香川大学農学部紀要 第57号 1∼94,1992
イチゴの花器および果実の発育に関する研究
−‘愛ベリー’の奇形果発生を中心として一
昔 田 裕
Studies on Flower and Fruit Developmentin StrawberTy,
With SpecialRefference to Fruit Malformationin‘Ai−Berry
Yuichi YosHIDA 目 次 第Ⅰ章 供試品種および調査形質の概要 3 3 3 4 4 4 4 4 5 7 7 9 ︿U l 第1節 供試品種の特性 1. 愛ベリー 2 宝交早生 3 贋紅 4“ とよのか 5. 女峰 6.. 盛岡16号 第2節 花芽の分化発育経過と発育段階の判定 第3節 イチゴの花序,花器および果実の呼称 1い 花序 2り 花器および果実 第4節 花器の話形質の調査方法 第Ⅰ章 ‘愛べリー’の奇形果発生要因 2 2 2 3 5 5 7 2 1 1 1 1 1 1 1 2 第1節 奇形果の発生様相 1 材料および方法 2一 結果および考察 第2節 窒素施肥盈と苗質が奇形果発生と収虚構成に及ぼす影響 1. 材料および方法 2‖ 結果および考察 第3節 雌ずいの発育様相と奇形果発生に及ぼす人工受粉の効果
1. 材料および方法 2‖ 結果および考察 3 3 5 5 7 1 2 2 2 2 2 2 3 3 第4節 雌ずいの形態と受精能力の開花後の変化 1. 材料および方法 2 結果および考察 第5節 総合考察 第6節■摘要 第Ⅲ章 雌ずいの分化発育速度と奇形果発生の品種間差異 第1節 開花時における雌ずいの発育様相並びに笥形果発生の品種間差異 1 材料および方法 2小 結果および考察 第2節 雌ずいの分化時期と発育速度の変異並びに品種間差異 1. 材料および方法 2. 結果および考察 第3節 線合考察 第4節 摘要 第Ⅳ章 ‘愛ベリー’の生長並びに雌ずいの分化発育と奇形果発生に及ぼす栽培環境の影響 第1節 窒素栄養の影響 1. 材料および方法 2. 結果および考察 第2節 温度の影響 1. 材料および方法 2t 結果および考察 第3節 日長の影響 1い 材料および方法 2. 結果および考察 第4節 総合考察 第5節 摘要 第Ⅴ章 総 括 引用文献 Summary
ー1−
緒
.⊥ ロ 大果系のイチゴ‘愛ペリー’は,‘露紅’の自殖系ב宝交早生’の交雑後代から選抜育成された品種であ り,育成元である愛三種苗(株)によって1988年に.‘愛ペリー’と命名,発表された(109).しかし,増殖小普 及段階で混乱が生じ,「クイーンベリー」(愛知),「秀霹」(徳島),「ニュー・ペソ、−」(高知),「サンベリ1−・」(三 重)等,各産地や出荷組合によって様々な名称で出荷されているのが現状である. ‘愛ペリー’ は,正常に結実すれば頂果房1番果が60∼80gとなり,2,3番果でも50g程度の果実が多く得 られ,これまでのいずれの品種よりも大果を産するい さらに,糖度,香気ともに高く,食味が優れている.第2 果房以下の果房では果実がやや小さくなるものの,通常の無加温促成長期栽培において,全収穫期間の収盈は5 t/10a程度は得られている‖ イチゴ栽培においては収穫労力が,作付規模を制限する大きな要因となる.しか し,上述のとおり,本品種は完全な果星型の品種であり,収穫労力は少ないまた,成り疲れ症状が発生しにく いため連続収穫が可儲であるなど,従来の品種にない優れた特性を数多く持っている.. しかし,促成栽培㌢こおいては俗に“先つまり果”,“巾着果”と呼ばれている果実先端部の肥大不良果や, “先青(さきあお)果”,“先白(さきしろ)果”とも呼ばれている果実先端部の着色不良果が多発する, イチゴの奇形果発生は昭和38年頃から施設栽培の普及に伴って大きな課題となった.しかし,花粉四分子形成 期の低温や高温,あるいほ低日照並びに,疲菌御等による花粉の機能低下と受粉不足がその発生要因であり,雌 ずいが不受精となる結果,奇形果が形成されるこ.とが明らかにされた(1,2,51,87,88,91,104,106,124,125).そ の対策として,フトーキシソ塀による単為結果の誘起も検討され,2,4,5−TP(2,4,5−trichlorophenoxyaceticacid)の 利用が提唱されたこ.ともある(27,53,58,85,135,136).耕種的な対応とともに,昭和40年代初期に受粉の促進を 目的としてミツバチやシマハナアブ等の花粉媒介昆虫の利用が実用化され(1,2),一応の解決を見ている 昭和49年から50年には,‘宝交早生’に雌ずいあるいは雄ずいが未発達な異常花が多発し,関心を集めたし かし,これらの異常花は雄ずい分化初期の高温とこの時期のGA3処理が大きな原因であることが明らかにされ た(44,83) また,‘宝交早生,,‘ダナ・−, 等の品種では,“鶏冠状果”や“縦精巣”と呼ばれている花床の発育異常に よる奇形果の発生が関心を集めたしかし,花芽分化期から発育期における断板や移植,また仁窒素施肥の過剰 などによる窒素栄養の急激な変化によって多発することが認知されるようになった.その結果,育苗方法や施肥 方法の改善などの耕種的な対応が図られ,実際的には解決をみている(56). それに対し,いわゆる“先青果”や“先つまり果”は,従来から‘ダナ・ノ などの大果系の品種で,−・部に発 生することが指摘されていた.しかし,発生する品種が限られており,頻度も高くないことから大きな課題とは されなかった‖ しかし,‘愛ペリー’の場合,他品種と比敬して発生頻度が極めて高く,特にり収益性に深い関 わりを持つ頂花房の大型の果実に多発するため,大きな関心が寄せられるようになった このタイプの奇形果は,現在の実用品種の中では‘愛ペリー’以外には問題とされていない従来は育種の過 程において,奇形果の発生しやすい形質は,欠陥形質として選抜の対象とされることはなかった このような欠 陥形質を持ってはいるが,現在の品種の中では,‘愛ペリー’の果実肥大特性や食味は非常に優れた形質であ る.また,この課題が解決されることによって,実用品種としての選抜基準が変更され,新しい形の品種の誕生 が期待されるそこで,‘愛ベリー・, の奇形果発生要因の解明を目的として,本研究に着手した その結果,花床頂部の雌ずいが開花時に未成熟であり,このことが奇形果発生の直接的な原因であること,ま た,開花時における花床基部と頂部の雌ずいの間の発育差は,花床上での雌ずいの分化時期の違いに起因し,こ−2一 の分化時期の差は他の品種と比較して‘愛ペソ、−,が特異的に大きいことを明らかにした.さらに,花芽,特に 雌ずいの分化発育に及ぼす,窒素栄養,温度ならびに日長の影響を明らかにし,奇形果発生に対する技術的対応 の方向性を示すことができた. 辞 謝 本論文の作成にあたってほ,前京都大学教授㈱浅平 端博士に懇切な御指導と御校閲を賜ったり また,前京都 大学教授藤本幸平博士には本研究の遂行並びに取りまとめにあたって終始御指導,御助言を賜った‖ ここに記し て,衷心より感謝の意を表する. 香川大学農学部園芸学大講座教官各位濫は多大な御指導,御助言並びに御便宜を賜ったまた,信州大学助教 授大井美知男博士(前京都大学農学部附属農場)並びに京都大学農学部助手後藤丹十郎氏には実験の遂行にあた り終始御助言と御協力を賜った‖ さらに,京都大学農学部附属農場疏菜花井部技官各位並びに香川大学農学部徳 田 孝技官の多大な御理解と御援助を頂いた..また,実験の遂行にあたっては鈴田 富氏(現愛知県安城農業改 良普及所)はじめ園芸学大帝座専攻生各位に多大な御協力を頂いたけ愛三種雷株式会社にほ種苗の入手に当たっ て多大な御協力卑頂いた一・ここに記して,深く感謝の意を表する・ 最後にり㈱浅平 端先生は本年10月4日午前8時38分にお亡くなりになった先生に・は京都大学入学以来,公 私ともにお世話になり,長年にわたり御指導頂いた.特に,本論文の作成に当たっては,御療養中にもかかわら ず,懇切丁寧に御校閲頂いた.本論文は昨年夏に完成していたにもかかわらず,著者の怠慢のため印刷が遅れ, 先生に.贈呈することができなかったことは誠に漸悦に耐えない 浅平先生の御冥福をお祈りするとともに,著者 の怠慢をお詫びし,この論文を恩師浅平 端先生に捧げる 本給文は京都大学審査学位論文を印刷に付したものである.
