生殖工学技術を利用した子ウシの性別制御に
関する研究
Studies on Producing Calves of Desired Sex Using
Reproduction Engineering Technologies
2016 年 1 月
目次 要約 ・・・・・ 1 略語一覧 ・・・・・ 5 第1章 緒論 1.1 わが国の畜産および酪農を取り巻く状況 ・・・・・ 6 1.2 わが国の胚移植技術の状況 ・・・・・ 6 1.3 ウシの雌雄産み分け技術 ・・・・・ 7 1.4 ウシ胚のガラス化保存 ・・・・・ 8 1.5 性選別精液を利用した雌雄産み分け ・・・・・ 9 第2章 緩慢凍結したウシバイオプシー胚の生存性と受胎性 2.1 緒言 ・・・・・11 2.2 材料および方法 ・・・・・13 2.3 結果 ・・・・・17 2.4 考察 ・・・・・18 2.5 要約 ・・・・・21 2.6 図表 ・・・・・22 第3章 ガラス化保存したウシ体外受精由来胚の加温条件が加温後の胚の生存 性に及ぼす影響 3.1 緒言 ・・・・・33 3.2 材料および方法 ・・・・・34 3.3 結果 ・・・・・36 3.4 考察 ・・・・・37 3.5 要約 ・・・・・39 3.6 図表 ・・・・・40
第4章 ガラス化保存したウシバイオプシー胚のストロー内希釈・直接移植後 の発生能 4.1 緒言 ・・・・・46 4.2 材料および方法 ・・・・・47 4.3 結果 ・・・・・49 4.4 考察 ・・・・・50 4.5 要約 ・・・・・52 4.6 図表 ・・・・・53 第5章 ガラス化保存後にストロー希釈したウシ性判別胚の生存性と受胎性 5.1 緒言 ・・・・・58 5.2 材料および方法 ・・・・・60 5.3 結果 ・・・・・62 5.4 考察 ・・・・・63 5.5 要約 ・・・・・66 5.6 図表 ・・・・・67 第6章 生体卵胞内卵子と性選別精液の体外受精によるウシ性判別胚の生産 6.1 緒言 ・・・・・76 6.2 材料および方法 ・・・・・77 6.3 結果 ・・・・・81 6.4 考察 ・・・・・82 6.5 要約 ・・・・・84 6.6 図表 ・・・・・85 第7章 総括ならびに結論 ・・・・・92 謝辞 ・・・・・97 引用文献 ・・・・・98
1 要約 性判別胚を用いて計画的な子ウシ生産を農家段階で実用化するためには、性 判別胚を超低温保存し、受胚牛の移植適期に合わせて移植できる技術体系の確 立が不可欠である。しかし、性判別胚はバイオプシー操作により胚の一部の細 胞が切除されるとともに透明帯が除去されている。このため、緩慢凍結により 保存された性判別胚において新鮮胚と同等の受胎性を確認した報告がある一方 で、インタクト(非バイオプシー)胚と比べて耐凍性や受胎性が低下すること も報告されている。ガラス化保存は、細胞外液に氷晶を形成させないために、 緩慢凍結に比べて高濃度の耐凍剤を含む溶液が用いられており、加温後は早期 に耐凍剤を希釈することが、耐凍剤の化学的毒性を抑制するために必要とされ ている。しかしながら、ガラス化保存ではダイレクトトランスファー法やワン ステップストロー法のように加温後直ちに受胚牛に移植した報告は少ない。 一方、ウシのX 精子または Y 精子を 90%以上の確率で含む性選別精液が市販 されているが、性選別精液はストロー1本あたりに充填された精子数が通常精 液に比べて少ないことなどから、人工授精後の受胎率や多排卵処理後に採取し た胚の移植可能胚率の改善が望まれている。また、ウシの生体内卵子吸引(以 下、OPU)-体外受精技術は、超音波画像診断装置を用いて生体卵巣から採取し た卵子を体外受精して胚を生産する技術であり、これまでの胚生産の中心であ った子宮灌流による体内受精由来胚の採取に代わる、あるいはこれを補完する 胚生産技術として開発が進められてきた。その中で、体内成熟卵子は、体外受 精後の発生能が体外成熟卵子に比べて高いが、体内成熟卵子と性選別精液を利 用した性判別胚の生産に関する報告や生産した性判別胚の受胎性に関する報告 は少なく、普及性や実用性に関する検討はほとんど行われていない。 本研究では、野外におけるウシの雌雄産み分け技術の開発を目的として、ウ シバイオプシー胚および性判別胚の緩慢凍結およびガラス化保存における保存 方法や融解、加温方法の影響を調査した。また、OPU 技術により採取した体内 成熟卵子と性選別精液の体外受精による性判別胚の生産について、胚の生産効 率と性判別胚を用いた子ウシ生産の状況を調査した。 第 2 章では、ウシバイオプシー胚の緩慢凍結方法が胚の生存性と受胎性に及 ぼす影響を調査した。過剰排卵処理を施した供胚牛から人工授精後7~8 日目に 採取した胚は金属刃でバイオプシーした後に凍結保存した。凍結方法は、10% グリセリンおよび20% 子ウシ血清(以下、CS)添加修正 PBS を凍結溶液とし て融解後に段階的に耐凍剤を希釈除去し胚の生存性を判定した後に受胚牛に移 植した 10G-SW、融解後にストロー内で 12.5%シュークロースおよび 20% CS 添加修正PBS と混合して一段階で耐凍剤を希釈除去し胚の生存性を判定した後
2
に受胚牛に移植した 10G-ISD、10% エチレングリコール(以下、EG)および
20% CS 添加修正 PBS を凍結溶液として、融解後に耐凍剤の除去操作を行わず
直ちに受胚牛に移植した10EG-DR、10% EG、0.1 mol/L シュークロースおよ
