分画し、western blot 法
により解析したところ、LR11 は虚血性神経障害による影響を受けなかった。LDLR および
ApoER2 は虚血後大脳皮質梗塞領域 (R-I) で減少した。これらの変化は in vitro で初代培養大脳
皮質神経細胞に NMDA 処置を行った際の結果と同様であった。
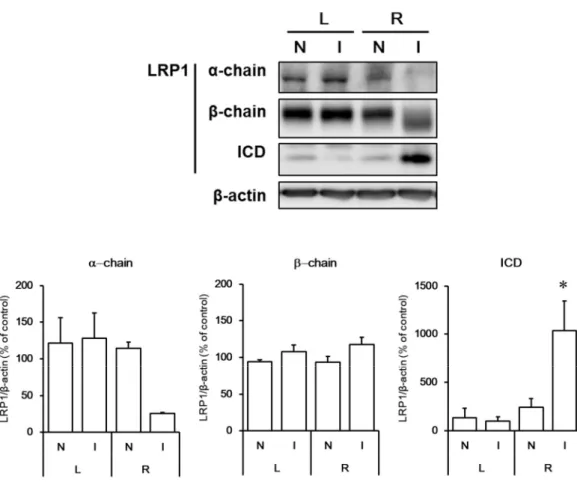
28 1-2-12 脳梗塞時の LRP1 の変化
ラット脳梗塞モデルの大脳皮質 LRP1 の変化について western blot 法によって解析した結果を
Figure 16 に示す。
再灌流 24 時間後の両側の大脳皮質の梗塞領域 (I) 、非梗塞領域 (N) を分画し、western blot 法
により LRP1 の解析を行ったところ、右脳梗塞領域で、in vitro と同様に α-chain の減少および
ICD の増加が観察された。
29 1-3 考察
本研究で用いた初代培養大脳皮質神経細胞の培養条件を蛍光免疫染色で検討した結果、神経細
胞マーカーである NeuN 陽性細胞の割合は 90% 以上であった。一方、主な混入細胞と考えられ
るアストロサイトのマーカー glial fibrillary acidic protein (GFAP) 陽性細胞の割合が 1% 未満であ ったことから、純度の高い神経細胞の培養条件になっていると考えられた。また、神経機能に重 要な役割を果たし、細胞死の誘導にも関わる NMDA 受容体のサブユニットの発現が培養 10 日 目に増加したことから、培養期間を通じて良好な培養条件が維持できたと考えられた。さらに NMDA 処置により細胞生存率が低下し、この低下は NMDA 受容体アンタゴニストである MK801 によって完全に抑制された。以上の結果より、NMDA 処置による神経細胞障害は NMDA 受容体を介して誘導されたことが示された。NMDA 受容体は Ca2+ を透過させることから 25)、こ の神経障害は Ca2+ 依存性であると考えられ、本研究では NMDA 処置による細胞内の急激な Ca2+ 濃度の上昇も確認した。このことは、神経細胞内で過剰な Ca2+ に応答したキナーゼやプロ テアーゼの異常な活性化の誘導を示唆した。 Hayashi らは初代培養網膜神経節細胞を使用した実験で、グルタミン酸誘導性神経細胞死に対 し、グリア細胞由来の LP が神経保護効果を発揮することを示した 24) 。しかしながら、本研究 では初代培養大脳皮質神経細胞死に対して、グリア細胞由来 LP は保護効果を示さなかった。こ のことは大脳皮質神経細胞で神経保護に必要なリポタンパク質受容体が存在していない、あるい は NMDA 障害によって保護機構が破綻している可能性が示された。大脳皮質神経細胞に少なく とも LR11、LDLR、ApoER2 および LRP1 が発現していることを確認し、とりわけ LDLR およ び ApoER2 は NMDA 処置後 4 時間目で減少傾向を示した。LDLR は脳を含むすべての組織に 発現しており、リポタンパク質の細胞内への取り込みを行っている。LDLR 欠損マウスで記憶学 習能力の低下が示されているものの 26)、LDLR の神経系での機能は未だ明らかにされていない。 これは、LDLR の神経系での機能が、他の受容体で代償できることに起因すると考えられている
27)。また ApoER2 欠損マウスは文脈的恐怖条件付け (contextual fear conditioning) と長期増強
(long-term potentiation : LTP) に障害をもたらすことから、シナプス可塑性に影響を与えると考えら
れている 28)。したがって、これら LDL 受容体の神経細胞での変化は神経障害後慢性期に惹起さ
れる記憶学習能力低下の初発要因となる可能性がある。
次に、NMDA 障害後の大脳皮質神経細胞の LRP1 の変化を検討した結果、細胞外ドメインであ
る α-chain が減少し、-chain から切り出される細胞内ドメイン (intracellular domain : ICD) が著
しく増加した。蛍光免疫染色法で各タンパク質の局在を検討した結果、通常、細胞全体に α-chain
および ICD を含む β-chain がスパイン様に観察されたが、NMDA 障害によって α-chain は神経
