博 士 学 位 論 文
神経前駆モデル細胞の神経様分化及び
前立腺癌細胞の神経内分泌様分化における
Ca
v3.2 T 型 Ca
2+チャネルの挙動と役割に関する研究
近 畿 大 学 大 学 院
薬学研究科薬科学専攻
深 海 和 樹
神 経 前 駆 モ デ ル 細 胞 の 神 経 様 分 化 及 び 前 立 腺 癌 細 胞 の 神 経 内 分 泌 様 分 化 に お け るCa v3 .2 T 型Ca 2+ チ ャ ネ ル の 挙 動 と 役 割 に 関 す る 研 究 深 海 和 樹博 士 学 位 論 文
神経前駆モデル細胞の神経様分化及び
前立腺癌細胞の神経内分泌様分化における
Ca
v3.2 T 型 Ca
2+チャネルの挙動と役割に関する研究
平 成 2 9 年 3 月近 畿 大 学 大 学 院
薬学研究科薬科学専攻
深 海 和 樹
目次
緒論 ... 1 第 1 章 H2S による Cav3.2 T 型 Ca2+チャネルの機能増強を介した NG108-15 細胞の神経様分化における Src キナーゼの関与 ... 3 Ⅰ 緒言 ... 3 Ⅱ 方法 ... 4 1)NG108-15 細胞の培養 ... 4 2)Whole-cell patch-clamp 法による Ba2+電流の測定 ... 4 3)形態学的検討 ... 5 4)Western blot 法 ... 6 5)試薬 ... 7 6)統計処理 ... 7 Ⅲ 結果 ... 8 1)未分化 NG108-15 細胞における NaHS 誘起 T-current 増強に対する ascorbic acid の抑制効果 ... 8 2)未分化 NG108-15 細胞における低濃度 NaHS の繰り返し添加及び ionomycin 処置による神経突起伸長 ... 8 3)未分化 NG108-15 細胞の NaHS 誘起神経突起伸長における T 型 Ca2+チ ャネル、HVA Ca2+チャネル、K ATPチャネル、TRPA1 チャネルの関与 ... 9 4)未分化 NG108-15 細胞における NaHS 及び ionomycin 誘起神経突起伸長 に対する Src、CaMKII、PKA、MEK 及び PI3K 阻害薬の抑制効果 ... 10 5)未分化 NG108-15 細胞の Src リン酸化レベルに対する NaHS の効果 12mibefradil 及び ascorbic acid による抑制効果... 13 7)NaHS 誘起 HVA Ca2+チャネル電流の増大に対する Src 阻害薬 PP2 の抑 制効果 ... 14 Ⅳ 考察 ... 15 第 2 章 神経内分泌様分化ヒト前立腺癌 LNCaP 細胞における H2S/Cav3.2 チャ ネルによる分泌反応制御 ... 19 Ⅰ 緒言 ... 19 Ⅱ 方法 ... 21 1)LNCaP 細胞の培養 ... 21 2)Whole-cell patch-clamp 法による Ba2+電流の測定 ... 21
3)Reverse transcription-polymerase chain reaction(RT-PCR)法 ... 22
4)形態学的検討 ... 23
5)MTT assay ... 24
6)Western blot 法 ... 24
7)siRNA による RNA 干渉法 ... 25
8)Prostatic acid phosphatase(PAP)assay ... 26
9)試薬 ... 26
10)統計処理 ... 27
Ⅲ 結果 ... 27
1)LNCaP 細胞における db-cAMP、IBMX、isoprenaline 及び IL-6 による神 経様突起伸長効果 ... 27
2)db-cAMP 誘起神経内分泌様分化 LNCaP 細胞におけるアンドロゲン非依 存的性質の獲得 ... 29 3)db-cAMP 誘起神経内分泌様分化 LNCaP 細胞における細胞増殖因子の発
現量増加 ... 30
4)LNCaP 細胞における db-cAMP 誘起神経内分泌様分化に伴った T-currents
の増大及びその増大に対する PKA 阻害薬の抑制効果 ... 31 5)LNCaP 細胞における isoprenaline 及び IL-6 誘起神経内分泌様分化に伴 った T-currents の増大 ... 33 6)LNCaP 細胞における抗アンドロゲン薬処置による T-currents の増大及 び神経突起伸長効果... 33 7)LNCaP 細胞における db-cAMP 誘起神経内分泌様分化に伴った Cav3.2 T 型 Ca2+チャネルの mRNA 及びタンパク発現増加 ... 34 8)db-cAMP 誘起神経内分泌様分化 LNCaP 細胞における H2S 合成酵素のタ ンパク発現量の検討... 35 9)db-cAMP 誘起神経内分泌様分化 LNCaP 細胞における H2S ドナーによる T-currents の増強とそれに対する T 型 Ca2+チャネル阻害薬の抑制効果 36
10)db-cAMP 誘起神経内分泌様分化 LNCaP 細胞に発現する T-currents に 対する H2S 合成酵素阻害薬の効果 ... 37
11)LNCaP 細胞の神経内分泌様分化に及ぼす H2S ドナー、CSE 阻害薬及
び T 型 Ca2+チャネル阻害薬の影響 ... 38
12)db-cAMP 及び IL-6 誘起神経内分泌様分化 LNCaP 細胞における prostatic
acid phosphatase(PAP)基礎分泌反応の増加... 39 13)db-cAMP 誘起神経内分泌様分化 LNCaP 細胞の PAP 基礎分泌反応の亢 進に対する H2S 合成酵素阻害薬の影響 ... 40 14)db-cAMP 誘起神経内分泌様分化 LNCaP 細胞の PAP 基礎分泌反応の亢
進に対する T 型 Ca2+チャネル阻害薬 mibefradil の影響 ... 41
ルのタンパク発現増加 ... 41
16)LNCaP 細胞における db-cAMP 誘起 Cav3.2 T 型 Ca2+チャネルタンパク 発現増加に対する Egr-1 のノックダウンの影響 ... 42
17)LNCaP 細胞における db-cAMP 処置による CREB のリン酸化増大 .. 43
18)LNCaP 細胞における REST のタンパク発現レベルに対する db-cAMP 処置の影響 ... 44 Ⅳ 考察 ... 45 第 3 章 ヒト前立腺癌由来 LNCaP 細胞の神経内分泌様分化に伴う Cav3.2 T 型 Ca2+チャネルの機能亢進に及ぼす高グルコースの影響:N 型糖鎖修飾の関与に ついて ... 50 Ⅰ 緒言 ... 50 Ⅱ 方法 ... 50 1) LNCaP 細胞の培養 ... 50 2) 形態学的検討 ... 51 3) Whole-cell patch-clamp 法 ... 51 4) Western blot 法 ... 51 5) 試薬 ... 52 6) 統計処理 ... 52 Ⅲ 結果 ... 52
1)LNCaP 細胞における db-cAMP 誘起 T-currents 増大及び神経様突起伸長 に対する高グルコースの影響 ... 52
2) LNCaP 細胞における db-cAMP 誘起 T-currents 増大及び神経様突起伸長 に対する N 型糖鎖生合成阻害薬 tunicamycin の効果... 53 3) LNCaP 細胞における db-cAMP 誘起 T-currents 増大に対する脱 N グリコ
シル化酵素 PNGaseF の効果 ... 55 4) db-cAMP 誘起神経内分泌様分化 LNCaP 細胞における Cav3.2 T 型 Ca2+ チャネルの発現増加及び Egr-1、REST、USP5 の発現に対する高グルコ ースの影響 ... 56 Ⅳ 考察 ... 57 総括 ... 61 引用文献 ... 64 略号一覧 ... 70 主論文 ... 72
緒論
電位依存性 Ca2+チャネルは、活性化される膜電位の違いにより低電位活性化 型(T 型)と高電位活性化型(N、P/Q、R、L 型)に分類される (1)。さらに T 型 Ca2+チャネルには、Ca v3.1、Cav3.2 及び Cav3.3 の 3 種類のアイソフォームが同 定されており、静止膜電位からわずかに脱分極することで活性化される性質か ら、主に神経興奮、ホルモンや神経伝達物質の分泌、筋収縮、細胞の増殖や分 化等の調節に寄与することが示されている (2)。 これら 3 つのアイソフォームの中で、特に Cav3.2 はその発現量や機能変化が 疾患への関連性を指摘する報告が数多く存在する (1)。興味深いことに、川畑ら のグループは、Cav3.2 のチャネル機能は、第 3 の生体内ガスメッセンジャーと して注目されている硫化水素(H2S)により増強されることを見出し (3-5)、さ らにこの反応が体性痛や神経損傷や抗癌剤による神経障害性疼痛及び内臓痛の 発症に寄与すること (3, 6-10)や神経前駆細胞の神経様分化を誘起することを明 らかにしている (11)。また、他のグループの報告により、ある種の癌細胞にお いて Cav3.2 を含む T 型 Ca2+チャネルや (12)、H2S 合成酵素 (13)の発現増加が認 められ、これらの発現増加が癌増殖等に関与することが示唆されていることか ら、癌においても H2S/Cav3.2 系が何らかの役割を担っていると考えられる。また、Cav3.2 のチャネル機能は、Ca2+/calmodulin-dependent protein kinase II(CaMKII)
や protein kinase A(PKA)等によるリン酸化調節 (14)や、asparagine 結合型(N 型)糖鎖修飾によって調節されている (15)。2 型糖尿病モデルマウスにおいては、
N 型糖鎖修飾の亢進よって Cav3.2 の膜発現の増加及びチャネル機能の増強がお
こり、これがニューロパチーの発症に寄与する可能性が指摘されている (16)。
一方、Cav3.2 のタンパク発現量は転写促進因子の 1 つである early growth response
transcription factor(REST)により抑制的に制御されている (17)。また、脱ユビ
キチン化酵素の一つである ubiquitin-specific peptidase 5(USP5)は、Cav3.2 のユ
ビキチン化を抑制することで、このチャネルのプロテアソーム分解を抑制し、 Cav3.2 のタンパク発現量の維持・増加に関与する (18)。 そこで本研究では、H2S 及び N 型糖鎖修飾による Cav3.2 T 型 Ca2+チャネルの 機能調節や Egr-1、REST 及び USP5 によるチャネル発現調節に焦点をあて、神 経前駆細胞の神経様分化及び前立腺癌細胞の神経内分泌様分化における Cav3.2 の挙動と役割について明らかにする事を目的として検討を行った。第 1 章では 神経前駆 NG108-15 細胞の H2S ドナーNaHS による Cav3.2 機能増強を介した神経 様分化に関与する細胞内シグナルについて検討した。第 2 章では前立腺癌のホ ル モ ン 療 法 抵 抗 性 獲 得 に 関 わ る 神 経 内 分 泌 様 分 化 前 立 腺 癌 細 胞 に お け る H2S/Cav3.2 系の分泌機能調節への関与について検討を実施した。第 3 章では、神 経内分泌様分化前立腺癌細胞に発現する Cav3.2 の機能が、細胞外グルコース濃 度上昇に伴う N 型糖鎖修飾の亢進により増強されるかについて検討を行った。
