早稲田大学大学院理工学研究科
博士論文
アカハライモリの腹色形成機構に関する研究 Studies on the making of the red ventrum of the Japanese newt, Cynops pyrrhogaster
松井 久実
物理学及応用物理学専攻 分子生殖生物学研究
2003 年 3 月
目次
第1章 序論
第2章 腹色とアカハライモリの生態 第1節 本研究に用いた個体群 第2節 幼体の成長
第3節 成体と幼体の腹色 第4節 章括
第3章 細胞形態学的研究 第1節 研究の背景 第2節 腹色を担う細胞
第3節 カロテノイド含有色素顆粒について 第4節 成長に伴う色素顆粒の変化について 第5節 章括
第4章 腹色を形成する色素の化学的研究 第1節 研究の背景
第2節 成体腹側皮膚におけるカロテノイドについて 第3節 カロテノイドの体内分布
第4節 成長とカロテノイドについて
第5節 餌限定条件下でのイモリのカロテノイドについて 第6節 章括
1
5 6 12 19 22
23 24 27 39 41 45
47 48 51 65 71 80 84
第5章 幼体期の食性 第1節 研究の背景 第2節 幼体期の食性
第3節 陸棲成体、他種幼体の食性 第4節 章括
第6章 総括
謝辞
文献
89 90 91 98 102
103
106
107
第 1 章
序論
動物の体色は、人間が外観で個々の動物を識別することが可能な形質の一つで、
これまで 2 つの方向から研究されてきた。一つは、体色の機能的な側面につい てである。これは、行動学、生態学の分野において明らかにされてきたもので、
捕食−被食関係における体色の機能(隠蔽色、警告色、擬態色)、求愛行動にお ける機能(婚姻色)、生理学的な機能(体温調節)などが代表的な事項として挙 げられる (Edmunds 1974; Endler 1978; McFarland 1981)。もう一つは体色 の決定機序についてである。動物の体色には発色を担う組織、細胞があり、特定 の色素を用いる定まった発色機構がある(Fox 1976)。この決定機序についての 研究は、細胞学、生理学、化学、生態学など様々な分野で行われている。
近年では、体色の発色機序と行動学的機能の間の相互関係についての報告がな され、近年の動物の体色に関する研究はより包括的なものになっている(Grether et al. 1999; Grether 2000; Hill et al. 2002)。このような研究の代表例として、
魚類の性選択と体色、カロテノイド色素の関連についての一連の研究が挙げられ る。グッピーの雄は、体表にオレンジ色の斑紋を持ち、この斑紋が大きく濃いも のほど繁殖相手として雌に選択される (Endler 1978)。このオレンジ色はカロ テノイドによるもので、グッピーは餌のラン藻からカロテノイドを得ている。体 表のオレンジ色の斑紋の大きさや濃さは餌のカロテノイドによって変化するため、
カロテノイドは、雌による性選択の際、雄の栄養状態を示す指標として働くと考 えられている(Grether et al. 1999; Grether 2000)。このような、カロテノイ ドなどの色素と、体色と行動の関係に関する研究は、様々な動物において報告さ れている(Bernardi et al. 2001; Termonia et al. 2001)。
両生綱有尾目に属するイモリ科には、鮮明な色の腹面を持つものが多く、警告 色としての機能が報告されている(Brodie 1977; Johnson and Brodie 1975)。
そのうち、日本固有種であるアカハライモリCynops pyrrhogaster はテトロド トキシンにより毒化し(Mosher et al. 1964; Yotsu et al. 1990)、背部が暗黒色、
腹部は赤色であることから、体色は警告色として機能すると推察されている
(Brodie 1977; 林 1996)。しかし、アカハライモリの腹色の発色機序について
ハライモリにおける腹色の占める位置について包括的に考察したものである。
本研究では、アカハライモリの腹色の発色機序について三方向からの解析を行 った(図 1)。1 つは発色を担当する細胞の特定を目的とした細胞形態学的研究 である。内容としては、発色担当細胞の位置や性質を明らかにし、また成長に伴 う色素顆粒の変化について解析した。2 つ目は発色色素の同定、成長に伴う変化、
個体における色素の体内分布、色素と体内における代謝などについて考察した化 学的研究である。3 つ目は、生態学的研究である。化学的研究により、アカハラ イモリの腹色は幼体期の食餌に由来する食餌由来色素であることが明らかになっ た。よって、幼体期の食性、成長と腹色の変化について解析を行った。
図1 アカハライモリ腹色の解析方法
細胞形態学的研究
化学的研究 生態学的研究
第 2 章
腹色とアカハライモリの生態
第 1 節 本研究に用いた個体群
第 2 章ではヒトの目から見たアカハライモリの腹色について述べた。本研究 の性質上、研究対象は野生動物であるが、アカハライモリについてはその生態に ついても不明な点が多い。特に、本研究では野生の幼体期の個体を研究対象とし たため、アカハライモリ幼体の生活史についても論じる必要があった。アカハラ イモリの生態については、水中で生活する成体については若干の報告があるもの の、陸上生活する幼体については殆ど情報がない。それは、幼体期のイモリが人 の目に付かないところで生活するため、今までに捕獲された絶対数が少ないため である。幸いにも著者は、複数の幼体が一時に捕獲できる地点を得、本研究に必 要なデータを得ることが出来た。そこで本章では、幼体の成長や年代構成など生 態学的な基礎データについて述べた。
本研究は、腹色の形成機序の解明を目的とするため、野生のアカハライモリの 個体群のうち、以下の 5 条件を満たすものを研究対象とした。
1. 個体群の規模が大きいもの。
2. 棲息場所の環境が安定していること。
3. 個体群の観察が容易であること。
4. 幼体の捕獲、観察が可能であること。
5. 腹側の斑紋パターンが単純であること。
著者が既知の個体群のうち、長崎県五島列島福江島岐宿地区(図 2)の個体群 がこの条件に合致した。一般的なアカハライモリの背面の色は黒一色に近い(図 3a-k)。よく観察すると黒の地色の上に黒の斑紋パターンが認められる場合もあ る。他方、福江島のアカハライモリ(以下、五島個体群)の背側の斑紋パターン は一般的なアカハライモリ個体群と異なり、オリーブグリーンから褐色の地色に 濃い色のスポットが散らばっている(図 3a-g)。対して腹面は一般的には、赤 の地色に黒の斑紋パターンを有する(図 3b-k)。腹面の赤い地色と黒の斑紋パ ターンの地方変異が報告されており、黒の斑紋パターンは斑紋の形状によりタイ
福江島岐宿地区
図2 調査地点所在図
長崎県五島列島
図 2 調査地点所在図
N
a
g
k
b
g
k
bar = 1cm
図 3 五島個体群の背面、腹面外観
アカハライモリ背面(a)、腹面(b)
また、地色である赤色にはいくつかの色相があるものの、一つの個体群での個 体差も大きく明瞭な地方集団間の変異は見られないとされる(Sawada 1963a, 林 1996)。五島個体群の腹面の斑紋パターンは Sawada の分類によるとタイプ I の黒の斑紋が殆ど無いタイプに属し、腹色の色相の判定が行いやすい(図 3b)。
アカハライモリは外部形態と腹部斑紋パターンの解析から、東北,関東,篠山,
渥美,広島,中間種族の 6 地方種族に区分されているが(篠山は兵庫県、渥美 は愛知県)、残念なことに五島個体群の解析はなされていない(Sawada 1963a)。