突起上で観察されなくなり、ICD は核の周辺に集積する傾向が観察された。LRP1 α-chain はリガ
ンド結合ドメインであることから、α-chain が切断されるとリガンドが結合した際の細胞内取り込
みやシグナル伝達が惹起されないと推察される。また、先に述べたように、ICD はシグナル伝達
やエンドサイトーシス誘起を司る NPxY モチーフを有しているため、この切断もまた、LRP1 の
30
ス/シグナル受容体で、神経細胞に多く発現している。例えば LRP1 は、神経細胞の後シナプス
部で NMDA 受容体と相互作用し、神経伝達を調節すると考えられている 29,30)。LRP1 のリガン
ド で あ る α2-マク ログロ ブリ ンや組 織型プ ラスミ ノーゲ ン活性 化因子 (tissue-plasminogen activator : t-PA) は、Src ファミリーキナーゼを介して tropomyosin receptor kinase (Trk) 受容体を活
性化し、神経栄養活性を持つ可能性が示されている 31)。さらに、神経細胞の LRP1 欠損は Akt リ ン酸化レベルの減少とカスパーゼの活性化を誘導し、アポトーシスを亢進する32)。神経細胞特異 的 LRP1 欠損マウスは、脳コレステロール量の減少および神経変性を誘導し、運動機能障害ある いは記憶障害を呈する 30,33)。これらの結果は、LRP1 が中枢神経系で重要な役割を果たしている ことを示している。したがって、LP が大脳皮質神経細胞で、保護効果を発揮できなかった理由の 一つとして、LRP1 が切断されることで惹起される α-chain の減少および細胞内断片である ICD の産生が LRP1 の機能不全を引き起こしている可能性がある。そこで、この大脳皮質神経細胞で 観察された LRP1 の切断が網膜神経節細胞でも起こるかを観察したところ、大脳皮質神経細胞で ICD が産生される条件では、網膜神経節細胞の LRP1 は変化しなかった。これらの本研究結果は、 大脳皮質神経細胞は NMDA 受容体を介した神経障害時に特異的な LRP1 切断機構を有し、かつ、 それが病態形成へ関与する可能性を初めて示したものである。 次に、脳梗塞モデル動物を用いて LRP1 の変化を検討した。その結果、虚血領域で LDLR お よび ApoER2 の著しい減少が確認された。さらに、初代培養大脳皮質神経細胞と同様に脳梗塞領
域の LRP1 α-chain は非梗塞領域と比較して減少し、ICD が著しく増加していた。In vivo 脳梗塞
31
32
第
2 章 虚血性神経障害時の LRP1 の変化に対する furin の役割
第 1 章では、脳の虚血性障害時に LRP1 の切断が起こることを明らかにした。
LRP1 は 600 kDa の前駆体タンパク質として合成され、TGN で furin によって、リガンド結合 ドメインを持つ 515 kDa の α-chain と、膜貫通ドメインおよび細胞内ドメイン(ICD) を含む 85 kDa の β-chain に切断され、この 2 つの chain は共有結合し、膜 1 回貫通型受容体として膜上
で機能する 36)。Furin は細胞内の TGN に存在し、神経成長因子や脳由来神経栄養因子などの前
駆体にも作用することで、その成熟化に関与するといわれている 17)。LRP1 に対しても同様に成
熟化に関与している。細胞膜上で機能する成熟した LRP1 は matrix metalloproteinase (MMP) や γ-secretase などの酵素によって段階的にさらに切断され α-chain の遊離および ICD の産生が誘導 される 37)。
細胞内で切り出された LRP1 ICD は核内に移行し、interferon (IFN) -γ の遺伝子発現の調節にか
33 2-1 実験材料・方法 2-1-1 実験動物 1-1-1 に準ずる 2-1-2 培養液の調製および 培養プレートのコーティング 1-1-2 に準ずる 2-1-3 ラット大脳皮質神経細胞の初代培養 1-1-3 に準ずる 2-1-4 大脳皮質神経細胞の障害の誘導 1-1-4 に準ずる