第 1 章 H
2S による Ca
v3.2 T 型 Ca
2+チャネルの機能
増強を介した NG108-15 細胞の神経様分化における
Src キナーゼの関与
Ⅰ 緒言
硫化水素(H2S)は、一酸化窒素(NO)や一酸化炭素(CO)に次ぐ第 3 のガ ス 状 メ ッ セ ン ジ ャ ー で あ る 。 生 体 内 に お い て は 主 に 、 L-cysteine か ら cystathionine--lyase(CSE)、cystathionine--synthase(CBS)、3-mercaptopyruvatesulfurtransferase(3-MST)/ cysteine aminotransferase(CAT)等の酵素により合成 される。さらに、最近、D-cysteine から 3-MST/D-amino acid oxidase(DAO)に
よって H2S が生成される新たな経路が発見され注目されている (19)。H2S の標 的分子は数多く報告されており、特にイオンチャネルの中では ATP 感受性 K+ チャネル(KATPチャネル)、T 型 Ca2+チャネル、L 型 Ca2+チャネル、TRPA1 チャ ネル、大コンダクタンス Ca2+活性化 K+(BK)チャネル等がある (20)。川畑ら のグループは、H2S が T 型 Ca2+チャネルのアイソフォームの一つである Cav3.2 T 型 Ca2+チャネルの機能増強を介して、体性痛及び内臓痛を促進するほか、神経 障害性疼痛の病態に関与することを明らかにしている (3, 7-10, 21)。 一方、神経モデル細胞である NG108-15 細胞において細胞内 cyclic AMP (cAMP)アナログである dibutyryl cAMP(db-cAMP)処置により誘起される神
経様分化に Cav3.2 T 型 Ca2+チャネルの機能増強が寄与することが報告されてい
る (22)。さらに、H2S ドナーである NaHS が NG108-15 細胞に発現する T 型 Ca2+
チャネルの活性を増強することで 、神経突起伸長や高電位活性化型(high
することを明らかにしているが (11)、その詳細な分子メカニズムは未解明のま まであった。そこで本章では、NG108-15 細胞における NaHS 誘起神経様分化に 関わる細胞内シグナルを、電気生理学的、形態学的及び生化学的手法を用いて 解析した。
Ⅱ 方法
1)NG108-15 細胞の培養 マウスの神経芽細胞腫(neuroblastoma)とラットのグリア細胞腫(glioma)と のハイブリット細胞である NG108-15 細胞を 10% fetal bovine serum(FBS) (Thermo Electron Corporation、Waltham、MA、USA)、0.1 mM hypoxanthine(Gibco、Carlsbad、CA、USA)、16 µM thymidine(Gibco)、1 µM aminopterin(Gibco)、50
unit/mL penicillin(Gibco)、50 µg/mL streptomycin(Gibco)を含む high glucose の
Dulbecco’s Modified Eagle’s Medium(DMEM)培地(Sigma-Aldrich、St. Loius、 MO、USA または和光純薬工業、大阪、日本)中で tissue culture dish(100 × 20 mm)
を用いて、37 ˚C、5% CO2の条件下で培養した。
2)Whole-cell patch-clamp 法による Ba2+電流の測定
NG108-15 細胞を poly-L-ornithine(Sigma)でコーティングした tissue culture dish
(35 × 10 mm)に 1.0 × 104 cells/dish で播種し、上述の DMEM 培地にて 1 日間培
養した細胞を実験に用いた。細胞外溶液には 10 mM BaCl2、10 mM HEPES、5.6
mM glucose に 、 Na+チ ャ ネ ル を 介 す る 電 流 を 阻 止 す る た め に 97 mM
N-methyl-D-glucamine (NMDG)、及び K+チャネルを介する電流を阻止するため
を介する電流を阻止するために 150 mM CsCl、を加え、pH を 7.2 に調整したも
のを使用した。Ba2+電流は、室温 (20-24 ˚C)で測定した。Ba2+電流の測定には、
Narahashi らによって設計された増幅器を用いた。測定の際の series-resistance compensation(Rs Comp)は 80% に、sampling interval は 5 kHz に設定し、保持 電位は-90 mV にした。データは、デジタイザー(Digidata 1322A、Axon Instruments、 Sunnyvale、CA、USA)を経由してデジタル化したものをコンピュータに入力さ
せ、pClamp8 software(Axon Instruments)を用いて解析した。T 型 Ca2+チャネル
依存性電流(T-currents)は、保持電位-90 mV から-20 mV のテスト刺激を与えた
時の Ba2+電流を測定し、この電流のピーク電流から 150 ms 後の電流を差し引い
た値として算出した。また HVA Ca2+チャネル電流(HVA-currents)は、保持電位
-90 mM から+10 mV のテスト刺激を与えた時に得られる電流において、T-currents
の影響が少ない刺激開始 75ms 後の電流として算出した。なお、T-currents の NaHS による増強効果に対する ascorbic acid の影響を検討する際は、control の T-currents を測定後、ascorbic acid を細胞外液へ添加し、10 秒後に NaHS を細胞外液へ添加 してから 2 分後に再び同じ細胞において T-currents を測定した。川畑らのグルー プが報告している NaHS 処置による HVA-currents の増大効果に対する Src 阻害薬 PP2 の抑制効果を検討する際は、PP2 を培養液に添加して 30 分後に NaHS(13.5 mM)を作用させ、NaHS 刺激から 2 日後に HVA-currents を測定した。
3)形態学的検討
NG108-15 細胞を poly-L-ornithine でコーティングした tissue culture dish(35 × 10
mm)に 1.0 × 104 cells/dish で播種し、1% FBS を含む上記の DMEM 培地にて 2-3
時間培養した後、NaHS、ionomycin 及び db-cAMP で刺激した。刺激開始 16 時間 後、光学顕微鏡下で観察し、細胞体の直径よりも長い神経突起をもつ細胞数を
計測し、視野内の全細胞数に対する割合を求めた。また、NaHS(1.5 mM)を 30 分間隔で複数回処置する場合、初回刺激時から 16 時間後に神経突起伸長効果を 評価した。なお、阻害剤はすべて刺激開始 30 分前に添加した。
4)Western blot 法
NG108-15 細胞は tissue cluture dish(100 × 20 mm)に 1.0 × 106 cells/dish または
6-well plate に 12 × 104 cells/well となるように播種し、1% FBS を含む上記の
DMEM 培地で 2 時間培養した。NaHS または db-cAMP で刺激後、sodium dodecyl
sulfate(SDS) buffer(62.5 mM Tris-HCl(pH 6.8)、2% SDS、10% glycerol)50 µL/dish
または 30 µL/well で溶解し、cell scraper を用いて 1.5 mL エッペンチューブに回 収した。回収した細胞溶解液を液体窒素及び超音波で処理した後、Lowry 法によ りサンプルのタンパク量を定量した。サンプル中のタンパク質量を 20 または 30 µg に調製した後、12.5% SDS-polyacrylamide gel を用いて電気泳動によりタンパ クを分離させた。分離させたタンパクを polyvinylidene difluoride (PVDF)膜 (Immobilon-P、Millipore Corporation、Billerica、MA、USA)へ転写し、抗 Src kinase 抗体 (Cell Signaling Technology、Beverly、MA、USA)及び抗 phospho-Src family kinase(Tyr416)抗体(Cell Signaling Technology)を、5% BSA、TBS、0.1% Tween-20 でそれぞれ 2000 倍及び 1000 倍に希釈して 4 ℃で一晩反応させた。次に、二次 抗体として HRP-linked anti-rabbit IgG 抗体(Chemicon International、CA、USA) を blocking buffer(TBS、0.1% Tween-20、5% w/v nonfat dry milk)で 1000 倍希釈 し、室温で 60 分反応させ、enhanced chemiluminescence(ECL)(Amersham Bioscuences、 Little Chalfont、Buckinghamshire、UK)で 1 分間反応させて発光さ せることにより、60 kDa に Src 及びリン酸化 Src のバンドを検出した。検出され たバンドは、Win Roof(三谷商事、福井、日本)を用いて数値化した。BAPTA/AM、
mibefradil 及び ascorbic acid は、NaHS 刺激の 30 分前から作用させ、その影響を 検討した。また、NaHS(1.5 mM)を 30 分間隔で複数回処置する場合、最終刺 激 30 分後にサンプルを回収した。
5)試薬
実験に使用した試薬は以下のとおりである。
NaHS はキシダ化学(大阪、日本)から購入した。L-ascorbic acid、db-cAMP、 mibefradil、nitrendipin、glibenclamide、KT5720、U0126 及び genistein は Sigma-Aldrich から購入した。Ionomycin、PP2、KN-93、BAPTA/AM は Calbiochem(Darmstadt、 Germany)から購入した。LY294002 は Tocris Cookson Ltd.(Ballwin、MO、USA) から購入した。 ω-conotoxin GVIA 及び ω-conotoxin MVIIC はペプチド研究所(大 阪、日本)から購入した。Allyl isothiocyanate(AITC)は東京化成工業(東京、 日本)から購入した。AP-18 は Enzo Life Sci. Inc.(Farmingdale、NY、USA)か ら購入した。NaHS、db-cAMP、ascorbic acid、mibefradil、ω-conotoxin GVIA 及び ω-conotoxin MVIIC は滅菌蒸留水に溶解して使用した。その他の試薬は dimethyl
sulfoxide(DMSO)(Sigma)で溶解して使用した。
6)統計処理
