首都大学東京 博士(理学)学位論文(課程博士)
論 文 名 シロイヌナズナ
NRT1/PTR FAMILY (NPF)
の植物ホルモン 輸送体としての機能解析(英文)著 者 千葉 康隆
審査担当者
主 査 委 員 委 員 委 員
上記の論文を合格と判定する 平成 年 月 日
首都大学東京大学院理工学研究科教授会 研究科長
DISSERTATION FOR A DEGREE OF DOCTOR OF PHILOSOPHY IN SCIENCE
TOKYO METROPOLITAN UNIVERSITY
TITLE
:Functional analysis of Arabidopsis NRT1/PTR FAMILY (NPF) as plant hormone transporters
AUTHOR: Yasutaka Chiba
EXAMINED BY Examiner in chief Examiner
Examiner
Examiner
QUALIFIED BY THE GRADUATE SCHOOL OF SCIENCE AND ENGINEERING TOKYO METROPOLITAN UNIVERSITY Dean
Date
Contents
Contents
ⅠAbbreviations
ⅡAbstract 1
Introduction 2
Results 5
Discussion 11
Materials and methods 15
References 18
Statement of ethical compliance 25
Acknowledgement 26
Figures 27
Tables 35
Abbreviations
ABC, ATP-binding cassette AD, activation domain DBD, DNA binding domain JA-Ile, jasmonoyl-isoleucine
LC-MS/MS, liquid chromatography-tandem mass spectrometry NPF, NRT1/PTR FAMILY
SD, synthetic dextrose
Y2H, yeast two-hybrid
Abstract
NPF (NRT1/PTR FAMILY) proteins were originally identified as nitrate or di/tri-peptide transporters. However, recent studies revealed that the members of this transporter family also transport plant hormones, including auxin (indole-3-acetic acid; IAA), abscisic acid (ABA) and gibberellin (GA), as well as secondary metabolite, such as glucosinolates. In the present study, I developed the modified yeast two-hybrid (Y2H) systems with the receptor complexes for GA and Jasmonates (JA-Ile) to detect the hormone transport activities of proteins expressed in the yeast cells. By using the Y2H receptor complex systems for ABA, GA and JA-Ile, I determined the activities of Arabidopsis NPFs for ABA, GA and JA-Ile transport.
Several NPFs induced the formation of receptor complexes under relatively low hormone concentrations. Hormone transport activities were confirmed for some NPFs by direct analysis of hormone uptake of yeast cells using liquid chromatography-tandem mass spectrometry (LC-MS/MS). These results suggested that at least some NPF proteins function as hormone transporters in vivo.
[Main results including figures and tables in this study have been accepted as an article
entitled “Identification of Arabidopsis thaliana NRT1/PTR FAMILY (NPF) proteins capable
of transporting plant hormones” by Yasutaka Chiba, Takafumi Shimizu, Shinya Miyakawa,
Yuri Kanno, Tomokazu Koshiba, Yuji Kamiya and Mitsunori Seo in Journal of Plant
Research on Dec. 05, 2014]
Introduction
Plant hormones are a group of naturally occurring endogenous bioactive small molecules that induce various physiological responses throughout plant life cycles (Kende and Zeevaart 1997; Santner et al. 2009; Santner and Estelle 2009). Physiological responses mediated by hormones are regulated by several steps including biosynthesis, catabolism (inactivation), transport, perception and signal transduction. The balance between biosynthesis and catabolism, in association with transport, determines local hormone concentrations at a particular cell. On the other hand, recognition of hormones by receptors and transduction of the signals regulate sensitivities or responsiveness of a cell to the hormones. Plant hormones exist in plant tissues at low concentrations and thus these processes must be regulated very precisely.
In animals, hormones are synthesized in specific cell types and are transported to target
cells to induce physiological processes. In the case of plant hormones, transport of auxin
(indole-3-acetic acid; IAA) has been well characterized (Blakeslee et al. 2005; Petrasek and
Friml 2009). The plant specific PIN transporter family and some members in subgroup B of
ATP binding cassette (ABC)-type transporter family (ABCB) function as IAA exporters,
whereas amino acid permease-like AUX/LAFX family mediates IAA uptake into cells (Habets
and Offringa 2014; Swarup and Peret 2012; Yang and Murphy 2009). Another example of
which transport has been extensively studied is abscisic acid (ABA) (Boursiac et al. 2013; Seo
and Koshiba 2011). ABA has been considered as a signal that is produced in roots upon water
deficit and transmitted to leaves to induce stomatal closure (Jiang and Hartung 2008). On
the other hand, immunohistochemical analysis with antibodies raised against ABA
biosynthesis enzymes AtNCED3, AtABA2 and AAO3 in Arabidopsis revealed that these
enzymes are localized in vascular tissues, suggesting that ABA is synthesized in the tissues
(Endo et al. 2008; Koiwai et al. 2004). This intern indicates that ABA synthesized in the
vascular tissues has to be transported to the guard cells to induce stomatal closure. So far, four ABA transporters have been identified in Arabidopsis, and two of them are subgroup G ABC transporters AtABCG25 and AtABCG40 (Kang et al. 2010; Kuromori et al. 2010). It has been suggested that AtABCG25 is expressed in vascular tissues and involved in the export of ABA from inside to outside of cells at the site of ABA biosynthesis whereas AtABCG40 mediates ABA uptake into guard cells to close stomatal. It is interesting to note that the recently identified transporters for strigolactone and cytokinin are also member of ABCG (Kretzschmar et al. 2012; Ko et al. 2014; Zhang et al. 2014b). In petunia, an ABCG protein is required for exudation of storigolactone from root tissues to media. The Arabidopsis AtABCG14 is involved in the transport of different types of cytokinins between the shoot and root thorough xylem and/or phloem. The third ABA transporter is a member of the NRT1/PTR FAMILY (NPF) proteins. There are 53 genes encoding NPF proteins in Arabidopsis and some members in this family have been identified as nitrate or peptide (di/tri-peptide) transporters (Leran et al. 2014; Tsay et al. 2007). NPF4.6 that was previously characterized as a nitrate transporter NRT1.2 (Huang et al. 1999), however, our group revealed that the same protein functions also as an ABA transporter (Kanno et al. 2012).
NPF4.6 is expressed in vascular tissues and possibly regulates the amount of ABA transported from vascular tissues toward the guard cells in association with AtABCG25. As the fourth Arabidopsis ABA transporter, a DTX/MATE protein called AtDTX50 was recently identified as an ABA efflux transporter expressed in vascular tissues and guard cells (Zhang et al. 2014a). One may conclude from these facts that the ABA transport system is highly complex and redundant.
As well as NPF4.6, some members of NPF proteins were shown to transport substrates
other than nitrate or peptides. At least three additional Arabidopsis NPF proteins, NPF4.1,
NPF4.2 and NPF4.5 transported ABA in yeast (Kanno et al. 2012). Interestingly, NPF4.1
transported gibberellin (GA; GA
3) as well (Kanno et al. 2012). NPF6.3/NRT1.1/CHL1, which had been identified as a nitrate transporter, functions as an IAA transporter (Krouk et al.
2010). NPF2.10/GTR1 and NPF2.11/NRT1.10/GTR2 are involved in the transport of glucosinolates (Nour-Eldin et al. 2012). Furthermore, considering that dipeptides are substrates of some NPFs, I speculated that the amino acid-conjugated bioactive hormone jasmonoyl-isoleucine (JA-Ile) might also be transported by NPFs. Transport of GA and JA-Ile within plants has been suggested (Matsuura et al. 2012; Pimenta Lange et al. 2012). These findings suggest that NPF proteins transport not only nitrate or peptides but also various other compounds.
In the present study, I determined whether there are other NPF proteins capable of
transporting plant hormones. To perform a comparative investigation, I developed modified
Y2H systems to detect GA or JA-Ile transport activities of proteins expressed in the system
by using receptor complexes for respective hormones as our group reported previously for
ABA. By using the systems, the activities of 45 out of 53 Arabidopsis NPF proteins for
transport ABA, GA and JA-Ile were examined. The results showed that a relatively large
number of NPF proteins could transport one or more kinds of plant hormones.