Furin 阻害薬として furin inhibitor 1 (Cayman chemical, 14965)、furin inhibitor 2 (SIGMA, SCP0148) および SSM3 (R&D SYSTEMS, 5253/10) を使用した。Calpain 阻害薬として calpeptin [calpain 1 および calpain 2 阻害薬 (Milllipore, 03-34-0051)]、-secretase 阻害薬として DAPT (Millipore, 565770)、MMP 阻害薬として GM6001 (Millipore, 3642069) を使用した。
2-1-5 大脳皮質神経細胞の蛍光免疫染色 1-1-7 に準ずる
使用した 1 次抗体および対応する 2 次抗体を Table 6 に示す。 Table 6. Antibodies for immunofluorescence staining
Primary antibody Dilution Secondary antibody (1:200) Microscope Rabbit anti-LRP1
(Epitomics, 2703-1) 1:2000
Alexa Fluor 488 donkey anti-rabbit
(Life technologies A21206) Confocal laser scanning microscope Goat anti-LRP1 (CⅡ)
(R&D systems, AF2368) 1:100
Alexa Fluor 594 donkey anti-goat (Life technologies, A11058)
34 2-1-7 ラット脳梗塞モデルの作製 1-1-15 に準ずる 2-1-8 ラット脳梗塞モデルの western blot 用サンプルの調製 1-1-17 に準ずる 2-1-9 脳梗塞モデルの脳組織凍結ブロックおよび薄切切片の作製 MCAO/R 24 時間後に灌流固定を行った。チオペンタール (0.1 mL / 100 g) (田辺三菱製薬株式会 社, ラボナール注射用 0.3 g) で麻酔し、ラットを仰臥位に固定し開胸した。その後、直ちに鉗子 で腹部大動脈を閉塞し、心尖部からヘパリンナトリウム (100 U / 0.1mL) (Nipro, 1182576040101) を 投与し、続いて、26 G 静注針 (Thermo Fisher) および Variable-Flow Peristaltic Pumps(流速 10 mL / min)[Thermo Fisher, pumpⅢ (model 3386)] を用いて 0.1 M PBS (pH 7.4, 4 ℃) (灌流液)50 mL
を灌流した。同時に右心耳を切開し、血液を排出させ、灌流液を流した後、4% paraformaldehyde
(PFA) 含有 0.1 M phosphate buffer (pH 7.4, PFA 溶液) 50 mL (10 mL / min) を灌流し脳を固定した。
灌流後、脳を摘出し、PFA 溶液中に 4 ℃で一晩 浸漬し後固定を行った。後固定後、10%、 20%
および 30% sucrose 含有 0.1 M phosphate buffer (sucrose buffer) に脳組織を浸漬し、置換した。脳
組織が沈降した後、30% sucrose buffer に 脳組織を浸漬した。その後、大脳半球を嗅球側前方より
3 部位 (0~4 mm, 4~8 mm, 8~12 mm) に分切し、凍結組織包埋剤 O.C.T. compound (サクラファイン
テックジャパン, 4583) を用いて液体窒素上で包埋後、脳組織凍結ブロックとした。CRYOSTAR
NX50 (Thermo Fisher) を用いて、脳組織凍結ブロックから 10 µm の 薄切切片を作製した。この 切片を MAS coating micro slide glass (Matsunami Glass Ind., Ltd.) に貼り付け、PBS で室温にて 30
分間洗浄した後風乾し、-20 ℃で保存した。
2-1-10 脳梗塞モデルの蛍光免疫染色
35 (DAKO, S3023) で封入した。
免疫染色後、共焦点顕微鏡 (OLYMPUS, FV1000-D IX81)で観察し、画像解析ソフトウェ ア
MetaMorph (Molecular Devices) を用いて解析した。
使用した抗体とその濃度を Table 7 に示す.