び20% CS 添加修正 PBS を凍結溶液として融解後に耐凍剤の除去操作を行わず 直ちに受胚牛に移植した10EG1S-DR、10% グリセリン 0.25 mol/L シュークロ ースおよび20% CS 添加修正 PBS を凍結溶液として融解後に耐凍剤の除去操作 を行わず直ちに受胚牛に移植した10G25S-DR、5%グリセリン 0.1 mol/L シュー クロースおよび20% CS 添加修正 PBS を凍結溶液として融解後に耐凍剤の除去 操作を行わず直ちに受胚牛に移植した5G1S-DR の 6 種類を用いてインタクト胚 とバイオプシー胚の生存性と受胎性を調査した。その結果、インタクト胚の受 胎率は凍結方法の間には有意差は認められなかったが、バイオプシー胚の受胎 率は10E1S-DR が新鮮胚、10G-SW および 10G25S-DR に比べて有意に低かっ た。また、いずれの方法においてもバイオプシー胚の受胎率はインタクト胚に 比べて低い成績となっており、バイオプシー胚に適した緩慢凍結方法について 更なる検討が必要と推察された。また、緩慢凍結したインタクト胚を一胚移植 により双子分娩が確認された事例から、ウシにおける自然発生的な一卵性双子 の発生様式を検討した。 第 3 章では、ガラス化保存したウシ胚をストロー内希釈後に直接移植する方 法を農家段階で実用化するために、加温温度、加温後の経過時間、希釈液の組 成が加温後の胚の生存性に及ぼす影響を検討した。媒精後7~8 日目に発生した
胚盤胞をガラス化溶液(以下、VSED: 25% EG+25% DMSO)で平衡した後にスト ロー内でガラス化保存し、加温後に胚の生存性と発育性を調査した。加温温度 は 20℃区と 30℃区において加温後の胚の生存率および透明帯脱出率に差は認 められなかった。また、加温後の保持時間 0 分に対して 5 分以降は有意に生存 率が低下し、0 分および 5 分に対して 40 分で有意に透明帯脱出率が低下した。 さらに、希釈液の組成により加温後の胚の生存率および透明帯脱出率に差は認 められなかった。これらのことから、ガラス化保存したウシ胚において、加温 温度の違いや希釈液の組成により生存性の低下は認められなかったが、移植終 了までの時間が延長した場合には胚の生存性が低下する可能性があることが示 された。 第 4 章では、ガラス化保存したウシバイオプシー胚を、農場内で加温し直ち に受胚牛に移植するために耐凍剤の希釈方法が受胎性に及ぼす影響を調査した。 人工授精後 7 日目の供胚牛から採取し性別判定のためにバイオプシーを施した 胚を、VSED 液で平衡した後にストロー内でガラス化保存した。加温後に耐凍 剤をストロー内で希釈しストローから取り出し胚の生存性を確認して移植した ISD 区の受胎率は 57.9%、耐凍剤をストロー内で希釈後に胚をストローから取
3 り出すことなく直ちに受胚牛に移植したDIR 区の受胎率は 62.5%であり、いず れもガラス化保存していないNV 区(56.3%)と有意差のない受胎率が得られた。 ストロー内希釈後のストロー内の EG 濃度は 6.9~7.7%、DMSO 濃度は 2.8~ 3.3%であった。これらのことから、ガラス化保存したウシバイオプシー胚は、 農場内での加温操作を行い直ちに受胚牛に移植することが可能であり、新鮮胚 移植と同等の受胎率の得られることが示唆された。 第 5 章では、ガラス化保存したウシ性判別胚を加温後直ちに受胚牛に移植す るために、耐凍剤のストロー内希釈が移植後の受胎性と子ウシ生産性に及ぼす 影響を調査した。過剰排卵処理を施した供胚牛から人工授精後7~8 日目に採取 した胚をマイクロブレードでバイオプシーし、雄特異的DNA の増幅の有無によ り性別を判定した。性判別胚は VSED 液で平衡し、希釈液と空気層を隔てて 0.25-mL ストロー内に充填し、液体窒素に投入した。加温後はストロー内でガ ラス化液と希釈液を混合し 3 分間保持した。評価区はストロー内希釈した胚の 生存性を判定した後に受胚牛に移植し、非評価区はストロー内希釈後の胚をス トローから取り出すことなく直ちに受胚牛に移植した。評価区および非評価区 の移植後の受胎率は 38.7%および 34.8%であり、両区の間に有意差は認められ なかった。しかし、評価区においてガラス化保存前の品質の劣る胚の生存性は 品質の高い胚に比べて有意ではないが低い傾向であった(P = 0.087)。受胎した 受胚牛の流産率は両区の間で有意差は認められなかった(13.9%、12.5%)。こ れらのことから、耐凍剤のストロー内希釈はガラス化保存したウシ性判別胚を 加温後直ちに受胚牛に移植するために有効な方法であることが確認された。 第6 章では、ホルスタイン種泌乳牛の卵巣より生体内卵子吸引により採取し た卵子と性選別精液を体外受精し、性判別胚の生産効率と性判別胚の移植によ る子ウシ生産状況を調査した。供試牛に多排卵処理(SOV 区)または卵胞発育 同調処理(FGT 区)を行った後に直径 5 mm 以上の卵胞から卵子を採取し、ホ ルスタイン種雄牛の性選別精液を用いて媒精し、胚盤胞期胚以降への発生を調 査した。また、媒精後7 および 8 日目の胚盤胞期胚および脱出途中胚盤胞期胚 を発情周期の同調した受胚牛に移植した。SOV 区および FGT 区における採取 卵子数に有意差は認められなかった。SOV 区では採取卵子のうち 62.1%が成熟 卵子であったが、FGT 区の採取卵子は全て未成熟卵子であった。媒精後 7~9 日の胚盤胞期胚率は試験区間に有意差は認められなかったが、供卵牛1頭当た りの胚盤胞期胚数はSOV 区が FGT 区に比べて有意に多かった(P < 0.05)。ま た、性判別胚の移植後の受胎率、在胎日数および産子の生時体重は試験区間に 有意差は認められず、産子の95.0%が雌であった。このことから、多排卵処理 後のホルスタイン種泌乳牛から採取した卵子と性選別精液を体外受精すること