Results
Construction of yeast two-hybrid systems to detect hormone dependent formation of receptor complexes
Our group previously developed a modified Y2H system to detect ABA transport activities of proteins expressed in the yeast cells by determining ABA dependent interaction between the ABA receptor PYR/PYL/RCAR and PP2C protein phosphatases (Kanno et al. 2012).
Arabidopsis PYR1 fused to the GAL4 DNA binding domain (DBD; DBD-PYR1), ABI1 fused to the GAL4 activation domain (AD; AD-ABI1) and a NPF protein were expressed in yeast strain PJ69-4A, in which marker gene ( HIS3 ) expression was activated by the ABA-dependent formation of DBD-PYR1/AD-ABI1 complexes. Next, yeast cells were inoculated in selection media. If NPF protein expressed in yeast had an ABA transport activity, it is expected that yeast cells would grow on selection media in the presence of relatively low concentration of ABA (Fig. 1A).
In our former assays, growth of yeast cells was determined after streaking on plates.
However, in the present study, I spotted a certain amount of cells on plates instead of streaking so as to compare the yeast growth semi-quantitatively. As our group reported previously, yeast cells expressing NPF4.6/NRT1.2/AIT1 grew on selection media containing 0.1 µM ABA whereas cells without the transporter did not. Yeast cells without NPF4.6/NRT1.2/AIT1 grew at relatively high concentrations (1 µM) of ABA, but not as rapidly as the cells expressing NPF4.6/NRT1.2/AIT1 (Fig. 1B).
It has been demonstrated that the receptors of GA, JA-Ile and IAA form protein
complexes in the respective hormone dependent manners: the GA receptor GID1 interacts
with DELLA proteins, the JA-Ile receptor COI1 interacts with JAZ proteins, and the IAA
receptor TIR1/AFB interacts with Aux/IAA proteins (Lumba et al. 2010). Therefore, I
expected that the Y2H systems with the receptor complexes for GA, JA-Ile and IAA will be useful for detecting hormone transport activities of NPFs (Fig. 2).
The Arabidopsis GID1a fused to GAL4 DNA binding domain (DBD; DBD-GID1a) and GAI fused to GAL4 activation domain (AD-GAI) were expressed in the yeast strain PJ69-4A of which marker gene ( HIS3 ) expression is activated by formation of the DBD-GID1a and AD-GAI complex (Fig. 2A). Our group has shown that NPF4.1/AIT3, which was originally identified based on its activity to transport ABA, could also transport GA (GA
3) when determined by a direct analysis with LC-MS/MS (Kanno et al. 2012). Thus, I tested whether the GA transport activity of NPF4.1/AIT3 could be detected by using the Y2H transport assay system with the GA receptor complex (Fig. 3). GA
1, GA
3and GA
4were used as substrates because they are recognized by the GA receptor. GA
1and GA
4are major bioactive GAs synthesized in higher plants. GA
3is a major GA produced in the fungus Gibberella fujikuroi but has biological activities like GA
1and GA
4when exogenously applied to plants. The three GAs have different affinity to the receptors (Nakajima et al. 2006), therefore I determine optimum concentrations of the GAs used for the assay. Significant growth of yeast without NPF4.1/AIT3 was observed on media containing 1 µM GA
1or GA
3, although cells expressing NPF4.1/AIT3 grew better than the control (Fig. 3A and B). In contrast, only cells expressing NPF4.1/AIT3 grew at 0.1 µM GA
1or GA
3. As expected from a higher affinity of GA
4to the GA receptor GID1a compared to those of GA
1and GA
3, the yeast containing DBD-GID1a and AD-GAI constructs grew well even when GA
4concentration was 0.1 µM both in the presence and absence of NPF4.1/AIT3. However, when GA
4concentration was 0.002 µM, I could observe a better growth rate of the yeast cells containing NPF4.1/AIT3 compared to the cells without NPF4.1/AIT3 (Fig. 3C). These results indicated that the Y2H system with the receptor complex could be used to detect GA transport activities of NPFs.
The Arabidopsis JA-Ile receptor COI1 fused to DBD and JAZ protein fused to AD were
introduced in the yeast cells to construct Y2H system (Fig. 2B). Although I tested two JAZ proteins, JAZ1 and JAZ3, both of them have been demonstrated to interact with COI1 in JA-Ile dependent manners, only JAZ3 induced the expression of the selection marker gene in the presence of JA-Ile. In my experimental conditions, the yeast containing DBD-COI1 and AD-JAZ3 grew better on the selection media in the presence of 100 µM JA-Ile compared to when 0 or 50 µM JA-Ile was present in the media (Fig. 4). I expected that this system will be useful to determine JA-Ile transport activities of NPFs, although I did not have positive controls to confirm the possibility.
I also attempted to construct Y2H systems to detect IAA dependent interaction between the TIR1/AFB receptor and Aux/IAA protein (Fig. 2C). I used AD-IAA1, AD-IAA3 and AD-IAA5 as interactors of DBD-TIR1. However, none of the combinations gave yeast growth on selection media even when IAA concentrations were 100 µM. Thus I could not apply the Y2H system with the receptor complex to detect IAA transport activities of NPFs.
Detection of hormone transport activities of NPF by the Y2H system
To determine whether there are unidentified Arabidopsis NPF proteins capable of transporting hormones, I cloned as many NPF cDNAs as possible. 45 out of 53 Arabidopsis NPF cDNAs were cloned into a yeast expression vector and introduced into the Y2H systems with hormone receptor complexes.
ABA transport activities of NPFs were detected by determining the growth of yeast containing DBD-PYR1 and AD-ABI1 on selection media in the presence of 0.1 µM ABA (Fig.
5). Although our group have already examined the ABA transport activities of 11 NPFs
(NPF4.6/NRT1.2/AIT1, NPF4.5/AIT2, NPF4.1/AIT3, NPF4.2/AIT4, NPF6.3/NRT1.1,
NPF4.4/NRT1.13, NPF4.3/NRT1.14, NPF6.2/NRT1.4, NPF6.4/NRT1.3, NPF6.1 and
NPF4.7) that were formerly categorized to group I according to phylogenetic relationship
(Tsay et al. 2007), these NPFs were included in the present assay so as to compare the activities of possibly all NPFs in the same condition. The growth of yeast spotted at different cell concentrations were compared. The yeast cells expressing NPF2.5, NPF4.1/AIT3, NPF4.5/AIT2, NPF4.6/AIT1 and NPF5.2/PTR3 grew better even when a lesser amount of cells (10
3) was inoculated, suggesting that these NPFs efficiently transported ABA into the yeast cells. Significant growth of yeast cells expressing NPF1.1, NPF8.2/PTR5, NPF5.1, NPF5.3 and NPF5.7 were observed at least when a higher concentration of cells (10
4) were inoculated, suggesting that these NPFs also had activities to transport ABA. Although our group previously showed a weak ABA transport activity of NPF4.2/AIT4, the activity was under detection level in this assay condition. Actually, the yeast expressing NPF4.2/AIT4 grew slightly better than control cells when much higher amount of cells (10
5) were inoculated and incubated for a long period (Fig. 6).
Next, I determined the activities of NPFs to transport GA
1, GA
3and GA
4using the Y2H system with DBD-GID1a and AD-GAI (Fig. 5). The yeast growth on selection media containing 0.1 µM GA
1suggested that NPF2.3, NPF2.5, NPF2.12/NRT1.6, NPF2.13/NRT1.7, NPF1.1/NRT1.12, NPF1.2/NRT1.11, NPF3.1/Nitr, NPF4.1/AIT3, NPF4.2/AIT4, NPF5.1, NPF5.2/PTR3 and NPF5.7 efficiently transported GA
1into the yeast cells. The yeast cells expressing NPF2.4, NPF2.6, NPF2.7/NAXT1, NPF2.10/GTR1, NPF8.2/PTR5, and NPF5.6 grew better than the control cells in the same condition, suggesting that they also have activities transport GA
1. Similar tendency of yeast growth was observed when 0.1 µM GA
3was used as a substrate except that the yeast cells expressing NPF2.6, NPF8.2/PTR5 and
NPF5.6 did not grow in this condition. Most of the NPFs that induced the yeast growth in the
presence of 0.1 µM GA
3or GA
1also induced the yeast growth in response to 0.002 µM GA
4,
although some NPFs such as NPF2.12/NRT1.6, NPF3.1/Nitr and NPF4.2/AIT4 were less
effective.