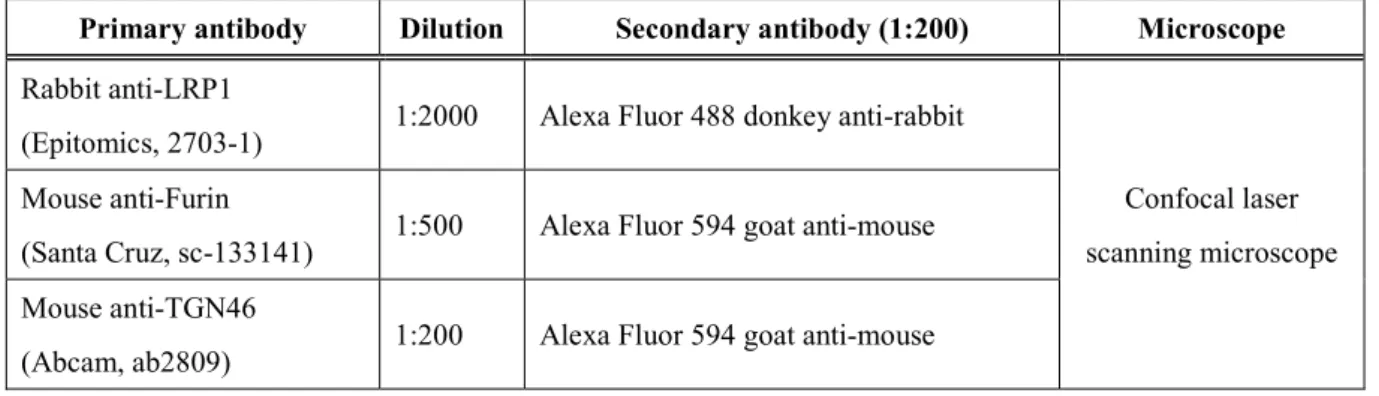
Table 7. Antibodies for immunofluorescence staining
Primary antibody Dilution Secondary antibody (1:200) Microscope Rabbit anti-LRP1
(Epitomics, 2703-1) 1:2000 Alexa Fluor 488 donkey anti-rabbit
Confocal laser scanning microscope Mouse anti-Furin
(Santa Cruz, sc-133141) 1:500 Alexa Fluor 594 goat anti-mouse Mouse anti-TGN46
(Abcam, ab2809) 1:200 Alexa Fluor 594 goat anti-mouse
2-1-11 Western blot 1-1-18 に準ずる。
使用した 1 次抗体および対応する 2 次抗体を Table 8 に示す。 Table 8. Antibodies for western blot
Primary antibody Dilution Secondary antibody (1:5000) Mouse anti-β-actin (SIGMA-Aldrich, A5441) 1:10000 Goat anti mouse IgG
Goat anti-LRP1 (C2) (R&Dsystems, AF2368) 1:2000 Mouse anti-goat IgG Rabbit anti-LRP1 (Epitomics, 2703-1) 1:5000 Goat anti-rabbit IgG Mouse anti-LR11 (BD, 611861) 1:1000 Mouse anti-goat IgG Rabbit anti-LDLR (Epitomics, 1956-1) 1:5000 Goat anti-rabbit IgG Goat anti-ApoER2 (Abcam, ab 51945) 1:1000 Goat anti-mouse IgG
36 2-2 実験結果
2-2-1 NMDA 処置後 LDL 受容体ファミリーの受容体の変化に対する calpain 阻害薬の影響
初代培養大脳皮質神経細胞での NMDA 処置 による LR11、LDLR および ApoER2 の変化に対
するcalpain 阻害薬の効果を検討した結果を、Figure 18 に示す。
NMDA 処置による LDLR および ApoER2 の減少が、calpain 阻害薬によって抑制された。 LR11 は NMDA 処置および calpain 阻害薬による影響を受けなかった。
37
2-2-2 NMDA 処置後 LRP1 の変化に対する calpain 阻害薬の影響
NMDA 処置 による LRP1 の変化に対する calpain 阻害薬の効果を検討した結果を、Figure 19 に示す。
NMDA 処置による LRP1 α-chain の減少および ICD の増加は calpain 阻害薬では抑制されなか った。
38
2-2-3 NMDA 処置後 LRP1 の変化に対する γ-secretase 阻害薬の影響
NMDA 処置による LRP1 の変化に対する γ-secretase 阻害薬の効果を検討した結果を Figure 20 に示す。
NMDA 処置による LRP1 α-chain の減少および ICD の増加は γ-secretase 阻害薬では抑制され なかった。
39
2-2-4 NMDA 処置後 LRP1 の変化に対する MMP 阻害薬の影響