−3一
策Ⅰ章 供試品種および調査形質の概要
本研究では,‘愛ベリー・,の対照品種として近年のイチゴ栽培の主要品種を適宜供試した また,本研究を遂 行する過程において,‘愛ペリ1−’ の奇形果発生が開花期までの花床上の雌ずいの分化発育と密接に関連するこ とが明らかになり,イチ・ゴの花芽,特に陳ずいの分化発育過程について検討する必要に迫られた 本章では,研究の結果を述べるに先だって,供試品種の特性を述べるさらに,雌ずいの分化発育経過の解明 にあたって用いた花芽発育段階の判定基準並びに本研究に.共通するイチづの各器官の呼称と雌ずい形質の調査方 法について詳述する第1節 供試品種の特性
‘愛ペソ、−’ の奇形果発生要因の解明に当たってほ,‘愛ペリー・’の対照品種として,両親系品種である‘宝 交早生’,‘霹紅’,現在の促成栽培の主要品種である‘とよのか’,‘女峰’,並びに寒冷地の露地栽培用品 種である‘盛岡16号,の5品種を適宜供試した“その他特記しない限り,供試品種ほ‘愛ペリー’である.以下 にそれぞれの品種の特性を述べる 1 愛ベリー ハウスメロ∵/前作の促成栽培用品種の育成を目的として,愛三種菌株式会社において育成された“‘濃紅’の 自殖系ב宝交早生’の交配実生から選抜され,1983年に‘愛ペリー’と命名された(109) 本品種の最大の特徴はその大束性にある‖ 最近育成されたイチゴ品種の果実は従来の品種と比較してかなり大 きいが,本品種の果実はこれらの新しい品種と比較しても著しく大きい.また,芳香に富み,甘味も強く,食味 の点でも非常に優れている 当初,名古屋地域で「愛交」という名称で試作されたが,育成者が品種登録を行なうために,‘愛ペリー’ と 命名した.しかし,試作段階から本品種を各地の特産品目とすべく,当初入手した産地において品種名とは別 に,出荷時の商品名がつけられた…その結果,普及当初は「アイベリー・」,「クイーンベリ1−・」(愛知),「秀雄」 (徳島),「ニューベリー」(高知),「サンベリ、−」(三重)等,各産地や出荷組合によって様々な名称で出荷され た“これらの事情により,育成者は品種登録を断念したため,正式な品種名はなかった.しかし,1988年に「疏 菜の新品種10」(109)誌上に,‘愛ペリー’ という品種名とともに育成経過および品種特性が発表され,この品 種名が定着しつつある. 根の発育は旺盛で吸肥力が強いい休眠覚醒のための低温要求盈は正確には知られていないが,経験的に‘凝 紅,に準じて,5℃以下で90時間程度の低温遭遇が必要と考えられている 果実の成熟には開花後50∼60日を要し,晩熱性である∩頂花房1番果は大きいものでは90g以上となり,2, 3番果でも50g以上の果実が得られることも少なくない.通常の無加温促成栽培では1月から収穫が始まり,中 休みも少なく6月上旬まセ収穫が可能であるい また,1月から5月までの稔収盈は約5t/10aときわめて多収 性を示す.果皮は‘靂紅,より明るい鮮紅色で,糖度,香りともに高い ただし,果実の表皮が軟らかいこと,うどんこ病,炭そ病,萎黄病にり病性であることの他に,本研究におい て主な検討課題とした“奇形果”の発生頻度が高いー4− 2. 宝交早生 ‘ァメリが にかわる生食加工兼用種の育成を目的として,兵庫県農業試験場宝塚分場において育成された一 ‘八雲,בタか−,の組合せから果数型の品種として選抜され,1960年命名,発表された(101)・ 休眠性は中程度,休眠覚醒のために必要な5℃以下の低温遭遇時間は,400∼500時間程度とされている電 照,GA3処理技術の開発によって,促成栽培に導入され,全国的に普及したが,近年新品種の普及にともな?て 栽培が減少している… 花房数,巣数はきわめて多いが,促成栽培でほ成り疲れを起こしやすい・・果実は比較的大きいが,最近育成さ れた品種と比較するとやや小さい.果皮色は,‘愛ペソノ よりやや淡い鮮紅色で糖度が高く,酸度ほ低い・頂 花房の1番果は,帯化しやすく∴鶏冠状果の発生頻度が高いが,それを除けば笥形果の発生は少ない平均果実 重は10∼15gに達し,粒ぞろいは良好である. 3… 麗 紅 ‘はるのか,の自殖系× 一福羽,の自殖系の取合せから,大果多収性の促成栽培用品種として千葉児戯業試験 場で選抜され,1976年命名発表された(89) 休眠は浅く,休眠覚醒のために必要な5℃以下の低温遭遇時間は50∼100時間である・また,軍勢が強く生態的 には促成栽培に適するが,花芽分化および果実の発育が遅いため,年内収盛は低い果皮色は‘愛ペソ・−’ より やや濃い鮮紅色で美しく,輸送性に富むが,食味がやや劣る.果実は大きく異形の乱れほ少ない 4. とよのか ‘ひみこ・בはるよい,の風合せから,早生・多収性で食味の優れた促成栽培用品種として野菜試験場久留 米支場で選抜され,1983年命名発表された(39) 休眠は浅く,休眠覚醒のために必要な5℃以下の低温遭遇時間は50∼100時間である・花芽分化期が早く,多収 性で促成栽培に適する.可溶性固形物含盈が高く,酸度は低いため,食味は優れ,芳香に富む果皮,果肉とも にやや硬く,輸送性に富む.果皮色はやや淡い鮮紅色であるただし,葉陰になると着色が悪く,色むらがでや すい一果形の乱れは少ないが,‘宝交早生’同様の鶏冠状果の発生がやや多い・ 5. 女 峰 〔一ダナーノ ×(‘はるのか,בダナー,)〕ב凝紅,の組合せから,早生・多収性で高品質の促成栽培用 品種として栃木県農業試験場で選抜され,1983年命名発表された(5)・ 休眠は涜く,‘贋紅,と同程度とみられるい花芽分化期が早く,早期収意が多い・ま声,連続開花性が強く, 促成栽培に適するい可溶性固形物含盈は高いが,酸度がやや高く,高温期にほ特にその債向が強い・果実はやや 小さいが,果皮色は鮮赤色で,果形はやや長い円錐型で美しい果形の乱れほ非常に少ないが,1∼2月の低温 期に開花した花は,奇形果を形成する場合がある‘ 6. 盛岡16号 (‘フェアファックス,בニッター・スバーグ,)ב千代田’の風合せから寒冷地の露地栽培用品種とし て,野菜試験場盛岡支場で選抜された(128)“ 休眠は‘ダナー,より長く,寒冷地の露地・早熟栽培に適する・大異性で,50g以上に達する場合もある−果 数は極めて少なく,果重塾の品種である.果皮色は鮮紅色で,果実が硬く,特に日持ちが優れるい耐病性が低 く,黒斑病に特異的に弱い.果形の乱れほ比較的少ない
−5− 第2節 花芽の分化発育経過と発育段階の判定 イチゴの花芽分化の判定は,江七(18)以降,ほとんどが,剣皮法によって実体顕微鏡下で行なわれている 上野(138)は,パラフィン切片法を用いて光学顕微鏡による観察を行なっているが,Durner・Poling(15) は,パデフィン切片法よりも剥皮法による実体顕微鏡下での判定のほうが信頼性は高いと述べている本研究に おいても,おもに剣皮法によって,実体顕微鏡下で判定したただし,光学的な実体顕微鏡では,解像力に限界 があり,詳細な形態観察や明瞭な写真を撮影することほ困難である.そこで,本研究においては,走査型電子顕 微鏡による観察と写共振影を併せて行なった 走査型電子顕微鏡観察用の試料作成方法ほ第1図に示した試料ほ採取後ただちに,2%または2.5%ダルク ルアルデヒドで固定し,4℃で貯蔵した..貯蔵した試料を適宜エクノい・・・・・ル系列で脱水し,臨界点乾燥を行なっ た‖後で述べるように,走査型電子顕微鏡による観察に.先だって,花器の諸形質,花芽発育段階等を実体顕微鏡 下で調査する場合には,水道水で水洗し,調査した後,脱水した‖ アルミニウム製試料台に固定し,白金一パラ ジウム(80−20%)を蒸着し,走査型電子顔傲鏡(日立,S−800型)で観察したい オスミウム酸による後固定と 臨界点乾燥時の酢酸イソアミルによる中間液処理は,予備実験において必要性が認められなかったため行なわな かった(148) 栄養生長状態のイチゴの茎頂は小さいが,その大きさ(2枚の美原基間の幅)は最新の菜原基の発育状態に よって異なる.すなわち,最新の菓原基が初生突起として分化した直後ほ比較的小さく,その後だんだんと大き くなり,新しい菓原基が形成される直前が最も大きい(第2臥左). イチゴの茎頂で,形態的な栄養生長から生殖生長への転換が始まると茎頂のドーム状組織は次第に肥大し,花 芽を形成するにいたる… しかし形態的な変化に先だって,生理的には花成誘導の過程が進行していると考えられ る.また,茎頂組織の形態的な変化は連続的であるため,花芽分化期(花芽形成開始期)の判定,すなわち,茎 頂が花芽形成を開始したか否かを茎頂の形態から判定するこ.とは非常に難しい.栽培現地においては,かなり細 かに分別されているようであるが,明らかに茎頂が肥大した段階でなければ,経験豊富な研究者あるいは技術者 であっても,花芽形成が始まったとは断言できない.本研究においては,明らかに.茎頂のド・−・ム状組織が肥厚し, Fixed withglutaraldehyde(20r25%inwater)
and stored at4℃
l
DehydratedinethanoIseriesfor60minineach
COnCentrationexceptthe fina1100%solution(24hr) 30%−50%−70%−80%−85%−90%一95%−100%−100%
王
Dried using thecriticalpointdrier(Hitachi,HCP−1) With CO2at43℃
