Cry1A
殺虫毒素
っ
誘導さ
2
種類の細胞死の機構
その役割の解析
Mechanisms and roles of two types of cell death induced by Cry1A insecticidal toxin
2014.3
東京農工大学大学院
生物システ
応用科学府
生物システ
応用科学専攻
目次
序論...9
第1章 イコ 消化管上皮細胞 起こ 濃度依存的 細胞死 い の解析
第1節 目的………..25
第2節 材料 方法………..13
1.2.1 供試昆虫
1.2.2 Cry1Aa毒素の活性化 精製
1.2.3 イコ 幼虫を用いたバイ アッセイ
1.2.4 イコ 幼虫組織切片の作製
1.2.5 TUNEL 染色
1.2.6 イコ 幼虫中腸のcDNA作製 ア タイ PCR
第3節 結果………..17
1.3.1 異 濃度のCry1Aa投与濃度 イコ 3齢幼虫の病状 の関係の継時的
観察
1.3.2 Cry1Aa毒素投与後の中腸上皮細胞の経時的観察
1.3.3 Cry1Aa処理 イコ 幼虫中腸 のアポトーシス関連遺伝子発現 ベ の比
較
第4節 考察………..21
1.4.2 消化管免疫誘導の分子機構
第2章 イコ ABC transporter C2のCry毒素殺虫作用機構上 の役割の解析
第1節 目的………..34
第2節 材料 方法………..36 2.2.1 昆虫培養細胞
2.2.2 cDNAのク ーニン 2.2.3 組換え型AcNPVの作製 2.2.4 Cry毒素の精製
2.2.5 LDHアッセイ
2.2.6 細胞傷害率の算出
2.2.7 抗血清の作製
2.2.8 BmABCC2変異体発現細胞表面上の毒素の染色
2.2.9 ウエスタン ッテ ン
2.2.10 免疫染色
2.2.11 BtR175発現細胞表面上の毒素の検出
2.2.12 SPRを用いたBtR175 Cry1A毒素の結合性状の解析 2.2.13 膜画分の調整
2.3.3 BtR175がSf9 与え 感受性付与能力のBmABCC2 の比較
2.3.4 コ ABCC2のCry毒素受容体 し の機能の確認
2.3.5 AaABCC2のCry4Aa受容体 し の機能の有無の調査 2.3.6 BmABCC2のCry9受容体 し の機能の有無の調査
第3章 ABCC2 BtR175の協調作用 い の解析
第1節 目的………..80
第2節 材料 方法………..82 3.2.1 昆虫培養細胞
3.2.1 Cry毒素の精製
3.2.2 イコ 幼虫を用いたバイ アッセイ
3.2.3 細胞傷害率の算出
3.2.4 SPRを用いたBtR175 Cry毒素の結合性状の解析
3.2.5 T7 ーターへのcDNAの連結
3.2.6 cRNAの合成 3.2.7 電気生理学的解析
3.2.8 ウエスタン ッテ ン
3.2.9 APN1発現Sf9細胞の免疫染色
3.2.10 受容体発現アフ ツ エ 卵母細胞の免疫染色
第3節 結果 考察………...87
い感受性
3.3.2 BmABCC2 BtR175の協調作用のCry毒素の活性発現 おけ 意味
3.3.3 BmABCC2 BtR175の2分子間 み た協調作用のCry毒素孔形成過程
与え 影響
第4章 BmABCC2 Cry毒素の結合動態及び相互作用領域の解析
第1節 目的……….112
第2節 材料 方法……….114 4.2.1 昆虫培養細胞
4.2.2 Cry毒素の精製
4.2.3 Cry毒素の細胞膜上 の検出
4.2.4 Flag-tagのcDNA配列C末端への連結 4.2.5 BmABCC2変異体cDNAの作製 4.2.6 組換え型AcNPVの作製
4.2.7 膜画分の調整 4.2.8 細胞傷害アッセイ 4.2.9 銀染色
4.2.10 ウエスタン ッテ ン
4.2.11 SPRを用いたBtR175及びBmABCC2 Cry1A毒素の結合性状の解析
4.3.2 細胞外 ー 領域2近傍がCry毒素 の結合領域 あ 可能性の検証
4.3.3 細胞外 ー 領域2 挿入さ ア ノ酸の種類の影響
4.3.4 BmABCC2細胞外 ー 変異体のCry毒素感受性 与え 影響
総合考察………141
謝辞………146
1 序論
陽性好気性芽胞形成細菌Bacillus thuringiensis 、1901年 石渡繁胤が イコ (Bombyx mori) の卒倒病菌 (sotto disease bacillus) し 報告した昆虫病原細菌 あ (石渡 1901) し し、石渡 その後も英文雑 B. thuringiensisを新種の微生物
し 記載し ったた 、1911年 シノ マ イ 同菌を同定したBerliner
っ 卒倒病菌 、B. thuringiensis さ 現在 (Berliner et al. 1915) その
後、B. thuringiensisの殺虫活性の本体が、芽胞形成時 作 結晶性タン ク質 (以
Cry毒素) あ こ が、Angus っ 発見さ た (Augus et al. 1954) 現在、Cry
毒素の遺伝子配列を Cry1 – Cry72 分類さ 400種類以上のCry毒素が同定さ
い http://www.lifesci.sussex.ac.uk/home/Neil_Crickmore/Bt/ B. thuringiesnsis 、 1菌株が1種類のCry毒素を生産す 限 、例え Bacullus thuringiensis HD-1 株
、Cry1Aa、Cry1Ab、Cry1Acの3種類の毒素を生産す いった う 、複数種類
の毒素を生産す こ 高い殺虫活性を有す う 昆虫 の間 共進化し きた
考え Cry毒素 、前駆体Cry毒素の分子量を 大きく130 kDa型 70 kDa
型の2種類 分類さ 130 kDa型 、 ョウ目 活性のあ Cry1A毒素、 エ目
活性があ Cry4A、Cry4B毒素、コウ ュウ目 活性を示すCry8C毒素、そし セン
ュウ目 活性があ Cry5毒素 があ こ 130 kDa型毒素 、昆虫の消化液
含ま テアー 限定分解さ 、N末端のア ノ酸20残 程度及びC末
端の約半分を失い、65 kDa前後の活性型毒素 殺虫活性を示す(Aronson et al. 1995)
一方 、70 kDa型 、 ョウ目、 エ目の両者 活性を示すCry2毒素、 エ目 活
性を示すCry11A、Cry11B毒素、コウ ュウ目 活性を示すCry3B毒素 があ 70
kDa型も テアー 限定分解を受け、N末端のア ノ酸数残 を失い、65 kDa
2
この う 、Cry毒素 そ が異 昆虫 対し 活性を示すわけ あ が、
こ 殺虫活性を持 Cry毒素の多く 、5箇所の高度 保存さ たア ノ酸配列領域
( ック)を持 こ が分 っ い (Hofte et al. 1989) この保存 ック 、Cry毒素
の殺虫活性 おい 何 重要 役割を持 考え い も 、B. thuringiensis
が産生す Cry毒素が共通の祖先 生ま 、進化し きたこ を示す証拠 もあ
1次構造 おけ 相同性の高さ 加え、Cry毒素 、3次構造 い も相同性が高
いこ が分 っ い 現在ま 3次構造の結晶構造解析が終わっ い 毒素 、
Cry3Aa (Li et al. 1991)、Cry1Aa (Grochulski et al. 1995)、Cry1Ac (Derbyshire et al. 2001)、 Cry2Aa (Morse et al. 2001)、Cry3Bb (Galitsky et al. 2001)、Cry4Ba (Boonserm et al. 2005)、 Cry4Aa (Boonserm et al. 2006)、そし Cry8E (Guo et al. 2009) の8種類 あ 全 の毒
素が インI インIIIの3 の イン 成 、 本的 3次元構造 、
全 のCry毒素 共通し い インI 、N末端部分 相当し、7個のα- ッ
クスを含 、こ のα- ックスが細胞膜 貫通し、 孔を形成す 考え い
インII インIII β-シートを含 構造 成 、宿主昆虫の消化管上皮
細胞上の分子 結合す 役割を持 考え い
Cry毒素 、限 た範囲の昆虫 特異的 活性を示し (van Frankenhuyzen 2009)、哺
乳類 無害 あ た 、1960年代 、人畜 無害 殺虫剤 (BT剤) し 米国
実際の農業現場 おい 使用さ 始 た さ 、1990年代以降、分子生物学の発達
共 遺伝子組換え作物の作出が可能 、Cry毒素の遺伝子を導入した耐虫性遺伝
子組換え作物 (B. thuringiensis Gene modified Crop; 以 Bt crop) が米国を中心 トウ
コシ、トマト、 イ 多くの作物 適応さ う った(Roh et al. 2007)
Bt cropの世界 おけ 作付面積 、1996年 、110万ha あったの 対し、2010年
、6,600万ha 急増し い (Tabashnik et al. 2013) 米国 おい 2012年度 生産
3
(http://www.ers.usda.gov/data-products/adoption-of-genetically-engineered-crops-in-the-us.asp
x)、綿 い 、米国、中国、イン 、 ースト ア 生産さ た綿の79 - 95%が
Bt crop あ こ が報告さ い (Tabashnik et al. 2013) したがっ 、Cry毒素 世
界 最も広く農業利用さ きた毒素タン ク質 あ 言えそう あ
この う Cry毒素を用いた生物的防除が1990年以降、急速 及した背 、
第二次世界大戦以後の化学合成農薬の開発 、そ も 人々の間 浸透した化学合
成農薬 対す 社会的危機感 あ Cry毒素 、タン ク質 あ がゆえ 土壌中
分解さ やすく、また紫外線 容易 失活す た 、化学合成農薬 おい 懸念さ
い 環境汚染や人体 有害 物質が作物 残留す 心配が回避 き また、自然
界 おい 何億年もの間、昆虫 共 共進化し きたCry毒素 対す 抵抗性 発達し
くい 考え 、化学農薬の使用 おい 問題視さ いた抵抗性害虫の問題も克服
き 期待さ たこ もその理由の1 あ し し、実際 、1985年 実験
室内 飼育さ いたシノ マ イ (Plodia interpunctella ) 初 のCry毒素
(Bt剤) 抵抗性害虫が認 た (McGaughey. 1985) その後も、コ (Plutella xylostella) おい Bt剤抵抗性が発達したこ が報告さ た (Tabashnik et al. 1999;
Janmaat and Myers. 2003) さ 、2007年 アフ 初 のBt Crop 対す 抵
抗性を発達させた (Busseola fusca) が報告さ 以来 (van 2007)、P. xylostella (Tabashnik et al. 2010)や、 トウ (Spodoptera frugiperda) (Storer et al. 2010)、そし 、
コウ ュウ目 あ 根切 シ (Diabrotica virgifera virgifera) おい 次々 抵 抗性の発達が報告さ い (Gassmann et al. 2011) この う Cry毒素抵抗性 (Bt剤や
Bt crop抵抗性を指す) の発達 、今後Cry毒素を用いた生物的防除手段の効果を高い
ベ 維持し、長期的 使用し いく上 解決す き重要 課題 言え
この う 、既 60年以上もの間生物的防除手段 し 利用さ きたCry毒素
4
の殺虫活性の本体がCry毒素 あ こ が明 った1956年のAugus の報告以
降、約60年もの間多くの研究者が、Cry毒素が昆虫を殺す 組みを明 し う
精力的 研究を行っ きた その成果 Cry毒素作用機構の大部分 解明さ た
う 一般的 考え え い が、現在の作用機構仮説 説明 き い現象が多く存
在す す わ 、 ン様タン ク質を異所発現させた細胞の感受性 必 しも
個体死を説明 き ほ の高さ い (Nagamatsu et al. 1999) また、APNを異所発
現させ もその細胞 毒素 対す 感受性が付与 き い の報告があ (Luo et al.