また、アカハライモリの種内分化については、アロザイム分析による遺伝的な系 統関係が調べられており、Sawada の分類では広島種族となっている西日本の集 団はさらに細分化されている(Hayashi 1988)。本研究における研究対象とした 五島個体群のイモリの系統関係については、複数のアロザイムの遺伝子座におけ る遺伝子頻度の解析がなされており、本個体群は九州北部産と比較していくつか の遺伝子座で顕著な遺伝子頻度の違いが認められるが、アカハライモリ全体の地 理的変異の中で見たときには九州北部産と共に西日本集団の中に含まれている
(Hayashi, personal communication)。
調査は標高 180 m、山の谷筋斜面の上部に作られた棚田とその周囲の草地で 2000 年 5 月上旬、11 月下旬、2001 年 5 月上旬,11 月中旬、2002 年 5 月上旬 に行った。山の斜面は南に面しており棚田の面積は約 50a で、田圃が作られて から 100 年以上は経過している。圃場整備は行われていない。棚田の横には一 年中水の枯れない約 4 m2のコンクリート製人工池が作られている。5 月には春 の求愛がこの池と水入れ後の棚田で見られた。11 月の調査期間は秋の求愛の季 節で雄の尾部には婚姻色を確認した。1998 年に水中の成体イモリの採集を行っ ている。この調査場所には他の両生類として、ツチガエル Rana rugosa、ヌマ ガエル Rana limnocharis、アマガエル Hyla japonica、シュレーゲルアオガエ ル Rachophorus schlegelii、カスミサンショウウオ Hynobius nebulosus が棲 息している。
図 4 に、水中において 2001 年 5 月に捕獲した成体雄 21、雌 38 個体の頭胴長
(SVL)分布を示した。ヒストグラムの階級は 1 mm 幅とした。平均 SVL(Mean
±SD)は雄では 47.70±2.34 mm、雌では 52.42±3.20 mm であった。最大 SVL は雄で 51.5 mm、雌では 58.5 mm、最小 SVL は雄で 41.1 mm、雌で 41.1 mm であった。
6 5 4 3 2 1 0 1 2 3 4 5 6 7
84 0 4 2 4 4 4 6 4 8 5 0 5 2 5 4 5 6 5 8 6
♂
♀
47.70
0 2.34(SD), N=21
52.42
±
3.20(SD), N=38
図4 五島個体群成体の頭胴長分布
±
図 4 五島個体群成体の頭胴長分布
SVL (mm)
(個体数)
第 2 節 幼体の成長
アカハライモリの生活史の概略を図 5a に示した。春に産卵された卵は 1 ヶ月 程度で孵化し、幼生となる。幼生は夏の終わりから秋にかけて変態、上陸する。
幼体は小さな無脊椎動物を捕食しながら陸上生活すると考えられている(佐藤 1943; 林 1996)。水場に戻り、繁殖に参加するようになるまで標高 500 m 以下 の 平 地 で は 3 年 、 高 地 で は 4 年 か ら 7 年 以 上 か か る と 推 察 さ れ て い る
(Marunouchi et al. 2000)。図 5b には、幼体の成長についての模式図を示し た。1 年目の夏から秋にかけて変態上陸した幼体は、冬がくると冬眠に入り、冬 眠の間の成長は中断する。2 年目には春のはじめから成長できるため、幼体の成 長は段階的なものとなる(Bell 1977)。よって、複数年齢で構成される幼体個体 群の体の大きさをプロットすると、幼体期間の年数に応じていくつかのピークが 出現する。
調査地において幼体を広く捜索したところ、この調査地における幼体は限定さ れた範囲の地表で捕獲された。地表への出現時間も限られていた。図 6 に 2001 年 11 月に捕獲した幼体の写真を示した。幼体の大きさはおおよそ 3 サイズに分 けられる傾向にあった。2000 年から 2002 年にかけての 5 月に捕獲した幼体の SVL 分布図を図 7 に、11 月に捕獲した幼体の SVL 分布図を図 8 に示す。図 9 には捕獲個体の各 5 月の頭胴長分布を示した。2000 年 5 月に捕獲した個体を全 て除去したところ、2000 年 11 月には図 10 のような SVL 分布が得られた。
繁殖水域
変態 幼体
卵 幼生
性成熟
繁殖に参加
数年かかる
変態
春 夏〜秋
卵→幼生
冬 成長
春 冬
冬眠 成長
春 冬眠 成長
図5 アカハライモリの生活史
a a a a
b b b b
図 5 アカハライモリの生活史
カスミサンショウウオ幼体カスミサンショウウオ幼体カスミサンショウウオ幼体 カスミサンショウウオ幼体
イモリ幼体イモリ幼体イモリ幼体 イモリ幼体
1cm1cm1cm 1cm
幼体小
幼体中 幼体大
図6 採集した幼体
(2001年11月)図7 3年間(5月)の幼体頭胴長分布
7
6
5
4
3
2
1
0
18 20 22 24 26 28 30 32 34 36 38 40 42 2000
2001 2002
(mm)
(個体数)(個体)
図8 2年間(11月)の幼体頭胴長分布
(mm)
(個体)
2000
2001
(個体数)
2000 2001 2002
図9 各5月の幼体頭胴長分布
(mm)
(個体数)
(個体数)
(個体数)
図10 各11月の幼体頭胴長分布
(mm)
(個体)
(個体)
2000
2001
(個体数)
(個体数)
第3節 成体と幼体の腹色
成体の腹色
林(1996)は、同一個体群の中でも腹色は多様であると述べている。そこで、
個体群内の腹色の多様性を調べる事を目的とし、水中で捕獲した成体イモリの腹 色を CMYK 方式の色チャート(視覚デザイン研究所)と対応させ、2002 年 5 月に五島個体群の成体 18 個体の腹色を調べた。CMYK 方式は印刷物の発色の指 標として用いられる表示方式である。表 3 中に示した数字は、黒 - シアン – マゼンダ - イエローの含有率を示している。結果を表 1b に示す。
幼体の腹色
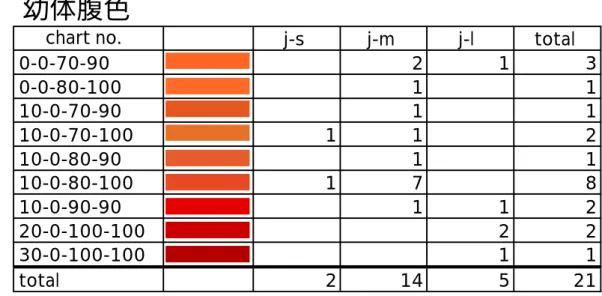
成体と同様の方法で、2002 年 5 月に 21 個体について腹色を調べた。図 11 に 幼体腹面の写真を示した。成体の大きさの最小値は 44.1 mm であるが、ここで は SVL41mm 以下の個体を幼体とし、頭胴長により j-s(幼体小)、j-m(幼体 中)、j-l(幼体大)の 3 群に分け、それぞれの腹色を解析した。 結果を表 1a に 示す。
結果
成体の腹色は、10-0-90-90, 20-0-100-100, 30-0-100-100 に集約した(表 1)。幼体では SVL 35 mm 以上の j-l (幼体大)で成体に見られる 10-0-90-90, 20-0-100-100, 30-0-100-100 の腹色を有した(表 1)。
幼体小 幼体中 幼体大
腹面
図11 幼体の腹色
(2001年11月)表 1 色チャートで示した成体, 幼体の腹色
j-s j-m j-l total
0-0-70-90 2 1 3
0-0-80-100 1 1
10-0-70-90 1 1
10-0-70-100 1 1 2
10-0-80-90 1 1
10-0-80-100 1 7 8
10-0-90-90 1 1 2
20-0-100-100 2 2
30-0-100-100 1 1
total 2 14 5 21
0-0-80-90 1
0-0-80-100 1
0-0-90-100 1
10-0-90-90 5
10-0-100-100 1
20-0-80-100 2
20-0-100-100 4
30-0-100-100 3
total 15
chart no.