l
Mounted on aluminum stubs and coated with Au−Pd (80−20%)using theioncoater(Eiko,SR−2)
I
Observed and photographedwith SEM(Hitachi,Sq800)
Fig2Vegetative(flower developmentalstage O)
Shoot apex of‘Ai−Berry’strawberry:Clearly uninitiated(1eft)and slightly enlarged but
Classifiedas uninitiated(right)
Barindicates200FLm
Fig1Processing methods of samples forObservation with scanning electron microscope(SEM)
ー6一
形態的に花芽形成を開始したと断言できる個体のみを花芽分化個体(花芽発育段階Ⅰ)とし,茎頂のトー・ム状離 織が肥厚していると思われるが,明瞭でない個体(第2図,右)ほ未分化個体(花芽発育段階0)と判定した
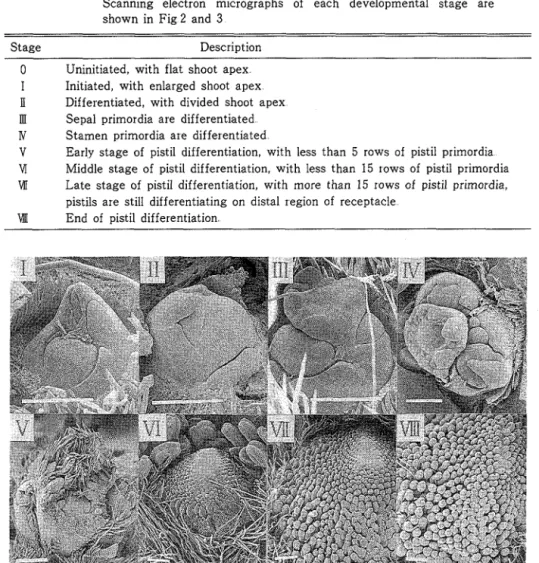
本研究において用いた花芽発育段階の分別,判定基準は第1表に,また,それらの基準に対応する花芽の形態 を第3図に示した
TablelClassification and description of 重lower developmentalstages Scanning electron micrographs of each developmental stage are ShowninFig2and3 DescI.iption g a St e
O Uninitiated,With11at shoot apex
Ⅰ Initiated,With enlarged shoot apex
I Di董ferentiated,With divided shoot apex
Ⅲ Sepalprimordiaare dif董erentiated Ⅳ Stamen primordia are differentiated
V Earlystage of pistildifferentiation,withless than5rows o王pistilprimordia
Ⅵ Middle stage of pistildi董ferentiation,Withless than15rows of pistilprimordia Ⅶ Late stage of pistildifferentiation,With more thar115rows of pistjlprimordia,
pistilsare stilldifferentiating on distalregion of receptacle Ⅶ End o董pistildifferentiation
Fig3 Primaryflowerbudsof‘Ai−Berry’strawberryateachdevelopmentalstage describedinTablel Barsindicate300FEm イチゴの花芽発育過程については,江口(18,19)がその概要を既に明らかにしている.しかし,発育段階の分
別基準やその呼称は研究者によって異なる(18,19,20,21,34,64,65,84,89)雄ずい分化期までだけについても,
森下・本多(84)は,分化開始期,分化期,がく片形成期,雄ずい形成期の4段階に分別しているのに対して, 成川ら(89)は,それらの間に,花房分化期,がく片初生期,花弁形成期の3段階を設けている 走査型電子顕微鏡による観察の結果,花弁の初生突起はがく片の間に生じるが,それに引き続いて,雄ずいのー7一 初生突起が形成され,その時間差はほとんどないと考えられた.また,それぞれの発育が進むまで,形態的に判 別することほ困難であった(第4図)そのため,雄ずい分化期までの発育段階については,森下・本多(84)に 準じて,Ⅰ:分化初期,Ⅰ:花房分化期,Ⅲ:がく片分化期,Ⅳ:雄ずい分化期の4段階に分別した. また,雌ずい分化期以降のイチゴ花芽の発育段階ほ,花粉・胚珠形成期(18,19,20,64,65)あるいは出菅期 (84)というように,雌ずいの分化発育段階ではなく,それ以外の花器や1香花原基の全体的な発育状態に・よっ て分別されている‖本研究においてほ,雌ずいの分化と発育について詳細に調査する必要を認めたため,従来の 分別基準は適用できなかった‖そのため,雌ずい分化期以降についてほ花床上での雌ずいの分イヒ程度によって∴ Ⅴ:雌ずい分化初期,Ⅵ:雌ずい分化中期,Ⅶ:雌ずい分化後期,Ⅶ:雌ずい分イヒ終了期の4段階に・分別し,そ の後の発育については分別しなかった‖ イチゴ雌ずいは花床基部から頂部へ,すなわち外側から中心へ向かって順次分化する(18,107).本研究におい ても,原則的にはこ.の法則に沿って雌ずいの分化ほ行なわれていたしかし,一部の個体においてほ逆転現象が 見られ,第5図に示したように,花床頂部から雌ずいの初生栗起が分化している場合があった‖ これらの個体の うら,雌ずいの分化が続いている場合は雌ずい分化後期,花床全体が雌ずいの初生突起で覆われている場合は雌 ずい分イヒ終了期と判定したり
Fig5Pistils differentiating from the apex of receptacle(flower developmental stage Ⅶ)in’Ai−Berry’strawberry Barindicates200pm Fig.4Petal(P)and stamen(S)primordia at earlystageassmallprotuberancesin‘Ai− Berry’strawberry
BarindicateslOOpm
第3節 イチゴの花序,花器および果実の呼称 】 花 序 イチゴの花序は,植物学上は有限花序の−・種の二出集散花序であり,花,花柄,小花柄の全体を含めて花房 (inflorescence)と呼ばれている(107).花房上の花の着生は,原則として第6囲に示すように規則正しく起こ るり基本的には,2節を有する主花柄の先端に1次の花(primaryflower)すなわち1香花(1stflower)が着生 する.その下の近接した2節からそれぞれ2節を葡する2次花柄分枝が伸びてその先端に2次の花(secondaryー8− flower)すなわち2,3香花(2nd,3rdflower)が着生し,以下3次の花(tertiaryflower)すなわち4∼7香花 (4th∼7thflower),4次の花(quaternaryflower)すなわち8∼15香花(8th∼15th董lower)と順次着生する すなわち,それぞれの花柄分枝は2本ずつ次の花柄を分枝し,頂端に・花を着生することを繰り返す.これが,通 常は.3∼5次の分枝で停止し,1花房上には7∼31花着生する それぞれの花柄分枝は2節を有す−るが,通常節間ほ基部から長一短一長の順で並び,中央の節間が短い(43) そのため対生しているように見えるが,発育段階をおって観察するとほぼ葉序と同じ2/5で着生しており,通 常は花柄基部に近い方の節から発生した分枝上の花の発育が優れ,先に開花する 斉藤(107)は,1香花が主花柄の頂卿こ着生するとし,花柄の分枝を小花柄と呼称している.しかし,小花柄 (pedicel)という術語は,通常直接花(果実)を着生する花柄部分をさすことから(144),本研究においてほ, 第7図に示したように,花房基部から花柄が分枝するまでの部分を主花柄(mainpeduncle),主花柄から分枝し た花柄の基部から次の分枝までの部分を2次花柄分枝 (secondarypedunclebranch),以下3,4次花柄分枝と し,それぞれの花(果実)の基部から花柄分枝の発生す る部分までをそれぞれの花の小花柄(pedicel)と呼称し た 上述のような,標準的な花序を持つ場合にほ,1次の 花がまず最初に開花し,ついで,2次の花が2,3番目 に順次開花する.通常は,引続き,基部に近い節から分 枝した2次の花(2香花)の花柄分枝上の3次の花が 4,5番目に開花し,3香花の花柄分枝上の3次の花 が,6,7番目に開花する‖以降,高次分枝になるにし たがって,開花の順序は乱れるが,同じ次数の花であれ ば原則として,早く開花した花の花柄から分枝した花柄 上の花ほど早く開花する上に,大きいことが多い.その ため花房内での花(果実)の順位をある程度統一・して比 較する必要がある しかし,品種または栽培条件によって,花房の形態は 大きく変化し,主花柄の第1節間が厚い場合(basal branching)や主花柄が3つ以上の節を持ち,2次の花 が3∼5花着生する場合,さらには,花房が複数発生す る場合も見られる.これらの場合,開花順に1,2,3 ・・・n香花とすると,4,5香花は2次の花であった り,3次の花であったりすることになる本研究におい ては,1花房内で最も早く開花する1次の花を1香花 (果),主花柄の最も基部から分枝した2次花柄分枝に 着生した2次の花を2香花(通常,最も早く開花する2 次の花),その次の節に着生した2次の花(通常,2番目 に開花する2次の花)を3香花(果)とした‖第8図に