1999) さ 、APN遺伝子の破壊 依存す 抵抗性系統の発生の例 知 い
い さ また、Cry毒素変異体 (Cry1AbMod、Cry1AcMod) 、 ン様タン
ク質 変異があ 害虫を殺すこ が き こ が報告さ い (Soberon et al. 2007)
こ のこ 、現在語 い 作用機構仮説 問題があ こ を示唆し い
考え
感受性昆虫体内 込ま たCry毒素 、ま 消化液中の テアー 限定
分解さ こ 、活性型の毒素 (Aronson et al. 1995) 現在提唱さ い 消化
管上皮細胞 ベ の作用機構仮説 “Pore formation model” (Bravo et al. 2004) や “Signal transduction model ” (Zhang et al. 2005; Zhang et al. 2006) があ 前者 、
Cry毒素が感受性昆虫体内 殺虫活性を示すの 複数の過程が必要 さ きた
昆虫消化管内 活性型 った毒素 、 ン様タン ク質 の相互作用を経
マー化し (Gomez et al. 2002; Jimenez et al. 2007)、更 他の細胞膜上の分子への結合
を介し 消化管上皮細胞の 筒細胞の細胞膜 挿入さ 、poreを形成す 中腸上皮細
胞 、Cry毒素 形成さ た 孔 大量 流入したイ ン 浸透 の上
、細胞が膨潤、破裂し、結果 し 昆虫を死 し さ い (Knowles
1987) 一方、後者の仮説 、活性型 った毒素がGタン ク質の活性化を介し
5
の 細胞死を細胞 誘発す さ し し、この仮説 培養細胞 おい
のみ適用 き こ が示さ たもの あ 、昆虫の消化管 も同 こ が起こ 否
い 検討さ い い
Cry毒素 っ 誘導さ 現象 、毒素が宿主昆虫を の う 機構 殺す い
う殺虫作用のみ フ ー スを当 た研究がほ あ し し、昆虫が死
ま 細胞 起こ 現象 、毒素 っ 物理的 引き起こさ 現象 け くそ
対抗し う す 宿主昆虫の免疫応答 も同時 起こ こ が考え 実際
、いく の研究 おい その う 昆虫のCry毒素 対す 免疫応答 も考え
え 反応 着目し、研究成果を報告し い ア イエ (Culex pipeiens)の幼虫 おい
、Cry毒素を摂 す こ っ 中腸上皮細胞 アポトーシスが誘導さ 、その
こ が昆虫の個体死の原因 っ い こ が示唆さ い (Smouse and Nishiura
1997) アポトーシス 、生物種間 高度 保存さ た 細胞死の1 あ
、DNAの断片化、ク マ ンの凝集、 ス ー の活性化 いった特徴 規定
さ (Kerr et al. 1972) 多様 動物種 おい 発生の過程 おけ 細胞数の制御の役
割を担い (Driscoll and Chalfie. 1992; Champlin and Truman. 1998; Mckay. 1997)、また、傷
害を受けた細胞 誘導さ アポトーシスを引き金 、幹細胞が分化、増殖す こ
細胞数を維持す こ 役立っ い の報告もあ (Jiang et al. 2009)
また、タバコス (Manduca sexta) ネッタイシマ (Aedes aegypti) Cry1Ab、 Cry11Aaを処理しRNA干渉 (RNA interference、以 RNAi) を用いた研究成果 も
MAPK p38 経路の活性化が、Cry毒素を摂食した昆虫のCry毒素 対す 抵抗性 重要
あ 報告した (Cancino-Rodezno et al.2010) さ 、近年、Bedoya-Pérez 、 A. aegypti おい RNAiを行った実験結果 unfolded protein response (UPR) 経路が Cry11Aa 対す A. aegyptiの免疫 し 機能し い こ を示した (Bedoya-Perez et al.
6
同時 そこ 回避し う 反応し い 期待さ た そこ 、第1章 、 イコ
幼虫消化管上皮細胞 起こ 、致死量の毒素を与えた際 細胞が短時間 死 いう
現象 け く、毒素 傷害 宿主側のその傷害 対抗し う す 反応の全
をみ こ 、昆虫消化管細胞 実際 起こっ い Cry毒素 っ 誘導さ 現象
を正しく理解す こ を目指した
前述した通 、Cry毒素殺虫作用機構仮説 “Pore formation model” (Bravo et al. 2004) “Signal transduction model ” (Zhang et al. 2005; Zhang, et al. 2006) が有力 あ 考え
い し し、 が正しい仮説 あ の 、また、全 の毒素 おい この
作用機構仮説が当 ま の 関し 未解明 あ し し が 、こ の仮説
いたCry毒素作用機構上 、Cry毒素 、宿主昆虫の消化管上皮細胞上 発現
す 受容体分子 相互作用す こ が必要 あ 、この受容体分子を明 す こ
、Cry毒素殺虫作用機構の全貌を知 うえ 非常 重要 あ 言え 本研究 主
用い い Cry1A毒素 い 、特 この受容体 い の研究が進 お 、
必 しも機能の証明が いものも含 いく の受容体、あ い 受容体候補 称さ
き分子がいく 報告さ い ン様タン ク質 、B. mori
(Nagamatsu et al. 1999)、M. sexta (Hua et al. 2004) 、ニセア タバコ (Heliothis virescens ) (Jurat-Fuentes et al. 2006)、 ー ッ アワノ イ (Ostrinia nubilalis)
(Flannagan et al. 2005) おい Cry1A毒素 対す 感受性を培養細胞 付与す こ の
き 分子 あ こ が示さ い また、H. virescens (Gahan et al. 2001) 、 タ
バコ (Helicoverpa armigera) (Xu et al. 2005)、ワタア シ (Pectinophora
gossypiella) (Morin et al. 2003)のCry毒素抵抗性系統 関す 抵抗性遺伝子の遺伝学的解
析 、 ン分子が正しく発現 き く こ が、抵抗性の1 の原因 あ
こ が示さ い 一方 、 コシ スフ イノシトー
7
ー N (Amino peptidase N、以 APN)、特 APN Class 1 (以 APN1) 、ショウ ョウ バエ (Drosophila melanogaster) 発現させ こ 、Cry1Ac 対す 感受性が生ま た の報告や (Gill et al. 2002)、APN1のサイ ンシン がCry毒素 対す 幼虫個体の 感受性を させ の報告があ (Yang et al. 2010; Tiewsiri et al. 2011) また、Cry1Ac
抵抗性のH. armigera おい APN1分子 変異が入っ いた の報告があ (Zhang et
al. 2009) し し、 ン様タン ク質の う 遺伝学的 APN1がCry毒素抵抗
性原因因子 同定さ た例 く、未 APN1の変異や発現量の減少 抵抗性の因果
関係 明瞭 部分が多く残っ い 一方、APN 同様のGPIアン ー型タン
ク質 あ ア フ スフ ター (Alkaline phosphatase、以 ALP) い 、Cry
毒素抵抗性昆虫 おい ALP 対す 結合性の が抵抗性 相関関係があ こ が
報告さ い が、異所発現実験 Cry毒素 対す 感受性を付与 き す
報告 み い (Jurat-Fuentes et al. 2004; Jurat-Fuentes et al. 2011) 2010年、Gahan
今ま Cry毒素殺虫作用機構上 一度も姿を現したこ の いABC ト ンスポ
ーターC2 (ATP-binding cassette transporters subfamilyC member2、以 ABCC2) が、 ン
ー マッ ン の結果 きCry1Ac抵抗性H. virescensの抵抗性原因因子 あ
し 報告さ た(Gahan et al. 2010) また、2012年 、 イコ おい もCry1Ab
抵抗性 イコ 系統の ン ー マッ ン 解析が行わ た結果 、ABCC2分子の
変異がCry1Ab抵抗性の原因 あ し 報告さ た (Atsumi et al. 2012) さ 、
ン様タン ク質やAPN 変異が認 ったCry1Ac抵抗性のP. xylostella
やイ クサ ンウワバ (Trichoplusia ni) おい ABCC2分子内 変異があ こ が新
た 見 (Baxter et al. 2011)、ABCC2分子がCry毒素殺虫作用機構上 重要 働き
をす 分子 あ こ が確定的 っ きた
そこ 、第2章 、ABCC2分子がCry毒素殺虫作用機構上 重要 働きをす 分
8
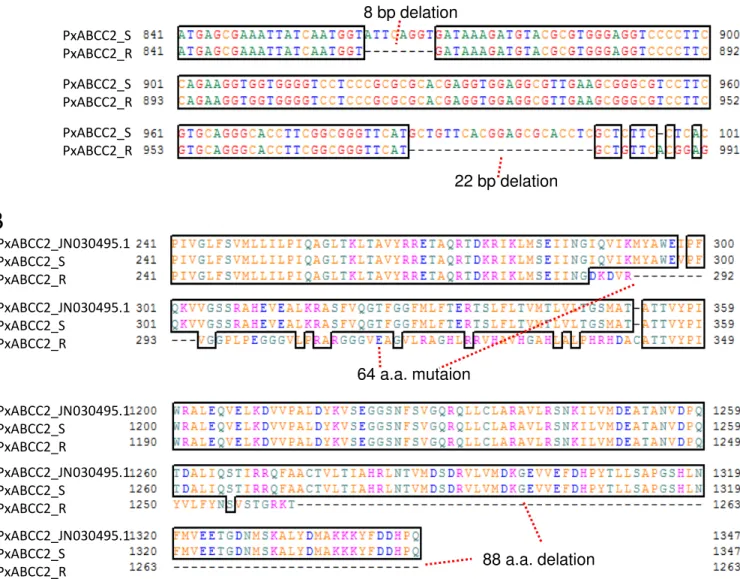
コ のABCC2 (BmABCC2) 及び ン様タン ク質 (BtR175) のcDNA配列を組
込 組換え型Autographa californica nuclear polyhedrosis virus (AcNPN)を用い 、Sf9
細胞膜上 受容体分子を発現させ、Cry1A毒素 対す 感受性をin vitroのバイ アッ
セイ っ 評価した また、 ョウ目 あ P. xylostella、 エ目 あ ヒトス シマ
(Aedes albopictus)のABCC2 い も受容体 し 機能す を調 こ 、Cry
毒素受容体 し ABCC2が機能し い 昆虫の範囲、及び、様々 Cry毒素の中の
の範囲ま がABCC2を利用 き の 、 い 考察した また、このSf9細胞を用
いたバイ アッセイの結果を BmABCC2の受容体 し の機能 (あ い 細胞
感受性を付与す 機能) をBtR175 比較す こ も試みた