4 で多数の性判別胚を得ることが可能であり、後継牛の計画的生産に利用できる ことが示唆される。 本研究の結果より、ウシバイオプシー胚および性判別胚の超低温保存におい て、ガラス化保存は新鮮胚移植と同等の実用的な受胎率が得られることが明ら かとなり、ガラス化保存した胚のストロー内希釈は超低温保存した性判別胚を 農家の庭先で融解し、一般的な胚移植と同様の簡単な操作により受胚牛への移 植を可能とすることを示した。さらに、ストロー内希釈後の胚の生存性および 受胎性に影響を及ぼす要因を確認したことにより、この技術の野外普及を進め るにあたっての注意点や技術普及のための課題を提示した。また、SOV 処理後 に生体内卵子吸引により採取した体内成熟卵子と性選別精液を体外受精する新 しい性判別胚の生産システムが後継牛生産に有効な手法であることを明らかに した。 本研究で実施したウシ性判別胚の超低温保存方法および性選別精液を利用し た性判別胚の生産に関する研究成果は、酪農経営における計画的な後継牛生産 の実現と今後の家畜繁殖技術の発展に寄与するものと考えられる。
5 略語一覧
BSA: bovine serum albumin ウシ血清アルブミン
CIDR: controlled intravaginal drug releasing device
腟内留置型黄体ホルモン製剤
CS: calf serum 子ウシ血清
DMSO: dimethyl sulfoxide ジメチルスルホキシド
EG: ethylene glycol エチレングリコール
FGT: follicle growth treatment 卵胞発育同調処理
FSH: follicle stimulating hormone 卵胞刺激ホルモン
GnRH: gonadotropin releasing hormone
性腺刺激ホルモン放出ホルモン
h: hour(s) 時間
LAMP: loop-mediated isothermal amplification ランプ法
LN: liquid nitrogen 液体窒素
min: minute(s) 分
NBS: new born calf serum 新生子ウシ血清
OPS: open-pulled straw オープンプルドストロー法
OPU: ovum pick-up 生体内卵子吸引
PCR: polymerase chain reaction
PG: prostaglandin F2α プロスタグランジン F2α 製剤
S: sucrose シュークロース
SOV: superovulation 多排卵処理 VSED: 25% EG (v/v), 25% DMSO (v/v) and 0.3% BSA (w/v) dissolved in PBS
6 第1章 緒論 わが国の畜産および酪農を取り巻く状況 わが国における畜産業は、平成24 年度の農業総産出額(8.5 兆円)の 30.4% (2.6 兆円)を占めており、中でも酪農業の産出額は農業全体の 9.1%(7.7 億円) を占めている(農林水産省大臣官房統計部、2013)。また、生産される牛乳およ び乳製品は国内消費量の約 65%(761 万トン)を占め(農林水産省大臣官房食 料安全保障課、2013)、国民の食生活を支える重要な産業として位置づけられて いる。 近年、バイオエタノール需要の増加や中国をはじめとする新興国の需要増大、 さらに生産国における作柄や投機資金の動きなどを受け、輸入飼料価格は高止 まりの傾向が続いており、今後さらに高騰することが懸念されている。そのよ うな情勢の中でも、穀物相場に翻弄されない足腰の強い畜産経営を目指すため に、国は「新たな食料・農業・農村基本計画(平成22 年 3 月)」の中で、自給 粗飼料、エコフィード等の利用により、平成32 年度までに飼料自給率を 38%ま で引き上げることを目標とした取り組みを進めている。一方、生産コストの低 減による酪農経営の安定化と牛乳・乳製品の安定供給を図るためには、乳牛の 生涯生産性の向上に努めながら、能力と体型の改良を着実に進める必要があり、 新たな改良手法や繁殖技術の活用に努めるとともに、基本的な繁殖・飼養管理 技術の高位平準化の取り組みが求められている。 一方、国内の酪農における最近の後継牛生産の状況は、平成25 年における乳 用牛に対する黒毛和種精液の交配の割合は、全国平均では 30.1%であるが、北 海道を除く都府県では 43.5%と特に高い状況である(社団法人日本家畜人工授 精師協会、2014)。このことは、都府県の酪農経営において一時的な副産物収入 を目的とした交雑種子牛の生産が増加し、更新のための育成牛の確保を北海道 からの導入に依存する傾向がより進んでいることを示している。その結果、初 妊牛価格は一層高騰し、酪農経営をさらに圧迫する状況が続いている。このこ とから、酪農経営における一層の生産性の向上を図るための方策のひとつとし て、後継牛の計画的な確保と後継牛確保に要する費用の節減を可能にする技術 体系の確立が求められる。 わが国のウシ胚移植技術の状況 ウシの胚移植技術は、優良後継牛の生産や黒毛和種肥育素牛の生産を実現す る技術として国内の農家に広く普及しており、近年では移植後の受胎率は全国 平均で50%に近づき、年間 20,000 頭を超える子ウシが生産されている(農林水 産省生産局畜産部 2012)。しかしながら、胚移植により生産される子ウシは全
7 国で生産される子ウシ総数のわずか1%程度を占めるに過ぎず、技術利用の一層 の拡大が求められている。 ウシの胚移植技術は、供胚牛の過剰排卵処理、人工授精、採胚、胚の保存、 受胚牛への移植の一連の技術から構成されている。現在、国内で実施される胚 移植のおよそ 8 割は凍結胚の移植で占められており、緩慢凍結した胚を受胚牛 の移植適期に合わせて農場内で融解し直ちに移植する方法が一般的である。す なわち、10% (V/V)程度のグリセリンやエチレングリコールと 0.1~0.3 mol/L 程度のシュークロースを含む溶液で平衡した胚を、プログラムフリーザーを用 いて毎分 0.3~0.5℃程度で緩慢に冷却した後に液体窒素中に投入して超低温保 存し、融解は液体窒素中から取り出したストローを空気中で10 秒程度保持した 後に、30℃程度の温水に浸して行い、耐凍剤の除去操作は行わずに、直ちに移 植器に装填して頚管経由法で受胚牛に移植する方法である。