Fig6.Diagram of normalin董lorescence of straw− berry
I:primary flower,I(:SeCOndaryflower,, 1,2.3‥:SequenCe Of flowering
Fig7 Termin0logyformainpeduncle,branchesof penduncles and pedicelsin strawberry
示したように.2次花柄分枝が3本以上発生した場合,花 器形質を調査する際には,2香花の花柄分枝の節から発 生した3次の花をそれぞれ下から4,5香花,3香花の 両厳に着生した花を6,7香花とし,2次の花の内3番 目以降に開花したものは調査対象から除外したい第Ⅳ章 においてポッ†植えの株を用いて果実の肥大を調査する 場合には,着果数を揃えるため2次の花柄分枝の内,基 部から2本を残し,それより上の節についた2次花柄分 枝(第8囲,破線部分)は摘除した イチゴの花房当りの開花数は,品種や栽培環境条件に よって大きな変動がみられる‖ これは先にも述べたよう にり おもに.花房の発生数(主花柄の発生本数)と2次以 下の花柄分枝が何次まで発生するかによって決定され Fig8Diagram of abnormalinflorescences of StraWberrywhichhavefour(1e董t)or three (Iight)secondary branches of peduncle: fruits shown with solid circles and per duncles shown with dottedlines were removedin the experimentsin chapter Ⅳ 1−5:SequenCe Of flowerlng る.花柄分枝が何次まで発生するかは.株によっても変異 が大きいが,少なくとも‘愛べり・−’についてみた場合には,最終の分枝上の花の大きさにはそれが3次分枝上 であろうと,5次分枝上であろうと大きな差が認められない.これを比較する場合にn次分妓上の花という表現 は適切とは考えられない.そこで,本研究においては,それぞれの花を大きさの上で大きく分別するため,低次 分枝上の花(主花柄あるいは2次花柄分枝の先端に着生した大きな花,すなわち大果となる素質を持つ)を上位 花,高次分枝上の花を下位花と呼称した. イチゴは秋期の低温と短日に反応して,クラウンの茎頂で花芽が分化し,頂花房(1stin董lorescence)を形成す る…その後20′、ノ30日遅れて,2∼3枚の葉を分化した腋芽が花芽を分化し,腋花房を形成する.通常の促成栽培 ではこれらの腋芽の内,先端の2芽を残して掃除し,2本の第1次腋花房を開花させる小 さらに,長期栽培にお いてはこれらの腋芽がそれぞれ1∼2本の第2次腋花房を着生し,これが繰り返される 本研究においても,整枝は慣行に従い,原則として第1次腋花房2本を残し,残りは腋芽の基部から摘険し た、また,第1次腋花房の内,先に開花した腋花房(通常ほ,上位節の腋芽,すなわち頂花房に.近い方の腋芽か ら発生した花房)を第2花房(2ndinflorescence)と呼び,その厳芽が花芽分化までに形成した真の数を花房間乗 数とした 2‖ 花器および果実 イチゴの雌ずい(pistil)は単一の心皮(carpel)からなり,第9囲に示した様な発育経過をたどる”すなわち, 雌ずいは最初花床上に乳頭上の突起として形成され(第9図A),発育が進むにしたがって,中央部が汚曲し始め る(第9図B)い その後,花柱部分が突出し,基部から両端が融合し始める(第9図C).さらに,全体が融合す るとともに花柱が伸長し始める(第9図D)い花柱が伸長し,子房が肥大するとともに,花柱基部がくびれ,花柱 基部と子房基部に毛じが形成される(第9図E,F)‖さらに,子房の肥大と花柱の伸長が引続き起こり(第9図 G),開花にいたる(第9図H) 子房中の胚珠は一・つで受粉受精し正常に発育すれば単一・の種子を奄する疫果(achene)となるハすなわち,花 床上で分化した心皮が発達して柱頭・花柱・子房の3部に分化し,単一の雌ずいが形成され,多数の雌ずいから なる雌ずい群(gynoecium)が花床上に形成されるそれぞれの雌ずいが受粉受精後発育して痩果となり,花床
Fig9Developing pistils of LAi−Berry’strawberry Barsindicate200FLm 範織が肥大して,いわゆる果実となるイチゴ果実は偽果であり,植物学的な果実は疫果であるが,通常は,疫 果が種子と呼ばれ,肥大した花床と多数の痩果を合わせて果実と呼ばれている(56,107) イチゴの場合,雌ずい・疫果ともに1心皮の発達したものであり,全発育段階を通じて心皮という呼称を当て ることができる.また,開花時には雌ずいと呼称し,受精後に疫果の呼称を用いている報告も見られるが(78), 雌ずいと疫果を分供しそのあいだに明確な境界を設けることほ難しい本研究においてほ,成熟した果実の痩果 ほ種子とし,花床上で初生突起として分化した段階から果実肥大期の間ほ,雌ずいまたほ雌ずい原基という呼称 を用いた 正常に受粉受精し,十分に.発育していると考えられるイチゴの種子の大きさは,果実成熟時には幅08′、ノ1.O mm,長さ12∼1.5mmである.それに対して,明らかに稔性を持たないと考えられる種子は,幅,長さともに1 /3∼1/2程度であるただし,発育がやや劣り,これらの中間的な大きさの種子も存在する.本研究におい ては,種子の発育程度をその大きさで判定し,幅05mm程度で明らかに発育不良であると考え.られる種子を不稔 種子とし,その他ほ稔実種子として特に分別しなかった 第4節 花器の詩形質の調査方法 本研究においては,‘愛ベリ1−’ の花床基部と頂部の雌ずいの発育差に着目し,この発育差が現われる要因の 解明を試みたい そこで,開花時における雌ずい並びに花床の発育程度を明らかにするためにり 以下に述べるよう な花器の諸形質を調査した‖調査にあたってほ,開花当日に花を採取し,ただちに2∼2.5%ダルクルアルデヒド で固定した.固定した試料は4℃で貯蔵し,適宜水道水で水洗後,実体顕微鏡下でマイクロメ・−・タ・−を用いて測
定した 雌ずい列数 花床上で分化した雌ずい数の指標とするために,開花 した花の花床上の雌ずい列の数を調査した.イチゴ雌ず いは,先に述べたようにり花床基部すなわち外側から, 頂部すなわち中心部へと順次分化する一都にほ乱れが 認められるが原則として,雌ずいは花床上で千鳥状に整 然と配置されている‖ これらの花軸と垂直に並んだ雌ず いを1列とし,第10図に示したように計測した 雌ずいの子房幅および花柱長 閑花時における雌ずいの発育程度の指標として,雌ず いの子房幅と花柱長を調査したい先に述べた雌ずい列の 内,最も基部の列に位置する雌ずいを花床基部の雌ずい とし,頂部に位置する雌ずいを花床頂部の雌ずいとし て,それぞれの子房幅,花柱長を第11図に示したように 測定した 花床径および花床高 開花時における花床の大きさの指標として,花床径と 花床高を第12図に示したように測定した‖花床径は雌ず いを含む花床の直径として,長径および澄径を測定し,そ れらの平均値を用いた.花床高ほ基部の雌ずいの子房の 下端から頂部の雌ずいの子房の先端までの長さとした
Fig 10 Pistils on receptacle and position of
rows of pistils on receptacleFig12 Methodsof measuringreceptacle
diameter(D)and height(H)
Fig.11Methodsofmeasuringovarydiameter(D)
一12− 第Ⅰ章 ‘愛ベリ1−’の奇形果発生要因 ‘愛ペリー・,は,頂果房1番果が正常に発育す れば60∼80gとなり,従来の品種には見られない 大栗性の品種である(第13図)いしかし,本来大果 となる素質を持つ各果房の上位果で,いわゆる “巾着果”・“先つまり果”と呼ばれる果実先端部 の肥大不良果や“先青果”・“先白果”と呼ばれる 果実先端部の着色不良果が多発する(第14図) このタイプの奇形果ほ,従来から‘ダナ1−ノ 等 の大果系の品種では,大筒を多肥栽培した場合に 発生しやすいことが指摘されていた.ただし,発 生頻度が低く,発生する品種も限られていたこと から,その発生要因についてほ明らかにされてい ない.栽培現地の事例から‘愛ペソ・−’において も‘ダナーノ と同様に大筒を多肥栽培した場合に 多発するようである.しかし,‘愛ペリー・’は新 しい品種であり,その生態的特性も明らかに.され ているとはいえず,奇形果の発生様相についても 明らかではない ‘愛ペソ、−, の奇形果発生を防止し,生産を安 定させるためにほ,まず,‘愛ペリー’の収盈構 成とともに,果実先端部の肥大不良による奇形果 の発生様相と発生要因を明らかにする必要があ る Fig13Normallarge董ruitof’Ai−Berry’(ca75g) Fig14Malformed fruitsof’Ai−Berry’,Which arecommonlycalledJSakitsumari一重ruit’ (1eft)or‘Sakiao−fruit’(right)