前述した有力 作用機構仮説の1 あ “Pore formation model” (Bravo et al. 2004)
おい 、Cry毒素が細胞毒性を発揮す の ン様タン ク質 ア ノ
ー (APN) いう異 2種類の受容体 毒素の異 部位 結合す こ が重
要 さ い (Bravo et al. 2004) また、H. virescens おい APN 同様のGPI
アン ー型のタン ク質 あ ALPが ン様タン ク質 結合し マー
化した毒素が第2の受容体 し 結合す こ Cry毒素の細胞膜への挿入が可能
の報告もあ (Jurat-Fuentes et al. 2006) さ 近年、Cry毒素がAPN 結合した
後 ン様タン ク質 結合し再びAPN 結合す いう”Ping pong binding
model ”も提唱さ い (Pacheco et al. 2009) 少 く も、 ョウ目昆虫の消化管の細
胞上 数種類の結合タン ク質が存在す の 事実 あ 、第2章 調 た う 単独
細胞上 発現す 系を調 た け Cry毒素の真の作用の工程を正しく理解し
い い可能性があ
そこ 第3章 、BmABCC2 BtR175の2分子を同一の昆虫培養細胞 発現させ
こ 、先行研究 主張さ い う 、 続きの作用工程 両分子が働き、
9
い 検討した また、アフ ツ エ 卵母細胞 両分子を発現させ、形成さ
Poreを電気生理学的手法 定量的 解析す こ 、本研究 Cry毒素が作
用す 過程 相互作用す 考え た2分子の受容体 、 の う し Cry毒
素の活性が高 の い も考察した
一方、第4章 、Cry毒素の受容体 し 機能す こ が分 ったBmABCC2が受
容体 呼ぶ き分子 あ も関わ 、Cry毒素 結合す 証拠が見 っ い い
点 着目し、BmABCC2 Cry毒素 の結合解離の動態を え こ を試みた その
結果、確 Cry毒素 BmABCC2が結合し い こ を示す証拠を得たた 、更
その結合動態をAPN1やBtR175 比較す こ 、BmABCC2 Cry毒素の結合動態
の う 特徴があ を考察した さ 、BmABCC2分子の の領域がCry毒素
10
第1章 イコ 消化管上皮細胞 起こ 毒素濃度依存的 細胞死 い の解析
第1節 目的
諸言 も既 述 た通 、感受性昆虫体内 込ま たCry毒素 、ま 消化液中
の テアー 限定分解さ こ 、活性型の毒素 (Aronson et al. 1995)
現 在 提 唱 さ い 消 化 管 上 皮 細 胞 へ の 作 用 機 構 仮 説 、“Pore formation model” (Bravo et al. 2004) や“Signal transduction model ” (Zhang et al. 2005; Zhang, et al. 2006)
があ 前者 、Cry毒素が感受性昆虫体内 殺虫活性を示すの 複数の過程が
必要 さ きた 昆虫消化管内 活性型 った毒素 、 ン様タン ク質
の相互作用を経 マー化し (Gomez et al. 2002; Jimenez-Juarez et al. 2007)、更
他の細胞膜上の分子への結合を介し 消化管上皮細胞の 筒細胞の細胞膜 挿入さ 、
poreを形成す 中腸上皮細胞 、Cry毒素 形成さ たpore 大量 流入した
イ ン そ 伴う水の流入 浸透 の上 、細胞が膨潤、破裂し、結果
し 昆虫を死 し さ い (Knowles 1987) 一方、後者の仮説 、活性
型 った毒素がGタン ク質の活性化を介し PKAを活性化す こ 、oncosis様
の 細胞死を細胞 誘発す さ この う Cry毒素の殺虫作用機構
い 、現在 も、全貌が解明さ お 、議論の余地が残さ い
こ 、実際の宿主個体 、毒素の作用 宿主の反応の総和 っ 宿主個体や
組織 変化を示す 考え す わ 、毒素 対す 宿主個体の感受性 例え 免
疫応答 っ 大き 影響を受け こ が予想さ し、そ こ 、免疫反応が組
織の崩壊を助長す 可能性さえも考え Cry毒素が属す Pore-forming toxin (以
PFT) 対す 宿主の細胞の応答 い 哺乳類 おい も多くの研究が行わ
い PFT 、Strephylococcus aureus, Streptococcus pygenes, Clostridium perfringens
11
あ (Huffman et al. 2004; Bischof et al. 2008) 例え 、CCL-185、CCL-138、HEK い う3種類のヒトの上皮培養細胞 Streptococus pneumoniaeの産生す Pneumolysin及び、 Bacillus anthracisの産生す AnthorolysinOを添加す mitogen-activated protein kinase
p38 (以 MAPK p38) 経路 いう免疫応答 関連のあ シ 伝達経路が活性化さ
た 報告し い (Ratner et al. 2006)
セン ュウや昆虫 おい 、Cry毒素 対す 中腸上皮細胞の宿主の免疫応答の解明
を目的 研究が始まっ い Cry5Bを摂食したC. elegansの研究 、マイク ア
イ 解析 Mitogen activated protein kinase (以 MAPK) p38 経路 c-Jun N-terminal kinase (以 JNK) 経路 いう 種類のMAPK経路が活性化した 結論付け
い さ 、そ の経路の活性化 関与す 因子をRNA interference (以 RNAi)
ノック ウンす 実験が行わ 、こ 2種類のMAPK経路がC. elegansの消化
管上皮細胞を Cry5B 与え 傷害 保護す こ 貢献し い 報告さ
た(Huffman et al. 2004; Bischof et al. 2008) 最近行わ たBravo のM. sexta Aedes aegypti Cry1Ab、Cry11Aaを処理しRNAiを用いた研究成果 もMAPK p38 経路の
活性化 Cry 毒素を摂食した感受性昆虫が生存す た 重要 あ 報告さ い
(Cancino-Rodezno et al.2010)
一方、いく のCry毒素を用いた研究 おい 昆虫の消化管細胞 アポトーシス
が誘導さ こ が報告さ い Culex pipeiensの幼虫を用いた実験 、Cry毒素
を摂 す こ っ 中腸上皮細胞 アポトーシスが誘導さ 、そのこ が昆虫の個
体死の原因 っ い こ を示唆し い (Smouse and Nishiura 1997) また、Heliothis
virescens の中腸の培養細胞を用いた実験系 、Cry毒素 っ 中腸上皮細胞のアポ
トーシスが誘導さ こ が示さ た (Loeb et al. 2000) 同 ー の後の研究成果
、Cry毒素を添加す こ 、アポトーシス 共 、幹細胞の分化が 進さ た
12
っ 傷害を受けた 、死 しまった細胞を更新す 意味を持 の い 述
い (Loeb et al. 2001)
ここま 述 きた毒素の作用機構も、宿主側の生体防御反応 関し も多く 培
養 細 胞 を す 様 々 in vitro の 実 験 系 を 利 用 し 示 さ き た も の あ
(Zhang et al. 2005; Zhang, et al. 2006 ; Loeb et al. 2000; Loeb et al. 2001) したがっ 、Cry
毒素 昆虫個体の死が実際 の う 出来事の組み 合わせ 起こっ い の
い 解答を出すた 、先 述 た2種類の毒素の作用機構仮説 宿主側の生
体防御反応の全 を想定したうえ もう一度昆虫個体の組織 起こ 出来事を詳細
観察す 必要があ いく の先行研究 おい 、致死量のCry毒素を暴露した
ョウ目昆虫 おい 組織学的 解析が行わ きた (Sousa et al. 2010; NIsitsutsuji-Uwo
1980; Abdel-Razek 2002) し しそ の多く 、毒素 っ 細胞傷害が起こ
のみフ ー スを当 、高濃度のCry毒素を投与した実験 あった 実際 起
こっ い 免疫反応が顕在化す の その効果が発揮さ 条件 観察した際 限
もし いの 、昆虫個体 起き い こ を正しく知 致死濃度の毒素
を投与した際の組織を観察す け 十分 あ もし い そこ 本章 、
イコ 幼虫 対し 致死濃度 く、sublethal (亜致死濃度 ; 死 死
い の濃度 あ い 、そ く容易 組織が治癒しう 濃度 ま 設
定を広げ 中腸組織 起こ 反応 い 観察し、昆虫消化管細胞 実際 起こっ い
13
第2節 材料 方法
1.2.1 供試昆虫
イコ (品種 錦秋×鐘和;上田蚕種株式会社) を25℃、明期16 時間、暗期 8 時
間の条件 、人口試料 (シ ク イト;日本農産工業) を給餌し 飼育した
1.2.2 Cry1Aa毒素の活性化 精製
前駆体型 Cry1Aa を発現す 組換え型大腸菌 、Bacillus thuringiensis var. kustaski HD-1-Dipel 株 ク ー ニ ン さ た cry1Aa 遺 伝 子 を 持 pES1 (Schenepf et al. 1985)を鋳型 、GST融合型発現ベクターpGEX-4T-3 (Amersham pharmacia Biotech) サ
ク ーニン した後、大腸菌株BL21 を形質転換し 作製さ た (Ikeda et al.2000)
前駆体型Cry1Aaを発現す 組換え大腸菌 、フ ー ストック 釣菌し、50 µg/ ml
アン シ ンを含 LB培地5 ml中 37℃、一晩培養した 培養液を50 µg/ mlアン
シ ンを含 LB培地250 ml 全量加え、37℃ 1.5時間培養した後、IPTGを終濃度1