The activities of NPFs to transport JA-Ile were also determined by examining their effects on the growth of yeast cells containing DBD-COI1 and AD-JAZ3 on selection media in the presence of 50 µM JA-Ile (Fig. 5). The yeast cells expressing NPF1.1/NRT1.12, NPF1.2/NRT1.11, NPF4.1/AIT3, NPF8.1/PTR1, NPF8.2/PTR5 and NPF5.7 exhibited significant growth either when a lesser or higher amount of cells was inoculated. In addition, the yeast expressing some other NPFs such as NPF2.4, NPF2.6, NPF2.7/NAXT1, NPF2.10/GTR1, NPF2.13/NRT1.7, NPF3.1/Nitr and NPF5.1 showed faint growth.
Direct examination of hormone transport activities of NPFs by liquid chromatography-mass spectrometry (LC-MS/MS)
The results presented in Fig. 5 suggested that several NPF proteins transported ABA, GA and/or JA-Ile. However, as the hormone transport activities were detected indirectly based on yeast growth, I could not exclude the possibility that the NPFs induced growth independently of their hormone transport activities. Our group had demonstrated that NPF4.6/NRT1.2/AIT1 transported ABA whereas NPF4.1/AIT3 transported ABA and GA
3, by directly measuring the hormones taken up by yeast cells using LC-MS/MS (Kanno et al.
2012). In the present study, I could confirm that NPF4.6/NRT1.2/AIT1 transported ABA, and
NPF4.1/AIT3 transported ABA and GA
3. In addition, as expected from the Y2H-based
transport assays described in Fig. 5, it was shown that NPF4.1/AIT3 transported GA
1, GA
4and JA-Ile whereas NPF4.6/NRT1.2/AIT1 did not (Fig. 7). Furthermore, I determined
hormone transport activities of several other NPF proteins. Based on the results shown in
Fig. 5, I selected 4 NPFs (NPF2.5, NPF5.1, NPF5.2/PTR3 and NPF8.2), 5 NPFs
(NPF1.1/NRT1.12, NPF1.2/NRT1.11, NPF4.2/AIT4, NPF5.1 and NPF8.2) and 3 NPFs
(NPF5.1, NPF5.7 and NPF8.2/PTR5) for determination of ABA, GA and JA-Ile transport
activities, respectively. All these NPFs showed expected hormone transport activities (Fig. 8)
indicating that the results of Y2H assays reflect actual hormone transport activities of NPF
proteins.
Discussion
In the present study, I demonstrated that the Y2H systems with the receptor complexes for GA and JA-Ile, as well as ABA, could be used to detect respective hormone transport activities (Figs. 1-5). As I expected, several NPF proteins could enhance the hormone dependent formation of receptor complexes in yeast cells in response to exogenous application of hormones (Fig. 5). LC-MS/MS analysis confirmed that yeast cells expressing the NPFs accumulated the expected hormones at significantly higher levels compared to control cells when incubated with the compounds (Figs. 7, 8).
Our group previously showed that 4 members of NPFs, NPF4.1/AIT3, NPF4.2/AIT4, NPF4.5/AIT2 and NPF4.6/NRT1.2/AIT1, had activities to transport ABA by the Y2H and/or direct assay with LC-MS/MS, and one of these NPFs, NPF4.1/AIT3 could also transport GA
3when determined by a direct assay with LC-MS/MS (Kanno et al. 2012). The present study
revealed that a relatively large number of NPFs other than NPF4.1/AIT3, NPF4.2/AIT4,
NPF4.5/AIT2 and NPF4.6/NRT1.2/AIT1 were capable of transporting hormones ABA, GA
and/or JA-Ile. Note that I cannot clearly define the presence or absence of hormone transport
activities for several reasons. First, the rate of yeast growth was determined at a specific time
point for each substrate: much numbers of NPFs tended to promote yeast growth if the yeast
were incubated for a longer time period whereas much less number of NPFs promoted the
yeast growth after a shorter time period. Second, I could not determine the levels of active
NPF proteins expressed in the yeast cells. Because our group previously found that the
addition of a tag to the NPF protein for immuno-detection inhibited its transport activities
(Kanno et al. 2012), I expressed NPFs without tag in the present assay. Also, it has to be
noted that the chemical nature of substrates used in the present assays differ in terms of
their affinity to the receptors, membrane permeability, stability and so on. Thus different
concentrations of substrate were used for the assay, and yeast growth was determined at
different time point depending on hormones. Therefore, I cannot simply compare relative strength of transport activities of NPFs for these hormones. Nevertheless, it would be possible to discuss substrate preferences or substrate specificities of NPFs to some extent. For example, NPF4.6/NRT1.2/AIT1 transported ABA but not GA (GA
1, GA
3, GA
4), JA-Ile, JA or IAA, suggesting that this protein has a high specificity for ABA (Fig. 5; Kanno et al. 2012).
Therefore, NPF4.6/NRT1.2/AIT1 has been shown to functions as an ABA transporter in vivo (Kanno et al. 2012). Similarly, NPF8.1/PTR1 was relatively specific to JA-Ile (Fig. 5). On the other hand, NPF4.1/AIT3 showed relatively broad substrate specificity for ABA, GA and JA-Ile (Fig. 5), however, the protein did not transport JA or IAA (Kanno et al. 2012).
NPF2.12/NRT1.6 and NPF5.6 are relatively specific to GAs, but the two proteins preferred different GA species (Fig. 5). These data suggest that there are some mechanisms for substrate recognition. In many cases, closely related NPFs showed similar substrate specificities each other (Fig. 5). However, NPFs capable of transporting a particular substrate are widely distributed in the family member, and thus phylogenetic relationship is not sufficient to predict the substrate of a particular NPF protein. Recently 3D structure of NPF6.3/NRT1.1/CHL1 was resolved and mechanisms by which nitrate is recognized by the protein as a substrate have been discussed (Parker and Newstead 2014; Sun et al. 2014). It would be interesting to study how NPFs recognize various substrates based on 3D structures.
Several NPFs have been characterized as nitrate or peptide transporters. It has been reported that nitrate transporters NRTs did not transport peptides whereas peptide transporters PTRs did not transport nitrate (Tsay et al. 2007). However, recent studies revealed that multiple compounds could be substrates for a NPF protein (Kanno et al. 2012;
Krouk et al. 2010; Nour-Eldin et al. 2012). NPF6.3 has been well characterized as a dual
affinity nitrate transporter NRT1.1/CHL, but the same protein was shown to function as an
IAA transporter (Krouk et al. 2010). Similarly, NPF4.6 that was originally identified as a low
affinity transporter NRT1.2 functions as an ABA transporter (Kanno et al. 2012). Although the functions of NPF2.10 and NPF2.11 have been unidentified until recently, they were identified as glucosinolate transporter GTR1 and GTR2, respectively (Nour-Eldin et al. 2012).
These data indicate that NPFs transport a variety of compounds. In the present study, I identified additional NPFs capable of transporting ABA, GA and or JA-Ile, and it is possible that at least some of them function as hormone transporters in vivo. Considering that a relatively large number of NPFs transported same substrate, the functions might be highly redundant. Nevertheless, several mutants defective in NPFs have been shown to have interesting phenotypes (Almagro et al. 2008; Fan et al. 2009; Guo et al. 2003; Hsu and Tsay 2013; Huang et al. 1999; Karim et al. 2007; Komarova et al. 2008; Li et al. 2010; Lin et al.
2008; Wang and Tsay 2011). Although the phenotypes have been discussed in the context of
nitrate of peptide transport, it is possible that the phenotypes are caused by the defects in
hormone transport. For example, mutants defective in NPF2.13/NRT1.7 (nrt1.7) showed
growth retardation upon nitrogen starvation (Fan et al. 2009). It is well known that
GA-deficient or GA-insensitive mutants are dwarf. Therefore, it could be possible that the
phenotype observed in nrt1.7 was caused by a defect in GA transport. It is interesting to
speculate that seed abortion observed in mutants defective in NPF2.12/NRT1.6 (Almagro et
al. 2008) might be related to GA transport, although the role of GA during seed development
is unclear. On the other hand, mutants effective in NPF5.2/PTR3 (ptr3) were susceptible for
pathogen infection (Karim et al. 2007). It is possible that the phenotype might be related to
ABA transport activity of NPF5.2/PTR3, because it has been reported that ABA is involved in
responses to pathogen infection (Lee et al. 2012). Consistent with our observation that
NPF2.10/GTR1 transporter GA, it was demonstrated very recently that NPF2.10/GTR1
functions not only as a glucosinolate transporter but also as a GA transporter in vivo (Saito et
al. 2015). In the case of NRT1.1, competition between two substrates, nitrate and IAA have
important role in physiological responses (Krouk et al. 2010). Likewise, it would be possible that competition between nitrate/peptide and hormones or competition between several hormones have important physiological roles.