NMDA 処置による LRP1 の変化に対する MMP 阻害薬の効果を検討した結果を Figure 21 に 示す。
NMDA 処置による LRP1 α-chain の減少および ICD の増加は MMP 阻害薬では抑制されなか った。
40
2-2-5 NMDA 処置後 LRP1 の変化に対する furin 阻害薬の検討
NMDA 処置による LRP1 の変化に対する furin 阻害薬の効果を検討した結果を Figure 22 に 示す。
Furin 阻害薬は、NMDA 処置による α-chain の減少および ICD の増加を抑制する傾向であっ た。
41
2-2-6 NMDA 処置後の LRP1 の局在変化に対する furin 阻害薬の影響
NMDA 処置による LRP1 の変化に対する furin 阻害薬の影響を蛍光免疫染色によって検討し た結果を Figure 23 に示す。
蛍光免疫染色の結果、western blot 法による結果と同様に、NMDA 処置による α-chain の減少
および ICD を含む β-chain の局在変化が furin 阻害薬によって抑制された。
42
2-2-7 NMDA 処置後の LRP1 および TGN46 の局在とそれに対する furin 阻害薬の影響 NMDA 処置後の LRP1 および TGN46 (トランスゴルジネットワークマーカー) の局在を、蛍光
免疫染色によって検討した結果を Figure 24 に示す。
NMDA 処置によって核周辺に凝集した LRP1 の ICD を含む β-chain は TGN46 と共局在し
た。また、furin 阻害薬によって、LRP1 および TGN 46 の核周辺への凝集は抑制された。
43
2-2-8 NMDA 処置後の LRP1 および furin の局在とそれに対する furin 阻害薬の影響
NMDA 処置後の LRP1 および LRP1 のプロセシングに関わる furin の局在を、蛍光免疫染色に よって検討した結果を Figure 25 に示す。
NMDA 処置によって核周辺に凝集した LRP1 の ICD を含む β-chain は furin と共局在した。
また、furin 阻害薬によって、LRP1 および furin の核周辺への凝集は抑制された。
44 2-2-9 脳梗塞時の LRP1 の局在変化
MCAO/R 後の LRP1 の局在を、蛍光免疫染色によって検討した結果を Figure 26 に示す。
右大脳皮質梗塞領域では核が変形し、さらに ICD を含む β-chain および TGN46 の共局在が左
脳と比較して多く観察された。
45 2-3 考察
本研究の NMDA による神経障害はカルシウム依存性であったことから、カルシウム依存性タ
ンパク質分解酵素の calpain がリポタンパク質受容体の切断に関与していると考えられた 25)。そ
こで NMDA 処置による大脳皮質神経細胞のリポタンパク質受容体の変化と、それに対する
calpain 阻害薬の効果を検討した。その結果、NMDA 処置による LDLR および ApoER2 の減少
は calpain 阻害薬によって抑制された。これに対し、NMDA 処置による LRP1 ICD の産生は
calpain 阻害薬による影響を受けなかった。次に、NMDA 処置による LRP1 の切断機序を明らか にするために、 furin、MMP および γ-secretase 阻害薬の影響を解析した。その結果、furin 阻害 薬が NMDA 処置後の α-chain の減少と ICD の産生を抑制した。LRP1 は γ-secretase によって
切断され、切り出された LRP1 ICD は核内移行し、IFN-γ の発現調節に関わるとされている 37)。
このことから、本研究で観察されたLRP1 ICD の産生が IFN-γ を含む何らかの遺伝子発現に関わ