mM う 加え,37℃、一晩培養し 毒素タン ク質を強制発現させた 菌体を
5,870 g,10 分 間 の 遠 心 分 離 回 し 超 音 波 処 理 し 菌 体 を 破 し た 後 、1% TritonX-100 室温15分間イン ュベートし、11,302 g、10分間遠心分離した後上清を
除去し,封入体を回 した 回 した封入体 、Elix水中 懸濁、11302 g、10分間遠
心分離し、上清を除去す 操作を 2 回繰 返した 上清回 後の封入体 、2.5 ml の
Elix 水 懸濁し、2 M Tris-HCl (pH8.3) を50 µl、1 M DTTを5 µl、1 M NaOHを50 µl
加え氷上 ッテ ン 後、15 分静置し、再び10,000 rpm 30分間遠心分離し上清
を回 し可溶化毒素溶液 した 可溶化毒素溶液 、透析 50 mM Tris-HCl (pH8.3)
バッフ ー置換し,28,933 g 60 分間遠心分離し 溶成分を沈殿させ除去した 上
14
吸着させた Tris-HCl (pH8.3) の濃度を200 mM 変え、40分間バッフ ーを流
し 中の非吸着物質を洗い流した後、0.5 mg/ mlのト シン溶液を2 ml
イン クトし、 中をト シン溶液 満たした後 を し、37℃、2
時間反応させた 再び を 付け、Tris-HCl (pH8.3) バッフ ーを40分間 150
mM – 500 mM う イ ー エント 流し、フ クションコ クター
活性化させた毒素を回 した 得 たフ クションを10% Sodium dodecyl sulfate – polyacrylamide gel 電気泳動 (以 SDS-PAGE; Laemmli et al. 1970) っ 解析し、60
kDa付近の単一 バン を含 フ クションを回 した こ を透析 Elix水
バッフ ー置換した後、凍結遠心エバポ ーターを用い 500 µl -1 mlの範囲の容量
ま 濃縮し、1 M PBSを得 たタン ク質溶液 等量加えた タン ク質の濃度
、SDS-PAGE タン ク質を分離、CBB 染色した 目的のバン の濃さを
ンシト ト ー法 測定した
1.2.3 イコ 幼虫を用いたバイ アッセイ
12時間絶食させた3齢脱皮直後の幼虫を各区30 試料 ッ 選別した 毒素
処理区 、Cry1Aa毒素を 0.05 - 1.0 µg/ g diet う Elix水 希釈し、各100 µl
う し、人工試料3 g 混 イコ 3齢起幼虫 与えた 対照区 、Elix
水100 µlを人工試料3 g 混 与えた バイ アッセイ 各区画3 連 行い 各時間
おけ 死亡個体数を計測した
1.2.4 イコ 幼虫組織切片の作製
1.2.3 同様の手法 毒素を摂食させた イコ 3齢幼虫及び対照区の幼虫を2時間、4
時間、24時間、72 時間のそ の時点 回 し、頭部 尾部を切断除去後、組織の
15
後、PBS 5分間洗い、70%エタノー 中 30分間、90%エタノー 中 30分間静置
し、新しい90%エタノー 中 4℃ 一晩静置した
そし 以 の組成の溶液中 そ 適正時間静置し、脱水、透徹を行った
1. 35%1- タノー ,50%エタノー /水 1時間 2. 55%1- タノー ,40%エタノー /水 1時間 3. 75%1- タノー ,25%エタノー /水 1時間 4. 100%1- タノー 1時間 5. 100%1- タノー 4 ℃ 一晩
以上の操作を行ったサン を60 ℃の恒温層 融解し おいたPathoprep 568 (病理組
織包埋用,WAKO) 等量の1- タノー を加えた溶液中 1.5 - 2時間静置した その
後、24 well マイク ート中 融解し おいたPathoprep中 1時間静置し、新しい
Pathoprep層へ移動す 操作を3回繰 返し、サン を新しいPathoprepへ移動させす
氷水へ浸し急速 固ま せた 得 た フ ン ック 木 貼 付け、
ク トー (Leica) 10 µmの厚さの連続切片 し、MASコートス イ ス (松
波硝子工業) 乗せ、40 ℃ 設定したヒート ック上 2 - 4時間伸展させ使用す
ま 4 ℃ 保存した 脱 フ ン 再加水をおこ うた 、伸展後のス イ
ス上の フ ン切片を 以 の溶液 指定時間 順 移した
16 8. PBS 5分
1.2.5 TUNEL 染色
1.2.4 得 た 脱 フ ン 及 び 加 水 処 理 済 み の フ ン 切 片 を DeadEnd™
Fluorometric TUNEL System (promega) の トコー 従い染色した 完成した切片 、
蛍 抗体体色防 液 (1.25%DABCO 1,4- ア シク [2,2,2] クタン,90% セ
ー / PBS) 封入し、蛍 顕微鏡 観察した
1.2.6 イコ 幼虫中腸のcDNA作製 ア タイ PCR
3齢 イコ 幼虫各30 分の中腸を摘出し、ISOGEN (ニッポン ーン) の トコ
ー 従いトータ RNAを調整した トータ RNA DNase I FPLCpure
TM (GE
ス ア) の トコー 従いDNase I 処理反応を行った後、ヒート ックを用い
75℃15分間処理す こ DNaseIを失活させた その後、エタノー 沈殿を行い、RNA
の ットを風乾させ、RNase free water 溶解した DNase処理が終わったtotal RNA
、High-Capacity cDNA reverse Transcription Kit (ア イ バイ システ ) を使い、
添付の トコー 従っ 逆転写反応を行いcDNAを合成した 作製したcDNA 、
RNase free water 適量 希釈し、SYBR Premix �� �����II (TaKaRa) を使った ア タ
イ PCRを行った 使用した イマー い Table 2 示した また、 ア タ
イ PCR反応 、Roter-GeneQ (Qiagen) を用い 行い、得 た結果 、delta-delta Ct
17 第3節 結果
1.3.1. 異 濃度の Cry1Aa投与濃度 イコ 3 齢幼虫の病状 の関係の継時的観
察
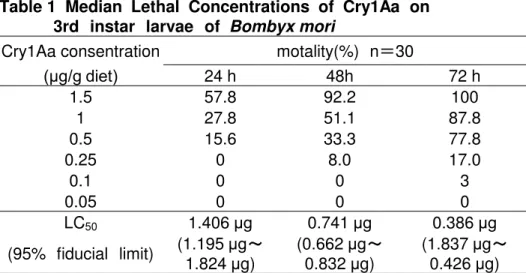
Cry1Aa 毒素投与濃度 生死や摂食停 の イコ 幼虫の反応 の関係を明
す 目的 摂食実験を行った (Table 1) 全 の濃度の実験区 Cry1Aa毒素処理後2 -
3 時間経過す イコ 3 齢幼虫 、Cry1Aa を含 人工飼料を食 のをや た
1.0 µg/g diet 区の イコ 幼虫 24時間後ま 27.8%が死亡し、72時間後 87.8%
が死亡した 0.5 µg/g diet区の イコ 幼虫 24時間後ま 15.6%が死亡し、72時間
後 、77.8%が死亡した 一方、0.25 µg/g diet区の イコ 幼虫の死亡率 、48時間
後 8.0%、72時間後 17% し く、死亡し い い個体 摂食阻害がみ た
が、その後の観察 おい 、死亡す 個体 観察さ った また、0.1 µg/g diet区
お び0.05 µg/ g diet区 、摂食阻害 対照区 比較し イコ 幼虫の体のサ
イ が一回 さく ったが、死亡個体数 72時間後ま 0.1 µg/g diet区 1
け(3%) (Table 1) あった その後の観察 おい も両区画 おい 死亡個体 こ 以上
出現し った
1.3.2 Cry1Aa毒素投与後の中腸上皮細胞の経時的観察
2 の作用機構仮説“Pore formation model” “Signal transduction model”のい
適合す 現象が中腸 おい 認 、あ い イコ 幼虫の応答の1 し
アポトーシスが起こ の を検討す た 、Cry1Aa 毒素投与後の イコ 3 齢幼虫
の中腸を経時的 固定し、切片を作製し DAPI染色お びTUNEL染色をし、蛍 顕
微鏡 観察した 対照区の組織 、大き 核を持 筒細胞 細胞中 空隙を持 杯
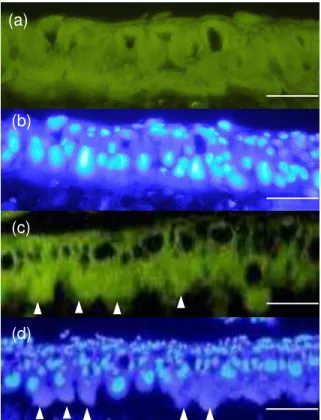
状細胞が、高密度 整然 一層の細胞層を形成し い 様子が観察さ た (Fig. 1a, b)
18
させた 2 時間後のサン 、多くの 筒細胞が膨潤し い 様子が観察さ た
(Fig. 1c, d) アポトーシスを起こし い こ を示すTUNEL陽性細胞 検出さ
った Fig. 1c 1.0 µg/g diet、摂食開始4時間後の切片 、2時間後 同様 膨潤し
た 筒細胞が存在す 場所も見 たが (data not shown) 多くの 筒細胞 破裂した
後 あ 、異常 形態をした細胞や細胞残骸が多数み た (Fig. 2a) 一方、アポ
トーシスを起こし い 細胞 観察さ った そし 同条件 おい 27.8%が死
った 24時間後の組織を観察した こ 、 筒細胞の存在頻度 し、し も形態
を保った 筒細胞 ほ 観察さ 、組織 崩壊状態 あった(Fig. 2b) 一方、2
時間後、4 時間後同様アポトーシスを起こし い 細胞 観察さ った(Fig. 2b)
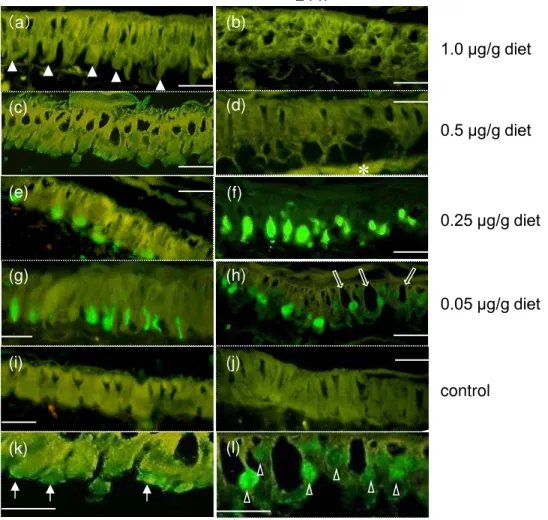
24時間後ま 15.6%が死亡した0.5 µg/g diet投与区 おい 、4時間後 多くの
筒細胞が膨潤し い のが観察さ た Fig. 2c) また、興味深いこ 、その う
細胞の多く 、核が管腔側 移動し お 、 ベ あ がTUNEL染色さ い
た(Fig. 2c, k) また、24時間後 中腸上皮組織の一部 正常 中腸上皮組織 み
た 筒細胞 杯状細胞の整然 した配列が乱 た領域がみ 、この時間条件 おい
組織の残っ い 細胞 アポトーシスを起こし い こ を示すTUNEL陽性の細
胞 検出さ ったが、多くの細胞残骸が囲食膜 上皮組織 の間 観察さ た Fig.