The Y2H systems with receptor complexes will be applicable for other plant hormones such as strigolactone and salicylic acid (Fu et al. 2012; Hamiaux et al. 2012; Jiang et al. 2013;
Zhou et al. 2013). It would be also possible to identify new plant hormone transporters other
than NPF by screening against cDNA libraries as was done for ABA (Kanno et al. 2012).
Materials and Methods
Construction of Y2H systems
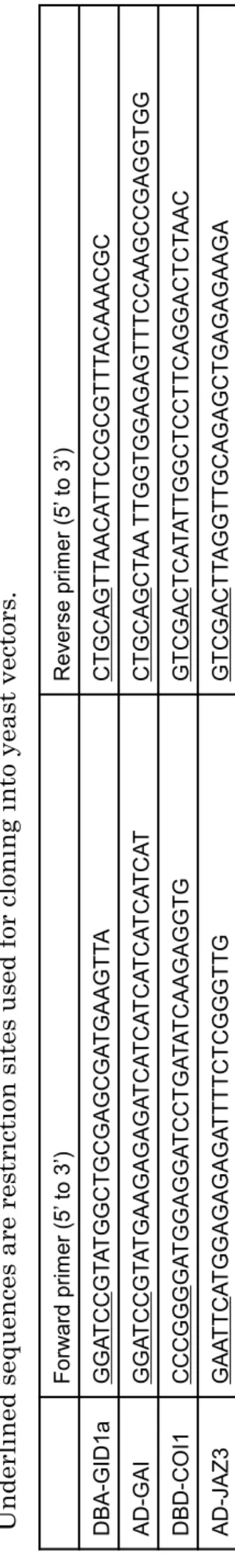
For expression of DBD-GID1a and DBD-COI1 in yeast, GID1a (At3g05120) and COI1 (At2g39940) cDNAs were amplified with primer combinations presented in Table 1 and cloned into pT7-Blue (Novagen). After sequencing, the GID1a and COI1 cDNAs were excised by digestions with Bam H I and Pst I , or with Sma I and Sal I, respectively, and cloned into the pGBT9 vector.
For expression of AD-GAI, AD-JAZ1 and AD-JAZ3 in yeast, the GAI (At1g14920), JAZ1 (At1g19180) and JAZ3 (At3g17860) cDNAs were amplified with primer combinations presented in Table 1. After sequencing, inserted cDNAs were excised by digestions with Bam H I and Pst I (for GAI ), or with EcoR I and Sal I (for JAZ1 and JAZ3 ), and cloned into the pGAD424 vector.
DBD-PYR1 and AD-ABI1 constructs were generated as described previously (Kanno et al.
2012).
Cloning of NPF cDNAs
NPF cDNAs were amplified with primer combinations listed in Table 2, and cloned into
pENTR/D-TOPO (Invitrogen) or pDONR207 (Invitrogen) before introduction into yeast
expression vector pYES-DEST52 or its derivative. For cloning into pDONR207, cDNAs
amplified with primers that contain a part of att B1 (forward primers) and att B2 (reverse)
sequences on their 5’ region were first cloned into pT7-Blue, and then re-amplified with
primers 5’-GGGGACAAGTTTGTACAaaaaagcaggct-3’ (forward primer) and
5’-GGGGACCACTTTGTACAagaaagctgggt-3’ (reverse primer) to add complete att B1 and
att B2 recombination sequences, respectively. Nucleotides shown in lowercase overlap with
the part of att B1 and att B2 recombination sequences added to the first primers. The amplified cDNA with recombination sequences were introduced into pDONR207 by BP reactions. After sequencing, NPF cDNAs were cloned into pYES-DEST52 of which GAL1 promoter had been replaced with ADH1 promoter (for Y2H assays and direct assays) or unmodified pYES-DEST52 (for direct assays).
Eleven NPF cDNAs in subgroup 4 and 6 were prepared as described previously (Kanno et al., 2012).
Y2H transport assay
The DBD, AD, and NPF constructs was transformed in the yeast strain PJ69-4A according to standard protocols publishes elsewhere. Yeast cells derived from 10 independent original transformants cultured overnight in liquid media [synthetic dextrose (SD); -Trp, -Leu, -Ura]
were collected by centrifugation and re-suspended in buffer (10 mM Tris-HCl, 1 mM EDTA, pH 7.5) so that OD
600became 1 (approximately 1×10
7cells/ml), 0.1 or 0.01. Ten micro l of cell suspensions (1×10
5, 1×10
4or 1×10
3cells) were inoculated on plates (SD, -Trp, -Leu, -Ura, -His) containing desired concentrations of hormones and incubated at 30℃ .
Direct transport assays with by LC-MS/MS
Direct hormone transport assays were performed essentially as described previously (Takeuchi et al. 2014), with some modifications. Vectors to express NPFs in yeast cells were constructed with the original pYES-DEST52 (Invitrogen) except NPF4.6.
NPF4.1, NPF4.2 and NPF4.6 cloned in pYES-DEST52 of which GAL1 promoter had been
replaced with ADH1 promoter were introduced in the yeast strain PJ69-4A. NPF1.1, NPF1.2,
NPF2.5, NPF5.1, NPF5.2, NPF5.7 and NPF8.2 cloned in unmodified pYES-DEST52 were
introduced in the yeast strain INV Sc 1.
For NPF1.1, NPF1.2, NPF2.5, NPF5.1, NPF5.2, NPF5.7 and NPF8.2, yeast cells were cultured in SD (synthetic dextrose) media containing 2 % (w/v) galactose and 1 % (w/v) raffinose instead of 2 % (w/v) glucose.
Hormones were added to reaction mixtures at 10 µM. ABA and GA
3were extracted and
purified from yeast cells as described previously (Kanno et al. 2012). GA
1, GA
4and JA-Ile
were extracted and purified as GA
3was. LC-MS/MS analysis was performed with a Nexcera
(Shimazu)/Triple TOF 5600 (AB SCIEX) system with an ZORBAX Eclipse XDB-C18 column
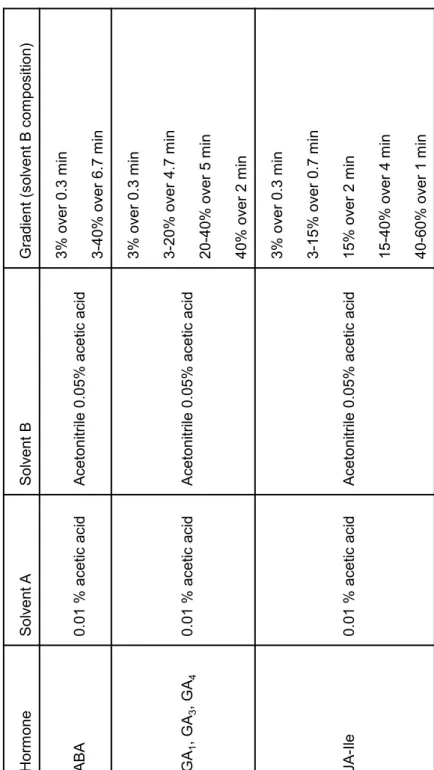
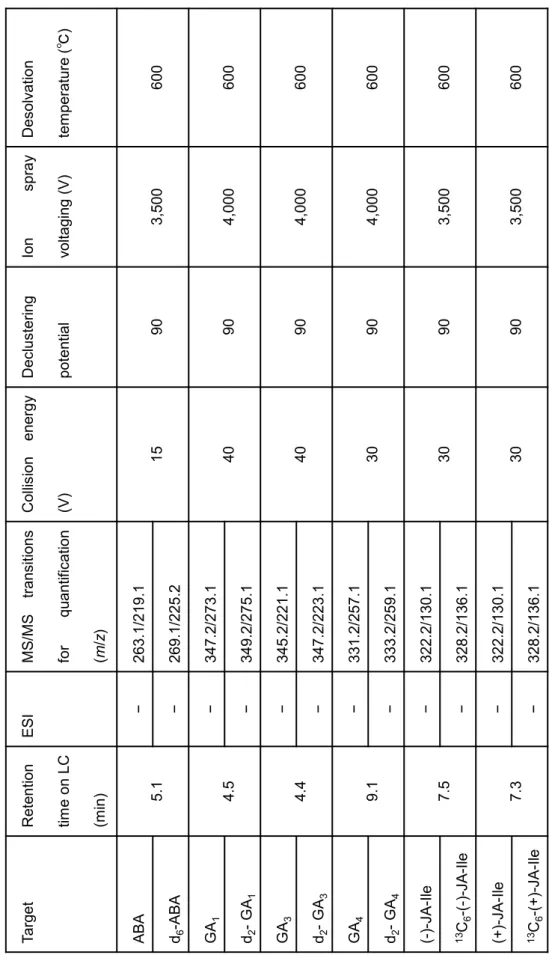
(Agilent). LC conditions and MS/MS parameters are shown in Tables 3 and 4.
