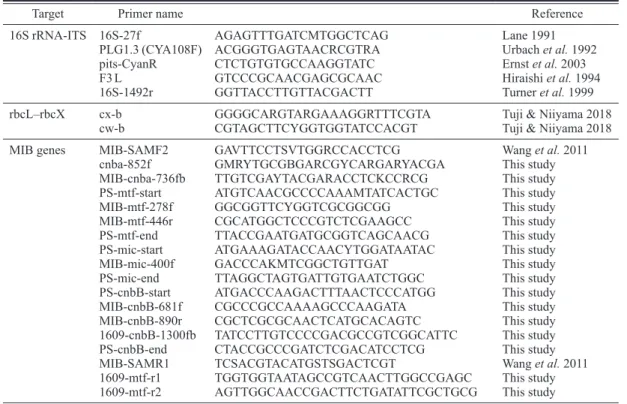
Table 2. Comparison of morphological, ecological and genetic characteristics of Micr ocoleus pseudautumnalis , M. autumnalis , and M. vaginatus . T axon names are iden - tical with thorse shown in Fig. 1 1. Taxa trichome(s) in sheath
cell width μm cell length μm cell L/W habitat motility
11 bp insert bad smell
reference M. pseudautumnalis 1 6.9–7.6(8.3) 2.0–5.0 0.5–2.5 planktic − + + this study M. autumnalis 1 4–7(9) 2–5(7) 0.5 or more on wet soil, mud, walls, rocks in rivers − Whitton 2002 (as “ Phormidium autum - nale ” ) M. autumnalis 1 (3.5)4–7 2–4(5) 0.5 periphytic on submersed substrate, in streams + − Komárek & Anagnostidis 2005 (as “ P. autumnale ” ) M. autumnalis 1 4–6 0.3–1 epipelic + − − Hašler et al. 2012 (as “ P. autumnale ” ) M. autumnalis 1 4.3–8.1 1.3–3.8 periphytic + − − Strunecký et al. 2013 M. vaginatus many (2.5)3–7 2–5(6.7) subaerophytic + − Komárek & Anagnostidis 2005 M. vaginatus many 3.5–7 0.5–1.5 on moist soil + − Whitton 2002 M. vaginatus many (4)5–7 0.3–1 epipelic + + − Hašler et al. 2012 M. vaginatus many 1.5–9.8 1.3–8.5
on various kinds of soils or walls, stream periphyton
+ + − Strunecký et al. 2013 M. vaginatus many (3.8)4.5–5.5 2–5(6.7) on arid soils ++ + − Boyer et al. 2002
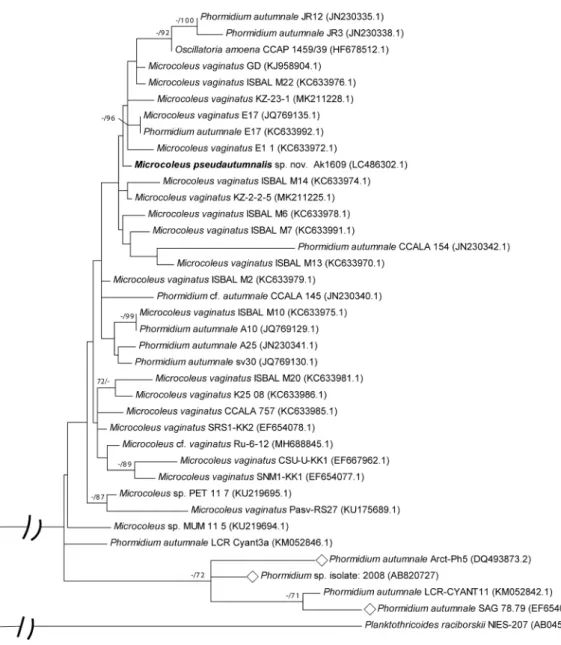
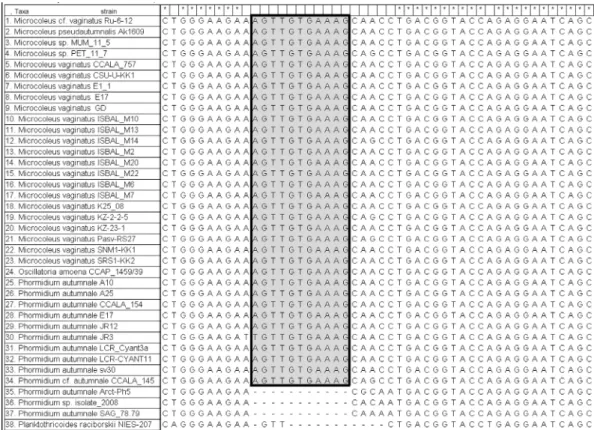
Table 3. The similarity (%) and the number of base dif ferences per partial 16S rRNA sequence (1036 positons) from 26 strains between sequences are shown. All positions containing gaps and missing data were eliminated.