このような超低温 保存胚の利用方法はダイレクトトランスファー法とよばれ、人工授精と同様の シンプルな融解操作および移植操作で利用できることから、受胚牛の発情周期 に合わせた移植や胚を供給するセンターから遠隔地で飼養される受胚牛への移 植に広く利用されている。 ウシの雌雄産み分け技術 胚移植の利用による付加価値を高めるために、2胚移植による双子生産、体 外受精を利用した肥育素牛の生産、性判別胚を利用した雌雄産み分け、一卵性 双子やクローン牛の生産など様々な技術が開発されている。なかでも産子の雌 雄産み分け技術は、確実な後継牛生産を可能にするとともに、後継牛生産に仕 向けない雌ウシから交雑種生産や胚移植による黒毛和種生産を行うことにより 子ウシ販売による収入確保を可能にする点でも酪農経営の収益性向上を可能に するものと考えられる。 家畜をはじめとする哺乳動物の性別は、受精に至った精子の性染色体によっ て決定する。すなわち、Y 染色体を持つ精子と X 染色体を持つ精子のどちらが 卵子と受精するかにより性別が決定され、あらかじめ性別を判定した胚を移植 に用いることで胚移植を利用した雌雄産み分けが可能となっている。これまで に、家畜胚の性別判定は、性染色体検査による方法(牛島ら 1989)、雄特異的 抗原を利用した方法(内海 1989)、X 染色体にリンクした酵素活性を利用した 方法(Iwasaki ら 1983)や胚の発育速度差や形態的品質を指標とした方法(富 永ら 1996)が試みられてきたが、性別判定までの所要時間の短さや判定精度の 高さなどから、現在ではバイオプシーした胚細胞から雄特異的DNA を検出し性 別を判定する方法が広く普及している(Bredback ら 1994;Itagaki ら 1996; Hirayama ら 2004)。
8 性判別胚を用いて計画的な子ウシ生産を農家段階で実用化するためには、性 判別胚を超低温保存し、受胚牛の移植適期に合わせて融解し移植できる技術体 系の確立が不可欠である。しかし、性判別胚はバイオプシー操作により胚細胞 の一部が切除されるとともに透明帯が除去されているため、緩慢凍結により保 存された性判別胚において新鮮胚と同等の受胎性を確認した報告(沼辺ら1995; 的場ら 2006)がある一方で、インタクト(非バイオプシー)胚と比べて耐凍性 や受胎性が低下する(Thibier と Nibart 1995; Hasler ら 2002; 富永 2005)こと も報告されている。しかしながら、性判別胚をガラス化保存した場合には新鮮 胚に近い受胎率の得られることが報告されている(Vajta ら 1997; Agca ら 1998; Tominaga 2004)。 ウシ胚のガラス化保存 ガラス化保存は、高濃度の耐凍剤を含む溶液に平衡した胚を極めて急速に冷 却することにより細胞内外に氷晶形成を伴わない超低温保存法であり、緩慢凍 結と比較して、植氷やプログラムフリーザーを用いた冷却が不要なばかりでな く、適切な条件で処理されれば、胚の生存性低下が極めて少ないことから、性 判別胚、体外生産胚、核移植胚等の耐凍性の低い胚や卵子の超低温保存に利用 されている(Kasai 1997; Tominaga 2004)。 哺乳動物胚のガラス化保存はRall と Fahy (1985)によりマウス 8 細胞期胚で の成功例が報告されて以来、さまざまな動物種の胚に対して耐凍剤の組成、冷 却方法、器具、加温後の希釈操作の検討が行われ、実験動物ではマウス、ラッ ト、ウサギ、家畜ではヒツジ、ブタ、ウマにおいて、胚のガラス化保存の成功 例が報告され(Kasai 1997; Kuwayama 2005)、ウシ胚では Massip ら (1986) が成功例を報告し、国内においても堂地 ら(1990)が同様の方法で成功例を報 告している。 ガラス化保存では、細胞外液に氷晶を形成させないために、緩慢凍結に比べ て高濃度の耐凍剤を含む溶液が用いられており、このため加温後は低濃度の耐 凍剤や糖類を添加した溶液に胚を移し替えて、できるだけ早期に耐凍剤を希釈 することが、耐凍剤の化学的毒性を抑制するために必要とされている(Kasai 1997)。これまでの多くの報告では、加温後にストローやシャーレ内で耐凍剤を 段階的に希釈し、胚の生存性を確認した後に、移植用ストローに再度胚を充填 して移植に用いている(堂地 ら 1990; Ishimori ら 1993; Massip ら 1987)。 しかし、この手法では実験室での胚の操作が必要であり、受胚牛が多数の農家 に分散して飼養されている国内の状況では、胚移植の実施が制限されることが 懸念される。 一方、緩慢凍結ではストロー内で耐凍剤を希釈するワンステップストロー法
9
(Leibo 1984; 鈴木ら 1984)や耐凍剤の希釈操作を行わないダイレクトトランス ファー法(Dochi ら 1998; Massip ら 1987; Voelkel と Hu 1992)は、融解後に 胚をストローから取り出すことなく直ちに受胚牛に移植するため、人工授精と 同様に農家の庭先で胚を融解し、効率的に胚移植を実施することが可能である。 しかしながら、ガラス化保存ではワンステップストロー法やダイレクトトラン スファー法のように加温後直ちに受胚牛に移植した報告は少ない(Leeun ら 1997; Saha ら 1996)。 これらのことから、ウシの雌雄産み分け技術を農家段階で普及するためには、 超低温保存された性判別胚を農家の庭先で融解し、一般的な胚移植と同様の簡 単な操作で受胚牛へ移植できる方法(Dochi ら 1998; Voelkel と Hu 1992; Massip ら 1987)の確立が望まれる。 性選別精液を利用した雌雄産み分け 家畜のX 精子および Y 精子の分別はさまざまな方法で試みられてきたが実用 的な技術は得られなかった。近年、フローサイトメーターとセルソーターを利 用して精子頭部のDNA 量の違いによりウシの X 精子および Y 精子を識別し分 取することが可能となった。