References
Almagro, A., Lin, S.H. and Tsay, Y.F. (2008) Characterization of the Arabidopsis nitrate transporter NRT1.6 reveals a role of nitrate in early embryo development. Plant Cell 20:
3289-3299.
Blakeslee, J.J., Peer, W.A. and Murphy, A.S. (2005) Auxin transport. Curr. Opin. Plant Biol. 8:
494-500.
Boursiac, Y., Leran, S., Corratge-Faillie, C., Gojon, A., Krouk, G. and Lacombe, B. (2013) ABA transport and transporters. Trends Plant Sci. 18: 325-333.
Endo, A., Sawada, Y., Takahashi, H., Okamoto, M., Ikegami, K., et al. (2008) Drought induction of Arabidopsis 9-cis-epoxycarotenoid dioxygenase occurs in vascular parenchyma cells. Plant Physiol. 147: 1984-1993.
Fan, S.C., Lin, C.S., Hsu, P.K., Lin, S.H. and Tsay, Y.F. (2009) The Arabidopsis nitrate transporter NRT1.7, expressed in phloem, is responsible for source-to-sink remobilization of nitrate. Plant Cell 21: 2750-2761.
Fu, Z.Q,, Yan, S., Saleh, A., Wang, W., Ruble, J., et al. (2012) NPR3 and NPR4 are receptors for the immune signal salicylic acid in plants. Nature 486: 228-232.
Guo, F.Q., Young, J. and Crawford, N.M. (2003) The nitrate transporter AtNRT1.1 (CHL1)
functions in stomatal opening and contributes to drought susceptibility in Arabidopsis. Plant
Cell 15: 107-117.
Habets, M.E. and Offringa, R. (2014) PIN-driven polar auxin transport in plant developmental plasticity: a key target for environmental and endogenous signals. New Phytol. 203: 362-377.
Hamiaux, C., Drummond, R.S., Janssen, B.J., Ledger, S.E., Cooney, J.M., Newcomb, R.D., and Snowden, K.C. (2012) DAD2 is an α/β hydrolase likely to be involved in the perception of the plant branching hormone, strigolactone. Curr. Biol. 22: 2032-2036.
Hsu, P.K. and Tsay, Y.F. (2013) Two phloem nitrate transporters, NRT1.11 and NRT1.12, are important for redistributing xylem-borne nitrate to enhance plant growth. Plant Physiol. 163:
844-856.
Huang, N.C., Liu, K.H., Lo, H.J. and Tsay, Y.F. (1999) Cloning and functional characterization of an Arabidopsis nitrate transporter gene that encodes a constitutive component of low-affinity uptake. Plant Cell 11: 1381-1392.
Jiang, F. and Hartung, W. (2008) Long-distance signalling of abscisic acid (ABA): the factors regulating the intensity of the ABA signal. J. Exp. Bot. 59: 37-43.
Jiang, L., Liu, X., Xiong, G., Liu, H., Chen, F., et al. (2013) DWARF 53 acts as a repressor of strigolactone signaling in rice. Nature 504: 401-405.
Kang, J., Hwang, J.U., Lee, M., Kim, Y.Y., Assmann, S.M., Martinoia, E. and Lee, Y. (2010)
PDR-type ABC transporter mediates cellular uptake of the phytohormone abscisic acid. Proc.
Natl. Acad. Sci. USA 107: 2355-2360.
Kanno, Y., Hanada, A., Chiba, Y., Ichikawa, T., Nakazawa, M., Matsui, M., Koshiba, T., Kamiya, Y. and Seo, M. (2012) Identification of an abscisic acid transporter by functional screening using the receptor complex as a sensor. Proc. Natl. Acad. Sci. USA 109: 9653-9658.
Karim, S., Holmstrom, K.O., Mandal, A., Dahl, P., Hohmann, S., Brader, G., Palva, E.T. and Pirhonen, M. (2007) AtPTR3, a wound-induced peptide transporter needed for defence against virulent bacterial pathogens in Arabidopsis. Planta 225: 1431-1445.
Kende, H. and Zeevaart, J. (1997) The Five "Classical" Plant Hormones. Plant Cell 9:
1197-1210.
Ko, D., Kang, J., Kiba, T., Park, J., Kojima, M., et al. (2014) Arabidopsis ABCG14 is essential for the root-to-shoot translocation of cytokinin. Proc. Natl. Acad. Sci. USA 111: 7150-7155.
Koiwai, H., Nakaminami, K., Seo, M., Mitsuhashi, W., Toyomasu, T. and Koshiba, T. (2004) Tissue-specific localization of an abscisic acid biosynthetic enzyme, AAO3, in Arabidopsis.
Plant Physiol. 134: 1697-1707.
Komarova, N.Y., Thor, K., Gubler, A., Meier, S., Dietrich, D., Weichert, A., Suter Grotemeyer, M., Tegeder, M. and Rentsch, D. (2008) AtPTR1 and AtPTR5 transport dipeptides in planta.
Plant Physiol. 148: 856-869.
Kretzschmar, T., Kohlen, W., Sasse, J., Borghi, L., Schlegel, M., Bachelier, J.B., Reinhardt, D.,
Bours, R., Bouwmeester, H.J. and Martinoia, E. (2012) A petunia ABC protein controls strigolactone-dependent symbiotic signalling and branching. Nature 483: 341-344.
Krouk, G., Lacombe, B., Bielach, A., Perrine-Walker, F., Malinska, K., et al. (2010) Nitrate-regulated auxin transport by NRT1.1 defines a mechanism for nutrient sensing in plants. Dev. Cell 18: 927-937.
Kuromori, T., Miyaji, T., Yabuuchi, H., Shimizu, H., Sugimoto, E., Kamiya, A., Moriyama, Y.
and Shinozaki, K. (2010) ABC transporter AtABCG25 is involved in abscisic acid transport and responses. Proc. Natl. Acad. Sci. USA 107: 2361-2366.
Lee, S.C. and Luan, S. (2012) ABA signal transduction at the crossroad of biotic and abiotic stress responses. Plant Cell Environ. 35: 53-60
Leran, S., Varala, K., Boyer, J.C., Chiurazzi, M., Crawford, N., et al. (2014) A unified nomenclature of NITRATE TRANSPORTER 1/PEPTIDE TRANSPORTER family members in plants. Trends Plant Sci. 19: 5-9.
Li, J.Y., Fu, Y.L., Pike, S.M., Bao, J., Tian, W., et al. (2010) The Arabidopsis nitrate transporter NRT1.8 functions in nitrate removal from the xylem sap and mediates cadmium tolerance. Plant Cell 22: 1633-1646.
Lin, S.H., Kuo, H.F., Canivenc, G., Lin, C.S., Lepetit, M., et al. (2008) Mutation of the Arabidopsis NRT1.5 nitrate transporter causes defective root-to-shoot nitrate transport.
Plant Cell 20: 2514-2528.
Lumba, S., Cutler, S. and McCourt, P. (2010) Plant nuclear hormone receptors: a role for small molecules in protein-protein interactions. Annu. Rev. Cell Dev. Biol 26: 445-469.