成体腹色 幼体腹色
j-s: SVL< 24.9 mm j-m: 25< SVL< 34.9 mm j-l: 35< SVL < 40.9 mm
a
b
18
第 4 節 章括
本章では、五島個体群における腹色形成時期を知るため、成体の大きさ、幼体 の大きさの分布と成長について、および肉眼で見た腹色についての解析を行った。
成体の大きさは、最小の個体でも雄雌とも SVL 44.1 mm で、陸地で捕獲した個 体(成体,幼体を含む)の SVL 最大値よりも大きかった。
幼体の体長分布は 3 または 4 つのピークを持つことから、五島個体群の幼体の性 成熟にかかる期間は 3 年から 4 年程度であると考えられた。アカハライモリの成 熟年齢は、棲息地の標高によって異なり、低地では 3〜4 年、山の上などの高地 では 7 年以上かかると推測されている(Marunouchi et al. 2001)。マダライモリ Triturus marmoratus においても高地と低地では性成熟年齢が異なることが報告 されている(Caetano and Castanet 1993)。本研究の調査地の標高は約 180m と低地であり、幼体が 0〜3, 4 才の年齢層を持つことは妥当である。今後、指骨 を用いた年齢査定法を併用し、五島個体群の幼体の成長を明らかにする。
腹色の評価に関して、ヒトの目で見た色は、視認対象物の本来持つ色を反映し ているとはいえない。反射スペクトルを測定する方法がより正確にアカハライモ リの腹色を捉えることができる。この方法は便宜的なものであることを注記する。
第 3 章
細胞形態学的研究
第 1 節 研究の背景
動物の体色は様々な機序で発色する。一つは皮膚およびその付属器官が特定の 光の波長のみを反射し、体色を作り出す場合がある。その中で知られるものには 鳥類の羽毛による体色発現や昆虫の金属光沢のある外皮などが挙げられる(Fox 1976)。また、血液中のヘモグロビンの色が赤い皮膚の発色に寄与する事も知ら れており、サルの尻の色やニワトリの頭冠が有名な例である(Fox 1976)。第 3 の例として、色素細胞による体色の発現が挙げられる。本研究の研究対象である アカハライモリは両棲綱有尾目に属し、一般的に両生類の体色発現は色素細胞が 担うことが明らかとなっている(Bagnara 1998)。
色素細胞とは、神経堤細胞(神経冠細胞)に由来し、細胞質内に色素を保有ま たは 生産し 、体色の 発現の 機能を 担って いる細 胞群であ る(Bagnara et al.
1979)。哺乳類,鳥類はメラニン生成により黒を発色するメラノサイトのみを保 有するが、鳥類以下の脊椎動物には、数種の異なる色調を担当する色素細胞が存 在する。なかでも両生類は、メラノフォア, イリドフォア, ザンソフォアの3種 の色素細胞の存在が知られている(表 2)。
水溶性・
食餌由来 フラビン
脂溶性・
食餌由来 カロテン キサントフィ
ル カロテノイ
ド顆粒
プリン結晶 プテリジ
ン フェオメラニ
ン ユーメラニン 保有色素
不溶性(アル カリで可溶)・
生合成 水溶性・
生合成 不溶性
生合成 色素の特徴
反射小板 プテリノソーム
メラノソーム 色素産生
保有場所
白〜青 黄〜赤
黒〜青, 黄, 赤 色調
イリドフォア ザンソフォア(エリスロフォア)
メラノフォア 色素細胞
メラノフォアはメラノソーム内に色素としてメラニンを持ち殆どの光波長を吸 収して黒化する。イリドフォアは反射小板といわれるプリン体結晶物を一定配列 で保持し、特定の波長を反射または散乱する(Bagnara 1998)。ザンソフォア はプテリノソーム内に水溶性色素プテリジン、およびカロテノイド顆粒に脂溶性 色素のカロテノイドを持ち、特定の波長を吸収する。ザンソフォアは複数の色素 の組み合わせや細胞内小器官の構造変化、蛋白の結合などにより様々な色を呈す ることが知られている。いずれの色素細胞も動物種によって形態学的・生化学的 多様性が報告されており、色調により黄色のザンソフォア(黄色素胞)、赤のエ リスロフォア(赤色素胞)またはザンソフォア、紫のビオレオフォア(紫色素胞)
な ど に 分 類 さ れ て い る 。( Bagnara and Taylor 1970; Frost et al. 1984;
Ichikawa et al. 1998; Nishioka and Ueda 1985; Yasutomi and Hama 1971;
Yasutomi 1973)。また、3 種の色素細胞が一つの構成単位を作り(真皮性色素胞 単位)、個々の色素細胞では発色できない体色を作り出すことも知られている
(Bagnara 1968)。両生類の色素細胞は特定のホルモンによって拡散収縮する が、神経支配は受けていない(Bagnara and Hadly 1973)。
色素細胞の識別や分類は、光学顕微鏡や電子顕微鏡を用いた細胞質内の色素、
または色素産生細胞の微細構造の区別と色素の化学的な解析により行われてきた。
両生類の色素細胞についても同様で、形態学的な研究は無尾類・有尾類で多数の 報 告 が あ る 。 ザ ン ソ フ ォ ア に 関 す る 代 表 的 な 研 究 は 、 ニ ホ ン ア カ ガ エ ル
(Yasutomi 1971; Yasutomi and Hama 1973)、ヤマアカガエル(Ichikawa et al. 1998)、シュレーゲルアオガエル(Nishioka and Ueda 1985)、アフリカツ メガエル(Yasutomi and Hama 1972)、スズガエル(Frost and Robinson 1984;
Eicheiburg and Obert 1979)メキシコサンショウウオ(Frost et al. 1984)、
ブチイモリ(Forbes et al. 1973)、アルペンイモリ(Pederzoli and Trevisan 1990)、セアカサンショウウオ(Bagnara and Taylor 1970)で行われている。
ザンソフォアには、プテリジンを持つもの、カロテノイドを持つもの、その両 方を持つものが存在する。プテリジンは水溶性色素で環状プリン体構造を示し、
一般に黄色を呈色する。しかし、ニホンアカガエルは赤のドロソプテリジンを持
ち(Yasutomi 1973)、シュレーゲルアオガエルは紫のプテリジンを有するプテ リノソームを持つ(Nishioka and Ueda 1985)。プテリジンはザンソフォアの 細胞小器官であるプテリノソーム中に存在し(Matsumoto 1965)、動物によっ てはザンソフォアの分化,発達と共にプテリノソームがラメラ構造を示すことが 知られ、4 つの分化段階に分類されている(Yasutomi and Hama 1971)。プテ リジンは動物体内で生合成される色素で、魚類においてその代謝経路が明らかに され(Zieglar et al. 2000)、代謝酵素やその機能も知られている(Gutlich et al.