すでに、国内においても性選別された凍結精液の ストローが市販され、人工授精(Hayakawa ら 2009)、多排卵処理による胚採 取(Hayakawa ら 2009)、体外受精および顕微授精(Hamano 2007)による胚 生産に利用可能なことが報告されている。しかしながら、非選別精液に比べて ストロー内に封入された精子数が少ないこと、生存性の低下や生存時間の短縮 により人工授精後の受胎率が低いこと(Andersson ら 2006; DeJarnett ら 2008; Schenk ら 2009; DeJarnette ら 2011)や、多排卵処理後に採取した胚の移植可 能胚率が低いこと(Hayakawa ら 2009; Peippo ら 2009; Larson ら 2010)など が指摘され、効果的な利用方法は確立されていない。 ウシにおける生体内卵子吸引(以下、OPU)技術は、超音波画像診断装置と 腟内挿入型探触子を用いてウシの生体卵巣から排卵前の卵子を採取し、体外受 精後に移植可能胚を生産する技術である(Pieters ら 1988)。従来の多排卵処理 後に子宮灌流により胚を生産する方法を補完する胚生産技術として利用可能な ことが報告されている。一方、繁殖障害牛(坂口ら1995)、妊娠牛(Imai ら 2006)、 肥育牛(大谷ら2005)、若齢牛(今井と田川 2006)など、従来の方法では胚生 産の対象とすることができない供卵牛おいても移植可能胚の生産が報告されて いる。また、FSH 投与による卵胞刺激処置(今井ら 2014)や体内成熟卵子の採
取(van de Leemput ら 1999; Dieleman ら 2002)などにより卵子の品質を向上 することで移植可能胚の生産が高まることが報告されている。これらのことか
10 選別精液を用いて胚生産の効率を改善する可能性があり、OPU 技術の利用によ り性選別精液を利用した新しい性判別胚システムを構築することが期待される。 本論文では、第 2 章において国内において広く普及している緩慢凍結を用い た際のウシバイオプシー胚の生存性と受胎性を明らかにし、第 3 章ではガラス 化保存したウシ胚を加温しストロー内で耐凍剤を希釈する処理における加温温 度、経過時間および希釈液の組成等の加温条件が胚の生存性に及ぼす影響を検 討し、第 4 章ではガラス化保存したウシバイオプシー胚を農場内で加温し直ち に受胚牛に移植するための耐凍剤の希釈方法を検討するとともにガラス化保存 胚と新鮮胚の受胎率を比較検討し、さらに第 5 章ではガラス化保存したウシ性 判別胚における耐凍剤のストロー内希釈が移植後の受胎性や子ウシ生産性に及 ぼす影響を明らかにするとともに性判別胚を用いたウシの雌雄産み分け技術の 野外普及の可能性を検討した。加えて、第6 章では OPU 技術により採取した体 内成熟卵子と性選別精液の体外受精による新しい性判別胚生産システムについ て、性判別胚の生産効率と性判別胚を用いた子ウシ生産の状況を明らかにした。
11 第 2 章 緩慢凍結したウシバイオプシー胚の生存性と受胎性 2.1 緒言 家畜の性別をコントロールし、計画的に雌雄を産み分けることができれば、 家畜の改良増殖の効率化と経営の安定化に対する効果は大きいと考えられる。 ウシの雌雄産み分け技術は胚の一部の細胞を切除し、性染色体検査や Y 染色体 特異的 DNA の検出などにより性別を判定し、残された胚(性判別胚)を移植に 用いることにより可能となっており(牛島ら 1991; 渡辺ら 1992; Bredbacka ら 1994; Hirayama ら 2004)、性判別胚を利用したウシの雌雄産み分けの農場段階で
の実用化が進められている(Thiber と Nibart 1995; 吉羽ら 1996; Hasler ら 2002)。
ウシの雌雄産み分け技術の野外利用をより一層推進するためには、胚の性別判 定の精度が高いこと、性別判定した胚の生存性や受胎性が低下しないことが求 められる。 性判別胚を用いて計画的な子ウシ生産を農家段階で実用化するためには、超 低温保存した性判別胚を受胚牛の移植適期に合わせて融解し移植する技術体系 の確立が不可欠である。一般に、胚移植の受胎率は新鮮胚に比べて凍結胚が低 いことが知られている(農林水産省生産局畜産部 2012)。胚の凍結保存におい て、グリセリンやエチレングリコールなどの耐凍剤を含む溶液に平衡した胚は プログラムフリーザーを利用して毎分 0.3~0.5℃程度の速度で緩慢に冷却され、 液体窒素中に保存される方法が用いられている(Dochi ら 1998; Massip ら 1987; Voelkel と Hu 1992)。性判別胚は、顕微操作により透明帯が切開され、 性別判定に必要な検査用細胞を切除されている。このため、胚を構成する細胞 数が減少するとともにバイオプシー操作による物理的な障害を受けている可能 性がある。さらに、凍結融解の過程で胚細胞が耐凍剤溶液や氷晶に直接曝され るなどの外的な感作を受けやすい状況にあると考えられる。これまで、性判別 胚やバイオプシー胚を寒天に包埋する方法(志賀ら 1991)や空透明帯へ再収納 する方法(Takeda ら 1987; Ushijima 2005)、数時間の修復培養(後藤ら 1995) や細胞賦活作用を有する培養液により形態を回復させる方法(富永 1990)など の処置後に移植や凍結保存する方法が試みられたが、実用的な手法は構築され ていない。 また、凍結保存された性判別胚において新鮮胚と同等の受胎性を確認した報 告(沼辺ら 1995; 的場ら 2006)がある一方で、バイオプシーを施していない胚 (以下、インタクト胚)と比べて耐凍性や受胎性が低下することも報告されて いる(Thibier と Nibart 1995; Hasler ら 2002 ; 富永 2005)。さらに、バイオプシ ー胚の形態的品質が凍結融解後の生存性や移植後の受胎性に影響を及ぼすこと