得られた実験値は、平均値±標準誤差で示した。有意差の検定には t 検定及びチ ューキー法を用い P<0.05 で有意差ありとした。
Ⅲ 結果
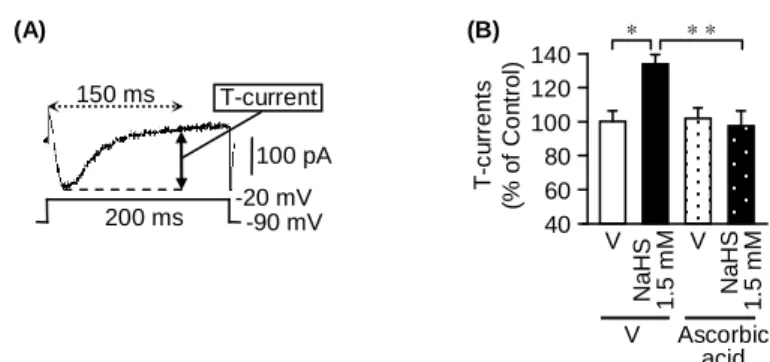
1)未分化 NG108-15 細胞における NaHS 誘起 T-current 増強に対する ascorbic acid の抑制効果
未分化 NG108-15 細胞に対して whole-cell patch-clamp 法により保持電位-90 mV から-20 mV のテスト刺激を 200 ms 間与えると、一過性の内向き電流が見られた
(Fig. 1A)。この電流は T 型 Ca2+チャネル阻害薬の mibefradil により完全に抑制
されることが報告されていることから、T 型 Ca2+チャネル依存性電流(T-currents) であることが確認されている (3)。未分化 NG108-15 細胞におけるこの T-currents は、H2S ドナーである NaHS を 2 分間作用させることにより増大した。(Fig. 1B) 一方、NaHS 誘起 T-currents 増強は、T 型 Ca2+チャネルのアイソフォームのうち Cav3.2 を選択的に阻害することが知られている ascorbic acid(1 µM)(23) の前処 置によりほぼ完全に消失した。 Ascorbic acid V N a H S 1 .5 mM V T -c u rr e n ts (% o f Co n tr o l) 40 60 80 100 120 140 * * * N a H S 1 .5 mM V (A) 100 pA 150 ms T-current -20 mV -90 mV 200 ms (B)
Fig. 1. NaHS-evoked facilitation of T-type Ca2+ channel-dependent currents (T-currents) and its reversal by
ascorbic acid in undifferentiated NG108-15 cells. (A) T-type Ca2+ channel-dependent-currents (T-currents) were determined as the difference between the peak current and the current 150 ms after beginning of the test pulse at -20 mV from a holding potential of -90 mV. (B) After the control T-current was measured, NaHS was applied at 2 min. Ascorbic acid, known to selectively inhibit Cav3.2 among three forms of T-type Ca2+ channels, was applied 10 s before addition of NaHS at 1.5 mM or vehicle. Data show the mean with S.E.M. from 4-10 (B) different cells. * p < 0.05, ** p < 0.01. V, vehicle.
2)未分化 NG108-15 細胞における低濃度 NaHS の繰り返し添加及び ionomycin 処置による神経突起伸長
ことで、神経突起伸長が促進されることが報告されている (11)。一方、H2S は 気体であるため、NaHS を培養液中に添加すると生成した H2S が急速に空気中へ 拡散・消失していくことが報告されている (24)。そこで、単回の添加では有意 な効果を示さない NaHS(1.5 mM)を 30 分間隔で 3 回もしくは 5 回繰り返し添 加した。その結果、NaHS(1.5 mM)を 3 回添加しても突起伸長促進効果は見ら れなかったが、5 回繰り返し添加した場合、高濃度の NaHS(13.5 mM)を単回 添加した場合と同様の突起伸長が誘起された(Fig. 2A)。さらに興味深いことに、 Ca2+イオノフォアである ionomycin を処置することによっても、NaHS 処置と同 程度の神経突起伸長が誘起された(Fig. 2B)。 30-min intervals V 3 Na HS 1 .5 mM 3 V 5 V e h icle 1 (A) (B) n.s. 0 10 20 30 Na HS 1 .5 mM 5 Na HS 1 3 .5 mM 1 * * * * V 0.1 0.3 Ionomycin (µM) * * * P ro p o rt io n o f ce lls wi th n e u ri te s (% )
Fig. 2. Effect of repetitive application of NaHS at a low concentration or ionomycin on neurite outgrowth in NG108-15 cells. Proportion of cells with neurites longer than the cell diameter was determined 16 h after the onset of stimulation with NaHS or ionomycin. (A) NaHS was applied at 13.5 mM once or applied at 1.5 mM repetitively at 30-min intervals 3 or 5 times in total. (B) Ionomycin, a Ca2+ ionophore, was applied once to the cells. Data show the mean with S.E.M. from 4-12 (A) or 4-8 (B) different experiments. * p < 0.05, ** p < 0.01. V, vehicle; n.s., not significant. 3)未分化 NG108-15 細胞の NaHS 誘起神経突起伸長における T 型 Ca2+チャネ ル、HVA Ca2+チャネル、K ATPチャネル、TRPA1 チャネルの関与 川畑らのグループは、未分化 NG108-15 細胞における NaHS 誘起神経突起伸長 は T 型 Ca2+チャネルの 3 つのアイソフォームのうち Ca v3.2 を選択的に阻害する
ascorbic acid (23) 及び Zn2+の前処置により抑制されることから、Cav3.2 チャネル
ル以外に、HVA Ca2+チャネル、K
ATPチャネル、TRPA1 チャネル等にも作用する
ことが報告されている (24-26)。そこで、各チャネルの阻害薬や活性化薬を用い て NG108-15 細胞の NaHS 誘起神経突起伸長に関与するイオンチャネルを検討し
た。その結果、NaHS による神経突起伸長は T 型 Ca2+チャネル阻害薬 mibefradil
により有意に抑制されたが(Fig. 3A)、N 型、P/Q 型及び L 型の HVA Ca2+チャネ
ル阻害薬混合液(ω-conotoxin GVIA、ω-conotoxin MVIIC 及び nitrendipin を含む)
や KATPチャネル阻害薬 glibenclamide は無効であった(Fig. 3B、C)。また、TRPA1
チャネル阻害薬 AP-18 は、NaHS 誘起神経突起伸長に対してわずかに抑制傾向を 示したが、TRPA1 チャネル活性化薬 AITC によって神経突起伸長は誘起されな かった(Fig. 3D、E)。 0 4 12 16 20 8 AP-18 15 µM (D) V V N a H S 1 3 .5 mM N a H S 1 3 .5 mM V * * n.s. (A) V Mibefradil 0.3 µM V V N a H S 1 3 .5 mM N a H S 1 3 .5 mM 0 10 20 30 * * * * P ro p o rt io n o f c e lls w it h n e u ri te s (% ) V N a H S 1 3 .5 mM N a H S 1 3 .5 mM n.s. HVA inhibitors (B) * * V 0 10 20 30 Gliben 30 µM (C) V N a H S 1 3 .5 mM N a H S 1 3 .5 mM V n.s. * * 0 10 20 30 V 1 10 AITC (µM) n.s. n.s. (E) 0 4 12 16 20 8
Fig. 3. Mibefradil-sensitive neurite outgrowth caused by NaHS in NG108-15 cells. Mibefradil, a pan-T-type Ca2+ channel inhibitor (A), an inhibitor mixture of HVA Ca2+ channels (HVA inhibitors, -conotoxin GVIA at 1 µM for N-type; -conotoxin MVIIC at 1 µM for P/Q-type; nitrendipine at 1 µM for L-type Ca2+ channels) (B), glibenclamide (Gliben), a KATP channel inhibitor (C), or AP-18, a TRPA1 inhibitor (D), were added 30 min before stimulation with NaHS. (E) Allyl isothiocyanate (AITC), an agonist of TRPA1, had no increase in neurite outgrowth. Data show the mean with S.E.M. from 6 (A, E), 10 (B, C), 7-9 (D) different experiments. ** p < 0.01. V, vehicle; n.s., not significant.
4)未分化 NG108-15 細胞における NaHS 及び ionomycin 誘起神経突起伸長に対 する Src、CaMKII、PKA、MEK 及び PI3K 阻害薬の抑制効果
神経由来 PC12 細胞において脱分極により誘起される神経分化には、細胞内
(27)。そこで、NG108-15 細胞における NaHS 誘起神経突起伸長に同様の機序が 関与する可能性を検討する為、Src 阻害薬 PP2 の効果を検討したところ、NaHS 及び ionomycin 誘起神経突起伸長は PP2 により完全に抑制された(Fig. 4A、B)。 また、PP2 は細胞膜透過性 cAMP アナログである db-cAMP により誘起される神 経突起伸長を部分的に抑制した(Fig. 4C)。さらに、NaHS 誘起神経突起伸長に おけるその他のシグナル分子の関与について各種阻害薬を用いて検討したとこ ろ、calmodulin-dependent protein kinase II(CaMKII)阻害薬 KN-93、protein kinase
A(PKA)阻害薬 KT5720 は、NaHS 誘起神経突起伸長に影響を与えなかった(Table.