第1節 奇形果の発生様相
‘愛ベリ1−,の奇形果ほ各果房の上位果ほど多発する慣向が認められている・奇形果発生要因の究明に当たっ ては,まず奇形果発生の実態を明らかにする必要がある.そこで,促成栽培における奇形果の形成過程ならびに 奇形果の形態と果房内での着生位置による奇形果発生頻度の差異を明らかにするために以下の調査を行なった 1. 材料および方法 1985年7月17日仮植,9月13日定植として,‘愛ペリー’ を無加温電照促成栽培した1施肥盈は,10a当たり に3要素(N−P205−Ⅹ20)それぞれ,苗床6−6−6kg,本圃10−12−10kgとした.昼間,最高気温25℃ を目標に換気扇による強制換気を行い,ハウス内皮低気温が10℃を下回り始めた12月上旬から夜間2重被覆し, 夜温保持に努めた.開花開始前の11月上旬よりミツバチを放飼し,他の管理は慣行に従った−13− ハウス中央部の約100株の頂花房から収穫期に達した果実を順次採取し,奇形果の形成過程ならびに奇形果の 形態について調査を行った. 本実験においては,いわゆる正常な花房(第6図に示したように主花柄の第1節間が伸長し,2本の2次花柄 分枝を有する花房)の発生した個体のみを調査対象とし,主花柄の第1節間が短い(basalbranching)個体や主 花柄が3つ以上の節を持ち,2次の花が3花以上着生した個体,あるいは複数の花房が発生した個体ほ調査対象 から除外した 2. 鯨果および考察 第15,16因に示したように,本品種においては,果実先端部に・不稔種子をともなう奇形果が多発する程度が 軽く先端部の不稔種子が少ない場合ほ,果実の形態に対する影響は小さいしかし,先端部の不稔種子数が多く なるに従って果実先端部の肥大は不良となり,不稔種子が10列程度になると,いわゆる“巾着果”と呼ばれる状 態になるい また,果実先端部の着色が遅れる“先青果’’ほ,頂部に不稔種子をともない,最も先端部ほいわゆる “先つまり果”となりそれからいくらか基部よりの部分が着色せず,その場合着色の遅れる部分には不稔種子と 正常な種子が混在している場合が多くみられた
Fig“15.Variancein the shapeo董malformed fruitsaccompanied by the undeveloped
achenes on the distalregion of receptaclesin LAトBerry’strawberry
Left to right:Degree of malformationlto5
Degrees of malformation are describedin Table2
Fig16 Developmentofachenesonthedistalregionofmalformed fruits
Left to right:Degreeof malformationlto4
Degreesof malformation are describedin Table2and fruit shapes
are shownin Fig15
ー14一
Table2.Classification and description of degrees of fruit malformation
accompanied by undeveloped achenes on the distalregion of
receptacleFmit shapes and undeveloped achenes on the distal
regionof receptaclesforeachdegreeareshowninFig15and16
Degree Descriptiondeveloped achenes on the distalregiono董
ut
e
Normal shape and color with well receptacle
Fruit shapeis almost normalb receptacle are undeveloped,Or th
0(noI・mal)
1
but achenes are well developed
Distalregion of receptacleis not enlarged with more undeveloped achenes
thanl,Or distalregion of receptacleis stillwhite or green and some
achenes on that part are undeveloped
Nearlyone third of rows of achenes on the distalregion oL receptacle are
undevelopedand the partof receptacleisnotenlarged
Half or more rows o董 achenes on the distal region ofIeCeptaCle are undevelopedandthe parto王receptacleis notenlarged
5(severe) Wholereceptacleisnot enlarged,OrlittlepartofIeCeptaCleisenlargedwith
董ew developed achenes
Fig.17Development ofpistilsafteranthesis
Left to r・ight:0,4,8,16days after anthesis
そこで,奇形の程度を数値化して表現するた め,正常な果実を0,著しい奇形果を5として, 第2表および第15図のような基準を設定し,奇形 指数として表わした 奇形果の発生は,第3表に示すとおり1番果で 最も著しく,4,5着果での発生はわずかであっ たい また,6番果以降の果実では奇形果の発生は ほとんど認められなかった“特に,1番果は果実 歪が下位果と比較して大きいが,その変異が大き く,正常に発育した果実は70g以上に達するが, 著しい奇形果は30g未満であった一果実の成熟日 数(開花から成熟までの日数)も正常な果実は55 日程度であったのに対して,奇形指数4あるいは Table3 DiHerencesin fruit development
according to the sequence of 1lowering inaninflorescencein ‘Ai−Berry’strawberry
Fruit Degree o董 Maturation
weight(g)malformationy period(day)Ⅹ
FlowerB
48.3aV
2.6a 39.2b 2.Oab 37.3b lい8b 26.3c O.5c 24.6c O.3c 68.5a 66一.7a 66.9a 52.9b 52.. 6b t nd S rd th tト 1 2 3 4 5 罫1st:primary,2ndand3rd:SeCOndary,4thand5th: tertiaryflower,reSpeCtively y DegreesofmalformationaredescribedinTable2 ⅩDaysfromanthesistoharvest W MeanseparationbyDuncan,smultiplerangetest, 5%levelー151 5の果実は70日以上必要とし,著しい奇形果ほど長かった ‘愛ペソ・ノ の上位花を開花時に観察すると,花床頂部の雌ずいは基部の雌ずいと比較して色調が異なり,明 らかに小さい.このように,肉眼でも花床頂部の雌ずいが未発育であると判断された花は,著しい奇形果を形成 することが多かった..頂花房1香花のうち,開花当日に花床頂部の雌ずいが小さく未発育であると判断された花 の発育経過を第17図に示した‖ イチゴの雌ずいは,正常に・受粉受精した場合,開花数日後には柱頭が褐変し,受 精の確認が可能である‖開花8日後に.おいても頂部の雌ずいの柱頭は褐変せず未受精であると考えられたま た,開花16日後にほ,花床基部が肥大し始め,雌ずいも大きく肥大していたのに対して,花床頂部の雌ずいの柱 頭は褐変せず,子房,花床細胞ともに発育が不良であった. これらのことから,花床頂部の雌ずいが開花時に未発育であり,受精能力を持たないため,第16図に示したよ うな不稔種子が果実先端部に形成されると考えられる,また,イチゴ果実の肥大には受粉受精した雌ずいからの 刺激が必要であることが知られていることから(94,95),花床頂部の雌ずいが正常に受粉受精しないために,花 床観織が肥大せず,奇形果が発生すると考えられる. 第2節 窒素施肥量と苗質が奇形果発生と収量構成に及ぼす影響 一愛ペソ・ノ は大異性で,典型的な異数型の品種であり,促成栽培に適した品種であると考えられるしか し,その生態的特性が明らかにされる前に普及したため,収盈構成や花芽分化期等の栽培上重要な特性に関する 詳しい調査は行なわれていない…そこで,奇形果発生の問題とともに,促成栽培における生産力と収最構成につ いて明らかにしておくことは本品種の活用の上から有意義であると考えられる 栽培現地の事例から判断して,‘愛ペリー’の奇形果ほ,一般的に圃場の排水性が良好な砂質土の地域でほ比 枚的発生が少なく,排水性が悪い粘質土の地域で多く発生するようである。