るならば、虚血性障害で誘導される神経変性に寄与する可能性がある。しかしながら本研究結果 では γ-secretase 阻害薬は ICD 産生に影響を及ぼさず、主に furin が LRP1 を切断する可能性を 示した。
LRP1 ICD は 2 つの NPXY モチーフ( 2 つのうち遠位モチーフは YXXL モチーフとオーバーラ
ップする)、2 つのジロイシンモチーフなど、多くのシグナル伝達モチーフを持つ。中でも YXXL
モチーフが最も優性なエンドサイトーシスシグナルであり、2 つの NPXY モチーフは DAB1、
FE65、JIP1、postsynaptic density (PSD) 95、SHC および CED-6 / GULP のような細胞質アダプター
タンパク質および足場タンパク質と相互作用する 40)。また、シグナル経路に関与するアダプター タンパク質である Shc の結合は、遠位 NPXY モチーフ内の Tyr 63 のリン酸化を必要とする。さ らに、プロテインキナーゼA または C による Ser および Thr のリン酸化は、エンドサイトーシ スおよびアダプター分子との会合を調節する。したがって、LRP1 ICD のリン酸化およびアダプタ ー分子との会合は、リガンド結合の際の物質輸送またはシグナル伝達経路における LRP1 の機能 に必須であると考えられている 41-43)。LRP1 変異体ノックインマウスを用いた研究では、LRP1
ICD の 2 つの NPXY モチーフのうち、近位の NPXY モチーフの変異と、furin 切断部位の変異
は肝臓でのそれぞれ異なる特徴的な表現型を示した。近位 NPXY モチーフの変異は臓器形成不
全により周産期死亡を誘導し、一方の furin 切断部位の変異は LRP1 成熟障害を引き起こし、von
kupffer star 細胞のリソソームの拡大を誘導した 44)。以上より、LPR1 ICD は細胞外ドメインへの
リガンド結合によりリン酸化やアダプター分子との会合を行い LRP1 の主要な機能であるエン
ドサイトーシスやシグナル伝達に重要な役割を担っていることが明らかである。
本研究は、NMDA 障害後に核周辺の領域に集積している LRP1 β-chain (ICD を含む) が、TGN
マーカーである TGN46 および furin と共局在することを示した。本研究結果のみでは、この
TGN 周辺領域での蓄積が細胞表面への輸送途中の LRP1 の蓄積なのか、もしくは細胞表面で切
断を受けた LRP1 β-chain が TGN に運ばれたものかを結論付けることはできない。しかしなが
ら、培養細胞の免疫染色の結果で、α-chain の染色が NMDA 処置によって消失したことから、お
46 TGN 周辺領域で蓄積したと考えられる。
細胞内のタンパク質分解機構の破綻は、アルツハイマー病やパーキンソン病などでみられる神
経細胞死に深く関わると考えられているものの、その詳細は未解明な点が多い。近年 Golgi
membrane associated degradation (GOMED) pathway というゴルジ体が関与する新たなタンパク質分
解機構が報告された 45)。GOMED は、何等かの原因によりゴルジ体から細胞膜への輸送が阻害さ
れることで、ゴルジ体の変形とゴルジ体タンパク質の分解が活性化される細胞障害機序である。 この現象は分泌されずに細胞内に貯留するタンパク質を分解するために機能していると考えられ
ている。GOMED が起こると、ゴルジ体の一部が変形し、不要になったタンパク質などを取り囲
み解離し、リソソームと融合して分解する。ゴルジ体の詳細な病態把握は今後の検討課題である が、本研究結果での LRP1 ICD と furin の局在を考え合わせると、NMDA 障害によって GOMED が誘発されている可能性が考えられた。