1)。また、KN-93 は db-cAMP 誘起神経突起伸長に対しても影響を与えなかった
(Table. 1)。MAPK/ERK kinase(MEK)阻害薬 U0126、phosphatidylinositol 3-kinase (PI3K)阻害薬 LY294002 及び非選択的 tyrosine kinase 阻害薬 genistein は、NaHS による刺激のない状態で、単独添加により神経突起伸長を促進した(Table. 1)。 (A) 0 10 20 30 40 50 PP2 1 µM V Na HS 1 3 .5 mM V Na HS 1 3 .5 mM V * * * * Io n o m y c in 0 .1 µ M (B) 0 5 10 15 20 25 V Io n o m y c in 0 .1 µ M V PP2 1 µM V * * * * (C) 0 10 20 30 40 50 db -cA M P 1 mM V db -cA M P 1 mM V * * * * * * PP2 1 µM V P ro p o rt io n o f ce lls wi th n e u ri te s (% )
Fig. 4. Effects of PP2, a Src inhibitor, on the neurite outgrowth induced by NaHS, ionomycin or db-cAMP in NG108-15 cells. PP2 was added 30 min before stimulation with NaHS, ionomycin or dibutyryl cAMP (db-cAMP) for 16 h. Data show the mean with S.E.M. from 8 (A), 8-12 (B) or 14 (C) different experiments. ** p < 0.01. V, vehicle.
CaM kinase II
-Non-selective tyrosine kinase PI3 kinase
MEK PKA
CaM kinase II
-Non-selective tyrosine kinase PI3 kinase MEK PKA CaM kinase II -Target of Inhibitor 39.4 2.3 ** KN-93 10 µM db-cAMP 1 mM 42.0 2.6 ** Vehicle (DMSO) db-cAMP 1 mM 43.7 0.8 ** Genistein 30 µM NaHS 13.5 mM 32.2 4.1 LY294002 10 µM NaHS 13.5 mM 46.8 2.4 **, † U0126 10 µM NaHS 13.5 mM 27.1 4.1 KT5720 1 µM NaHS 13.5 mM 34.4 7.2 * KN-93 10 µM NaHS 13.5 mM 34.3 2.1 ** Vehicle (DMSO) NaHS 13.5 mM 45.7 1.9 ** Genistein 30 µM Vehicle (water) 34.7 1.7 ** LY294002 10 µM Vehicle (water) 44.9 1.7 ** U0126 10 µM Vehicle (water) 16.9 1.8 KT5720 1 µM Vehicle (water) 25.9 4.0 KN-93 10 µM Vehicle (water) 20.3 1.5 Vehicle (DMSO) Vehicle (water) Proportion of cells with neurites (%) Inhibitors Stimulators CaM kinase II
-Non-selective tyrosine kinase PI3 kinase
MEK PKA
CaM kinase II
-Non-selective tyrosine kinase PI3 kinase MEK PKA CaM kinase II -Target of Inhibitor 39.4 2.3 ** KN-93 10 µM db-cAMP 1 mM 42.0 2.6 ** Vehicle (DMSO) db-cAMP 1 mM 43.7 0.8 ** Genistein 30 µM NaHS 13.5 mM 32.2 4.1 LY294002 10 µM NaHS 13.5 mM 46.8 2.4 **, † U0126 10 µM NaHS 13.5 mM 27.1 4.1 KT5720 1 µM NaHS 13.5 mM 34.4 7.2 * KN-93 10 µM NaHS 13.5 mM 34.3 2.1 ** Vehicle (DMSO) NaHS 13.5 mM 45.7 1.9 ** Genistein 30 µM Vehicle (water) 34.7 1.7 ** LY294002 10 µM Vehicle (water) 44.9 1.7 ** U0126 10 µM Vehicle (water) 16.9 1.8 KT5720 1 µM Vehicle (water) 25.9 4.0 KN-93 10 µM Vehicle (water) 20.3 1.5 Vehicle (DMSO) Vehicle (water) Proportion of cells with neurites (%) Inhibitors Stimulators
Table 1. Effects of various inhibitors on the NaHS-induced neurite outgrowth in NG108-15 cells. Cells were stimulated with NaHS, dibutyryl cAMP (db-cAMP) or vehicle (water) for 16 h in the absence or presence of inhibitors. Inhibitors were applied 30 min before stimulation with NaHS or db-cAMP. Data show the mean with S.E.M. from 6-18 different experiments. * p < 0.05, ** p < 0.01 vs. vehicle + vehicle, †p < 0.05 vs. vehicle + NaHS.
Statistical analysis was performed by using the Tukey's test.
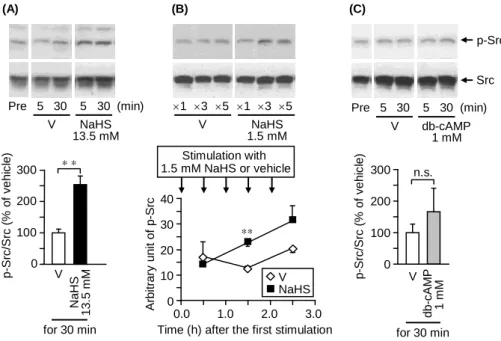
5)未分化 NG108-15 細胞の Src リン酸化レベルに対する NaHS の効果
NaHS 誘起神経突起伸長が Src 阻害薬によって阻害されたことより、次に NaHS
が Src のリン酸化を誘起するか否かを検討した。その結果、NG108-15 細胞の Src リン酸化レベルは高濃度の NaHS(13.5 mM)処置 30 分後において、vehicle 処 置群と比べて有意に増加していた(Fig. 5A)。さらに、低濃度の NaHS(1.5 mM) を繰り返し処置することによっても、Src リン酸化は増加した(Fig. 5B)。一方、
(B) (C) 0 100 200 300 Na HS 1 3 .5 mM V p -S rc/ S rc (% o f ve h ic le ) for 30 min (A) A rb itr a ry u n it o f p -S rc 0 0.0 1.0 2.0 3.0 V NaHS
Time (h) after the first stimulation Stimulation with 1.5 mM NaHS or vehicle 10 20 30 40 ** 0 100 200 300 db -cA M P 1 mM V p -S rc/ S rc (% o f ve h ic le ) for 30 min 1 3 5 V NaHS 1.5 mM Pre 5 30 V 5 30 NaHS 13.5 mM (min) p-Src Src 1 3 5 Pre 5 30 V 5 30 db-cAMP 1 mM (min) * * n.s.
Fig. 5. Phosphorylation of Src in response to NaHS, but not dibutyryl cyclic AMP (db-cAMP), in NG108-15 cells. The top panels show the representative western blots and the bottom panels show the quantified data by densitometry. (A) The cells were stimulated with NaHS at 13.5 mM or vehicle (V) for 5 or 30 min. (B) NaHS at 1.5 mM were repetitively applied at 30 min intervals, 1, 3 or 5 times in total, and the cells were collected for the analysis 30 min after the final application. (C) The cells were stimulated with db-cAMP at 1 mM for 5 or 30 min. Data show the mean with S.E.M. from 4 (A), 3 (B) or 8 (C) different experiments. ** p < 0.01. V, vehicle; n.s., not significant.
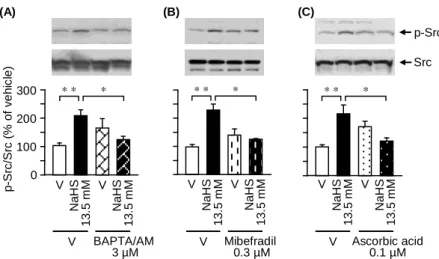
6)未分化 NG108-15 細胞の NaHS 誘起 Src リン酸化に対する BAPTA/AM、 mibefradil 及び ascorbic acid による抑制効果
NaHS 誘起 Src リン酸化に Cav3.2 チャネルを介した Ca2+流入が関与するか否
かを調べるため、細胞内 Ca2+キレーターの BAPTA/AM、mibefradil 及び ascorbic
acid の影響を検討した。その結果、NaHS により誘起されたリン酸化 Src の増加 は、BAPTA/AM、mibefradil あるいは ascorbic acid の前処置により完全に抑制さ れた(Fig. 6)。
V N a H S 1 3 .5 mM V V Ascorbic acid 0.1 µM V Mibefradil 0.3 µM V BAPTA/AM 3 µM N a H S 1 3 .5 mM (A) (B) (C) p -S rc/ S rc (% o f v e h ic le ) p-Src Src 0 100 200 300 * * * V N a H S 1 3 .5 mM V N a H S 1 3 .5 mM * * * * * * V N a H S 1 3 .5 mM V N a H S 1 3 .5 mM
Fig. 6. Effects of BAPTA/AM, mibefradil or ascorbic acid on the NaHS-caused phosphorylation of Src in NG108-15 cells. The cells were stimulated with NaHS for 30 min in the absence or presence of BAPTA/AM, an intracellular Ca2+ chelator, mibefradil, a T-type Ca2+ channel inhibitor, or ascorbic acid, known to selectively inhibit Cav3.2 among three T-type Ca2+ channel isoforms. All three reagents were added 30 min before stimulation with NaHS. Data show the mean with S.E.M. from 3-6 different experiments. * p < 0.05, ** p < 0.01. V, vehicle.