Matsuura H, Takeishi S, Kiatoka N, Sato C, Sueda K, Masuta C, Nabeta K (2012) Transportation of de novo synthesized jasmonoyl isoleucine in tomato. Phytochem. 83:25-33
Nakajima, M., Shimada, A., Takashi, Y., Kim, Y.C., Park, S.H., et al. (2006) Identification and characterization of Arabidopsis gibberellin receptors. Plant J. 46: 880-889.
Nour-Eldin, H.H., Andersen, T.G., Burow, M., Madsen, S.R., Jorgensen, M.E., Olsen, C.E., Dreyer, I., Hedrich, R., Geiger, D. and Halkier, B.A. (2012) NRT/PTR transporters are essential for translocation of glucosinolate defence compounds to seeds. Nature 488: 531-534.
Parker, J.L. and Newstead, S. (2014) Molecular basis of nitrate uptake by the plant nitrate transporter NRT1.1. Nature 507: 68-72.
Petrasek, J. and Friml, J. (2009) Auxin transport routes in plant development. Development 136: 2675-2688.
Pimenta Lange MJ, Knop N, Lange T (2012) Stamen-derived bioactive gibberellin is essential for male flower development of Cucurbita maxima L. J. Exp. Bot. 63:2681-2691
Saito, H., Oikawa, T., Hamamoto, S., Ishimaru, Y., Kanamori-Sato, M., et al. (2015) The
jasmonate-responsive GTR1 transporter is required for gibberellin-mediated stamen
development in Arabidopsis. Nat. commun. 6: 6095
Santner, A., Calderon-Villalobos, L.I. and Estelle, M. (2009) Plant hormones are versatile chemical regulators of plant growth. Nat Chem Biol 5: 301-307.
Santner, A. and Estelle, M. (2009) Recent advances and emerging trends in plant hormone signalling. Nature 459: 1071-1078.
Seo, M. and Koshiba, T. (2011) Transport of ABA from the site of biosynthesis to the site of action. J. Plant Res. 124: 501-507.
Sun, J., Bankston, J.R., Payandeh, J., Hinds, T.R., Zagotta, W.N. and Zheng, N. (2014) Crystal structure of the plant dual-affinity nitrate transporter NRT1.1. Nature 507: 73-77.
Swarup, R. and Peret, B. (2012) AUX/LAX family of auxin influx carriers-an overview. Front.
Plant Sci. 3: 225.
Takeuchi, J., Okamoto, M., Akiyama, T., Muto, T., Yajima, S., et al. (2014) Designed abscisic acid analogs as antagonists of PYL-PP2C receptor interactions. Na.t Chem. Biol. 10: 477-482.
Tsay, Y.F., Chiu, C.C., Tsai, C.B., Ho, C.H. and Hsu, P.K. (2007) Nitrate transporters and peptide transporters. FEBS Lett. 581: 2290-2300.
Wang, Y.Y. and Tsay, Y.F. (2011) Arabidopsis nitrate transporter NRT1.9 is important in
phloem nitrate transport. Plant Cell 23: 1945-1957.
Yang, H. and Murphy, A.S. (2009) Functional expression and characterization of Arabidopsis ABCB, AUX 1 and PIN auxin transporters in Schizosaccharomyces pombe. Plant J. 59:
179-191.
Zhang, H., Zhu, H., Pan, Y., Yu, Y., Luan, S. and Li, L. (2014a) A DTX/MATE-type Transporter Facilitates Abscisic Acid Efflux and Modulates ABA Sensitivity and Drought Tolerance in Arabidopsis. Mol. Plant.
Zhang, K., Novak, O., Wei, Z., Gou, M., Zhang, X., Yu, Y., Yang, H., Cai, Y., Strnad, M. and Liu, C.J. (2014b) Arabidopsis ABCG14 protein controls the acropetal translocation of root-synthesized cytokinins. Nat. Commun. 5: 3274.
Zhou, F., Lin, Q., Zhu, L., Ren, Y., Zhou, K., et al. (2013) D14-SCF
D3-dependent degradation of
D53 regulates strigolactone signalling. Nature 504: 406-410.
Statement of ethical compliance
DNA recombination experiments
The study was approved by the RIKEN Yokohama Safety Center (approval number, P2012-002-07). All experiments were performed in accordance with the Genetic Recombinant Experiment Safety Control Regulations at RIKEN.
Conflict of Interest
There is no conflict of interest to declare.
Acknowledgements
I would like to express my deepest gratitude to Prof. Tomokazu Koshiba who offered
continuing support and constant encouragement. I would also like to thank Dr. Mitsunori Seo
and Dr. Takafumi Shimizu whose meticulous comments were an enormous help to me.
In ocu la ti on on selecti on medi a Number of cells used for inoculat ion
(µM ABA) 0.1 10
31 0 10
4EV NPF4.6 EV NPF4.6 EV NPF4.6
: NP F
: ABA . 1 Dete cti on of ABA tr ans por t ac ti vi ti es of N P F p rotei ns us ing Y 2H sys tems . AB I1 fu sed GAL 4 AD a nd P Y R 1 fu sed to GAL 4 DBD a nd N P F p rotei n were ex pr es sed in y ea st c el ls . T he y ea st c el ls wer e ul ated on selecti on media c onta ini ng ABA . Af ter i noc ul ati on, g rowth of y ea st c el ls were ob ser ved . N P F 4. 6 cDN A c loned i nto the y ea st ex pr es sion v ec tor p Y ES -DEST 52 in whi ch the GAL 1 pr omoter had b een r ep la ced wi th th e 1 pr omoter , or the empty v ec tor (E V ) wa s intr od uc ed i nto the y ea st str ai n PJ 69 -4 A c onta in ing DBD -P Y R 1 and AD -ABI 1 tr u cts . Sel ec ti on medi a (SD, - T rp , -L eu , -U ra , -H is) c onta ini ng 0 , 0. 1 or 1 µ M ABA were i noc ul ated 10
3or 10
4c el ls . P hoto s ta ken 3 d ay s after i noc ul ati on .
A B
In tr od u cti on of th e rece p tor , P P 2 C an d a NP F Ob ser va ti on y eas t cell s gr owth
: GA : JA -I le : IAA or mone r ec ep tors a nd thei r inter ac tors us ed in the p res ent stu dy . sys te m wi th the r ec ep tor c omplex es for GA . GAI fu sed GAL 4 AD a nd GI D 1a fu sed to GAL 4 DBD were c ons tr uc ted in y ea st sys te m wi th the r ec ep tor c omplex es for JA -Il e. J AZ 1 or J AZ 3 fu sed GAL 4 AD a nd COI 1 fu sed to GAL 4 DBD were c ons tr uc ted st c el ls . sys te m w ith the r ec ep to r complex es for IAA . IAA 1, I AA 3 or IAA 5 fu sed GAL 4 AD a nd T IR 1 fu sed to GAL 4 DBD wer e uc ted in y ea st c el ls .
A B C
(µM GA
1) 0.1 1 0 10
310
4(µ M GA
3) 0.1 1 0 10
310
40.002 0.1 0 (µM GA
4) 10
310
4EV NPF4.1 EV NPF4.1 EV NPF4.1
3 C on str uc ti on of a Y 2H s yste m to d etec t GA tr ans por t activ it ies of N P F p roteins . N PF 4. 1 cDN A c lon ed i nto the yea st es sion v ec tor p Y ES -DEST 52 of whi ch the GAL 1 pr omoter had b een r ep la ced wi th the ADH 1 pr omoter , or the empty v ec tor wa s intr od u ced i nto the y ea st str ai n PJ 69 -4 A c onta in ing DBD -GI D 1a a nd AD -GAI c ons tr uc ts . Sel ec ti on medi a (SD, - T rp , - , - U ra , - H is) cont ai ni ng GA
1( 0, 0 .1 or 1 µ M) ( A) , GA
3( 0, 0 .1 or 1 µ M) ( B ) or GA
4( 0, 0 .0 02 or 0 .1 µ M) ( C) were i no cul ated wi th 10
4c el ls . P hotos were ta ken 3 d ay s (A a nd B) or 2 d ay s (C ) after i noc ul ati on .