2d 24時間後ま イコ 幼虫が1 も死 った0.25 µg/g diet 0.05µg/g diet
区の 2 時間後の観察 、 筒細胞の膨潤も TUNEL 陽性の細胞も観察さ った
(data not shown) し し、4時間後の組織 おい 、 筒細胞の膨潤 み っ
たが、TUNEL陽性 あ 筒細胞の大型の核が多数観察さ た (Fig. 2e, g) こ の
細胞の形態 通常の 筒細胞 あま 変わ ったが、核 ー ン側 移動し 細
胞の先端 (apical side) あった また、縦長の形態を 核も見 た (Fig. 2e, g) 24
時間後の組織の観察 も引き続き 筒細胞 あ こ を示す大型の核を持 TUENL陽
19
ア ポ ト ー シ ス 様 の 細 胞 ほ 観 察 さ っ た が 、 逆 ほ の 筒 細 胞 が
TUNEL陽性を示す場所も観察さ た (Fig. 3c) おTUNEL陽性細胞がほ 観察
さ った領域の中腸上皮細胞 おい 概し 筒細胞が少 く っ いた (Fig.
3h) また杯状細胞の空隙が対照区 おけ 中腸上皮細胞の杯状細胞 のもの も広が
っ いた (Fig. 2h) そし 、アポトーシス様細胞が高頻度 み 部位 おい
TUNEL陽性細胞が中腸上皮組織 脱落し い 様子が観察さ た (Fig. 3d こ
、ここ 観察さ た間隙の広がった杯状細胞が多く存在し い 領域 おい 筒
細胞がす アポトーシス様の反応 脱落した後 あ こ を示唆し い 考え
た さ TUNEL陽性細胞のほ が、中腸上皮細胞の中 最も大き 核を持
成熟した 筒細胞 あ こ 、DAPI 核染色の の比較 更 確
た(Fig. 3a) また、場所 っ 、こ 成熟した 筒細胞の核の他 、未成熟
筒細胞の比較的 さ 核も弱くTUNEL染色さ い 様子が観察さ た(Fig. 2h, i)
24時間後 最もTUNEL陽性細胞が多く観察さ た0.05 µg/g dietのCry1Aa毒素投与区
72時間後の組織をTUNEL染色した 同標本 おい 、24時間後 高頻度 見
たTUNEL陽性細胞 観察さ く 、アポトーシスを起こし減少し いた う
見えた 筒細胞の数も対照区 (Fig. 4b) 同程度ま 復帰し、広がっ いた杯状細胞の
空隙も正常時の状態 戻っ い 様子が観察さ た (Fig. 4b)
1.3.3 Cry1Aa処理 イコ 幼虫中腸 のアポトーシス関連遺伝子発現 ベ の比較
中腸上皮組織 観察さ たアポトーシス の う 細胞内シ 伝達経路が
関与し い の を調 目的 、Bm-Apaf1、Bm-MAPKp38、Bm-Jun発現量の変化を
解 析 し た ま た 、 組 織 の 更 新 伴 う 腸 管 の 肝 細 胞 の 分 化 増 殖 の 進 を 制 御 し い
Bm-Socs い も同様の解析を行った ribosomalL31 遺伝子を フ ンス遺伝子
20
PCR 相対定量した Cry1Aa毒素0.1 µg/g dietの区 1.0 µg/g dietの区の両区 おい
そ の遺伝子の発現量を比較した こ 、Bm-Socs い 、1.9倍 2.5倍、
Bm-Jun両区 おい 3.4倍 3倍、そし Bm-MAPKp38 い 、1.9倍 1.1倍、
21
第4節 考察
1.4.1 消化管上皮細胞 起こっ い 出来事
3日以内 ほ が死 がそ ほ 高い濃度条件 言え い1.0 µg/g diet お び
0.5 µg/g diet Cry1Aaを投与した イコ 幼虫の組織の観察 、4時間以内 筒
細胞が膨潤し い 様子が観察さ 、24 時間後 組織が崩壊し い 様子が観察さ
た Fig. 1c, d, Fig. 2a, b 先行し 致死濃度のCry毒素摂食 イコ 幼虫の顕微鏡観
察を行っ い Nisitsutsuji-Uwo の ー 、中腸上皮細胞の特 筒細胞が膨潤す
様子が毒素処理 60~90分後 顕著 観察さ いう本研究 類似した結果を
報告し い (Nisitsutsuji-Uwo.1980) こ のこ 、まさ Pore formation model の
仮説 考え い 浸透 依存的 細胞の膨潤、破裂(Knowles 1987) いう現象が消
化管 起こったこ を示唆し い 一方、Smouse and Nishiura の ー (Smouse
and Nishiura 1997) 、Cry毒素 っ 誘導さ たアポトーシスがCulex pipeiensの個
体死の原因 っ い 結論付け い 本実験 、 イコ 幼虫が、死
Cry毒素濃度条件 あ 0.5 µg~1.0 µg/ g diet 、膨潤し い 細胞 おい ベ
TUNEL 染色さ 細胞を比較的 頻度 検出した け 、TUNEL 陽性 く
膨潤、破裂し い 細胞が主ほ あった (Fig.2 a, c, b, d) この結果 、Cry毒
素 中腸の崩壊 そ 続く昆虫の個体死 、アポトーシスが主要因 っ い
考え った(Fig. 6) また、前述した う ン様タン ク質を発現させ
た培養細胞 、PKAの 流 oncosis様の 細胞死 細胞が死 こ が示唆さ
い が(Zhang et al. 2005)、oncosis様の 細胞死 際立った形態的特徴
報告 い いた 、Zhang の主張す う oncosis 様の 細胞死が同時
進行し た 否 い 定 った
こ 対し 、多くの幼虫が最後ま 生き残 濃度のCry1Aa毒素を処理した条件
22
う 細胞が膨潤す 様子が観察さ った(Fig. 2e-j, Fig. 4) そし 注目す き 、
Cry1Aa毒素処理 4時間後及び24時間後 おい TUNEL陽性のアポトーシスを起
こし い 考え 細胞が観察さ たこ あ (Fig. 2e-h, Fig. 3b,c) こ 、
この組織 観察さ た顕著 アポトーシス様の反応のほ 、中腸上皮細胞を構成
す 細胞 最も大き 核を持っ い 筒細胞 起こっ いた(Fig. 2e-h, Fig. 3b,c) 中
腸上皮細胞 筒細胞上 、Cry毒素の受容体 し 働く 分子が存在し い 考え
い こ (Hofmann et al. 1988)、ここ 観察さ たアポトーシスを起こし い
細胞 、Cry毒素 膨潤を伴わ い ベ の弱い 害を受けた細胞 あ 可能性
が考え また、特 24時間後の組織の観察 、 筒細胞がアポトーシスを起
こし い こ 加え、杯状細胞の空隙が広がっ い 様子が観察さ た(Fig. 2c,d)
この う Cry毒素摂食後の感受性昆虫の消化管上皮組織の杯状細胞の形態変化 、
イコ 、そし Alabama argillacea の幼虫中腸 も報告さ い (Sousa et al.2010;
Abdel-Razek 2002) この現象 、 筒細胞がアポトーシス っ 死 脱落したこ
中腸上皮細胞を構成す 細胞数が減少したこ 伴うもの あ 、中腸が組織崩
壊す のを防 こ 役立っ い う 見えた さ 、sublethal 条件の Cry1Aa
毒素摂食後72時間経過後の組織の観察を行った こ 、24時間後 頻繁 み た
TUNEL 陽性細胞が検出さ 、上皮組織の 筒細胞の頻度 対照区 同程度ま 復帰
し、広がっ いた杯状細胞の空隙も対照区 同様の状態 戻っ いた(Fig. 4) こ
のこ 、Cry1Aa毒素の弱い作用を受けたが膨潤す ほ った 筒細胞 、4
時間後 24時間後、あ い そ 以降ま け アポトーシスを起こし、脱落し、
その後 組織の更新が起こったこ を示唆し い (Fig. 6) In vitroの実験 あ が、
H.virescens の 中 腸 の 培 養 細 胞 、Bacillus thuringiensis AA1-9 も し く 、Bacillus
thuringiesnsis HD-73を添加した こ 、アポトーシスが誘導さ 筒細胞、杯状細胞の
23
Loeb et al. 2001) また、in vivo も、Drosophila Pseudomonas aeruginosa いう細菌
を摂食させ 、傷害を受けた成熟したenterocyteがアポトーシスを起こし、幹細胞の
分化が 進す こ を中腸上皮細胞 のCaspase3の活性を 述 い (Apidianakis,
et al. 2009) したがっ アポトーシス 、新た 細胞の増殖分化 ンクした傷害を受
けた中腸上皮細胞の速や 排除 組織更新 寄与す もの の もし い
1.4.2 消化管免疫誘導の分子機構
Cancino-Rodezno MAPK p38 の RNAi を M. sexta A. aegypti い 行い、
Cry1Ab毒素及びCry11Aa毒素 対す 感受性が、そ 、8倍及び10倍増加したこ
を示し、MAPKp38がCry毒素を摂食した昆虫が生存す た 必須 あ いう考
えを示した (Cancino-Rodezno et al. 2010) このMAPK p38経路、そし 同 くMAPK
の1 あ JNK経路の 流 、アポトーシスの引き金 Caspase-6、Caspase-9、
Caspase-3が活性化さ こ が報告さ い (Porta et al. 2011; N'Guessan et al. 2005)
一方、Cancino-Rodezno の実験 おい Cry1Ab毒素を摂食したM. Sexta のMAPKp38 の発現量が7日後 約7倍増加したの 対し (Cancino-Rodezno et al. 2010)、 々の結 果 Bm-MAPK p38 い 0.1 µg/g dietの区画 1.9倍、1.0 µg/g dietの区画 1.1
倍 あ 、増加 軽微 あ その意味 明 あった 一方、Jun い 、0.1 µg/g
dietの区画 3.4倍、1.0 µg/g dietの区画 3倍程度の発現量の増加がみ (Fig. 5)、
この経路が何等 の役割を果たし い 可能性が考え た Apaf1 トコン
ア おい シトク C 結合し、Caspase9 を活性化す 過程 関与し お 、先行
研究 おい アポトーシスが起こ 際 転写 ベ の活性化が起こ こ がヒトや
Drosophila おい 報告さ い (Zhou et al. 1999; Fortin et al. 2001) 近年、 イコ
もアポトーシス経路 関連した遺伝子の解析が積極的 行わ い 、他の種 報告
24
2010) 具体的 、death receptor pathway、epidermal growth factor pathway、そし Apaf1
が関与し い mitochondrial apoptotic pathway 関わ 遺伝子が同定さ い 、得
Apaf1が関与す mitochondrial apoptotic pathway い 、 イコ 由来の培養細胞
あ BmE cells おい スト スを感 た際 トコン ア シトク Cが有
意 放出さ たこ この経路が イコ おい アポトーシスが誘導さ 際
利用さ い こ が示唆さ た(Pan MH 2009) 本研究 おい 、 イコ のApaf1
oltholog あ Bm-Apaf1遺伝子の転写 ベ を解析した こ 、アポトーシスを頻繁
起こし いた0.1 µg/g diet いうsublethal Cry1Aa毒素処理区 おい Cry1Aa摂食
開始 6時間後 の転写が対照区 比較し 約7倍活性化さ たこ が確認さ 、中