7)NaHS 誘起 HVA Ca2+チャネル電流の増大に対する Src 阻害薬 PP2 の抑制効 果 未分化 NG108-15 細胞は、細胞内 cAMP 増加により誘起される分化によって神 経突起伸長に加えて、HVA Ca2+チャネル電流(HVA-currents)の増大が誘起され ることが報告されている (22)。同様に、川畑らのグループも、未分化 NG108-15 細胞を NaHS で 2 日間処置すると神経突起伸長の亢進とともに、HVA-currents の増大が見られることを報告している (11)。そこで、whole-cell patch-clamp 法に て NG108-15 細胞における NaHS 誘起 HVA-currents 増大に対する Src 阻害薬 PP2 の効果について検討した。Fig. 7A に示すように、NG108-15 細胞において、保持 電位を-90 mV に設定し、-120 mV から+40 mV のランプ刺激を与えると、-30 mV 付近で T-currents、+10 mV 付近で HVA-currents と考えられる膜電流が見られ、 後者は 2 日間 NaHS を処置することにより明らかに増大した。この NaHS 処置後 の+10 mV における膜電流の増大は、Src 阻害薬 PP2 を前処置することで消失し た(Fig. 7A)。さらに Fig. 7B に示すように、保持電位-90 mV から+10 mV のパ
ルス刺激を与えた時に得られる電流において、T-currents の影響が少ない刺激開 始 75ms 後の電流を HVA-currents として算出し、その変化を調べたところ、NaHS 処置により HVA-currents の有意な増大が確認され、その効果は PP2 前処置によ り完全に抑制された(Fig. 7C)。 -120 mV +40 mV 850 ms Ca lc iu m c h a n n e l-d e p e n d e n t c u rr e n ts ( nA ) Ramp pulse (mV) -0.10 -0.05 0.00 0.05 0.10 -120 -100 -80 -60 -40 -20 0 20 40 DMSO + Water DMSO + NaHS PP2 + Water PP2 + NaHS HP: -90 mV (A) V Na HS 1 3 .5 mM V Na HS 1 3 .5 mM V PP2 (B) HV A cu rr e n ts ( nA ) 0.00 0.02 0.04 0.06 0.08 0.10 (C) +10 mV 200 ms HP: -90 mV 75 ms HVA current 0.05 nA * * *
Fig. 7. Effects of PP2 on the upregulation of HVA Ca2+ channel-dependent currents in NG108-15 cells exposed
to prolonged treatment with NaHS. The cells were treated with NaHS at 13.5 mM for 2 days in the absence or presence of PP2, a Src kinase inhibitor, at 1 µM. PP2 was applied 30 min before treatment with NaHS. The randomly chosen cells were stimulated with a ramp voltage (from -120 mV to +40 mV, 850-ms duration) (A) or a square pulse of +10 mV (200-ms duration) (B, C) from a holding potential of -90 mV. HVA currents (C) were determined as the currents at 75 ms after the beginning of the test pulse (B). Data show the mean with S.E.M. from 32-36 (A) and 31-39 (C) different cells. * p < 0.05, ** p < 0.01. V, vehicle.
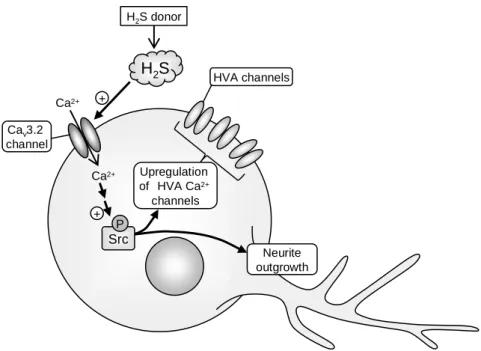
Ⅳ 考察
神経前駆 NG108-15 細胞に対して H2S ドナーである NaHS を処置することによ
り誘起される神経突起伸長や HVA-currents の増大は Src 阻害薬 PP2 により抑制 された(Figs. 4A、7C)。これより、NG108-15 細胞の NaHS 誘起神経様分化には、 Src シグナルが関与することが考えられる。NG108-15 細胞において NaHS は Src
のリン酸化を誘起したが、これは細胞内 Ca2+キレーターBAPTA/AM、T 型 Ca2+
チャネル阻害薬 mibefradil や T 型 Ca2+チャネルのうち Ca
る ascorbic acid によって抑制された(Figs. 5、6)。一方、NaHS 誘起神経突起伸 長も BAPTA/AM、mibefradil 及び ascorbic acid により抑制されることがすでに報 告されている (11)。これより、NG108-15 細胞の NaHS 処置により誘起される神 経様分化には、Cav3.2 T 型 Ca2+チャネルの機能増強を介した細胞内 Ca2+濃度上昇 に続く Src リン酸化が関与していることが示唆された(Fig. 8)。 Ca2+ Ca2+ H2S Cav3.2 channel + Upregulation of HVA Ca2+ channels HVA channels Neurite outgrowth H2S donor Src P +
Fig. 8. Possible mechanisms of the NaHS/H2S-induced neuronal differentiation in NG108-15 cells. The
NaHS-derived H2S enhances the activity of Cav3.2 T-type Ca2+ channels and causes Ca2+ influx, and
subsequent activation of Src causes neuronal differentiation characterized by neurite outgrowth and upregulation of HVA Ca2+ channels in NG108-15 cells.
Ca2+イオノフォアである ionomycin 処置により誘起される神経突起伸長も同様 に Src 阻害薬 PP2 により抑制されたことからも、この仮説を支持する知見であ る(Fig. 4B)。一方、db-cAMP 誘起神経突起伸長も、PP2 により部分的に阻害さ れた(Fig. 4C)。しかしながら、db-cAMP 処置 NG108-15 細胞では、リン酸化 Src は検出されたが、vehicle 処置細胞と比べ有意なリン酸化 Src の増加は見られな かった(Fig. 5C)。これより、db-cAMP 刺激によって Src の活性化レベルは増強 されないが、ある程度 Src が活性化された状態でなければ分化は起こらないこと
が示唆された。 NG108-15 細胞において、単回処置で T-currents を増強する低濃度 NaHS(1.5 mM)では、有意な Src リン酸化は見られなかった(Fig. 5B)。一方、単回処置 で有意な神経突起伸長効果や Src リン酸化を示した NaHS 濃度(13.5 mM)は非 常に高濃度であり、生理的及び病態生理学的には到達しない濃度であると考え られる。このような高濃度を要した原因として、H2S が気体であるため NaHS を 培養液中に添加すると生成した H2S が急速に空気中へ拡散・消失してしまうこ とが考えられる。実際、NaHS を 37˚C の培養液中へ添加した場合、H2S の半減 期は 6.2 分であることが示されている (24)。一方、低濃度 NaHS(1.5 mM)を 3 ~5 回繰り返し添加することによって、vehicle 処置群と比べて有意な神経突起伸 長や Src リン酸化が誘起された(Figs. 2A、5B)。これより NaHS による神経突起
伸長や Src リン酸化には、持続的な Cav3.2 T 型 Ca2+チャネルの活性化による細 胞内 Ca2+流入が必要であることが考えられる。 PC12 細胞の脱分極により引き起こされる神経突起伸長においても Src の活性 化が関与し、この Src 活性化の上流シグナルには、脱分極による細胞内 Ca2+濃度 上昇後の CaMKII の活性化及び下流シグナルには MEK/ERK 系が寄与することが 明らかとなっている (27)。一方、NaHS 誘起神経突起伸長は、CaMKII 阻害薬 KN-93 や PKA 阻害薬 KT5720 により影響を受けなかった。さらに、MEK 阻害剤 U0126、PI3K 阻害薬 LY294002 及び非選択的 tyrosine kinase 阻害薬 genistein は、 単独添加により神経突起伸長を促進した。これより、今回の検討によって、NaHS 誘起神経突起伸長において CaMKII 及び PKA は関与しないこと及び MEK/ERK、 PI3K 及び tyrosine kinase の関与については明らかにできなかった為、NG108-15
細胞の細胞内 Ca2+濃度上昇後の Src 以外の細胞内シグナルは未解明のままであ
を介して神経突起伸長を促進することが報告されている (28, 29)。そのため、 NaHS 誘起神経突起伸長に対する U0126 以外の MEK/ERK 阻害剤による影響や ERK のリン酸化の状態を確認することで、NaHS 誘起神経突起伸長における MEK/ERK 系の関与を明らかにできる可能性がある。 H2S は Cav3.2 T 型 Ca2+チャネル以外に、HVA Ca2+チャネル、KATPチャネルあ るいは TRPA1 チャネル等にも作用する (24-26)が、NaHS 誘起神経突起伸長には T 型 Ca2+チャネル以外のイオンチャネルの関与は認められなかった。また、NaHS 誘起 T-currents 増大や神経突起伸長は T 型 Ca2+チャネル阻害薬 mibefradil や T 型 Ca2+チャネルのうち Ca v3.2 を選択的に阻害する ascorbic acid によって抑制される ことから、NG108-15 細胞において H2S は Cav3.2 T 型 Ca2+チャネルを活性化する ことで神経突起伸長を誘起することが強く示唆される。 神経系における H2S による Cav3.2 チャネルの機能調節は、体性痛や内臓痛な どの様々な痛覚伝達やヒスタミン非依存性の痒み伝達に関与することが明らか となっており、注目を集めている (3, 7-9, 21, 30)。今回の検討では、NG108-15 細胞において H2S は、Cav3.2 T 型 Ca2+チャネルからの Ca2+流入を促進し、Src 型 チロシンキナーゼの活性化を介して神経様分化を誘起することを明らかとした。 この知見は、H2S/Cav3.2 系が疼痛の誘発だけではなく、神経分化や神経損傷後の 軸索再生に寄与する可能性を示唆するものであり、H2S の神経系における役割を 理解する上で新たな知見となった。
第 2 章 神経内分泌様分化ヒト前立腺癌 LNCaP 細胞
における H
2S/Ca
v3.2 チャネルによる分泌反応制御
Ⅰ 緒言