12
が報告されている(山中ら 1995; 牛島ら 1995)。
本研究の目的は、ウシバイオプシー胚の緩慢凍結方法が融解後の生存性や移 植後の受胎性に及ぼす影響を明らかにすることである。
13 2.2 材料および方法
2.2.1 供試材料
神奈川県畜産技術センターで飼養するホルスタイン種経産牛および黒毛和種 経産牛から胚を採取した。供胚牛は、発情周期の任意の時期に腟内留置型黄体 ホルモン製剤(EAZI-BREED, Inter Ag, New Zealand, 以下 CIDR)を留置し、7 日 目から卵胞刺激ホルモン(アントリン R10、川崎三鷹製薬、東京、以下 FSH) を、朝夕 2 回、3 日間、漸減投与し、過剰排卵処理を施した。FSH 投与量は、ホ ルスタイン種では合計 36 AU(7、7、6、6、5、5 AU)、黒毛和種では合計 20 AU (5、5、3、3、2、2 AU)とした。FSH 投与開始後 3 日目の朝に PGF2α類縁体 (エストラメイト、住友製薬、大阪、以下 PG)0.75 mg を投与し、CIDR を除去 して発情を誘起した。発情確認後に人工授精を 2 回行った。胚の採取は、発情 後 7 日目にバルーンカテーテルを用いて子宮頚管経由で行い、後期桑実胚から 拡張胚盤胞のコード 1 および2の品質(Robert と Nelson 1998)の胚を供試した。 また、一部は体外受精由来胚を供試した。と畜場で採取したウシ卵巣を実験 室に持ち帰り、18 G 注射針を付けた 5 mL 注射筒で小卵胞を吸引して未成熟卵子 を採取した。卵子は 5%子ウシ血清(以下、CS)を添加した TCM199(GIBCO、 12340)で 22 時間の成熟培養の後に、最終濃度 500 万/mL に調整した精子ドロッ プに導入して体外受精した。体外受精後 6 時間目に卵子を回収し、5% CS 添加 TCM199 中で培養し、体外受精後 7~8 日目に胚盤胞に発育した胚を供試した。 2.2.2 バイオプシー操作 供試胚は、0.2 mol/L シュークロース添加 PBS 中に胚を保持し、シャーレ底に 胚を付着させた後にバイオプシーした(山中ら 1995)。バイオプシー操作は倒 立顕微鏡下で行い、マイクロマニピュレーターに接続した金属刃(FEATHER BIO-CUTNo.715)により胚容積の 10~20%を切断してバイオプシー胚を作製し た。なお、初期胚盤胞以降の発育の胚では、内部細胞塊を避け栄養膜細胞のみ を切断した。 2.2.3 バイオプシー胚の修復培養 バイオプシー胚の修復培養は、5% CS 添加 TCM199 ドロップ(50 μL)中で、 38.5℃、5% CO2、95%空気、湿度飽和の条件で卵丘細胞と共培養、または 38.5℃、 5% CO2、5% O2、90% N2の条件で 3~5 時間または 16~20 時間行った。 2.2.4 胚の凍結保存および融解 バイオプシーを行っていないインタクト胚および修復培養後のバイオプシー
14 胚を次の方法で凍結保存し融解した。 1)10%グリセリン-段階希釈(以下、10G-SW) 10%グリセリンおよび 20% CS 添加修正 PBS を凍結溶液として胚を室温で 30 分間平衡し、図 2.1 に示すとおり 12.5%(W/V)シュークロースおよび 20% CS 添加修正 PBS 、20%CS 添加修正 PBS とともに 0.25 mL ストローへ充填した。 胚を充填したストローはプログラムフリーザー(プラナー、R-204)の冷却槽に 投入し、室温から1℃/分で-7℃まで冷却し、2 分後に植氷、さらに 8 分間保持し た後に、-30℃まで 0.3℃/分で冷却し、液体窒素中へ投入した。 融解はストローを空気中で 10 秒間保持後、35℃温水中で氷晶が消えるまで保 持して行い、ストロー内の 10%グリセリンおよび 20%CS 添加修正 PBS 層のみを シャーレ内に押し出し、胚を 6%、3%、0%グリセリンおよび 0.3 mol/L シューク ロース添加した修正 PBS にそれぞれ 5 分間浸せきし、20% CS 添加修正 PBS 中 で洗浄して耐凍剤を段階的に希釈除去した(図 2.2)。 2)10%グリセリン-ストロー内一段階希釈(10G-ISD) 10%グリセリンおよび 20% CS 添加修正 PBS を凍結溶液として胚を室温で 30 分間平衡し、図 2.1 に示すとおり 12.5%(W/V)シュークロースおよび 20% CS 添加修正 PBS 、20% CS 添加修正 PBS とともに 0.25 mL ストローへ充填した。 胚を充填したストローは 10G-SW と同様の手順で冷却した。 融解はストローを空気中で 10 秒間保持後、35℃温水中で氷晶が消えるまで保 持して行い、図 2.3 に示すとおりストロー内容液を混合し、ストローを垂直に 10 分間保持した後に、胚をシャーレ内に押し出し、20% CS 添加修正 PBS 中で 洗浄して耐凍剤を一段階で希釈除去した(鈴木ら 1984 )。 3)10%エチレングリコール-直接移植(10E-DR) 10%エチレングリコールおよび 20% CS 添加修正 PBS を凍結溶液として、バイ オプシー胚を室温下で 15 分間平衡し、0.25 mL ストローへ充填した(図 2.4)。 冷却は-7℃に保持したプログラムフリーザーにストローを投入し、2 分後に植氷、 8 分間保持後、-30℃まで 0.3℃/分で冷却し、液体窒素中に保存した。融解はスト ローを空気中で 10 秒間保持後、30℃温水中で氷晶が消えるまで保持して行った (Dochi ら 1998)。 4)10%エチレングリコール 0.1 mol/L シュークロース-直接移植(10E1S-DR) 10%エチレングリコール 0.1 mol/L シュークロースおよび 20% CS 添加修正 PBS を凍結溶液として、バイオプシー胚を室温下で 15 分間平衡し、0.25 mL ス トローへ充填した(図 2.4)。冷却は-7℃に保持したプログラムフリーザーにスト ローを投入し、2 分後に植氷、8 分間保持後、-30℃まで 0.3℃/分で冷却し、液体 窒素中に保存した。融解はストローを空気中で 10 秒間保持後、30℃温水中で氷 晶が消えるまで保持して行った。