1996; Iino et al. 1996, 2000; Kato 1971; Pelletier et al. 2001)。
ザンソフォアに含まれるもう一つの色素としてカロテノイドが挙げられる。カ ロテノイドは脂溶性色素で細胞中では脂肪滴に溶け込んで存在すると考えられて いる。8 つのイソプレンが共役二重結合し、両端に C6の環状構造を有するのが 基本構造である。動物はカロテノイドを合成する事ができないと考えられている。
それは今までのところ、イソプレンを 8 つ結合する経路は動物では見つかって いないためである。形態学的見地から見たカロテノイドについては報告が少ない。
それはカロテノイドが脂溶性色素であることから顕微鏡試料作製中に溶出すると 考えられていること、また体内で合成されない食餌性色素であることから解析が 困難な事が推察される。
アカハライモリにおいては、過去にプテリジン色素について解析されており
(Obika 1963; Bagnara and Obika 1965)、幼生,変態個体ではアカハライモ リの皮膚は数種のプテリジンを有するが、成体では多量の水溶性色素であるリボ フラビンを有するもののプテリジンは検出されないとある。また、Hama(1963)
もアカハライモリザンソフォアの発生過程におけるプテリジン類の変化について 報告している。
本章はアカハライモリの腹色の発色機序について細胞形態学的な側面から解析 したもので、腹色発色細胞の特定とその細胞の特徴を明らかにすることを目的と している。本章は 5 節で構成され、アカハライモリ成体における腹色発色細胞 の特定、その細胞の色素顆粒について、および成長に伴う色素顆粒の変化につい
第 2 節 腹色を担う細胞
第 1 節において、両生類の体色発色機序の概要を述べたが、アカハライモリ の腹色について具体的な解析は次のように行った。新鮮組織の凍結切片を光学顕 微鏡、で観察し細胞の位置や性質を特定、また透過型電子顕微鏡を用いて色素顆 粒の特定を行った。また、孵化後から餌限定条件下で飼育した人工飼育個体は、
腹側皮膚が黄色いイモリとなる。この黄色い個体の腹の色素細胞についても同様 に観察し、腹の赤い成体との形態学的差異について検討を加えた。
材料と方法
成体のイモリは広島県内の中国山地側の止水域で 1998 年 5 月に採集したもの、
および長崎県五島列島福江島で採集した個体を用いた。採集後、人工照明下で昼 8 時間、夜 16 時間の条件で室温飼育した。材料の採取は採集後 2 ヶ月以内に行 った。週に一度フタホシコオロギ Gryllus bimaculatus 2 齢虫および冷凍アカ ムシ(ユスリカ幼虫)を与えた。人工飼育の成体は次のように作製した。状態の よい雌を選んで hCG(25 unit/個体)を 2 日おきに腹腔内に注射し受精卵を 得た。孵化した幼生は 20〜25℃の室温で飼育し、孵化三日後より毎日、孵化直 後のブラインシュリンプ Artemia sp.を充分量与えた。変態後の幼体には孵化 直後のフタホシコオロギを与え、成長と共に与えるコオロギの大きさを大きくし た。フタホシコオロギは、広島大学理学研究科附属両生類研究施設内で繁殖飼育 されているものを用いた(Nishioka and Matsuura 1977)。
材料は、光学顕微鏡観察用、電子顕微鏡観察用にそれぞれ採取した。ジエチル エーテルで麻酔後、外科用剪刀で皮膚を約 5mm×10mm の大きさに切り出した。
採取した部位は、体幹背側部、体幹腹側部の黒斑領域、非黒斑領域の 3 部位で、
光学顕微鏡観察用および電子顕微鏡観察用に材料を分割した。
光学顕微鏡観察用の材料は、無固定で OCT コンパウンド(Tissue-Tek)に 包埋し、液体窒素中で急速凍結後、-20℃に設定したクリオスタットで厚さ8 µm の、皮膚に垂直な連続切片を作製した。切片は無染色で観察し、必要に応じてメ
チルグリーンで核染色を施した。組織化学的なカロテノイド同定法として硫酸試 験法(Lison 1936)を行った。観察は光学顕微鏡(オリンパス BH-2)を用い て行い色温度及び色フィルター条件を同一にして写真撮影を行った。イリドフォ アの検索は、斜上から光を照射し反射光を検出した。
電子顕微鏡観察用試料は、2.5%グルタールアルデヒド/Millonig's 燐酸緩衝 液 pH.7.4 中で4℃2時間固定し、固定液中で細切後、0.12 Mサッカロース添 加燐酸緩衝液で 15 分4回洗浄、1%オスミウム/0.42 Mサッカロース添加燐酸 緩衝液で4℃2時間固定した(Millonig 1961)。0.12Mサッカロース添加燐酸緩 衝液で 15 分 4 回洗浄後、エタノール上昇系列で 10 分ずつ脱水、無水エタノー ルで 15 分 2 回脱水後、プロピレンオキサイド(関東化学)で 30 分 2 回置換、
EPON812(TAAB)樹脂を加えて置換、プロピレンオキサイドを一晩室温で蒸 発させた後、平板包埋シリコンに水平静置した(Richardson et al. 1960)。樹 脂の重合は電子顕微鏡用包埋恒温器 TD-700B(堂阪イーエム)内で 37℃一晩、
60℃2日間の条件で行い完全に重合させた。包埋ブロックごとにガラスナイフ で 1μm の厚切り切片を作製、Methylene blue - AzurⅡ染色液で染色し、光 学顕微鏡で観察後超薄切部位を選択した。ウルトラミクロトーム(Sorvall MT- 6000)を用いガラスナイフで切削面を研磨後、ダイヤモンドナイフを用いて約 90 nm の厚さに薄切した。切片は 300 メッシュの銅グリッド(VECO)に拾い、
風乾した後、1%酢酸ウラン/70%エタノールで 40 分染色した。その後、70%エ タノールで 1 回、蒸留水で2回洗浄、蒸留水で 15 分間流水水洗後、1%クエン 酸鉛水溶液で4分間染色し、蒸留水で 30 分流水水洗して、風乾した(Raynord 1965)。観察は日立 H-300 電子顕微鏡で行った。
腹色とカロテノイド顆粒タイプの相関を調べる目的で次のように解析を行った。
野生成体、人工飼育成体のそれぞれ 2 個体分の異なる 2 つの組織ブロックから 複数の切片を切り出し、透過電子顕微鏡下で 2000 および 3000 倍の倍率で撮影 した。ネガフィルムを用いてカロテノイド顆粒を約 1000 個タイプ別に計数し、
対応のある 2 群の比率の差の検定を用いて統計学的有意差を検定した。
結果
図 12 に人工飼育個体の外観を示した。養成敷幼生時期のアカハライモリの腹 面は半透明で腹部臓器が透けて見えている。変態直後の個体では、変態後数日で 腹面は不透明となり、薄いクリーム色を呈する。成体(変態後 3 年)になると、
腹面の色は黄色を呈する。
野生個体の背側皮膚では表皮性メラノフォア、メラノフォア、ザンソフォアが 確認された。図 13 は、背面がオリーブ色を呈しているイモリ(五島個体群)の 凍結切片像で、真皮最上層にメラノフォアとザンソフォアが並列した。ザンソフ ォアは黄色色素を含有していた。背面が黒いイモリでは真皮最上層はメラノフォ アに覆われ、ザンソフォアは確認できなかった。体側皮膚の腹側面では真皮最上 層にザンソフォアが赤色素を伴っており、メラノフォアは収縮してザンソフォア の下部に位置した(図 14b)。