前立腺癌は男性ホルモン(アンドロゲン)に依存して増殖する性質を持つ。 そのため、アンドロゲン依存的な前立腺癌には、薬物療法もしくは外科的処置 によって体内のアンドロゲン量を減少させる、あるいはアンドロゲン受容体を 遮断するホルモン療法が非常に有効とされている。しかし、治療を一定期間継 続すると、多くの患者において、癌細胞はホルモン療法に対して抵抗性を示す ようになり、癌が再燃する。この前立腺癌のホルモン療法抵抗性の獲得に関与 するメカニズムを解明し、これを克服する新たな治療法を確立することが急務 である。現在までの研究により、前立腺癌のホルモン療法抵抗性獲得には、① 前立腺癌細胞自身によるアンドロゲン様分子の産生、②アンドロゲン受容体 (androgen receptor、AR)変異の結果認められる他のステロイドホルモンや AR 拮抗薬に対する増殖反応、③AR の異常な発現増加、④恒常的活性化型 AR の出 現、⑤AR 以外の増殖シグナルの獲得、などが関与する可能性が報告されている (31)。 ホルモン療法抵抗性獲得の要因の 1 つである⑤AR 以外の増殖シグナルの獲得 には、前立腺癌細胞の「神経内分泌様分化」が関与する可能性が報告されてい る (32)。ホルモン療法を行っている患者において、神経内分泌様分化前立腺癌 細胞の出現と癌増殖の程度が相関すること、また長期的なホルモン療法を行う ことで前立腺癌細胞の神経内分泌様分化が亢進することが報告され、ホルモン 療法抵抗性と神経内分泌様分化の関係が注目されるようになった (33, 34)。興味深いことに、神経内分泌様分化前立腺癌細胞自身は増殖活性が低く、AR を欠損 しているため、従来のホルモン療法ではこの細胞の機能を抑制することはでき ない。一方、神経内分泌様分化前立腺癌細胞は様々な細胞増殖因子を産生、分 泌することで周囲の癌細胞の増殖を促し、その結果、腫瘍全体がホルモン療法 に抵抗性を示すと考えられている (35-37)。 実 験 的 に は 、 培 養 し た 前 立 腺 癌 細 胞 で は 、 培 地 中 ア ン ド ロ ゲ ン 除 去 、 interleukin-6 (IL-6)刺激あるいは細胞内 cAMP 量を増加させる薬物などによっ て神経内分泌様分化が誘導される (38)。興味深いことに、細胞内 cAMP 量を増 加させることで神経内分泌様細胞へ分化を誘導したアンドロゲン依存的ヒト前
立腺癌 LNCaP 細胞は、Cav3.2 T 型 Ca2+チャネルを高発現し、Cav3.2 を介して細
胞内へ流入した Ca2+ に依存して細胞増殖因子等の分泌がおこることが報告さ れている (39, 40)。一方、硫化水素(H2S)は、Cav3.2 を含むいくつかのイオン チャネルの機能を変化させることで、痛みの情報伝達や神経分化を促進するほ か、分泌反応の調節にも関与することが報告されている (25, 41-45)。さらに、 ホ ル モ ン 療 法 抵 抗 性 を 示 す 前 立 腺 癌 細 胞 は H2S 合 成 酵 素 で あ る cystathionine--lyase(CSE)あるいは cystathionine--synthase(CBS)を発現して いることが報告されており (46)、H2S が前立腺癌において何らかの病態生理学 的役割を果たしている可能性がある。そこで、第 2 章では db-cAMP 誘起神経内 分泌様分化 LNCaP 細胞の分泌機能調節における内因性 H2S/Cav3.2 T 型 Ca2+チャ ネルの役割に焦点をあて検討した。また、Cav3.2 のタンパク発現量は転写促進
因子である early growth response 1(Egr-1)により促進的に、転写抑制因子であ る repressor element-1 silencing transcription factor(REST)により抑制的に制御さ れることが報告されていることより (17)、db-cAMP 処置によって神経内分泌様
タンパク発現増加への関与を検討した。
Ⅱ 方法
1)LNCaP 細胞の培養ヒト前立腺癌由来 LNCaP(lymph node carcinoma of prostate)細胞(独立行政法 人理化学研究所、茨木、日本)を 10% FBS、50 unit/mL penicillin、 50 µg/mL streptomycin を含む Roswell Park Memorial Institute-1640 (RPMI-1640)培地(和
光純薬工業)中で、tissue culture dish(100 × 20 mm)を用いて、37℃、5% CO2
の条件下で培養した。
2)Whole-cell patch-clamp 法による Ba2+電流の測定
LNCaP 細胞を poly-L-ornithine でコーティングした tissue culture dish(35 × 10
mm)に 2.0 × 104 cells/dish で播種し、上述の RPMI-1640 培地にて 3 時間培養し
た後、分化誘導剤(db-cAMP、IBMX、isoprenaline、IL-6、bicalutamide、flutamide) を添加し 3-5 日間培養した細胞を実験に用いた。細胞外溶液及び細胞内溶液は、
第 1 章で記した同様の方法で調製した。Ba2+ currents は、第 1 章と同様の条件及
び増幅器を用いて測定した。保持電位は-80 mV にした。データは、デジタイザ ー (Digidata 1322A もしくは 1440A) を経由してデジタル化したものをコンピ ュータに入力させ、pClamp8 もしくは 10.2 software(Axon Instruments)を用いて 解析した。それぞれの阻害薬は刺激開始の 30 分前に添加した。なお T-currents
は、保持電位 -80 mV から-20 mV のテストパルス刺激を与えた時の Ba2+ currents
を測定し、この電流のピーク電流から 150 ms 後の電流を差し引いた値として算 出した。
3)Reverse transcription-polymerase chain reaction(RT-PCR)法
Cav3.2 、neurotensin、parathyroid hormone-related peptide(PTHrP)の mRNA 検
出では LNCaP 細胞を poly-L-ornithine でコーティングした 6-well plate に 3.0 × 105
cells/well で播種した後、db-cAMP と IBMX を添加し 3 日間培養した細胞を実験 に用いた。Prostate specific antigen(PSA)の mRNA 検出では LNCaP 細胞を同様
の 6-well plate に 3.0 × 105 cells/dish で播種し、FBS 中に含まれるステロイドホル
モンの影響を除く為 10%の charcol/dextran-stripped fetal bovine serum(CSS) (Thermo Fisher Scientific、 横浜、 日本)を含む RPMI-1640 培地にて 3 時間培 養し、db-cAMP と IBMX を添加した (39)。その 2 日後に db-cAMP と IBMX を 加えた新しい 10% CSS を含む RPMI-1640 培地で培地交換を行い、さらに 3 日間 培養した細胞を用いた。培養開始から 5 日目の細胞の培地を 10% CSS を含む新 しい培地に交換し、1 時間後に dihydrotestosterone(DHT)で刺激を行い、その 6 時間後の細胞を実験に用いた。
上記の細胞を回収し、total RNA を抽出した。RNA LA PCR kit(AMV)ver.1.1 (タカラバイオ株式会社、滋賀、日本)を用いて、1 µg の total RNA を 42℃で 50 分間及び 70℃で 15 分間、逆転写反応させて cDNA を得た後、PCR 法により
増幅させた。プライマーは、Cav3.2(α1H)(PCR product size: 193 bp);5’-TTG GGT
TCC GTC GGT TCT-3’(forward)、5’-ATG CCC GTA GCC ATC TTC A-3’(reverse)、
neurotensin(PCR product size: 131 bp);5‘-GCA TGC TAC TCC TGG CTT TC-3’
(forward)、5’-CCA AGA GGG AAC ATG TGC TT-3’(reverse)、PTHrP(PCR product
size: 93 bp);5‘-GTC TCA GCC GCC GCC TCA A-3’(forward)、5’-GGA AGA ATC
GTC GCC GTA AA-3’(reverse)PSA(PCR product size: 194bp);5’-GGT GAC CAA
GTT CAT GCT GTG-3’(forward)、5’-GTG TCC TTG ATC CAC TTC CG-3’(reverse)、
GAA GGT GAA GGT CGG AGT C -3’(forward)、5’- GAA GAT GGT GAT GGG ATT TC -3’(reverse)を用いた。PCR 反応は、変性 94℃ 30 秒、アニーリング 55℃ 30 秒、伸長 72℃ 1 分を 1 サイクルとし、GAPDH は 25 サイクル、その他は 35 サ イクル反応させた。反応液中の PCR 産物の確認は、2%アガロースゲルで電気泳 動後、ゲルを臭化エチジウムで染色し、紫外線を照射して行った。
Real-time PCR 法の場合は、Cav3.2、neurotensin 及び PTHrP の最終 cDNA 量が
100 ng、PSA と GAPDH の最終 cDNA 量が 25 ng となるように diethylpyrocarbonate
(DEPC)水で希釈した。PCR-96-PE plate(BM ビーエム株式会社、東京、日本) に Power SYBR Green PCR Master Mix(Applied Biosystems、東京、日本)と最終 濃度 0.2 µM のプライマー、それぞれのサンプルと DEPC 水を入れた。その後、 ABI PRISM 7000 Sequence Detector (Applied Biosystems)もしくは Light Cycler 480 (Roche-Biochem、東京、日本)を用い Step1 で 50℃ 2 分を 1 サイクル、Step2 で 95℃ 10 分を 1 サイクル、Step3 で 95℃ 15 秒、60℃ 1 分を 40-45 サイクルの PCR 反応を行った時の蛍光を測定した。
4)形態学的検討
LNCaP 細胞を poly-L-ornithine でコーティングした tissue culture dish(35 × 10
mm)に 1.0-2.0 × 104 cells/dish で播種した後、分化誘導剤(db-cAMP、IBMX、
isoprenaline、IL-6、bicalutamide、flutamide)を添加し 2 日間培養した細胞を実験 に用いた。光学顕微鏡下で細胞を観察し、50 µm より長い神経様突起をもつ細胞 数を計測し、視野内の全細胞数に対する割合を求めた。なお、阻害剤は刺激開 始 30 分前もしくは db-cAMP 刺激 2 日後に添加した。
5)MTT assay 未分化 LNCaP 細胞の検討の場合は poly-L-ornithine でコーティングした 48 well-plate に 1.0 × 104 cells/well となるように播種し、24 時間培養した後、培地の 全量が 200 µL になるように培地を回収した。神経内分泌様分化 LNCaP 細胞の 検討の場合は細胞を播種した後、db-cAMP(1 mM)と IBMX(100 µM)を加え 2 日間培養した。その後、同濃度の db-cAMP と IBMX を含む新しい培地で培地 交換を行い、さらに 3 日間培養した後、培地の全量が 200 µL となるように培地 を回収した。その未分化あるいは神経内分泌様分化 LNCaP 細胞に対し阻害薬を 添加して、37℃、48 時間インキュベート後、MTT Cell Proliferation Kit(Cayman Chemical Company、MI、USA) を用いて細胞数を測定した。MTT 試薬(5 mg/mL) を各 well に添加し 37℃、4 時間インキュベートした後、Crystal Dissolving Solution で可溶化した。その後、各サンプルを回収し、遠心分離(300 G、5 min)を行っ た後、595 nm (対照波長 750 nm)で吸光度を測定した。