腸 観察さ たアポトーシスがBm-Apaf1 を介す mitochondrial apoptotic pathwayを利
用し い 可能性があ 考え た さ 興味深いこ 、アポトーシスが組織
おい 全く観察さ った1.0 µg/g diet おい も同遺伝子が、sublethal Cry1Aa
毒素濃度条件 あ 0.1 µg/g diet 6時間後 検出したもの 同程度BmApaf1が活性化
さ た いう結果が得 た(Fig. 5 and 6) このこ 、1.0 µg/g diet いう イコ が
致死す 条件 も、Cry1Aa 毒素 アポトーシス 、転写 ベ 誘導さ
い 可能性を示唆し い また、 イコ が致死 条件 あ 0.5 µg/g diet お
い 膨潤し い 細胞 ベ のTUNEL陽性細胞が観察さ た 中腸上皮組織の
顕微鏡観察の結果 Fig. 2c も矛盾し い し し、実際 アポトーシスの過程が細
胞 進 いく前 、Cry毒素 形成さ Pore のイ ン流入 細胞の膨
潤、破裂の方が先行し 起こ た 、アポトーシスが進行す 前 細胞が受動的 死
しまっ い の もし い
さ 近年、他の生物種 おい 、Cry毒素同様、宿主の細胞 スト スを与え 様々
因子が、幹細胞の分化増殖の引き金 あ JAK/STAT経路を活性化す いう報告が
25
、JAK/STAT 経 路 の 活 性 化 を 意 味 す Bm-Socs の 転 写 ベ を 解 析 す こ JAK/STAT経路の活性化の程度を評価した こ 、0.1 µg/g diet 区画 1.9倍、1.0 µg/g diet 2.5 倍 いう結果が得 た(Fig. 5 and 6) 2009 年の Jiang の研究結果
Drosophilaの腸細胞 おい アポトーシスがJAK/STAT経路活性化のト ー こ
が示さ い こ (Jiang et al. 2009)、本研究 観察さ たCry毒素 っ 誘
Table 1 Median Lethal Concentrations of Cry1Aa on 3rd instar larvae of Bombyx mori
Cry1Aa consentration motality(%) n=30
(μg/g diet) 24 h 48h 72 h
1.5 57.8 92.2 100
1 27.8 51.1 87.8
0.5 15.6 33.3 77.8
0.25 0 8.0 17.0
0.1 0 0 3
0.05 0 0 0
LC50 1.406 μg 0.741 μg 0.386 μg
(95% fiducial limit) (1.195 μg~ 1.824 μg)
(0.662 μg~ 0.832 μg)
(1.837 μg~ 0.426 μg)
(b)
(d) (c) (a)
Fig. 1. Reaction of midgut epithelial cells treated with lethal dose of Cry1Aa.
Mock (a, b) or diet treated with lethal dose of Cry1Aa (1.0 µg/g diet) (c, d) midgut were fixed 2 h after treatment, and paraffin sections were prepared and stained by the TUNEL (a, d) method or counterstained with DAPI (b, d). Nuclei of the midgut epithelial cells were stained light blue with DAPI (b, d). Arrowheads indicate swollen columnar cells of which cytoplasm protruded into the lumen (c, d). Sections were examined under the
Fluorescence microscopes. The scale bars represent 20 μm.
Fig. 2. Reaction of midgut epithelial cells treated with different concentration of Cry1Aa.
Bombyx mori larval midgut treated with 4 steps of Cry1Aa (0.05 µg/g diet – 1.0 µg/g diet) were fixed 4 h (a, c, e, g, i) or 24 h (b, d, f, h, j, k, l) after treated, sections were made and stained by TUNEL. The sections were observed under the fluorescence microscopes. The closed allow head indicates wreckage of the burst cells (a). The light green fluorescein indicates the TUNEL positive nuclei (e, f, g, h).The closed allows indicate weakly TUNLE positive nuclei (k). The open allow heads indicateTUNEL positive nuclei of immature cell (I). The open allows indicates goblet cells with enlarged cavity(h). Asterisk indicates cell debris accumulated on the peritorophic membrane.The scale bars represent 20 μm.
a. Cry1Aa 1.0 μg/g diet b. Cry1Aa 1.0 μg/g diet c. Cry1Aa 0.5 μg/g diet d. Cry1Aa 0.5 μg/g diet e. Cry1Aa0.25μg/g diet f . Cry1Aa0.25μg/g diet g. Cry1Aa0.05μg/g diet h. Cry1Aa0.05μg/g diet i. control j. control k. magnified figure of fig. h l. magnified figure of fig. f
*
(h)
4 h 24 h
control
0.05 μg/g diet 0.25 μg/g diet 0.5 μg/g diet 1.0 μg/g diet
(e)
50 μm
(b) (f) (h) (g) (f) (i) (d) (j) (c) (f) (a (d) (f)
(k) ((l) l)
*
Fig.3. The spatial pattern of apoptotic cells
Midgut of 3rd instar larvae of Bombyx mori treated with sublethal concentration of Cry1Aa (0.05 µg/g diet) were fixed 72 h after treatment, then the section were stained with DAPI (a) and by TUNEL (b, c, d) The allows indicates cell nucleus stained by TUNEL staining (b). The allow head indicates nuclei disintegrated and shifted to the lumen side due to apoptosis (d). The scale bars represent 20 μm (a, b, d),50μm (c).
(a)
(c)
(b)
(d)
(a)
(b)
Fig.4. Bombyx mori midgut epithlial cells t 72 h after treatment
Midgut of 3rd instar larvae of Bombyx mori were fixed 72 h after treated with Cry1Aa (0.05 µg/g diet).The sections were conducted TUNEL staining (a), mock control (b). The scale bars represent 20 μm.
Fig.5. Analysis of host defense related genes expression
The expression levels of each genes on 3rd instar larval midgut 6h after treatment with Cry1Aa were detected by ⊿⊿Ct method by real time RT-PCR. The cDNA were prepared from 30 larval midguts. Three replicates were
performed. Open bars indicate gene expression of the insects administered with Cry1Aa at 0.1 μg/g diet relative to the control insects. Closed bars indicate gene expression of the insects administered with Cry1Aa at 1.0 μg/g diet relative to the control insects.
0 1 2 3 4 5 6 7 8
Socs Jun MAPKp38 Apaf1
R
elat
iv
e
ge
ne
ex
po
ression
to
the
co
ntorol
i
ns
ec
ts
Fig.6. The model of Bombyx mori midgut epithelial cell’s reaction against Cry1Aa In the lethal condition of Cry1Aa administration, midgut epithelial cells were swelled and busted, and then midgut tissue collapsed. Although apoptosis happened in parallel, swelling and bust were seemed to progress faster than apoptosis. In the sublethal or lower condition of Cry1Aa administration, apoptosis seems to lead to renewal of columnar cells layer in the midgut for the repair of damage.