しかし,奈良県の水田地帯のような 粘質土の地域でも,元肥施肥盈を桂端に少なくし,無施肥の苗床で無仮植育苗した小首を花芽分化直前に定植す ることによって発生を軽減することができるとされている イチゴの生育収量に及ぼす窒素施肥の影響については多くの報告があるが,窒素施肥盈の適正値は,栽培地域 や作塾,品種によって大きく異なっている‖ただし,地力窒素の重要性を指摘し,有機質肥料や緩効性窒素肥料 の使用が望ましいという報賃が多い(11,37,38,62,71,74,105,130,131,132)い先に述べたように,‘愛ベリー’ では,元肥として多盈の窒素を施肥した場合に奇形果が多発する傾向が認められているまた,‘ダナ、ノ にお いても,大苗を多肥栽培した場合奇形果が多発するとされており,奇形果発生と窒素施肥,簡質との関係を明ら かにする必要がある 無仮植育苗ほ当初萎黄病回避対策と青首労力の軽減を目的として行われてきた育苗方法である しかし,その 他にも,‘宝交早生,の栽培において生産性の高位安定仁鶏冠状果の発生低下,花数の減少による小果の減少等 多くの長所が認められている(72)そこで,無仮構育苗を中心として∴革質と元肥窒素施肥最との関係から, ‘愛ペリー,の奇形果発生および収盈構成について調査した 1 材料および方法 第4表に記した条件で育苗した6種の筒と本圃への窒素施肥畳2水準を組み合わせて合計12処理区を設けた, 供試個体数は1処理区40株として,1986年9月20日に定植した,栽植密度は9000株/10aとし,瞳幅110cm,株
-16
Table4Growing conditions before planting
G少g Ⅹ
fNursery Z
tnt Fujimi June dddd − 0− O i2 F T 7 8 7 8 P P 工 ⊥ H H Iune July August hly August Takatsuki Takatsuki Takatsuki Takatsuki TakatsukiZ pot:Pottedon Tuly20inlO5cmpot,Field:Nottransplanted
y Fu5imi:Grownat1200m,Takatsuki:Grownat20minelevation
X AppliedtonurseryonJulylasammoniumsulphate
間20cmで2粂植えとした.栽培期間中の温度管理は,最低夜温6℃を目標に加温し,日中ほ最高気温25℃を目標 に換気扇による強制換気を行った..その他の栽培管理は慣行に従い,11月25日より間欠照明(10分/1時間)によ る電照を行った 育苗方法はポット育苗と無仮植育筒の2種類とし,ポット育苗区は山上げによる花芽分化促進処理の南無に よって2処理区(PF,二PT),無仮植育苗区ほ苗齢2水準と甫床への窒素施肥の有無によって4処理区(L7, L8,H7,Ⅱ8)として合計6の育苗処理区を設けた ポット育苗区は7月20日に本葉4∼5枚の苗を10..5cm黒ポリポットに鉢上げした用土はマサ土:ピーtモ スを1:1に混合し,苦土石灰と過燐酸石灰をそれぞれ用土1ゼ当り0.5gずつ混和した8月10日まで液肥(0 Ⅹ−F−1,N−P205−Ⅹ20=15−8−17)500倍液を週1回80mlずつ施与し,以後施肥は行わなかった ポット育苗区のうち,山上げを行うPF区は,8月18日に長野県富士見市の標高約1200mの地点に山上げし,9 月19日に山下ろしを行い,山上げしないPT区偲,大阪府高槻市の京大農場(標高約20m)でそのまま育苗を続 けた. 無仮植育濁区については,4月から5月にかけて発生した1次の子株を親株として用い,6月10日に瞳幅2 m,株間50cmで無肥料の苗床に定植した活着後UF化成(N−P205−K20=16−10−14)を親株1株当た り30g株元に施与した親株から6月中に発生したランナーほ7月1日に全て取り除き,無施肥で育酉を続ける L区と苗床に窒素追肥を行うⅡ区を設けた.H区腰7月1日に窒素成分で6kg/10aとなるように硫安を苗床全 面に散布し,耕起したそれぞれの処理区の内,7月下旬に着地発根した1次の子株をL7区」H7区,8月下 旬に着地発根した3次の子株をL8区,H8区とした 定植後の窒素肥培水準を異にするため,本隙に元肥として硫安で10kg/10a窒素を施与する多肥区と窒素施肥 を行わない少肥区の2水準の処理区暑設けた.ただし,P20515kg,Ⅹ2010kg,たい肥8t(それぞれ10a当 たり)は共通として,過燐酸石灰と硫酸加里で施与した‖なお,多肥区のみ活着後,窒素成分で3kg/10aとなる ように硫安で追肥を行ったい12月上旬以降は窒素施与処理の条件にかかわらず,追肥として液肥(くみあい2 号,N−P205−E20=10−4−8)を適宜施与した 奇形果発生については,第2表の基準に基づいて3月末までに収穫した全果実約11000個について判定し,同 時に個々の果実について果実重を測定した。.ただし,奇形程度5に相当する無肥大果と約6g未満と思われる小 果については,収穫対象外とし,約6g以上の果実について調査を行った−17− 2. 結果および考察 り定植時の苗の特性 定植時における各育苗処理区の苗の形質を第5表に示した.株重,クラウン径ともにH7>L7>PF>PT >H8>L8の順であった.ポッt育苗でほ高冷地で生育適温に近く気温が推移したと思われるPF区で地上部 盈がPT区と比べて大きかった.無仮植育苗区では発生時期が同じであっても窒素施肥により生育が促進され た.株重については.1次の子株であるL7,H7匡の間の差は小さかったが,3次の子株であるL8区とH8区 の間には大きな差が認められたいクラウン径についてほ,1次の子株でも窒素施肥の影響が強く現れたT/R 比はポット青首区∵で1.5∼2であったのに.対七て,無仮櫓育筒区は4前後で施肥畳,発生時期による大きな差ほ 認められなかった
Table5Characteristicsof nursery plants at planting time
Freshweight(g)T/R Crow㌣ N。。f
TreatmentZ
rati。
dヤ聖erl。a,eSy
Total Top Root ▲auU(mm) 9 3 2 2 2 6 <U O 1.〇〇 2 9 1 1 1 1 1 5 2 0 3 8 1 4 1 0 1 4 2 1 3 4 4 4 3 5 7 3 2 5 6 6 5 1 5 2 3 4 00 2 5 2 3 9 7 5 1 1 1 1 2 1 6 9 5 5 7 7 9 5 3 6 6 3 1 1 2 2 1 F T 7 8 7 00 P P L T⊥ T] H + 一 3 1 0 ︵XU 一 9 5 0 5 1
Z Treatments are describedin Table4
yTotalnumber ofleavesincluding those removed at digging time and
dead leaves 2)開花様相 頂花房開花期はPF区が最も早く,他の育苗処 理区と比べて約10日早かった.しかし,無位植育 苗区とPT区の間にほ大きな差は認められなかっ た.窒素施肥盈間には有意な差が認められ,多肥 区で少肥区.より1て日早かった第2花房開花期 についてもPF区カミ早かったが,他の育苗処理区 との差は約5日と頂花房と比較して小さくなっ た小 また,多肥区と少肥区の間にはほとんど差が 認められなかった(第18図,第6表).第19図に示 したように,頂花房分化後第2花房までに分化し た花房間乗数は,多肥区で2‖6枚,小肥区で2小3枚 となり∴窒素多肥によって増加したい PF区では
l・‡,_‖こ_‡三
P F PT L7 L8 H「 H8 ︼uむ∈︸d巴↑ 10 20 30 10 20 30 Nov Dec DateFig18 Effects of nitrogen application and nursery conditions on flowering in LAトBerry StraWberry
0:Okg−N/10a or●:10kg−N/10a was applied as ammonium sulphate,before plantingin
Culturing field
Treatments are describedin Table4 Bars indicate SE
花房間乗数が多肥区で3.5枚,少肥区∴でも2い9枚と
他の育苗処理区より多かったまた,最も小さな箇であったL8区でも花房間乗数は多肥区で32枚と多くなっ た‖しかし,少肥区∴では2い5枚となり,他の青首処理区との間の差は小さく,小首ほど花房間葉数は多くなり,窒
18 2 3 SむAd望−〇.〇Z 0 0 2 S︼むき○−−−〇.〇Z
P F P T L・7 L8 H7 H8
TreatrnentFig20 Effects of nitrogen application and nurser’y conditions on number of flowersinlst inflorescencein’Ai−Berry’strawberry Left(open):Okg−N/10a,right(solid):10kgp N/10a,aSdescribedin Fig18
Treatments are describedin Table4