6)Western blot 法
Cav3.2、Egr-1、REST、CREB、p-CREB、CSE 及び CBS のタンパク検出では
LNCaP 細胞を poly-L-ornithine でコーティングした 6 well-plate に 3.0 × 105
cells/well で播種した後、db-cAMP と IBMX を添加した。これらの細胞を SDS
buffer で溶解した後に回収した。このサンプルを用いて Cav3.2、Egr-1、REST、
CREB、p-CREB、CSE、CBS 及び GAPDH の検出を行った。Lowry 法もしくは Bradford 法によりサンプルのタンパク量を定量した。CREB、p-CREB、CBS 及 び GAPDH は 10 µg、CSE は 50 µg のタンパク質量のサンプルを 12.5%
SDS-polyacrylamide gel を用いて、Cav3.2、Egr-1 及び REST は 30 もしくは 40 µg
より分離させた。分離させたタンパクの転写及びブロッキングは第 1 章と同様 の方法で実施した。以下の一次抗体を 5% nonfat dry milk-TBS/T でそれぞれの倍
率に希釈して 4℃で一晩反応させた。一次抗体は、抗 Cav3.2 抗体(1:500)(Santa
Cruz Biotechnology Inc.、Santa Cruz、CA、USA)、抗 Egr-1 抗体(1:1000)(Santa Cruz
Biotechnology Inc.)、抗 REST 抗体(1:1000)(Merck Millipore、東京、日本)、抗
CREB 抗体(1:1000)(Cell Signaling Technology)、抗 CREB(phospho-Ser 133)
(p-CREB)抗体(1:1000)(nano Tools、Teningen、Germany)、抗 CSE 抗体(1:1000)
(Sigma Genosys、St. Louis、MO、USA)、抗 CBS 抗体(1:3000)(Abnova Corporation、
Taipei、Taiwan)、抗 GAPDH 抗体(1:5000)(Santa Cruz Biotechnology Inc.)を用
いた。二次抗体として Cav3.2、Egr-1、REST、CREB、CSE 及び GAPDH の検出
には HRP-linked anti-rabbit IgG 抗体(1:1000)(Merck Millipore)、CBS、p-CREB の検出には HRP-linked anti-mouse IgG 抗体(1:1000)(Cell Signaling Technology) を用い 5% nonfat dry milk を含む TBS/T buffer でそれぞれの倍率で希釈し、室温 で 90 分反応させ、ECL(Amersham Bioscuences)もしくは Chemi-Lumi One Super (Nacalai Tesque)で 1 分間反応させて発光させることにより、各タンパク質の バンドを検出した。検出されたバンドは、Win Roof または ImageJ を用いて数値 化した。
7)siRNA による RNA 干渉法
LNCaP 細胞は poly-L-ornithine でコーティングした 6 well-plate に 3.0 × 105
cells/well となるように播種し、penicillin 及び streptomycin を含まない、10% FBS を含む RPMI-1640 培地で培養した。一晩培養した後、DharmaFECT3 Transfection Reagent(Thermo Fisher Scientific K.K.、東京、日本)を含む 40 pmol の siGENOME SMARTpool Human EGR1 ( Thermo Fisher Scientific K.K ) 及 び siGENOME
Non-Targeting siRNA#2(Thermo Fisher Scientific K.K)を添加した。3 日間培養後、 同濃度の siRNA 存在下で db-cAMP(1 mM)及び IBMX(100 µM)を添加し、 さらに 24 時間培養した細胞を回収した。Western blot 法を用いて本サンプルの
Cav3.2 と Egr-1 のタンパク発現量を検出した。
8)Prostatic acid phosphatase(PAP)assay
LNCaP 細胞を poly-L-ornithine でコーティングした 6 well-plate に 3.0 × 105
cells/well で 10% FBS を含む上記の培地に播種した後、分化誘導剤(db-cAMP、 IBMX、IL-6)を添加した。2 日間培養した後、分化誘導剤を含む FBS-free の新 しい培地に交換し、さらに 3 日間培養したときの上清中に含まれる PAP 量、及 びこの培養後に 1% Triton-X 処置をして得られた全細胞中の PAP 量(全細胞中の PAP 量)を tartrate-resistant acid phosphatase (TRACP)& alkaline phosphatase (ALP)assay kit(タカラバイオ、滋賀、日本)を用いて測定した。各サンプル を回収し、遠心分離(300 G、10 min)を行った後、上清 25 µL ずつを 96-well plate に移し、そこに pNPP substrate を 25 µL ずつ加えた後、37℃で 45 分間反応させ た。その後、各 well に 25 µL ずつ 0.5 N NaOH を加えて反応を停止させた後、405 nm で吸光度を測定した。なお、阻害剤は培地交換時に添加した。
9)試薬
実験に使用した試薬は以下のとおりである。
NaHS はキシダ化学から購入した。Na2S、db-cAMP、3-isobutyl-1-methylxanthine
(IBMX)、isoprenaline、KT5720、dihydrotestosterone(DHT)、bicalutamide、flutamide、
mibefradil、DL-propargylglycine (PPG)、β-cyano-L-alanine(BCA)、aminooxyacetic
protein(AKAP)St-Ht31 inhibitor peptide(AKAPI)は Promega(Madison、WI、 USA)から購入した。IL-6 は和光純薬工業から購入した。IL-6 は滅菌 phosphate
buffered saline(PBS)で溶解して使用した。Flutamide はエタノール(EtOH)(和
光純薬工業)で溶解して使用した IBMX、KT5720、bicalutamide、NNC は DMSO (Sigma)で溶解して使用した。その他の試薬は滅菌蒸留水で溶解して使用した。 10)統計処理 得られた実験値は、平均値±標準誤差で示した。有意差の検定には t 検定及び チューキー法を用いP<0.05 で有意差ありとした。
Ⅲ 結果
1)LNCaP 細胞における db-cAMP、IBMX、isoprenaline 及び IL-6 による神経様 突起伸長効果
前 立 腺 癌 細 胞 の 神 経 内 分 泌 様 分 化 の 特 徴 と し て 、 神 経 様 突 起 伸 長 、 neuron-specific enolase 等の神経マーカーの発現増加や細胞増殖能の低下等が報 告されている (38)。そこで、まず神経様突起伸長を指標として、様々な分化誘 導剤により LNCaP 細胞の神経内分泌様分化が誘起されるか否かを確認した。未 分化 LNCaP 細胞に、細胞内 cAMP アナログである db-cAMP、phosphodiesterase 阻 害薬である IBMX、アドレナリン受容体アゴニストである isoprenaline、炎症性 サイトカインである IL-6 を添加して 48 時間後に神経様突起伸長効果を評価した。 その結果、LNCaP 細胞において、db-cAMP と IBMX 併用添加することにより
db-cAMP の濃度依存的な神経様突起伸長の亢進がみられた(Fig. 9A、B)。この
した方がより強力であった(Fig. 9B、C)。また、db-cAMP 添加時と比べて程度 は弱いものの、isoprenaline+IBMX や IL-6 による刺激によっても、同様の神経 様突起伸長が誘起された(Fig. 9D、E)。一方、db-cAMP 誘起神経様突起伸長効 果 は 、 PKA 阻 害 薬 KT5720 あ る い は PKA の 足 場 タ ン パ ク で あ る A-kinase-anchoring protein (AKAP)と PKA の結合を阻害する AKAP St-Ht31
inhibitor peptide(AKAPI)の前処置により、部分的であるが有意に抑制された(Fig.
9F、G)。 Vehicle db-cAMP 1 mM plus IBMX (B) P ro p o rt io n o f c e lls w it h n e u ri te s (% ) V + V IBMX 0 10 20 30 40 50 db-cAMP (mM) 0 .1 0 .3 1 V * * * * * (C) 0 10 20 30 40 50 db-cAMP (mM) 0 .1 0 .3 1 V * * * * (F) 0 10 20 30 40 50 V K T 5 7 2 0 V V K T 5 7 2 0 db-cAMP 1 mM * * * * (G) 0 10 20 30 40 50 V AKAPI V V AKAPI db-cAMP 1 mM * * * * 50 100 V IL-6 (ng/ml) 0 4 8 12 16 (E) ** 0 .0 1 Isoprenaline (µM) V 0 .1 1 0 4 8 12 16 V + V IBMX (D) * * ** * * * P ro p o rt io n o f c e lls w it h n e u ri te s (% ) (A)
Fig. 9. Effects of dibutyryl cyclic AMP (db-cAMP), IBMX, isoprenaline, and IL-6 on the neurite outgrowth in LNCaP cells. (A) Representative photographs of LNCaP cells treated with 1 mM db-cAMP plus 100 µM IBMX or vehicle (water plus DMSO) for 48 h. Scale bars indicate 50 µm. (B-E) Neurite outgrowth caused by 48-h stimulation with db-cAMP plus 100 µM IBMX (B), db-cAMP (C), isoprenaline plus IBMX (D) or interleukin-6 (IL-6) (E). (F, G) Effects of KT5720, a protein kinase A inhibitor, and AKAPI, an AKAP inhibitor, on the db-cAMP-induced neurite outgrowth in LNCaP cells. Cells were treated with 10 µM KT5720 (F) or 10 µM AKAPI (G) 30 min before stimulation with db-cAMP for 48 h. Data show the mean with S.E.M. for 4-7 (B, C, E-G) and 9-12 (D) different experiments. * p < 0.05, ** p < 0.01. V, vehicle.