15 5)10%グリセリン 0.25 mol/L シュークロース-直接移植(10G25S-DR) 10%グリセリン 20% CS 添加 m199 にバイオプシー胚を室温で 10 分間平衡し、 10%グリセリン 0.25 mol/L シュークロースおよび 20%CS 添加 m-199 に移して直 ちに 0.25 mL ストローへ充填した。ストロー内は胚を含む 10%グリセリン 0.25 mol/L シュークロースおよび 20% CS 添加 m199 を挟んで 0.25 mol/L シュークロ ース 20% CS 添加 m-199 を充填した(図 2.5)。冷却は- 6℃に保持したプログラ ムフリーザーにストローを投入し、1 分後に植氷、さらに 9 分間保持した後に、 -25℃まで 0.3℃/分で冷却し、-25℃で 5 分間保持した後に液体窒素中へ投入した。 融解はストローを空気中で 10 秒間保持後、30℃温水中で氷晶が消えるまで保持 して行った。 6)5%グリセリン 0.1 mol/L シュークロース-直接移植(5G1S-DR) 5%グリセリン 20% CS 添加 m-199 にバイオプシー胚を室温で 10 分間平衡し、 5%グリセリン 0.1 mol/L シュークロースおよび 20% CS 添加 m-199 に移して直ち に 0.25 mL ストローへ充填した。ストロー内は胚を含む 5%グリセリン 0.1 mol/L シュークロースおよび 20% CS 添加 m-199 を挟んで 0.1 mol/L シュークロース 20%CS 添加 m-199 を充填した(図 2.5)。冷却は- 4℃に保持したプログラムフリ ーザーにストローを投入し、1 分後に植氷、さらに 9 分間保持した後に、-25℃ まで 0.3℃ /分で冷却し、-25℃で 5 分間保持した後に液体窒素中へ投入した。融 解はストローを空気中で 10 秒間保持後、30℃温水中で氷晶が消えるまで保持し て行った。 2.2.5 胚の生存性判定 10G-SW および 10G-ISD の融解後の胚は、5% CS 添加 TCM199 で卵丘細胞と の共培養を行い、融解後1時間に形態を観察し、生存性判定を行った。 10G25S-DR および 5G1S-DR で凍結保存した体外受精由来胚は、融解後に 20% ウシ胎子血清および 0.1 mmol/L メルカプトエタノールを添加した TCM199 (GIBCO、12340)で培養し、48 時間後の胞胚腔の形態を観察し、生存性判定を 行った。 2.2.6 胚の移植 10G-SW および 10G-ISD では融解後 1 時間の形態観察により生存と判定され た胚を 20% CS 添加修正 PBS とともに 0.25 mL ストローに充填して移植に用い た。10E-DR、10E1S-DR、10G25S-DR および 5G1S-DR では融解後に耐凍剤の除 去操作を行わず、ただちに受胚牛へ移植した。 受胚牛はホルスタイン種未経産牛および経産牛を用い、発情後 7~8 日目に黄 体側子宮角へ頸管経由法で移植を行った。妊娠鑑定は胚移植後 35 日以降に直腸
16 検査法で行った。 2.2.7 統計処理 統計処理は、コンピューターソフト SPSS(SPSS 11.5J, SPSS Inc., 東京)を用 い、生存胚率および受胎率について Pearson のχ二乗検定または Fisher の直接確 率計算法を用いて危険率 5%未満を有意な差が有りとした。
17 2.3 結果 10G-SW および 10G-ISD で凍結保存したインタクト胚およびバイオプシー胚 の生存性を表 2.1 に示す。インタクト胚の融解後の生存胚率は 10G-SW が 90.0%、 10G-ISD が 68.0%で、バイオプシー胚の生存胚率は 10G-SW が 80.0%、10G-ISD が 58.3%であり、どちらの胚も凍結方法の間に有意差は認められなかった(P > 0.05)。 10G25S-DR および 5G1S-DR で凍結保存したインタクト胚およびバイオプシー 胚の融解後の生存性を表 2.2 に示した。インタクト胚の生存胚率は 10G25S-DR が 83.9%、5G1S-DR が 86.7%であり、バイオプシー胚の生存胚率は 10G25S-DR が 100.0%、5G1S-DR が 71.4%であり、いずれも有意差は認められなかった(P > 0.05)。 インタクト胚およびバイオプシー胚の凍結方法が受胎性に及ぼす影響を表 2.3 に示す。インタクト胚の移植後の受胎率は 38.9~63.6%であり、凍結方法の間に 有意差は認められなかった(P > 0.05)。バイオプシー胚の移植後の受胎率は 6.7 ~46.2%であり、10E1S-DR の受胎率は新鮮胚、10G-SW および 10G25S-DR に比 べて有意に低かった(P < 0.05)。また、10E1S-DR においてバイオプシー胚の受 胎率はインタクト胚に比べて有意に低かった(P < 0.05)。