P F P T L 7 L8 H7 H8
Treatment
Fig19Effectsof nitrogen application and nursery conditions on number ofleaves between lst and2ndinflorescence(bar・S)andinter− vals between flowering o董1st and2ndin− florescence(circles)in‘AトBerry’strawberry Lelt(open):Okg−N/10a,right(solid):10kg− N/10a,aSdescribedinFig18
Treatrnents are describedin Table4
Table6Analysisof variance for flowering date,number ofleaves betweenlst and2ndin董lorescence and number ofilowers Perlstinflorescence
Flowering date No ofy No of leaves flowers Source of VarlanCe lst
2nd IntervalZ
Nitrogen(N) ** ns ns ** 柑 Treatment(T) ** ** ** ** ** N x T ** ns ns * **ZIntervalsbetweenfloweringoflst and2ndinflorescence
y Noofleavesemerged betweenlstand2ndinflorescence
ns,*,**Non−Significantorsignificantat5%orl%levels,reSPeCtively 素施肥盈の影響を受けやすい傾向が認められた.頂花房と第2花房の開花期の差も同様の傾向を示し,花房間実 数の多い処理区で長かった、 第2花房の花芽分化についてほ頂花房の花芽分化が早まることによって早くなることが予想されるしかし, ‘とよのか,に低温暗黒処理を行った場合でも第2花房の開花期については無処理区との差は小さくなる(31)‖ 第2花房の花芽分化期は通常の促成栽培では頂花房の約1カ月後とされているが仁夜冷育苗などによって花芽分 化を促進した場合には,気温が比較的高く,日長も長い時期に当たる・さらに,定植活着後の日数が相当経過 し,窒素肥料を施与せずともたい肥由来の窒素や地力窒素を吸収し,窒素栄養状態が高く保たれるであろう“そ のために低温短日条件に対する反応が鈍くなり,頂花房の花穿分化が促進されて生理的に花芽分化しやすい状態 の株であっても特に分化促進を行わなかった株との差が小さくなったものと思われる・実際に,‘とよのか’で はNFTにおいて南濃虔培養液管理による第2花房開花期の遅れが認められていることから(146),第2花房に ついても頂花房同様に,高温長日条件下での花芽分化には窒素栄養が強く影響すると考えられる−19一 策20図に示したように,頂花房の花数はポッ†育苗区で多くなる慣向が認められた無仮植育苗区∴ではポら† 育苗区に比べて少なかったが,定植時の首が大きいはど花数ほ多くなったまた,多肥区で有意に多くなったが (第6表),18,H8区に見られるように小首でほ窒素施肥畳の影響は小さかった イチゴの開花数については,苗齢の進んだ大乱 また高窒素栄養に.よって増加することが知られている(7, 11,42,71,74,140,141).ポッ†育苗区での花数の増加ほ,苗として細板の盈が多く,定植後の活着が早いため窒 素吸収が盛んに行われたためであろう.小さな商で窒素施肥畠の影響が小さかった原因は,花芽分化時の甫の大 きさが花数を決定するのか,または小さな筒であれば窒素吸収が少なくなるためであるかの何れかもしくは両方 の原因によるものであろう 3)収量および果重別収量構成 第21図に株当たりの全収盈,30g以上の果実収塵と平均果実重を,第22図に全収穫果数,30g以上の果実数及 び30g以上の正常果数を示したい また,収盈構成要因に.関する分散分析結果を第7表に示した ︵叫︶呈晋∈霊n上品巴むAく ー00 600 500 400 300 200 ︵一udち\叫︶p一む⋮ト 8 6 ・4 召dち\s召上払︼dこ〇.〇Z 盲d石\∽ち︼〓〇.〇uヨ○↑ 0 5 0 2 P F p T L、7 L.8 H7 H8 Treatment
Fig22 Effectsof nitrogen application and nursery COnditions on number of malformed(open bars)and normal(solidbars)fruitsover30g and total number of harvested fruits (circles)in’Ai−Berry’strawberry
Left‥Okg−N/10a,right‥10kg−N/10a!
as describedinFig18
Treatments are describedin Table4 P F P T L7 L8 H7 H8
T−reatment
Fig21。Effects of nitrogen application and nursery COnditionsontotal(bars)andlarge(≧30g) fruit(solid bars)yields,and average fruit Weight(circles)in‘AiTBerry’strawberry Left:Okg−N/10a,right:10kg−N/10a, as describedin Fig18
Treatments are describedinTable4
Table7.Analysis of variance foryield components Yield Noof fruits harvested Normal Large Normal Large varlanCe fruits fruits Total fruits fruits
(≧30g)(≧30g) (≧30g)(≧30g)
Nitorgen * ** ns * ** * ** Treatment * ns ** * ** ns * ns,*,** Non−Significant or significant at5%orl%1evels,reSpeCtively
株当たり収畳ほⅡ7区を除けば少肥区で高かっ たが,その差は約12gで統計的に有意な差ではな かったしかし,青首処理区間にほ有意な差が認 められ,L8区では株当たり600g以上となり, 最も高い収屋が得られた.無仮植青首区の中でほ H7区が約520gと最も低く,無仮植育商でほ小 苗ほど収盛が多くなる傾向が認められた ポット 育蔑では,開花収穫始めが早く収穫期間の長かっ たPF区で多く約540gとなり,無仮植育苗の大 苗とほぼ同様の収盈が得られたが,PT区ほ約 510gと全育商処理区の中で最も低い収量であっ た 30g以上の果実収急にほ,育筒処理区間に有意 な差は認められなかった.しかし,少肥区で316 gで全収盈の57%であったのに対し,多肥区でほ 竃d石\sち︼〓〇.〇Z <10 <20 <30 <40 <50 <60 60≦ fruit weight(g)
Fig23Effects of nitrogen application on distribution Of fruit weightin‘Ai−Berry strawberry Composite of alltreatment Left(open):Okg−N/10a,right(solid):10kg−N/10a, as describedin Fig18 277gで51%と低かった小30g以上の収穫果数も少肥区で7.4個で全収穫果数の32%,多肥区で6.5個で26%と少 肥区が有意に多かった..また,育筒処理区間には有意な差が認められ,L8,H8区でほ76個であったのに対し て,⊥7,H7区ではそれぞれ65,6。6個となり,無仮植育苗では小西ほど大栗収畳が増加する傾向が認められ たい PF,PT区ほそれぞれ6い2,6‖4個で,ポッt青首区は無仮植の大筒より幾分劣った.それに対して∴全収 穫果数は多肥区で25小2個,少肥区∴で23。、1個と多肥区で有意に増加したが,育苗方法間には差が認められなかっ た… この,収穫果数の増加は第23園に示したように,20g未満の果実の増加によるものであり,30g以上の果実 数は窒素多肥によって明らかに減少した 収盈と平均果重の時期的な変勃について,代表的な変動を示したPI㌧工8,H7区の結果を舞24図に示し た,収穫始めは開花期の早かったPF区.が早く,その他の処理区間には大きな差は認められなかったH7区以 外では多肥区の方が早かった。全処理区ともに,第2果房の収穫盛期に当たる2月中旬に収唾のピー・クが認めら れたい30g以上の大栗の割合ほ多肥区でほ少肥区より早くから低下し,特にト“成り疲れ”と見られる収盈の低 下が認められた3月にほ大きく減少した..しかし,少肥区の小嘗(L8,E8)とPF区では3月においても30
g以上の大果の占める割合と平均果実重ほ比較的高く維持されたり全処理区ともに,2月以降10g未満の小果の