2)db-cAMP 誘起神経内分泌様分化 LNCaP 細胞におけるアンドロゲン非依存的 性質の獲得 未分化の前立腺癌細胞は、男性ホルモンであるアンドロゲン依存的に増殖す る性質をもつため、ホルモン療法や抗アンドロゲン薬処置により細胞増殖が抑 えられる。一方、神経内分泌様分化前立腺癌細胞は、細胞増殖能が低いことに 加え、アンドロゲン受容体の発現が低下しておりアンドロゲンシグナルが減少 している (38)。神経内分泌様分化前立腺癌細胞はこのようなアンドロゲン非依 存的性質を獲得することで、ホルモン療法や抗アンドロゲン薬に抵抗性を示す。 そこで、アンドロゲンシグナルの指標である prostate specific antigen(PSA)の mRNA 発現量を real-time PCR 法により測定することで、未分化及び db-cAMP 誘 起神経内分泌様分化 LNCaP 細胞におけるアンドロゲンシグナルの変化を確認し た。さらに、未分化及び db-cAMP 誘起神経内分泌様分化 LNCaP 細胞の細胞増 殖に対する抗アンドロゲン薬 bicalutamide の抗腫瘍活性の変化について MTT 法 を用いて検討した。db-cAMP+IBMX で 5 日間処置することで神経内分泌様に分 化した LNCaP 細胞では、未分化細胞と比べて、PSA mRNA 発現量が有意に増加 していた(Fig. 10A)。一方、アンドロゲンの一種である dihydrotestosterone(DHT) 処置 6 時間後にみられるアンドロゲン依存的 PSA mRNA 発現量の増加は、未分 化細胞に比べ db-cAMP 誘起神経内分泌様分化 LNCaP 細胞において有意に低下 していた(Fig. 10B)。さらに、bicalutamide を 2 日間処置することにより、未分 化 LNCaP 細胞では濃度依存的な細胞増殖の抑制がみられたが、db-cAMP 誘起神 経内分泌様分化 LNCaP 細胞では有意な増殖抑制効果は見られなかった(Fig. 11)。
(A) P S A /G A P DH m R NA db -cA M P + I B M X V 0 1 2 3 4 * (B) P S A /G A P DH m R NA (f o ld in cr e a s e o f b a s a l le ve ls ) 0 4 6 10 DHT 1 nM 8 2 db -cA M P + I B M X V DHT 10 nM db -cA M P + I B M X V DHT 100 nM db -cA M P + I B M X V * * *
Fig. 10. Expression levels of prostate specific antigen (PSA) mRNA in undifferentiated- and db-cAMP-induced neuroendocrine-like differentiated LNCaP cells, and effects of dihydrotestosteron (DHT), one of androgens, on the PSA mRNA levels. (A) PSA mRNA levels were analyzed in LNCaP cells treated with db-cAMP at 1 mM plus IBMX at 100 µM (neuroendocrine-like differentiated cells) or vehicle (V: water plus DMSO) (undifferentiated cells) for 5 days. (B) Dihydrotestosteron (DHT)-evoked increase in PSA mRNA levels was analyzed in the neuroendocrine-like differentiated (db-cAMP plus IBMX) and undifferentiated (V) LNCaP cells 6 h after application of DHT at 1 to 100 nM. Data show the mean with S.E.M. from 18 (A) and 6 (B) different experiments. * p < 0.05. V, vehicle. (A) (B) V 1 5 10 50 Bicalutamide (µM) M T T a ct iv it y ( % c o n tr o l) 0 20 40 60 80 100 120 V 1 5 10 50 Bicalutamide (µM) 0 20 40 60 80 100 120 db-cAMP + IBMX V M T T a ct iv it y ( % c o n tr o l) ** * ** *
Fig. 11. Effects of an androgen receptor antagonist, bicalutamide, on proliferation in the undifferentiated LNCaP cells or in neuroendocrine-like differentiated LNCaP cells after stimulation with db-cAMP plus IBMX for 5 days. (A) Undifferentiated LNCaP cells were cultured for 24 h, and then stimulated with bicalutamide or vehicle (V: DMSO) for 2 days. (B) The neuroendocrine-like LNCaP cells, which were differentiated by treatment with db-cAMP at 1 mM plus IBMX at 100 µM for 5 days, were stimulated with bicalutamide or vehicle (V, DMSO) for 2 days. Data show the mean with S.E.M. from 6 (A) and 9 (B) different experiments. * p < 0.05, ** p < 0.01. V, vehicle.
3)db-cAMP 誘起神経内分泌様分化 LNCaP 細胞における細胞増殖因子の発現量 増加
神経内分泌様分化前立腺癌細胞は、neurotesin や parathyroid hormone-related
peptide(PTHrP)等の細胞増殖因子を発現・分泌することが報告されている (47)。
ると考えられている。そこで、db-cAMP 誘起神経内分泌様分化 LNCaP 細胞にお ける neurotesin や PTHrP の mRNA 発現量を real-time PCR 法により測定した。そ の結果、db-cAMP+IBMX で 3 日間処置することで神経内分泌様に分化した LNCaP 細胞では、未分化細胞に比べ、neurotensin 及び PTHrP の mRNA 発現量の 有意な増加が確認された(Fig. 12)。 (B) N e u ro te n sin /GA P D H m R N A 0 0.004 0.008 0.012 0.016 0.020 db -cA M P + I B M X V P T H rP /GA P D H m R N A 0 0.2 0.4 0.6 0.8 1.0 db -cA M P + I B M X V (A) * * * * * *
Fig. 12. Upregulation of neurotensin and parathyroid hormone-related peptide (PTHrP) at mRNA levels in neuroendocrine-like differentiated LNCaP cells after stimulation with db-cAMP + IBMX for 3 days. Quantitative reverse transcription-polymerase chain reaction (qRT-PCR) analysis was used to determine the neurotensin or parathyroid hormone-related peptide (PTHrP) and glyceraldehyde-3-phosphate dehydrogenase (GAPDH) mRNA levels. Data show the mean with S.E.M. for 12 (A) or 7 (B) different experiments. *** p < 0.001. V, vehicle.
4)LNCaP 細胞における db-cAMP 誘起神経内分泌様分化に伴った T-currents の 増大及びその増大に対する PKA 阻害薬の抑制効果
LNCaP 細胞では db-cAMP と IBMX の処置による神経内分泌様分化に伴い T
型 Ca2+チャネル電流(T-currents)及び Ca
v3.2 T 型 Ca2+チャネルの mRNA 発現増
加が起こることが報告されている (39)。そこで、まず db-cAMP 誘起神経内分泌 様分化 LNCaP 細胞における T-currents について whole-cell patch-clamp 法を用い て確認したのち、その db-cAMP 誘起 T-currents の増大に対する PKA 阻害薬の影 響について検討した。その結果、db-cAMP+IBMX を 4-5 日間処置することで神 経内分泌様に分化した LNCaP 細胞では、vehicle 処置細胞に比べて膜電位-20~-10
mV あたりをピークとする Ca2+チャネルを介した Ba2+ currents の有意な増大が確
と考えられる電位である-20 mV のテストパルス刺激を行った時に得られる電流
から、HVA Ca2+チャネル電流を除くため刺激開始 150 ms 後の電流を差し引いた
電流を T 型 Ca2+チャネル電流(T-currents)として算出した(Fig. 13B)。その結
果、vehicle 処置に比べ db-cAMP+IBMX 誘起神経内分泌様分化 LNCaP 細胞では、
明らかに T-currents が増大していた(Fig. 13C)。一方、db-cAMP+IBMX 誘起 T-currents の増大は、PKA 阻害薬 KT5720 及び AKAPI の前処置により部分的に 抑制された(Fig. 14)。 (A) (B) (C) 50 pA -20 mV -80 mV 200 ms 150 ms T-current 40 -2.0 0 (mV) -80 -40 * * * * * * * * * * * * * * * * * * * (pA/pF) 1.0 , Vehicle , db-cAMP + IBMX Test pulse: -20 mV -1.2 -0.9 -0.6 -0.3 0 T -c u rr e n ts ( pA /p F ) db-cAMP + IBMX V *
Fig. 13. Upregulation of T-type Ca2+ channel-dependent currents (T-currents) in neuroendocrine-like
differentiated LNCaP cells. (A) Current-voltage relationship for calcium channel-dependent currents in LNCaP cells treated with db-cAMP at 1 mM and IBMX at 100 µM for 4-5 days. (B) T-type Ca2+ channel-dependent currents (T-currents) were determined as the difference between the peak current and the current 150 ms after beginning of the test pulse at -20 mV from a holding potential of -80 mV. (C) T-currents in LNCaP cells treated with db-cAMP plus IBMX for 4-5 days. Data show the mean ± S.E.M. for 22-26 (A, C) * p < 0.05, ** p < 0.01, *** p < 0.001. V, vehicle.
(A) (B) T -c u rr e n ts ( pA /p F ) -2.0 -1.5 -1.0 -2.5 -0.5 0 V K T 5 7 2 0 V V K T 5 7 2 0 db-cAMP + IBMX -2.5 -2.0 -1.5 -1.0 -0.5 0 T -c u rr e n ts ( pA /p F ) V AKAPI V V AKAPI db-cAMP + IBMX * * * * * * *
Fig. 14. Effects of protein kinase A (PKA) inhibitors on the up-regulation of T-currents induced by db-cAMP plus IBMX in LNCaP cells. Whole-cell patch-clamp technique was performed 5 days after the addition of db-cAMP at 1 mM plus IBMX at 100 µM. KT5720 at 10 µM (A) or AKAPI at 10 µM (B) was applied 30 min before addition of db-cAMP plus IBMX. Data show the mean ± S.E.M. for 16-22 (A) and 30-32 (B) different experiments. * p < 0.05, ** p < 0.01. V, vehicle.