A B C
Number of cells used for inoculation
Num b e r o f ce lls u se d fo r ino cu la tio n Num b e r o f ce lls u se d fo r ino cu la tio n
(µM JA -Ile) 50 Empt y vect or
100 0 10 3
10 4 ig . 4 Cons tr uc ti on of a Y 2H sys tem to d etect JA -Il e tr ans por t ac ti vi ti es of N P F p rotei ns . he y ea st str ai n P J6 9- 4A c onta ini ng DBD -COI1 a nd AD -J AZ3 cons tr uc ts wa s tr ans for med wi th the empty yea st ex pr es sion tor pY ES -DEST 52 of w hi ch the G AL1 p romoter had b een r ep la ced w ith the ADH 1 pr omoter . Sel ec ti on media (SD, -T rp , - eu , - U ra , - H is) contai ni ng 0, 50 or 1 00 µ M J A -Il e were inoc ul ated w ith 103 or 1 0
4 c el ls. P hotos were ta ken 5 day s after ul ati on. Numb e r o f c e lls u se d fo r in o cu la tio n
ig . 5 Detec ti on of ho rmon e tr ans por t activ iti es of N PF s. he y ea st str ai n P J6 9- 4A c onta ini ng the Y 2H rec ep tor complex systems for AB A, G A or JA -Il e wa s tr ans for med wi th N P F As cl oned i nto the y ea st ex pr es sion vec tor pY ES -DEST 52 of w hi ch the G AL1 p romoter had b een r ep la ced wi th the ADH 1 omoter . Sel ec ti on media (SD, -T rp , - L eu , - U ra , - H is) contai ni ng 0.1 µ M AB A, 0 .1 µ M G A
1, 0 .1 µ M GA
3, 0 .0 02 µ M G A
4or 5 0 J A -Il e were inoc ul ated wi th 1 0
3or 1 0
4c el ls. P hotos w er e ta ken 2 day s (GA
4), 3 da ys ( AB A, G A
1a nd GA
3) ,or 5 da ys ( JA - e) a fter i noc ul ati on. T he sc al e ba r ind ic ates a br anc h lengt h cor res pond ing to 0 .1 su bsti tuti on per site.
ig . 6 ABA tr ans por t ac ti vi ti es of N P F 4. 1/AIT 3, N P F 4. 2/AIT 4, N P F 4. 5/AIT 2 and N P F 4. 6/AIT 1. he y ea st str ai n P J6 9- 4A c onta ini ng the Y 2H rec ep tor complex systems for AB A w as tr ans for med wi th N P F 4.1 /AIT 3, PF 4. 2/ A IT 4, N PF 4. 5/ A IT 2 or N PF 4. 6/ A IT1 c DN A c lon ed i nto the yeast exp res si on vec tor p Y E S -DES T5 2, of which the GAL1 omoter had b een r ep la ced wi th the ADH 1 pr omoter . Sel ec ti on media (SD, -T rp , - L eu , - U ra , - H is) contai ni ng 0.1 µ M AB A inoc ul ated wi th 1 0
5cel ls, and ph otos were ta ken 4 d ay s a fter i noc ul ati on.
Contro l
NPF4.1
NPF4.2
NPF4.5
NPF4.6
0
2
4 EVNPF4.1
AB A
0123 EVNPF4.1GA
1 0123 EVNPF4.1GA
3 0123 EVNPF4.1GA
4 00.5 EVNPF4.1
JA -Ile
p m oles/1
7 × 10 cells/
min
0
1
2
3 EVNPF4.6
AB A
012 EVNPF4.6GA
1 012 EVNPF4.6GA
3 012 EVNPF4.6GA
4 00.5 EVNPF4.6
JA -Ile
PF4.1
p m oles/1
7 × 10 cel ls/
min
7 Di rec t mea sur ements of hor mone tr ans por t activ iti es by L C -M S/M S. A B A , G A ( GA
1, GA
3and GA
4) and J A -Il e tr ans por t vi ti es (p m ol es /1 0
7c el ls/mi n for AB A a nd G A, f m ol es /1 0
7c el ls/mi n for J A -Il e) of N P F 4.6 /NRT1 .2 /AIT 1 and N P F 4.1 /AIT 3 were ned i n the pr es enc e of 1 0 µM of the potenti al su bstr ates. Amounts of substr ates ta ken up b y y ea st cel ls tr ans for m ed wi th pty v ec tor (E V) a re shown as contr ol . Mean va lues of three bi ol og ic al r ep li ca tes ar e g iv en w ith sta nd ar d dev ia ti ons . es a bov e ea ch gr ap hs a re the res ul ts of Y 2H ass ay s sh own in F ig . 5 .
p m oles /1 × 10
cel 7
ls/m
in in ls/m cel 7 /1 × 10 oles p m
p m oles /1 × 10 cel 7
ls/m in
00.5
GA
4 01 EVNPF2.5AB A
012 EVNPF4.2GA
1 012 EVNPF4.2GA
3 012 EVNPF4.2GA
4NPF1.2 NPF2.5 NPF4.2
01 EVNPF5.1AB A
00.5
GA
4 00.5
JA -Ile
0246 EVNPF5.2AB A
00.51
JA -Ile
NPF5.2 NPF5.7
01 EVNPF8.2AB A
00.5
GA
400.5
GA
4 00.5
JA -Ile F ig . 8 Di rec t m ea su rements of hormon e tr ans por t ac ti vi ti es b y LC -M S/MS. AB A, G A (GA
1, GA
3a nd G A
4) and J A -Il e tr ans por t ac ti vi ti es of N P F s were determi ned i n the pr es enc e of 1 0 µM of the poten ti al s ubs tr at es . A mou nts of su bstr at es ta ken up by yea st cel ls tr ans for med wi th the empty vec tor (E V) a re sh own as contr ol . M ea n va lues of three bi ol og ic al rep li ca tes ar e g iv en w ith stand ar d devi ati ons . P ic tur es a bov e ea ch gr ap hs a re the res ul ts of Y 2H a ss ay s sh own in F ig . 5 .
Forward primer (5’ to 3’) Reverse primer (5’ to 3’) A-GID1aGGATCCGTATGGCTGCGAGCGATGAAGTTACTGCAGTTAACATTCCGCGTTTACAAACGC -GAI GGATCCGTATGAAGAGAGATCATCATCATCATCATCAT CTGCAGCTAA TTGGTGGAGAGTTTCCAAGCCGAGGTGG D-COI1CCCGGGGATGGAGGATCCTGATATCAAGAGGTG GTCGACTCATATTGGCTCCTTCAGGACTCTAAC -JAZ3 GAATTCATGGAGAGAGATTTTCTCGGGTTG GTCGACTTAGGTTGCAGAGCTGAGAGAAGA
T ab le 1 P ri mer c omb inat ions us ed to cons tr uc t Y 2H sys tems for GA a nd J A -Il e rec ep tor complex es . U nd er li ned seq uenc es a re res tr ic ti on si tes us ed for c loni ng into y ea st vec tors .
AGI code Other name Forward primer (5’ to 3’) Reverse primer (5’ to 3’) 1 At3g16180 CACCATGGAGAACCCTCCGGATCAATGATCAACCAATTGGTCTT 2 At1g52190 CACCATGGAGAACCCTCCCAATGATGCAAGACCTATTGTTCCTT 3 At5g11570 aaaaagcaggctATGGATCAAGAAGCTCTTCTCagaaagctgggtCACTTGTTATATTTATATACTACATTTACC 1 At3g45720 aaaaagcaggctATGGCTGGTTTAGTATTATCTAATGagaaagctgggtGTTGTGTCTTAGTTTGTAACATC 2 At3g45690 aaaaagcaggctATGGCTCGTTCAGTCGACAagaaagctgggtGTGTCTTAGGTTGTATCGTC 3 At3g45680 CACCATGGCTAGTTTAGTCTCTGGTTACATTGTACCATCTTTTG 4 At3g45700 CACCATGGCTAATTCAGACTCTGGACAGAACACGTTAGTGTCCT 5 At3g45710 CACCATGGCTGATTCAAAATCTGGCTAGGTTTTAACATCTTTAG 6 At3g45660 CACCATGTCTGGTTCAGTTTCCCGTTAGTGAGAGACATTTGCAT 7 At3g45650NAXT1 CACCATGGCTAGTTCAGTTACTGGTCAGTGAGAGACATTTGCAG 8 At5g28470NRT1.9 CACCATGGACGTCGAATCTTCATCATCTAAACATCTTTAGTGTC 9 At1g18880 CACCATGGAGGTTGAGAAGACAGATTACACTGACACCTTATCAA 10At3g47960GTR1 CACCATGAAGAGCAGAGTCATTCTTCAGACAGAGTTCTTGTCTT 11At5g62680GTR2/NRT1.10aaaaagcaggctATGGAGAGAAAGCCTCTTGAACagaaagctgggtTCAGGCAACGTTCTTGTCTTG 12At1g27080NRT1.6 CACCATGGGAGTTGTTGAGAATCGCTAACTTGGAGATTGATCGT 13At1g69870NRT1.7 CACCATGGTTTTGGAGGATAGAAATCATTTCATCGATTTCTTCG 14At1g69860 aaaaagcaggctATGGACAATGAGAAAGGGACAAagaaagctgggtGTT TGACAT GTTGGAAATGAGG 1 At1g68570NitraaaaagcaggctATGGAGGAGCAAAGCAAGAACAAagaaagctgggtCAACGCCAAGACTCATTCATC
le 2 P ri mer c omb inat ions us ed to a mp li fy N P F c DN As (N P F 1, 2, 3) er li ned seq uenc es a re ov er hang s for i ntr od uc ti on into pE N T R/D -T OP O. leoti des shown i n l owerc ase l et ters a re part of a ttB 1 (f or fo rw ar d pr imers ) and a ttB2 (f or r ev er se p ri mers ) r ec omb ina ti on seq uenc es . uenc es hi ghl ig hted b y dotted und er li nes i n the rev er se pr imers a re 3 ’ untr ans la ted r eg ions of cDN As .