割合が増加し,それにともなって,平均果実重の低下が認められたが,その傾向は多肥区でより顕著であった イチゴの収量構成についてほ今までに多くの報告があり,従来から大苗を用いて,花数を確保することに重点 がおかれていたように思われる‖特に,露地栽培では発生時期が早く,クラウンの充実した苗を用いることに よって収量が高まることが報告されている(74)半促成栽培においても,大筒を用いて頂花房の花数を確保する ことによって,早期収盈が高まるとされている(8)これらのことから,促成栽培においても,クラウン径1 cm程度の大苗を育成する必要があると考えられてきたのであろうしかし,本実験において,最も高い収盈が得 られたのほわずか6.5gの苗を定植した18区であった伏原・高尾(31)は短日夜冷育苗による促成栽培におい て大苗と小首の間に初期収盈の差はないとしており,促成栽培での筒賀の問題については,苗の齢および大きさ と本田での栽培管理条件との関連について品種の特性をふまえた上で検討する必要があろう21 □<10g Ⅲ刀<30g ■30g≦ ︵β]ま首⊆\琶︼−品d︼空々 60 40 20 0 60 40 20 0 6 0 0 5 00 50
1一l
︵宇扁ち\叫︶p︻む叫ト 0 150 100 50 0Dec Jan Feb j Mar Dec Jan Feb Mar
Month Month
Fig24 Seasonalchangesinyields(bars)andaverage fruitweight(circles) in‘Ai−Berry’strawberry
Treatments are describedin Table4
Degree of malformation 4 6 0 0 言dち\苫︼−P心∈︼○︼一d∈−〇・〇Z 0 0 4 6 ︵次︶ごコ︼〓d∈︼OCちuOモOd <10 <20 <30 <40 <50 <60 60≦ Fruit weight(g)
Fig25Effects of nitrogen application on fruit malformation(bars) and proportion of normalfruits to the number of harvested fruit (circles)in each fruit weight categoryin‘Ai−Berry’strawberry Composite of a11treatment
−22− 4)奇形果発生 第7表に示したように30g以上の正常果収盈と 正常果数は少肥区で有意に増加した.また,青首 方法間にも有意な差が認められ,無仮植青首の小 常で多くなる傾向にあったい第25図に示したよう に,株当りの奇形果発生数ほリ30g以上の果実で は少肥区が/10∼30gの果実では多肥区が多くな る傾向にあったい また,多肥区においては奇形指 数4あるいは3の著しい奇形果が少肥区よりやや 多く発生した.特に,10∼30gの果実でその憤向 が強く,少肥区で奇形果発生が抑制される傾向が 認められた.しかし,全収穫異数も同様の傾向に あったため(第22図),果実重毎の正常果率,平均 奇形指数に.ほ,多肥区と少肥区の間にほとんど差 が認められなかったい さらに,頂花房花数と30g 以上の正常果収盈,正常果数との間に.それぞれⅠ =−0‖8333(P<0..001),Ⅰ・=−0‖8543(P< ︵訳︶s霊コ七−d∈旨⊂−〇⊂○−亡Od O O 0 0 0 8 4 6 O uO一芸E︼○−︻d∈10む巴叫む凸 P F P T L7 L8 H7 H8 Treatment
Fig26Effectsof nitrogen application and nurserycon− ditions on fruit malformation(bars)and propor− tion o董normalfruits to the number of harvested fruits(circles)in‘AトBerry’strawberry
Left(open):Okg−N/10a,right(solid):10kg−N/10a, as describedin Fig18
Treatments are describedin Table4 Degrees of malformation are describedin Table 2 0い001)の強い負の相関が認められ,頂花房花数 を少なくさせることが,正常な大果収盈の増加につながることが示された ただし,全収穫具についで比較した場合,無仮構育筒の小筒(18,H8)でやや平均奇形指数が小さく,正 常果率も高くなる傾向にあったが,育苗処理区周に,統計的に有意な差は認められなかった(第26図) 前節で示したように,榎虔の奇形果は果実肥大が不良となり,果実畳も小さくなる・多肥区では,本来大果と なる素質のある1∼3香花の上位の花が奇形果となるために肥大不良となり,全体の大果収畳も少なくなったの であろう√しかし,‘愛ペソ・−・, の奇形果発生については,全収盈,全収穫果数の中で比較すると,小果数の増 加や奇形果の肥大不良などの他の要因との関係から比較が難しくなると考えられ,個々の花についてさらに詳細 な検討が必要である
第3節 雌ずいの発育様相と奇形果発生に及ぼす人工受粉の影響
第1節で述べたように,奇形果が多発する‘愛ペリー,の上位花では,花床頂部の雌ずいは基部の雌ずいと比 牧して,小さく,色調も異なる.イチゴの雌ずいは花床基部すなわち外側から頂部すなわち中心部へと順次分化 する.このことから,花床頂部の雌ずいは基部の雌ずいと比較して発育が遅れており,開花時に受精能力を持た ないのであろうと推察されるいその結果,不稔種子が花床頂部に形成され,奇形果が発生するのであろう これらのことから,開花後花床頂部の雌ずいが十分に成熟するまで人工受粉を行い,受粉受精を促すことに よって奇形果発生を回避し得る可儲性が考えられる‖ 本実験では,奇形果発生と開花時における雌ずいの発育との関係を明らかにするため,開花後の追加的な人工 受粉が奇形果発生に及ぼす影響について調査した.また,雌ずいをTTC(2,3,5−triphenyltetrazoriumchloride) で染色し,柱頭の呼吸活性から雌ずいの発育程度を判定したー23− 1. 材料および方法 1985年7月17日に15×15cmで仮植した苗を9月13日に睦幅120cmで2条植えとして,株間20cmで定植し,無 加痘電應促成栽培した.施肥量は,10a当たりに3要素(N−P205−Ⅹ20)それぞれ,苗床6−6−6kg, 本囲10−10−10kgとし,硫安,過燐酸石灰,硫酸加里で施与した.昼間,最高気温25℃を目標に換気扇による強制換 気を行い,夜間ほ2重被覆し,夜温保持転努めた.開花開始前の11月上旬よりミツバチを放飼し,他の管理ほ慣 行に従った. ハウス中央部の約30株の頂花房から開花後−2′ 0,2,4,6,8,10日目に頂花房1∼3香花(果)と10 ∼12香花(4次花)を採取し,花弁,がく片∴堆ずいを取り除いた後1%TTCによる染色を行い(113),開花 後の日数と柱頭の呼吸活性の関係から,雌ずいの発育状況について調査した二 開花後の日数と雌ずいの受精能力との関係を明らかにするため,頂花房1∼3香花について,開花0,2, 4,8日後にそれぞれ約10花に袋掛けをし,奇形果の発生と種子の稔実について調査した.なお,袋掛けを行う までは毎日毛筆による人工受粉を実施した.また,開花8日後まで毎日毛筆による受粉を行う無袋区を設け,無 処直の対應区と比較した. 本実験においては,本章第1節と同様に,いわゆる正常な花房(第6図に示したように主花柄の第1節間が伸 長し,2本の2次花柄分故を有する)の発生した個体のみを調査対象とした. 2. 結果および考察 TTCによる染色の結果,肉眼で十分に発育していると思われる雌ずいの柱頭は赤紫色に着色し,呼吸が旺盛 な状態にあることを示していた.開花2日前の花では柱頭はすべて着色せず,1香花では開花当日でも花床の基 部側1/2程度の雌ずいしか着色していなかった.2日後,4日後となるにつれて花床頂部まで着色するように なり,8日後には染色した全ての花で頂部まで着色していた(第27囲).ただし,2,3番花については開花4日 後に頂部まで着色しているものもあり,6日後にははとんどの花で花床頂部まで着色していた.開花時の雌ずい の大きさは,花床の基部ほど大きく頂部ほど小さかった.特に,1香花の花床頂部の雌ずいは小さい上に色も薄 く,発育途上であると考えられた.開花10日後でも雌ずいの大きさの差は認められ,基部の子房はすでに肥大し 始めていたのに対して,頂部ではようやく開花時の基部の子房程度の大きさに達しているだけであった.10∼12 香花は開花2日後にはほとんどの花で花床頭部まで着色しており,子房の大きさも頂部と基部の差はわずかで
Fig,27.Changesin respiratoryactivityof the stigmain thelst(primary)flower of LAトBerry’strawberry.
Left toright:0,2,4,6,8,10days after anthesis.
Reacted withl%TTC,0.2M succinic acid and O.1M phosphate buffer (pH7.0)for2hr at37℃.