野生個体の腹側皮膚正中面では赤色素を伴うザンソフォアと、イリドフォアが 真皮上層に観察された(図 14c, d)。Lison(1936)の硫酸試験法では、赤色素 の分布する領域において青色の反応が認められた。ザンソフォアはコラーゲン層 に沿って観察され(図 15a)、ザンソフォアの内部は多数の色素顆粒で占められ ていた。色素顆粒は 2 種のカロテノイド顆粒とプテリノソームの 3 種類が観察 され、色素顆粒の殆どはカロテノイド顆粒であった(図 15b)。カロテノイド顆 粒は直径が約 0.45 µm で非常に電子密度が高く、内部まで均一な電子密度を持 つもの(均一型カロテノイド顆粒)と、外周は電子密度が高く内部は抜けたよう に見えるもの(リング型カロテノイド顆粒)があり、リング型が多く観察された
(図 15c)。プテリノソームの直径は約 0.9 µm で、カロテノイド顆粒の間に散 見された(図 15d)。
人工飼育成体では、腹部正中面皮膚に黄色素を伴うザンソフォアと、イリドフ ォアが観察され(図 16b)、これらの細胞は野生個体と同様の位置にあった。ザ ンソフォアの内部は野生個体と同様に多数のカロテノイド顆粒で占められていた が、種類は均一型が優占していた(図 16d)。
卵からの飼育個体
野生個体
1cm
幼生(St. 55)
変態直後
図12 人工飼育個体の腹面
腹面
腹面
1 cm
1 cm
x a m
e p i
図13 背側皮膚の色素細胞
e m
epi: 表皮細胞
em: 表皮性メラノフォア xa: ザンソフォア
メラノフォア m:
bar = 10 µm
メチルグリーン核染色を施している epi: 表皮細胞
em: 表皮性メラノフォア xa: ザンソフォア
m: メラノフォア 核染色:メチルグリーン
a b
s g
e p i m
xa ir ir e p i
c
xa ir
d
e p i xa
ir
10 m 10 m
10 m
s g
図14 腹側皮膚のザンソフォア
s g
a: 野生成体腹面 b: 体側面皮膚垂直断 c: 腹部正中面垂直断 d: 切片c反射光観察 epi: 表皮細胞 xa: ザンソフォア m: メラノフォア ir: イリドフォア
sg: 皮膚腺
a
e p i
xa
c l p g
10 m b 10 m
h c g
rcv
c 1 m d 1 m
p t
h c g
rcv
図15 ザンソフォア色素顆粒の微細構造
a: 樹脂包埋切片 b: ザンソフォアの色素顆粒(透過電顕像)
c: カロテノイド顆粒 d: プテリノソ−ム
epi: 表皮細胞 xa: ザンソフォア pg: 色素顆粒 hcg: 均一型カロテノイド顆粒 rcv: リング型カロテノイド顆粒 pt: プテリノソーム
a b
e p i
ir xa s g
c
xa e p i p g
p g
10 m
10 m
d
r c v
h c g
図16 人工飼育個体のザンソフォア
a: 野生成体腹面 b: 腹部正中面垂直断 e: 樹脂包埋切片 f: ザンソフォアの色素顆粒(透過電顕像)
epi: 表皮細胞 xa: ザンソフォア ir: イリドフォアsg: 皮膚腺 pg: 色素顆粒
10 m
a: 野生成体腹面 b: 腹部正中面垂直断 c: 樹脂包埋切片 d: ザンソフォアの色素顆粒(透過電顕像)
epi: 表皮細胞 xa: ザンソフォア ir: イリドフォアsg: 皮膚腺 pg: 色素顆粒
野生個体と人工飼育個体では含有するカロテノイド顆粒のタイプの違いが示唆 されたことから、カロテノイド顆粒 1000 個中のリング型、均一型の割合を解析 した(表 3)。野生個体では 84.0%のカロテノイド顆粒はリング型、16.0%は均 一型であった。対して、人工飼育個体では 92.4%が均一型、7.6%がリング型で あった。野生個体ではリング型が優占すること、人工飼育個体では均一型が優占 することは、統計学的に有意であった(p<0.01)。
薄層クロマトグラフィー(TLC)によりカロテノイドの含有を確認した(図 17)。
野生個体、人工飼育個体共に 1 種のカロテンと複数のキサントフィルが確認さ れた。抽出カロテノイドの量は野生個体が明らかに多かった。
合計数 リング型 均一型
野生個体 1022 858 164
(n=2) 100.0% 84.0% 16.0%
飼育個体 956 72 884
(n=2) 100.0% 7.6% 92.4%
p<0.001
*
*
*
表3 カロテノイド顆粒の含有頻度
S W
β-カロテン
L
カンタキサンチン
ルテイン
S : 標品
W: 野生個体腹側皮膚 L : 飼育個体腹側皮膚
図17 カロテノイドのTLC
展開開始点
考察
色素細胞により体色がつくられる場合、体色は複数の色素細胞の相互作用によ って生じる。メラニン、プテリジン、カロテノイド、プリン体結晶が存在する場 合、メラニンは広いレンジにわたる光波長を吸収する。プテリジン、カロテノイ ドは特定の波長領域を吸収し、その他の色素は特定の波長を反射、透過する。プ リン体結晶は光を一定の角度に反射および散乱する。よって、これらの色素の皮 膚内での立体配置、つまり色素細胞の配置が、その動物の体色を決定する事にな る。アカハライモリの場合、背側においても腹側においても、メラノフォアとザ ンソフォアは皮膚内で同一深度に位置していた。しかし、背側と腹側ではザンソ フォアの色調は異なっていた。イリドフォアはメラノフォアとザンソフォアの下 方に位置していた。すなわち、皮膚に入射した光波長は、メラノフォアでは吸収、
ザンソフォアでは一部の光波長が吸収され、さらにイリドフォアにより体外側へ 反射、再度メラノフォア,ザンソフォアのフィルターを通過する事になる。これ は、体色の発色を増強すると考えられ、また、他にも生理学的な意義をもってい るとも考えられる。
本節から、野生個体でも人工飼育個体でも、ザンソフォアは同様に分化し、た くさんのカロテノイド顆粒を形成することが明らかになった。ザンソフォアには プテリジン、カロテノイドが存在するが、透過電顕像から、ザンソフォアの内部 はほとんどカロテノイド顆粒で占められており、アカハライモリの腹色にはカロ テノイドが大きく関与している事が示唆される。さらに、タイプ別に見ると野生 個体のザンソフォアでは有意にリング型カロテノイド顆粒が含有されていた。一 方、人工飼育個体では有意に均一型カロテノイド顆粒が多く、両者にはカロテノ イド組成や量に差があることが示唆された。一般に脊椎動物のザンソフォア中に 見られるのは均一型のカロテノイド顆粒である。リング型のカロテノイド顆粒は 過去の報告でも観察されているが、リング型カロテノイド顆粒の形成については 明確な論議はされていない(Robinson and Charlton 1973, Yasutomi 1973)。
リング型カロテノイド顆粒の形成についてはさらなる解析を必要とするが、カロ テノイド顆粒のタイプの違いは腹色の違いを反映していると考えられる。
カロテノイドは脂溶性色素で、生体内では遊離型として、およびエステル化な どを受けて存在していると考えられている。脂質に溶けて存在する事も考えられ る。脂肪酸の組織内での存在状態について組織学的に解析した報告では、電顕試 料作製時の飽和脂肪酸と不飽和脂肪酸の組織内への残存には違いがある事が示唆 されている(Angermüller and Fahimi 1982)。電顕試料作製時には固定剤とし て四酸化オスミウム(OsO4)が用いられるが、不飽和脂肪酸は四酸化オスミウ ムによって組織内に固定されるという。飽和脂肪酸は四酸化オスミウムの作用を 受けず、組織内から流出する。