1234567891011121314151617181920212223242526 1
Microcoleus pseudautumnalis Ak1609—99.9099.8199.8199.8199.8199.8199.8199.7199.7199.7199.7199.7199.5299.5299.5299.5299.4299.3299.2399.1398.9498.7598.6598.2691.99 2
Microcoleus vaginatus ISBAL M61—99.9099.9099.7199.7199.9099.7199.8199.6199.7199.8199.8199.6199.6199.6199.4299.3299.2399.3299.2399.0398.6598.7598.1791.99 3
Microcoleus vaginatus SRS1-KK221—100.0099.8199.81100.0099.8199.9099.7199.6199.9099.9099.5299.5299.7199.5299.2399.3299.2399.3299.1398.5598.6598.0792.08 4
Microcoleus cf.
vaginatus Ru-6-12210—99.8199.81100.0099.8199.9099.7199.6199.9099.9099.5299.5299.7199.5299.2399.3299.2399.3299.1398.5598.6598.0792.08 5
Microcoleus vaginatus E1 12322—100.0099.81100.0099.7199.9099.5299.9099.7199.3299.3299.7199.7199.2399.5299.0399.3299.1398.5598.4698.0792.18 6
Microcoleus vaginatus E1723220—99.81100.0099.7199.9099.5299.9099.7199.3299.3299.7199.7199.2399.5299.0399.3299.1398.5598.4698.0792.18 7
Microcoleus vaginatus ISBAL M14210022—99.8199.9099.7199.6199.9099.9099.5299.5299.7199.5299.2399.3299.2399.3299.1398.5598.6598.0792.08 8
Microcoleus vaginatus ISBAL M222322002—99.7199.9099.5299.9099.7199.3299.3299.7199.7199.2399.5299.0399.3299.1398.5598.4698.0792.18 9
Microcoleus vaginatus ISBAL M232113313—99.6199.5299.8199.8199.4299.6199.6199.4299.1399.2399.1399.2399.2398.6598.5598.1791.99 10
Microcoleus vaginatus KZ-23-1343311314—99.6199.8199.6199.2399.2399.6199.6199.1399.4299.1399.2399.0398.4698.3698.0792.18 11
Microcoleus vaginatus K25 083344554554—99.5299.5299.3299.3299.3299.2399.1399.0399.2398.9498.7598.4698.4698.0792.08 12
Phormidium autumnale sv3032111111225—99.8199.4299.4299.8199.6199.1399.4299.1399.4299.2398.4698.5597.9792.18 13
Phormidium cf.
autumnale CCALA 145321133132452—99.4299.4299.6199.4299.1399.2399.1399.2399.0398.4698.5597.9791.99 14
Microcoleus vaginatus SNM1-KK15455775768766—99.2399.2399.0398.9498.8498.9498.8498.6598.2698.3697.7891.60 15
Microcoleus sp. PET 11 754557757487668—99.2399.0399.1398.8499.3299.0399.0398.6598.5598.1791.99 16
Microcoleus vaginatus ISBAL M10543333334472488—99.4299.3299.2398.9499.6199.4298.6598.7598.1791.99 17
Oscillatoria amoena CCAP 1459/39565533536484610106—98.9499.8199.3299.0398.8498.4698.7598.3691.99 18
Microcoleus sp. MUM 11 56788888899999119711—98.7598.6599.5299.3299.1399.0398.6591.80 19
Phormidium autumnale JR127877557586106812128213—99.1398.8498.6598.4698.9498.5592.18 20
Phormidium autumnale CCALA 1548788101081099899117117149—98.5598.4698.1798.7598.2691.99 21
Phormidium sp. isolate: 200898777777881168121041051215—99.6199.0398.9498.5591.99 22
Phormidium autumnale LCR Cyant3a1110999999810138101410612714164—98.8499.1398.7592.18 23
Phormidium autumnale Arct-Ph51314151515151515141616161618141416916191012—98.7598.7591.31 24
Phormidium autumnale LCR-CYANT11141314141616141615171615151715131310111311913—99.4291.99 25
Phormidium autumnale SAG 78.7918192020202020201920202121231919171415181513136—91.80 26
Planktothricoides raciborskii NIES-20783838282818182818381828183878383838581838381908385—