NMDA 処置後の LRP1 の切断に furin が関わる可能性があるものの、furin の阻害による LRP1 の切断抑制が大脳皮質神経細胞の生存に影響するのか、また furin 阻害による他の機構が 神経保護機構に影響するのかを明らかにするには、さらなる検討が必要である。
また、LRP1 のリガンドである α2-マクログロブリンはマウス初代培養大脳皮質神経細胞で
LRP1 に結合すると NMDA 受容体を介するカルシウム流入を誘導し、シグナル誘導に関与するこ
とが報告されている 46)。ラット初代培養海馬神経細胞を用いた研究では、LRP1 リガンドである
t-PA によって、LRP1 を介した NMDA 受容体の活性化が引き起こされ、この t-PA によるカルシ
ウムシグナルの誘導には、LRP1 と NMDA 受容体の PSD95 を介した複合体の形成が必要なこと
が明らかになっている 47)。さらに PC12 細胞および初代培養神経細胞を用いた研究で、 t-PA あ
るいは α2-マクログロブリンの LRP1 への結合が Src family kinase を介して tropomyosin receptor kinase (Trk) 受容体の活性化を誘導し、Akt および extracellular signal-regulated kinase (ERK) 経路を
介する神経栄養活性を持つことも報告されている 31)。これらの結果は LRP1 が神経細胞のシグ
ナル伝達や生存に重要であることを示している。一方、初代培養大脳皮質神経細胞を用いた実験
で、t-PA は単独では細胞死を誘導しないものの、NMDA 刺激による Ca2+ 流入を増強し、神経細
胞死の程度を増悪させ、また NMDA による興奮性神経毒性の in vivo 実験で、リコンビナント
t-PA の脳内への注入は NMDA による神経障害領域を増大させた 48)。さらにラット脳梗塞モデル
での脳内 t-PA 活性の上昇は、血液脳関門 (blood-brain barrier : BBB) の破綻や神経細胞死を誘導
し、虚血領域の拡大を招いた。この t-PA による変化は神経細胞の LRP1 の阻害によって抑制さ
れたことから、LRP1 を介すると考えられた 49)。これらのことから LRP1 は神経細胞の NMDA
受容体を介したシグナル伝達や Trk 受容体を介した神経生存に関わるだけでなく、脳梗塞を含む
病態時の神経細胞死誘導にも重要な働きをしていることが示されている。
本研究において in vitro 初代培養細胞に対する NMDA 処置と in vivo ラット脳梗塞モデルの リポタンパク質受容体の変化は同様であった。また、脳梗塞モデルの免疫組織染色の結果から、
初代培養細胞と同様に LRP1 β-chain または ICD が TGN に局在することが示された。リポタン
パク質受容体は、神経細胞以外のグリア細胞にも発現がみられるため、本研究の in vivo 実験にお
47
脳の神経細胞で LRP1 の切断および局在の変化は誘導されていることが推察される。本研究では
ICD の明らかな核への移行は観察できなかったものの、この局在の変化は LRP1 の脳梗塞時の役 割あるいは神経生存に関わる重要な役割を示すのかもしれない。
本研究は脳梗塞後のリポタンパク質受容体の病態生理学的変化を明らかにし、特に神経細胞の LRP1 および furin の NMDA 障害による変化と神経生存に関わる役割を示した (Figure 27)。今
後、LRP1 ICD の病態生理学的役割や furin の in vivo での機能を解明することで、新たな脳梗塞
治療薬開発に貢献できることを期待する。