ここで話をカロテノイドに戻す。カロテノイドは多数の共役二重結合を有する 物質である。カロテノイドにも様々な構造があるが、基本となるカロテン骨格は 8 つの共役二重結合を含んでいる。これまで、カロテノイドは電顕試料作製時に 組織内から流出すると考えられており、組織内残存の可能性について議論された 例はない。そこで、第 3 節では、カロテノイドに対するオスミウムの作用につ いて検討し、カロテノイド顆粒のタイプとカロテノイドの関係について考察した。
第 3 節 カロテノイド含有色素顆粒について
これまでの定説では、カロテノイドは脂肪滴中に存在し、形態学的研究では標 本作製の際に流出すると考えられている(Yasutomi 1973)。一方、Angermüller and Fahimi(1982)らは、炭素二重結合を有する不飽和脂肪酸は四酸化オスミ ウムによって標本中に固定されることを報告している。飽和脂肪酸は四酸化オス ミウムの作用を受けない。カロテノイドは共役二重結合を多数有する物質である ことから、四酸化オスミウムの作用を受け、標本中に残存する可能性がある。そ こで、修飾基の異なるカロテノイドに対する四酸化オスミウムの作用を試験した。
材料と方法
β-カロテン(Wako)、カンタキサンチン(津島博士からの寄贈)、ルテイン
(Sigma)、アスタキサンチン(Sigma)について試験した。各カロテノイドの 溶液を 2 枚の濾紙(Whatman No.5, Whatman)にそれぞれスポットし、風乾 後、1 枚は 1%四酸化オスミウム水溶液(日新 EM)に浸し、室温で 30 分間反 応させた。反応後、透過電顕試料作製時に用いるプロピレンオキサイド(関東化 学)で洗浄し、濾紙上のスポットの残存を確認した。
結果
アスタキサンチン,ルテイン,β-カロテンの結果を図 18 に示した。β-カロ テン,ルテインは四酸化オスミウムの作用を受けて濾紙上に固定され、有機溶媒 に溶出しない事が分かった。対して、アスタキサンチンでは、四酸化オスミウム の作用を受けず溶出した。また、カンタキサンチンはアスタキサンチンと同様に 溶出した。アスタキサンチンモノエステル,ジエステルは四酸化オスミウムの作 用を受けて濾紙上に固定された。四酸化オスミウムを作用させなかった濾紙では、
全てのカロテノイドが溶出した。
考察
カロテノイドは種類によって、透過電子顕微鏡下で検出できる事が明らかとな った。ケト基を持たないカロテノイドは透過電子顕微鏡下で電子密度の高い物質 として観察されると考えられた。ケト基を持つカロテノイド(以下ケトカロテノ イド)は四酸化オスミウムの作用を受けず、流出する事が考えられた。ケトカロ テノイドはβ-カロテンよりも二重結合の数は多いが流出する。すなわち、ケト 基を有する事により、四酸化オスミウムの酸化を受けないことが考えられた。
-カロテン ルテイン
アスタキサンチン ß
OH HO
R O
R OH HOO
図18 四酸化オスミウムのカロテノイド固定力
作用前
作用後有機溶媒洗浄
有機溶媒洗浄のみ OsO4
第 4 節 成長に伴う色素顆粒の変化について
第 2 節ではアカハライモリのザンソフォアがどのような状態で存在している のか、どのような色素を含んでいるのか明らかにした。両生類のザンソフォアは 一般に、幼生期と変態後では発色色素の組成が変化することが知られている
(Bagnara 1998)。そこで、本節では人工飼育個体、野生個体の両者を用いて、
成長に伴うザンソフォアの色素顆粒の変化の過程を解析した。
材料と方法
幼生および変態直後の幼体は、五島個体群の成体から得た受精卵から発生させ た。状態のよい雌を選んで hCG(25 unit/個体)を腹腔内に注射し受精卵を 得た。孵化した幼生は 20〜25℃の室温で飼育し、孵化三日後より毎日、孵化直 後のブラインシュリンプ Artemia sp.を充分量与えた。幼生の発生段階の判断 には、岡田のイモリの発生段階基準改訂図表を用いた(Okada and Ichikawa 1947)。野生幼体は 2001 年 5 月に採集し、SVL により幼体小(SVL< 24.9 mm)、
幼体中(25 mm < SVL < 34.9 mm)、幼体大(35 mm < SVL)の 3 群に分け た。材料の採取は採集後 2 週間以内に行った。解析方法は第 2 節に準じた。
結果
幼生期に見られる色素細胞は背側,腹側共にメラノフォアとザンソフォアの2 種類であった。イリドフォアは変態以降に観察された。
ステージ 44(孵化後約2日後、採餌開始時)において、ザンソフォアはプテ リノソームを多数持つ細胞として真皮内の緩い結合組織内に存在していた。カロ テノイド顆粒はほとんど見られなかった。プテリノソームの大きさは直径約 1µm で、内部はほぼ無構造、しばしば微少な繊維状の物質が見受けられた。ステージ 55(幼生中期)になるとザンソフォア中の多数のプテリノソームの間隙にカロ テノイド顆粒の形成が散見された。ステージ 59(変態前期)ではカロテノイド 顆粒の数が増加した(図 19a)。カロテノイド顆粒の最大直径は約 0.78 µm で、
ステージ 55 と同様に中心部の電子密度が高かった(図 19a)。ステージ 60(変 態期)ではさらにカロテノイド顆粒の数は増加し、小胞の最大直径は約 0.85µm、
プテリノソームとほぼ同じ大きさであった。電子密度は全体的に高くなっており、
電子密度の高い中心部が拡大した(図 19b)。
図 20 には、成長に伴うカロテノイド顆粒の数とサイズ(直径)の変化を示し、
図 19 と併せ結果を述べる。図 19b にはステージ 60(変態期)の腹側皮膚ザン ソフォアを示した。カロテノイド顆粒の数は増加し(図 20)、直径はプテリノソ ームとほぼ同じ大きさになった。図 19c-e は野生幼体のザンソフォアを示して いる。幼体小では(図 19c)、多数のプテリノソームの間に多数の小さなカロテ ノイド顆粒が存在している(図 20)。プテリノソームの大きさは成長を通じ、ほ とんど変化しなかった。
幼体中(図 19d)のザンソフォアでは、カロテノイド顆粒の直径は幼体小よ りも大きくなっている(p< 0.05)(図 20)。写真上で穴のあいたように見える箇 所は、イリドフォアの反射小板が存在した所で、切片の染色過程で溶出している。
幼体大のザンソフォアは、カロテノイド顆粒の直径、数共に幼体(中)のものと 有意差はなかった(図 20)。
図 19f には 野生成体のザンソフォアを示した。成体になり初めて、リング型 カロテノイド顆粒が認められた。カロテノイド顆粒のサイズ、数は幼体(中、大)
と有意差はなかった(図 20)。
図 20 の表において、成長に伴うカロテノイド顆粒の、単位区画あたりの面積 を示した。面積は成長と共に増加した。
a: st. 59 b: 変態時 (st. 60) c: 幼体小d: 幼体中e: 幼体大f: 成体
黒矢頭: カロテノイド顆粒(均一型)白矢頭: カロテノイド顆粒(リング型)
白矢印: プテリノソ−ム
図19 成長に伴うザンソフォア色素顆粒の変化
a
c