lethal dose of Cry1Aa
apoptosis of columnar cells Repair of midgut epitherium Disruption of midgut epitherum
Activated Cry1Aa toxin
Swelling and burst of columnar cells
Bm Apaf1 Cry1Aa doses in
sublethal to non-lethal
Table 2. oligonucleotides used for real-time PCR
oligonucleotide sequence Accession Number Bm-MAPKp38-F 5' ttggattgctggatgtgtttac
AB208585 Bm-MAPKp38-R 5' catcgtacggacacagaagct
Bm-Socs-F 5' cagtgtccagagatatgtgagc
BGIBMGA009619 Bm-Socs-R 5' taagcagtgcagacgcag
Bm-Jun-F 5' cacagcatggagaccaccttc
BGIBMGA004164 Bm-Jun-R 5' gttcacgaatcgctgcac
Bm-Apaf1-F 5' atactggtgcttcacggtatg
BGIBMGA011028 Bm-Apaf1-R 5' gcgaattgcaagaccgatgac
Libo L31-F 5' ccaagagcaatcaaagaaatc
BGIBMGA000959 Libo L31-R 5' gtcagaaatgttcccttcacgt
34
第2章 イコ ABC transporter C2のCry毒素殺虫作用機構上 の役割の解析
第1節 目的
序章や第 1 章 第1 節 既 述 た通 、Cry毒素作用機構仮説 最も多くの論文
引用さ い の 、“Pore formation model” (Bravo et al. 2004) “Signal transduction model ” (Zhang et al. 2005; Zhang, et al. 2006) あ こ の仮説 いたCry毒素作
用機構上 、Cry毒素 、宿主昆虫の消化管上皮細胞上 発現す 受容体分子 相互
作用す こ が必要 あ 、Cry1A毒素 い 、既 複数の分子が実際 受容体
し 機能 す 可 能 性が示 さ い その 1 あ ン様タ ン ク 質 、
Bombyx mori (Nagamatsu et al. 1999)、Manduca sexta (Hua et al. 2004) 、Heliothis virescence
(Jurat-Fuentes et al. 2006)、Ostrinia nubilalis (Flannagan et al. 2005) おい Cry1A毒素
対す 感受性を培養細胞 付与す こ の き 分子 あ こ が示さ い また、
実際 昆虫個体を用いたCry毒素抵抗性の研究 おい 、H. virescens Gahan et al. 2001
、Helicoverpa armigera (Xu et al. 2005)、Pectinophora gossypiella (Morin et al. 2003)の抵
抗性遺伝子の遺伝学的解析 、 ン分子が正しく発現 き く こ が、抵
抗性の1 の原因 あ こ が示さ い 一方 、 コシ スフ イノ
シトー (glycosylphosphatidylinositol、以 GPI)アン ー型のタン ク質の1 、ア ノ ー N (Amino peptidase N、以 APN)、特 APN Class 1 (以 APN1) 、
Drosophla melanogaster 発現させ こ 、Cry1Ac 対す 感受性が生ま た の報
告や (Gill et al. 2002)、APN1のサイ ンシン がCry毒素 対す 幼虫個体の感受性を させ の報告があ (Yang et al. 2010; Tiewsiri et al. 2011) また、Cry1Ac抵抗性の
H. armigera おい APN1分子 変異が入っ いた の報告があ Zhang et al 2009
し し、 ン様タン ク質の う 遺伝学的 APN1がCry毒素抵抗性原因因子
35
っ い い 一方、APN 同様のGPIアン ー型タン ク質 あ ア フ
スフ ター (Alkaline phosphatase、以 ALP) い 、異所発現実験 Cry
毒素感受性を付与す 分子 あ いう報告 いが、Cry 毒素抵抗性昆虫 おい
ALP 対 す 結 合 性 の が 抵 抗 性 相 関 関 係 が あ こ が 報 告 さ い
(Jurat-Fuentes et al. 2004; Jurat-Fuentes et al. 2011) そし 、2010年、Gahan 今ま
Cry毒素殺虫作用機構 の関連が一度も報告さ たこ の いABC ト ンスポータ
ーC2 (ATP-binding cassette transporters subfamilyC member2、以 ABCC2) がCry1Ac抵
抗性H. virescenseの抵抗性原因因子 あ こ が ン ー マッ ン の結果 し 報
告さ た(Gahan et al. 2010) また、2012年 、 イコ おい もCry1Ab抵抗性
イコ の ン ー マッ ン 解析が行わ た結果 ABCC2分子がCry1Ab抵抗
性 あ こ が報告さ た Atsumi et al. 2012 さ 、先行研究 おい ン
様タン ク質やAPN 変異が認 ったCry1Ac抵抗性のP. xylostella やT. ni
おい もABCC2分子内 変異があ こ が新た 見 Baxter et al. 2011 、ABCC2
分子が Cry 毒素殺虫作用機構上 重要 働きをす 分子 あ こ が期待さ
った
そこ 、本章 、 イコ のABCC2 あ BmABCC2がCry毒素作用機構上
の う 役割をす 分子 あ の を解明す こ した ABCC2 分子 、細胞膜上
発現す 分子 あ こ 、Cry毒素受容体 し 機能す こ が期待さ た そ
こ 、Cry1A 毒素抵抗性 あ 昆虫培養細胞 Sf9 細胞 BmABCC2 分子を発現させ、
Cry毒素感受性を調査す こ BmABCC2が受容体分子 あ 否 を評価す こ
した また、前述した、従来 Cry1A 毒素殺虫作用機構 おい 重要 考え
きた ン様タン ク質 の受容体 し の機能の大きさを比 こ 、Cry毒
素感受性を生み出すの 重要 受容体 の分子 あ を1 の実験系を用い 評
36
第2節 材料 方法
2.2.1 昆虫培養細胞
Sf9 細胞 ( ス ン トウ近縁種 Spodoptera frugiperda 卵巣由来) を培養フ スコ (tissue-treated, IWAKI) 中 , 10 % (v/v) 牛胎児由来血清 (FBS, JRH Bioscience) を添加
したSf-900 II SFM (GiBCO, Invitrogen) 28℃ フ スコ底面 付着させ 生育さ せた
2.2.2 cDNAのク ーニン
Enhanced green fluorescent protein (以 EGFP)をコー す cDNA pA3hr5(-)EGFP
vector ( 山 口 大 学 林 淳 教 授 分 与 ) を 鋳 型 し 、
5'-tcggTCTAGAatggtgagcaagggcgagga-3' 5'-gatgAGGCCTtacattgtacagctcgtcca-3'の イ
マ ー を 用 い PCR 反 応 増 幅 し 、3 種 の cDNA を 同 時 発 現 き ベ ク タ ー
pBAC4x-1 (Novagen) のXbaI 、StuIサイト 組み込 完全長BtR175 (以 BtR-Full; AB026260)をコー す cDNA 、3 齢 イコ (錦秋×昭和) 中腸の cDNA を鋳型 5'-caagcgtAAGCTTatgggagttgacgttcg-3' 5'-acgaatcacgagctcgggtatggcaaccatga-3'の イ
マーを用い、EGFPを組み込 pBAC4x-1(pBAC4x-1-EGFP)のHindIII XhoIサイト 組 み 込 シ BtR175 毒 素 結 合 領 域 (Toxin binding region; 以 BtR175-TBR) のcDNA断片 、3齢 イコ (錦秋×昭和) 中腸のcDNAを鋳型 そ
、5'-caagcgtAAGCTTatgggagttgacgttcg-3' 5'-acgaatcacgagct cgggtatggcaaccatga-3'、 5'-tggttgccatacccgagctcgtgattcgtcc-3' 5'-cctacagCTCGAGtttctggaattgatttgc-3'の イ マ ー
をも い PCR反応 増幅し、overlap extension PCR cDNA断片を連結し、
pBAC4x-1-EGFPのHindIII XhoIサイト 組み込 APNをコー す cDNA 、
3 齢 イ コ ( 錦 秋 × 昭 和 ) 中 腸 の cDNA を 鋳 型 そ
37
マーを用い PCR 反応 増幅し、pBAC4x-1-EGFP のXhoI HindIII サイト 組
み込 BmABCC2_S BmABCC2_Rをコー す cDNA 、そ 、 イコ
(Ringetsu, race 606, https://www.gene.affrc.go.jp/ex-nises/bombygen/indexJ6-eng.html)また 、
イコ
(Chinese No. 2, race 401, https://www.gene.affrc.go.jp/ex-nises/bombygen/indexJ4-eng.html)由
来 の 中 腸 cDNA を 鋳 型 5'-cgtGCGGCCGCatgaatagtgatgggagag-3' 5'-tgaGCGGCCGCtttttctgtatttctacc-3'の イ マ ー を 用 い PCR 増 幅 し 、 pBAC4x-1-EGFP.のNotIサイト 組み込 BmABCC2_S
+234Y
及びBmABCC2_R -234Y
のcDNA配列 、3 のPCR断片をoverlap extension PCR 連結させ、作製した そ のPCR断片 、pBAC4x-1-EGFP BmABCC2_Sまた BmABCC2_Rを組み込 ものを鋳型 そ 、F1 (5'-cgtGCGGCCGCatgaatagtgatgggagag-3') and R1 (5'-taccacccagaggtagtgcaa-3'), F2 (5'-tgtaagcggagggaaactggt-3') and R2 (5'-acatgttataaatattatacttctttcagt-3'), and F3 (5'-atgatagcactcaggaagtct-3') and R3 (5'-tgaGCGGCCGCtttttctgtatttctacc-3')の イ マ ー を 用 い PCR反 応 を 行 い 増 幅 し た overlap extension PCR っ 各断片を連結させたBmABCC2_S
+234Y
及び
BmABCC2_R-234Yの 完 全長cDNA配列 、pBAC4x-1-EGFPのNotIサ イ ト 組 み込
PxABCC2_S PxABCC2_RのcDAN配列 、そ の遺伝子がク ーニン さ 組み
込 ま た ス DNA (pGEM-T easy) を 鋳 型 そ 、