AGI code Other name Forward primer (5’ to 3’) Reverse primer (5’ to 3’) 1 At3g25260AIT3 N.A. N.A. 2 At3g25280AIT4 N.A. N.A. 3 At1g59740NRT1.14 N.A. N.A. 4 At1g33440NRT1.13 N.A. N.A. 5 At1g27040AIT2 N.A. N.A. 6 At1g69850NRT1.2/AIT1 N.A. N.A. 7 At5g62730 N.A. N.A. 1 At2g40460 CACCATGGAGGCTGCAAAAGTTTACACACAAGTTAGATACTAAGAGGAGATGTGTCTAAGGC 2 At5g46050PTR3 CACCATGACAGTAGAAGAGGTAGGAGACGTTATTCAGTCTCTTTCATTTCCACCTCTTTCAC 3 At5g46040 CACCATGACAGTAGAAGAGGTAGGAGATGATTATACTTACTCATTGTAGTTATCTACCATATTTGGTTCTTTCTG 4 At3g54450 aaaaagcaggctATGGCAGATTCGACCTCCCagaaagctgggtTGATCAAACTTATCTTTCGCTTTCAAC 5 At2g38100 CACCATGTCAGTTCTGTCATGGGCCTACACAGAAGCGATTATCC 6 At2g37900 CACCATGGAGCATAAGAATATTGGTGCAGAAGTCAGGAGAGCCCTTGGCTTG 7 At3g53960 CACCATGGAGCACAACAAGGTTGATACAGAACTTACGTAAACTTGGACGTGTTATTCGTCC 8 At5g14940 CACCATGGCTGGAGGAGAGAAAAGAAGTTAACAATTTGTATCACATTGATCATCATCATCAC 9 At3g01350 CACCATGGATTTAGAACAGAAGACAAGAGGACTCATTTAGGGGCTTCTTCTTTTTCATCG 10At1g22540 CACCATGTCGATCTCCGGCGCTGTTAACTGGTGTCGAGCCTTTTAGAGAC 11At1g72130 CACCATGGCTATCACCTACTCCTCCGTTAAAAGGTGTTTGATCTGCTGTAGAC 12At1g72140 CACCATGTCGACATCCATCGGCGATAATGCTACTTTGGGCTGTTGTAGAGATAGGA 13At1g72125NRT1.16aaaaagcaggctATGACGACGACTTCCAAAACTTCagaaagctgggtCTACACTACGTCCACCCGG 14At1g72120NRT1.15CACCATGACGACTACTTCAGAAATTTCTCTTCAAGAAGCTACACTCGATCCACTCGACG 15At1g22570 aaaaagcaggctATGAAGATACCAGAGGAAGAAGagaaagctgggtTTAGACTTGGTCTAGCCGAC 16At1g22550 CACCATGGCGATAGCCGAAGAAGAAGCTTAGACTTGATCTACACGGCGATACAC
le 2 (c onti nued ) P ri m er comb inat ions us ed to a m pl ify N P F c DN As (N P F 4, 5) er li ned seq uenc es a re ov er hang s for i ntr od uc ti on into pE N T R/D -T OP O. leoti des shown i n l owerc ase l et ters a re part of a ttB 1 (f or fo rw ar d pr im er s) a nd a ttB2 (f or r ev er se p ri m er s) re com bi na ti on seq uenc es . not am pl ified in thi s stu dy .
le 2 (c onti nued ) P ri mer c omb inat ions us ed to a mp li fy N P F c DN As (NP F 6, 7, 8) er li ne d se quenc es are over hangs f or i ntr od uc ti on i nto p E N TR/D -TOP O. leoti des shown i n l owerc ase l et ters a re part of a ttB 1 (f or fo rw ar d pr imers ) and a ttB2 (f or r ev er se p ri mers ) r ec omb ina ti on seq uenc es . not amp li fied in thi s stu dy .
AGI code Other name Forward primer (5’ to 3’) Reverse primer (5’ to 3’) 1 At1g13400 N.A. N.A. 2 At2g26690NRT1.4 N.A. N.A. 3 At1g12110NRT1.1 N.A. N.A. 4 At3g21670NRT1.3 N.A. N.A. 1 At5g19640 CACCATGGCCGCTATGGATCCGTCAAACTTGTTCCAACTCAGAAACGG 2 At4g21680 CACCATGGATCAAAAAGTTAGACAGTTTGAGGTCAGACTTCCTCCTCTTCAGTTAC 3 At1g32450 aaaaagcaggctATGTCTTGCCTAGAGATTTATAAagaaagctgggtTTAGACTTTAGAATCCTTCTCT 1 At3g54140PTR1 CACCATGGAAGAAAAAGATGTGTATACGCTCAATGTGCTCGACCAACAG 2 At5g01180 PTR5 CACCATGGAAGATGACAAGGATATATACACAAAAGTCAAAGCGCATGCCCGG 3 At2g02040PTR2 CACCATGGGTTCCATCGAAGAAGAAGCTACGACGAAGCTTTCTTTTGC 4 At2g02020PTR4 aaaaagcaggctATGGCTTCCATTGATGAAGAAAGagaaagctgggtCCAGAAGAAGAGATCAAACCTT 5 At1g62200PTR6 aaaaagcaggctATGGTGAATTCGAATGAAGAAGAagaaagctgggtTTACAAAGCCTTCTTCTTTGTGTHormoneSolvent ASolvent BGradient (solvent B composition) ABA0.01 % acetic acidAcetonitrile 0.05% acetic acid3% over 0.3 min 3-40% over 6.7 min GA1, GA3, GA40.01 % acetic acidAcetonitrile 0.05% acetic acid
3% over 0.3 min 3-20% over 4.7 min 20-40% over 5 min 40% over 2 min JA-Ile 0.01 % acetic acidAcetonitrile 0.05% acetic acid
3% over 0.3 min 3-15% over 0.7 min 15% over 2 min 15-40% over 4 min 40-60% over 1 min
T ab le 3 Cond iti ons for L C
TargetRetention time on LC (min)
ESIMS/MStransitions forquantification (m/z)
Collisionenergy (V)
Declustering potential
Ionspray voltaging (V)
Desolvation temperature (℃) ABA 5.1−263.1/219.1 15903,500600 d6-ABA−269.1/225.2 GA1 4.5−347.2/273.1 40904,000600 d2- GA1−349.2/275.1 GA3 4.4−345.2/221.1 40904,000600 d2- GA3−347.2/223.1 GA4 9.1−331.2/257.1 30904,000600 d2- GA4−333.2/259.1 (-)-JA-Ile 7.5−322.2/130.1 30903,500600 13C6-(-)-JA-Ile −328.2/136.1 (+)-JA-Ile 7.3−322.2/130.1 30903,500600 13C6-(+)-JA-Ile −328.2/136.1