b
d
e f
bar=5µm
図20 成長過程におけるカロテノイド顆粒と プテリノソームの変化
Diameter ( µ m)
Number /100 µ m
2140 120 100 80 60 40
20 0
1 0.9 0.8
0.7 0.6 0.5 0.4 0.3 M S -j M - j L - j A d
s t. 44 s t. 55 s t. 59
Number of carotenoid granules Size of carotenoid granules
19.49 17.63
15.78 9.76
0.77 0.47
0.35 0.12
Total area ( m π
2) (S/2)2N
0.46 ± 0.02 0.47 ± 0.05
0.45 ± 0.02 0.35 ± 0.02
0.85 ± 0.05 0.78 ±
0.70 ± 0.10 0.07 0.83 ±
0.03 S
( m) (Mean –
SEM)
101.67 ± 24.48 Large juveniles
n=3
99.25 ± 16.11 Medium juveniles
n=3
0.22 ± 0.06 Larvae (st.44)
n=2
0.92 ± 0.18 Larvae
(st.55) n=2
117.33 ± 17.13 Adult
n=2
101.50 ± 13.21 1.35 ± 0.58
0.99 ± 0.62 N
(Mean – SEM)
Small juveniles
n=3 Metamor-
phosis n=4 Larvae
(st.59) n=3
第 5 節 章括
第 2 節では、腹色を担う細胞について細胞形態学的な解析を行った。腹色を 構成する細胞は真皮最上層に位置するザンソフォアと呼ばれる色素細胞で、細胞 内部にはたくさんのカロテノイドの色素顆粒を有していることが分かった。この カロテノイド顆粒はリング型,均一型の 2 タイプの形態に分けられ、野生個体 ではリング型の色素顆粒がそのほとんどを占めた。食餌を限定して卵から人工飼 育したイモリでは均一型のカロテノイド顆粒がほとんどを占めた。
第 3 節では、形態学的なカロテノイドの観察状態を明らかとした。カロテノ イドの種類によっては、電子顕微鏡観察下で電子密度の高い物質として検出され、
ケト基を持つ遊離型のカロテノイドは溶出して包埋操作時に樹脂と置換される事 が考えられた。
第 4 節では、成長に伴うザンソフォアの色素顆粒の変化を形態学的に観察し た。一般的にザンソフォアは水溶性色素を含むプテリノソームと脂溶性色素を含 むカロテノイド顆粒を有することが多いが、飼育下のアカハライモリのザンソフ ォアは幼生期にはプテリノソームを有し、変態後、成長に伴ってカロテノイド顆 粒へ置き換わっていく、また、カロテノイド顆粒のサイズ、数が変化していくこ とが明らかになった。カロテノイド顆粒の数とサイズの変遷は、野生幼体におい ても同様の結果となった。
色素細胞による体色の発色機構は遺伝的に進化の道筋において決定されてきた ものである。アカハライモリの近縁種で色素細胞についての報告があるものには、
アルペンイモリSalamandra atra aurorae (Pederzoli and Trevisan 1990)、ブ チイモリNotophthalmus viridescence (Forbes et al. 1973)、があり、両者と も表皮性ザンソフォア(エリスロフォア)を有することが報告されている。ブチ イモリのエフトは背面全体が赤くなる。この時期のザンソフォアのカロテノイド 顆粒はリング型である。性成熟して体色が変化すると、ザンソフォアのリング型 カロテノイド顆粒は消失する(Forbes et al. 1973)。
第 4 章
腹色を形成する色素の化学的研究
第 1 節 研究の背景
第 3 章より、アカハライモリの腹色はザンソフォアによって発色し、細胞内 はカロテノイド顆粒で占められている事が明らかとなった。ザンソフォアにはか なりの量のカロテノイドが蓄積されている事が推察される。そこで本章では、ア カハライモリの腹色に対するカロテノイドの関わりを明らかにすることを目的と して、腹側皮膚におけるカロテノイド組成,量,他組織との組成差・分布量の差 異などについて高速液体クロマトグラフィー(HPLC)およびマススペクトルメ トリー(MS)を用いてカロテノイドの分析を行った。
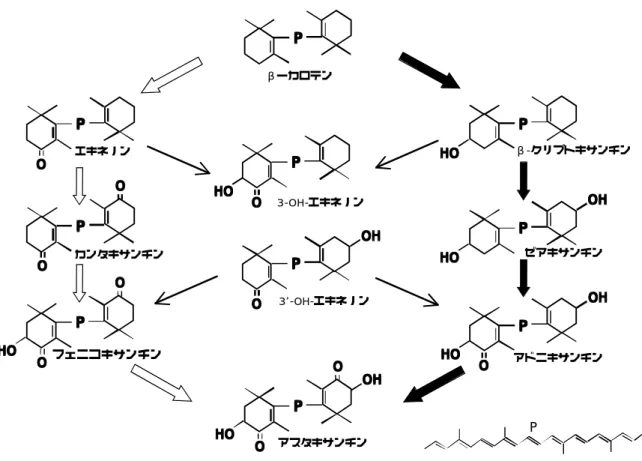
図 21 に代表的なカロテノイドの骨格と吸収スペクトルを示した。カロテノイ ドは 8 個のイソプレンが結合したテトラテルペノイドで、天然物だけで 700 種 以上が知られる。長い共役二重結合を持つため、一般に 400〜500 nm 付近の可 視光を吸収し黄色から赤を呈する。また、カロテノプロテインとの結合により青 や紫を発色する場合もある。色調はC鎖の共役二重結合の数で異なり、共役二重 結合が連なるものほど吸収波長は長波長に寄る。動物はカロテノイドを生合成で きないため、食餌から得る事になり、動物の持つカロテノイドの組成は餌となる 植物や動物、共生微生物の持つカロテノイドの種類の影響を受けることになる。
カロテノイドの両端の側鎖は 6 種類が知られている。α-エンドとε-エンドを 両端に有するカロテン(α-タイプとする)としてα-カロテン、両端にβ-エン ドを有するもの(β-タイプとする)にはβ-カロテンがあり、またそれぞれ側 鎖が酸化されたものをキサントフィルと称する。摂取されたカロテノイドはその まま、または酸化、還元、一部分の脱離など代謝を受けて体内移行し、肝臓や脂 肪組織、皮膚などに不均一に蓄積される(Britton 1983; Goodwin 1986; Parker 1996)。しかし、カロテノイドの組織移行、選択、蓄積の機構は明らかではない。
β- タイプカロテノイド
448
β-カロテン
470
ケトカロテノイド
α- タイプカロテノイド
442
ルテイン
β- タイプカロテノイド
OH R
アスタキサンチン
HO
O OH R HOO
図 22 緑藻類ヘマトコッカス藻におけるβ-カロテン 酸化経路
(Misawa et al. 1999)PPP P
PPPP OOO O
OHOHOH OH
PPP P OOO O
O O O O OHOHOH OH
HOHOHO HO PPP
P OOO O
PPP P OOO O
O O O O
PPP P OOO O
O O O O
HOHOHO HO
PPP P OOO HOHOHO O HO
PPP P HOHOHO
HO
PPP P OOO HOHOHO O HO
OHOHOH OH PPP
P
OHOHOH OH
HOHOHO HO βーカロテン
3-OH-エキネノン
3 -OH-エキネノン
アスタキサンチン エキネノン
カンタキサンチン
’
β-クリプトキサンチン
ゼアキサンチン
アドニキサンチン
P フェニコキサンチン
第 2 節 成体腹側皮膚におけるカロテノイドについて
腹側皮膚が含有しているカロテノイドの同定,組成,および量を解析すること を目的として、高速液体クロマトグラフィー、高性能分離用薄層クロマトグラフ ィー(以下 HPTLC)、およびマススペクトル測定を行った。
材料と方法