5’-GCTAggatccATGGAAAACGGAAGCGGAGC-3’
5’-CGCaagcttAGGATGGTCGTCGAAGTAT-3’ ま た 、
5’-GCTAggatccATGGAAAACGGAAGCGGAGC-3’
5’-ATTaagcttCGTCTTGCGCCCCGTGCTA-3’の イマーを用い PCR反応を行い獲得
し た 獲 得 し たPCR断 片 、pBAC4x-1-EGFPのBamHI XhoIサ イ ト 組 み 込
38
合成したcDNA を鋳型 、5’-AAATgcggccgcATGGATAGCTCGCGGAAG-3’ 5’-CCGActcgagTTGGCCTTCCTTGCTAAAAC-3’の イ マ ー を 用 い PCR反 応 を 行 い
増幅した このPCR断片 、pBAC4x-1-EGFPベクターのNotI XhoIサイト 組み込
そ のcDNAを組み込 cDNA 大腸菌株DH5α 込ませ形質転換した
2.2.3 組換え型AcNPVの作製
組換え型Autographa californica nuclear polyhedrosis virus (AcNPN) を作製す た 、24
well plate (Iwaki) 播種した4.0×10 4
/ wellのSf9細胞 対し、25 ngのBacMagic DNA
125 ngのpBAC4x-1ト ンスフ ーベクターを共導入した BacMagicDNA ト ンス
フ ーベクターを共導入した細胞 、力価の高いウイ スを得 た の培養操作 、
Bac MagicTMDNA kit (Novagen) の トコー 従っ おこ った
2.2.4 Cry毒素の精製
Cry1Aa1 (AA22353) 毒素 い 、第1章2節 (2.2) 使用したもの 同様の組換
え型大腸菌を用いた Cry1Ab8 (AAA22551) 毒素のDNA配列 、Atsumi et al. (2005)
使 用 さ た Cry1Ab8 の DNA 配 列 が 組 み 込 ま た ス DNA を 鋳 型
5´tcaatCCCGGGacatggataacaatccgaacatc3´ 5´gtccaatgctGCGGCCGCttattcctcca の イマ
ーを用い PCR反応 増幅した 獲得したDNA配列 、pGEX4t-3ベクターのSmaI
NotIのサイト 組み込 Cry1Fa毒素 い 、Fentes et al. (2006)の記載通
活性化、精製さ たタン ク質をDr. Jurat-Fuentes 分与いた いたものを用いた
Cry1Ac(AAA22331)毒素のcDNA配列 、B. thuringiensis HD-73の ス DNAを鋳
型 5'-gcaaGTCGACatggataacaatccgaacat-3' and 5'-atcGCGGCCGCctattcctccataaggagt-3'
イマー PCR反応 増幅した 獲得したDNA配列 、pGEX-4t-3ベクター
39
のサイト 組み込 Cry9Aa毒素の配列 、Bacillus genetic stock center 寄せ
た Cry9Aa 遺伝子配列を組み込 pSB1402 ス (BGSC No. ECE130)を鋳型
5’-ACGCgtcgacATGAATCAAAATAAACACGG-3’
5’-CGTgcggccgcTTACTTTTCTGTTTCAACG-3’の イマーを用い PCR反応 増
幅し、pGEX-4t-3ベクターのNotI, SalIサイト 組み込 Cry毒素遺伝子を組み込
pGEX-4t-3ベクター 、大腸菌株BL21 形質転換し第1章、2.2 同様の方法 大
腸菌を培養し、Cry毒素の封入体を精製した 精製し、200 mM Tris-HCl (pH8.3) 透析
したCry毒素前駆体タン ク質 、終濃度 0.2 mg/mlのト シンを加え37℃ イン
ュベートす こ 活性化した 活性化し凝集したタン ク質 、28,933 g, 10 min,
4 ℃の遠心分離 っ 沈殿 し 回 し、PBS 3回洗った後、再びNaOH 可溶化
し、PBS 透析した PBS 透析したタン ク質 、 ンシト ト ー法 濃度を
定量し 用いた Cry9Da2毒素 、Sinkawa et al., (1999) 記載さ い Cry9Da2のみ
を産生す B. thuringiensis japonensisis株(AF032733, genebank)を培養し、同論文 あ 2
層分配法を用い 精製した毒素を他の毒素 同様の方法 活性化、定量したものを用い
た(Sinkawa et al. 1999)
2.2.5 LDHアッセイ
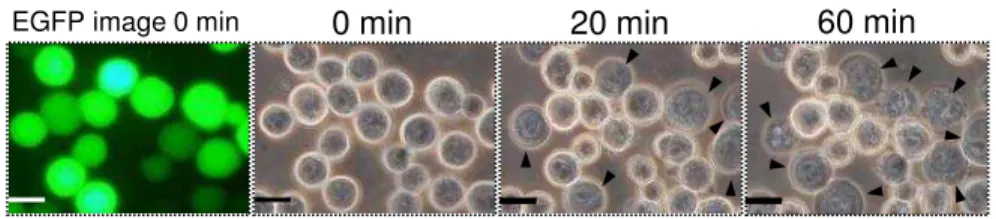
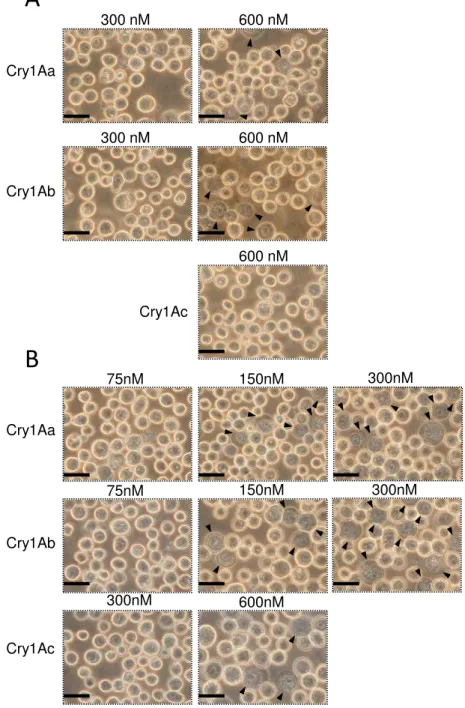
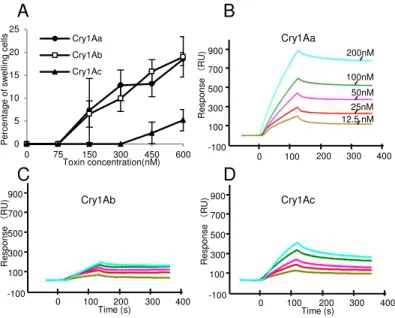
Sf9細胞 そ のAcNPVを感染させ、72時間、25℃ イン ュベートした 細
胞 、PBS 洗い、96 well ート 播種し、Cry1Aa 毒素を添加し、25℃ 1時間
イン ュベートした イン ュベート後の細胞 、Cytotoxicity Detection KitPLUS (Roche)
の トコー 従い、放出さ た乳酸脱水素酵素 (Lactate Dehydrogenase; 以 LDH)
を ート ー ーを用い 測定した
40
Sf9細胞 そ のAcNPVを感染させ、72時間、25℃ イン ュベートした 細
胞を回 し、細胞の懸濁液を バー ス上 各100 µl添加し、静置させ こ 細胞
を バー スへ貼 付けた 細胞を貼 付けた バー ス 、PBS 洗い死細胞や
活 力 を 失 っ た 細 胞 を 除 去 し た 後 、Cry 毒 素 溶 液 を 入 た 2 穴 の ス イ ス
(Matsunami) 細胞を接着させた面がCry毒素溶液 触 う 向き 被せた ス
イ ス 、60 分 25 度 イ ン ュ ベ ー ト し 、 蛍 顕 微 鏡 (BX50 Olympus light microscope) 、BX-FLA epifluorescence unit (Olympus) を用い 観察した 細胞傷害
率 、1視 中の膨潤した細胞の割合を算出す 操作を5視 分行いその 均値を用い
計算した
2.2.7 抗血清の作製
BtR175抗血清を作製す た の抗原 、Hara et al.(2003) 使用さ たBtR175の毒素
結合領域 TBR を発現す 大腸菌 発現させたタン ク質を用いた BmABCC2抗血
清を作製す た の抗原を作製す た 、本章 2.2 作製した pBAC4x-1-EGFP
BmABCC2_S の 配 列 を 組 み 込 を 鋳 型
5’-GCAAgtcgacTCGTTGGAACGCATTCAAAA-3’
5’-ATCgcggccgcTTTCACTGCAGACAAATAC-3’の イマーを用い PCR 反応を行っ
た 増幅したPCR断片 、pGEX4t-3ベクター SalI、NotIサイトを用い 組み込
組換えた ス 、大腸菌株BL21 形質転換し、第1章2.2 同様の手法 タン
ク質を生産させ、封入体を精製した 精製した封入体 、 422 エ クト エ
ューター(Bio-Rad)を用い エ クト エシューション法 目的タン ク質の断
片を切 出し、PBS 透析し、濃度を定量し、抗原タン ク質 した BtR175 抗血清
を得 た 、抗原タン ク質をJcl:ICRマウス6終齢雌個体 5回免疫し、全採血し、
41
用いた 抗血清 0.005%ア 化 ト ウ 終濃度 50%の セ ー を加え -
20℃ 保存し以後の実験 用いた BmABCC2抗血清 い 、マウス ウサ の両
方を作製した マウス血清 、BtR175 同様の方法 作製した ウサ 抗血清 、株
式会社 蛋白精製工業 外注し、作製さ たものを用いた
2.2.8 BmABCC2変異体発現細胞表面上の毒素の染色
Sf9上 発現させた
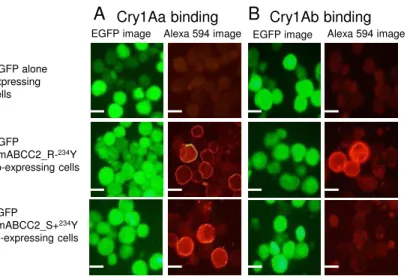
BmABCC2_R-234Y
BmABCC2_S+234YのCry1Aa, Cry1Ab結合能
を 調 た 、 そ の 組 み 換 え 型 AcNPV を 感 染 さ せ た 細 胞 、
NoStickHydrophobic Microtubes (Scientific Specialties Inc., Lodi, USA) 回 し、PBS 細
胞を洗った そ 20 mM Cry1Aaもしく Cry1Ab 10分間室温 反応させ、4%PFA
10分間固定した 固定した細胞 、PBS 3回洗い抗Cry1Aaウサ 抗血清 16時
間 4℃ 反応させ、PBS 3 回洗った後 、Alexa 594-conjugated goat anti-rabbit IgG (Molecular Probes, Eugene, OR) 1時間室温 反応させPBS 3回洗った後、封入し、
2.6 記載した顕微鏡 観察した
2.2.9 ウエスタン ッテ ン
2.2.6 同様の手法 そ のAcNPVを感染させたSf9細胞 、1.5 ml ュー
回 し、PBS 3回洗った後、SDS-PAGE sample bufferを加え煮沸し、SDS-PAGEのサ
ン した SDS-PAGE の操作 、10%ポ アク ア を用い 分離し、
分離後の PVDF膜 Perkin Elmer Life Sciences 転写した タン クを転写した
PVDF 膜 、5%ス ク/TNT 1 時間、室温 振盪し が ッ ン し、
42
2.2.10 免疫染色
発現細胞上のBtR175-TBR BtR175-fullの免疫染色 以 の う 行った 2.6 同
様の手法 そ のAcNPVを感染させ培養した細胞 、MAS-GP type A Micro slide
glass上 播種した 4%PFA/PBS 10分間固定した後、抗BtR175マウス抗血清を添加
後、16時間、4℃ イン ュベートし、PBS 3回洗った 2次抗体 、
Alexa-555-conjugated-anti-mouse IgG (Molecular Probes) を用い、1時間室温 反応させた
後封入し、2.2.6 記載した蛍 顕微鏡 観察した
2.2.11 BtR175発現細胞表面上の毒素の検出
Cry1AaのBtR175 対す 結合能を調 た 、2.6 同様の手法 、BtR175を発現
させたSf9細胞をMAS-GP type A micro slide glass (Matsunami) 播種し、4%PFA 固 定した PBS 3回洗った後、10 nM のCry1Aa 毒素 16時間4℃ 反応させ、抗Cry1Aa ウサ 抗血清 1 時間反応させた PBS 3 回洗った後、Alexa 594-conjugated goat anti-rabbit IgG (Molecular Probes) 1時間室温 反応させ、封入し、2.2.6 記載した蛍
顕微鏡 観察した
2.2.12 SPRを用いたBtR175 Cry1A毒素の結合性状の解析
大腸菌 発現させたBtR175-TBRの封入体の精製、可溶化 、Hara et al. (2003) 同様
の方法 行った 精製したBtR175 pH 4.0の10 mM 酢酸アン ニウ 終濃度 50 µg/
ml う 希釈し、CM5センサー ッ (GE Healthcare) ア ン ッ ン 法
固相化した Cry1A毒素 そ 各70 µl う PBST 希釈し5濃度
用意し、BtR175-TBRを固相化した CM5 センサー ッ 120 秒間イン クト