五島系統の成体イモリ雄 4 匹, 雌 4 匹の、下顎から総排泄孔までの腹側皮膚
(Mean±S.E.M. = 0.16±0.03 g)を切除後、分析時まで冷凍保存した。各 2 匹 ずつ、薄層クロマトグラフィー(TLC)分析、HPLC 分析にそれぞれ供した。
採材はイモリ採集後 2 週間以内に行った。TLC 分析はカロテノイド成分の予測 のために行った。以下 HPLC 分析の条件について述べる。抽出操作は暗室にお いて行い、抽出後直ちに分析した。抽出時に室温で解凍後、剪刀で細切しアセト ン抽出を抽出液の色が無色透明になるまで繰り返した。抽出液は共栓フラスコに 集め、37℃温湯下で減圧濃縮した。脱水のためにエタノールを加え、再び減圧 濃縮する操作を 2 回行った。粗抽出物を少量のクロロホルム-メタノール(3:1 v/v)に溶解、HPLC に供した。分析に使用した HPLC のシステムは HPLC の 条件を分析系 I として表 4 に示した。この条件は極性の高いカロテノイドから順 に分画されるため、キサントフィル類の良好な分離が得られることが報告されて いる(Takaichi and Ishidsu 1992)。
カラム: Bondapak C18 (25×100mm, Waters) 温度 :室温
流速 :2ml/min
移動相:100% methanol
検出 :Photo diode arraytectordi MCPD-3600, 大塚電子)(
表4 H PLCの条件I
µ
HPLC の各画分は既知カロテノイド標品との溶出時間の比較、画分の最大吸 収波長、吸収スペクトルの形状より推定し、MS による質量数データを合わせて 各カロテノイドを同定した。標品としてβ-カロテン(Wako)、アスタキサンチ ン、カンタキサンチン、ゼアキサンチン(ともに Roche)を用いた。試料は個 体別に一回の HPLC 分析に全量供し、各画分を分取した。β-カロテンより後に 溶出されてきたピークについてはカロテノイドエステルであると判断し、20- 27 分の画分と 32 分以降の画分の 2 つに分画した。個体別に分析した HPLC の結 果より、各画分の面積を測定し、分子吸光係数はアスタキサンチンε470 = 123/
cm/mM(Buchwald and Jencks 1968)を用いてカロテノイド量を定量した。
表 4 の条件ではカロテン類が同じ画分として溶出されるため、画分 7(14.5 分: β-カロテン画分)については分析系 II を用いて、更にカロテン同定のため の HPLC を行った。分析条件は表 5 に示した。この条件はカロテン類の分離が 良好な分析系である(Takaichi 2000)。
カロテノイドエステル画分については、Silica-gel HPTLC(Kieselgel 60, 10
×10 cm with concentrating zone 10×2.5 cm, MERCK)により精製した。展 開溶媒はアセトン – n - ヘキサン(15:85, v/v)で行い、展開後該当位置のバ ンドを削取し、フィルター(非水系フィルター 0.45 µm)を用いて溶出、回収 した。
カラム:Novapak C18 (8×100mm, Waters) 温度 :室温
流速 :2ml/min
移動相:1.75% methanol, 1.75% dichloromethane, 1.75%water,94.75% acetonitrile
検出 :Photo diode array ditector (MCPD-3600, 大塚電子)
表5 H PLCの条件II
MS 測定には Hitachi M-200 質量分析計を用いた。HPLC によって分画した 試料を画分毎に合一し、減圧濃縮後、少量のクロロホルム-メタノール(3:1, v/v)
に溶解した。試料を FD エミッターに風乾しながら充分量塗布し、MS に供した。
MS のイオン化には FD-MS(field - desorption mass spectrometry)を用い た。FD-MS ではカロテノイドのフラグメンテーションやエステル結合の開裂は 起きないことが既に分かっている(Takaichi 1993)。測定は加速電圧 4kv、ヒ ーティングカレントを 20 mA/10 min で上昇、ヒーティングカレント 10-15mA でカロテノイドが検出された。ポリエチレングリコールの質量数を補正コントロ ールとした。
カロテノイドエステルのカロテノイド部分は、ケン化によりカロテノイドと脂 肪酸塩に分離し、カロテノイドの質量数から同定した。ケン化は 6%水酸化カリ ウム/メタノール, 60℃10 分で行った。反応後、クロロホルムと水を加えて、脂 肪酸塩を水層に、カロテノイドをクロロホルム層に分離した。
ケトカロテノイドは必要に応じて、画分のα,β-タイプ属性を判断するため、
水素化ホウ素ナトリウム(NaBH4)を用いて還元試験を行った。
結果
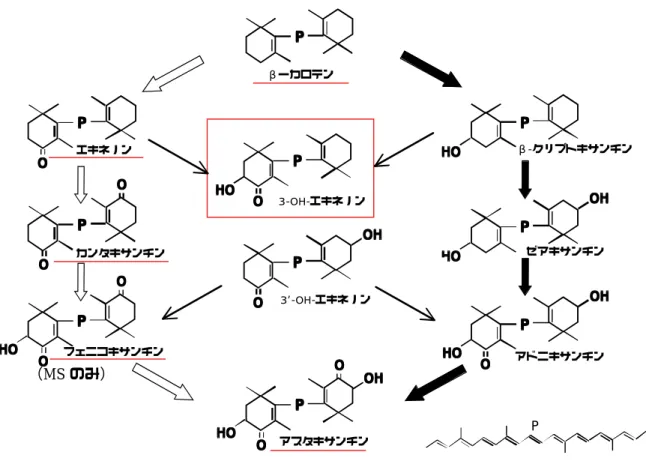
1.カロテノイドの同定
図 23 に成体アカハライモリの腹側皮膚カロテノイドの HPLC 分析結果の一例 を示した。また、図 24 には 16 分までに溶出したピークの拡大を示した。分析 の結果、9 種類のカロテノイドを分離した。画分の溶出時間、数、スペクトルの 形状は 4 個体とも同一で、各画分の量的な差はあるものの組成の個体差は見ら れなかった。MS 測定値と合わせ、各画分は次のように同定した。図 25˜27 に、
MS 測定値を示した。
画分 1: 溶出時間 3.3 分、スペクトル形状カルボニル型、λmax 470 nm(メタ ノール中)、質量数 596。標準試料と 3 項とも一致し、アスタキサンチンと同定 した。
画分 2: 溶出時間 3.9 分、スペクトル形状α型、λmax 422、442、470 nm、
質量数 568。溶出時間、質量数からはβ-タイプのゼアキサンチンまたはα-タ イプのルテインであると予想された。吸収スペクトルの形状は、第 3 ピークの 高さが第 2 ピークに対し 57%であった。メタノール溶媒中のルテインとゼアキ サンチンの吸収スペクトルの第 3 ピーク/ 第 2 ピークの割合の標準値はルテイ ン 61.7%、ゼアキサンチン 27.4%である(Takaichi and Shimada 1992)。従 って、画分 2 のスペクトル形状はルテインに近く、ルテインと同定した。
画分 3: 溶出時間 5.0 分、スペクトル形状カルボニル型、λmax 470 nm、質量 数 564。標準試料と 3 項とも一致し、カンタキサンチンと同定した。
画分 4: 溶出時間 6.2 分、スペクトル形状カルボニル型、λmax 470 nm、質量 数 566。NaBH4 還元試験によってβ-タイプであることが確認された。3’-OH エキネノンの溶出時間は 4.4 分であることから(Takaichi 2001)、ケト基と水 酸基は同側にあると考えられ、3-OH エキネノンと同定した。
画分 5: 溶出時間 8.1 分、スペクトル形状カルボニル型、λmax 470 nm、質量 数 550。NaBH4還元試験によってβ-タイプであることが確認された。よってエ キネノンと同定した。
画分 6: 溶出時間 10.0 分、スペクトル形状カルボニル型、λ 470 nm、微