18 2.4 考察 本研究は、6 種類の凍結方法についてインタクト胚とバイオプシー胚における 融解後の生存性や移植後の受胎性を調査したものである。その結果、インタク ト胚の受胎率は凍結方法の間には有意差は認められなかったが、バイオプシー 胚では 10E1S-DR が、新鮮胚、10G-SW および 10G25S-DR に比べて有意に受胎 率が低かった。 性判別胚は、顕微操作により透明帯が切開され、性別判定に必要な検査用細 胞を切除されている。このため、胚を構成する細胞数が減少するとともにバイ オプシー操作による物理的な障害を受けているものと考えられる。さらに、凍 結融解の過程で胚細胞が耐凍剤溶液や氷晶に直接曝されるなどの外的な感作を 受けやすい状況にあると考えられる。 バイオプシー胚の凍結胚移植において、沼辺ら(1995)、後藤ら(1995)、吉羽ら (1996)はインタクト胚に比べて受胎率が低いことを種々の凍結方法で報告して いる。また、富永(2005)は 1.8 mol/L エチレングリコールに 0.1 mol/L トレハロー スまたはシュークロースを添加した溶液で凍結保存したバイオプシー胚につい て大規模な移植試験を国内 21 県で行ったところ、実用的な受胎率は得られなか ったことを報告している。本研究では富永(2005)の報告と同様の凍結方法で ある 10E1S-DR において、インタクト胚では他の凍結方法と差のない受胎率が得 られたが、バイオプシー胚では新鮮胚、10G-SW および 10G25S-DR に比べて有 意に受胎率が低く、富永(2005)の報告と同様の結果であった。 一方、的場ら(2006)は 1.5 mol/L エチレングリコールと 0.1 mol/L シュークロー スを添加した溶液を用いて、小出ら(2009)は 10%グリセリンと 0.25 mol/L シュー クロースを含む溶液を用いて緩慢凍結したバイオプシー胚の直接移植において、 新鮮バイオプシー胚あるいは凍結インタクト胚と同等の受胎率が得られること を報告している。本研究では、小出ら(2009)の報告に準じた 10G25S-DR におい てインタクト胚、バイオプシー胚ともに有意な差ではないが他の凍結方法に比 べて融解後の生存率と移植後の受胎率が最も優れており、バイオプシー胚では 小出ら(2009)の報告に匹敵する受胎率が得られた。 Tominaga ら(2007)はバイオプシーした体外受精由来胚の凍結保存において、 5%グリセリン溶液に 0.05 mol/L または 0.1 mol/L シュークロースを添加すること で融解後に 80%以上の胚の生存を確認し、高田ら(2005)および Takada ら(2007) はインタクト胚において 5%および 6%グリセリン溶液を用いた直接移植でそれ ぞれ 45.1%および 46.7%の受胎率を得たことを報告している。本研究では Tominaga ら(2007)の報告を参考にした 5G1S-DR において、インタクト胚、バイ オプシー胚ともに有意ではないが受胎率は低く、Tominaga ら(2007)や高田ら
19 (2005)の報告を支持する成績は得られなかった。 Iwasaki ら(1994)は、凍結保存したバイオプシー胚の二重染色において内部細 胞塊の内部に死滅細胞が存在することを確認し、このことが受胎率低下の要因 となる可能性を報告している。本研究においては、10G-SW で凍結保存したバイ オプシー胚において融解後の形態的評価により生存性を確認した胚を移植に用 いたにも関わらず有意ではないがインタクト胚に比べて受胎率が低い結果とな った。これらのことから、凍結保存したバイオプシー胚においては凍結融解の 過程で形態的には判断できない傷害により胚の発育性の低下を引き起こしてい るものがあると考えられる。また、的場ら(2006)は 1.5 mol/L エチレングリコー ルと 0.1 mol/L シュークロースを添加した溶液を用いた緩慢凍結において発育ス テージの若い後期桑実胚から初期胚盤胞を用いることで、胚盤胞期胚以降の胚 に比べて高い生存性を得る可能性があることを報告している。本研究では移植 頭数が限られていることから、これらの点についての検討は行っていないが、 今後、大規模な移植試験において検証を行う必要があると考えられる。 加えて、バイオプシー胚の受胎率はバイオプシー操作の適否、すなわちバイ オプシー後の胚の品質により変動することが報告されている(牛島ら 1995)。本 研究では、現在、国内で広く利用されている金属刃(松本ら 1987)を利用して 胚のバイオプシー操作を行ったが、胚の品質低下を抑えるバイオプシー器具の 開発(上田ら 2009; 秋山ら 2010)やバイオプシー操作の改良(Vajta ら 1997; 橋 谷田ら 2007)についても検討する必要があると考えられる。 本研究の 10G-ISD で凍結保存したインタクト胚の1例において、図 2.7 に示す とおり融解後に透明帯内に2個の細胞塊が観察され、この胚を受胚牛に移植し たところ、移植後 261 日目に一卵性双子(♀♀)を分娩した(図 2.8)。ウシで は自然発生的な一卵性双子の発生は着床以後であり、胚盤胞の内部細胞塊が2 個の原始線条を形成した結果、2個体が生ずるとされている(Mclaren 1982; Hafenz 1992)。本例において観察された2個の細胞塊は、輪郭が明瞭であり、形 態的に凍結保存によって受けた傷害は少なく、供胚牛から採取時に既に2個の 胚盤胞が透明帯内で発生しており、融解後のシュークロース液を用いた耐凍剤 除去操作の過程でそれらが収縮し、2個の明瞭な細胞塊として観察されたもの と推察される。また、双子はぞれぞれ固有の絨毛膜と羊膜を持っていたことか ら一卵性双子の発生は発生初期に起こったものと考えられた(Renfree 1982)。こ れらのことから、透明帯脱出以前のウシ胚で一卵性双子の発生が起こる可能性 があることが示唆された。 これらのことから、本研究で用いた凍結方法の中では 10G25S-DR がバイオプ シー胚の凍結方法として最も適していると推察された。しかしながら、いずれ の凍結方法においてもバイオプシー胚の受胎率はインタクト胚に比べて 10~
20
20%程度低率であり、受胎率の向上を目指してバイオプシー胚に適した超低温保 存方法の開発がさらに必要と考えられる。
21 2.5 要約 ウシバイオプシー胚の緩慢凍結方法が胚の生存性と受胎性に及ぼす影響を調 査するために、過剰排卵処理を施した供胚牛から人工授精後7~8 日目に採取し た胚を金属刃でバイオプシーした後に凍結保存した。凍結方法は 10%グリセリ ンおよび 20% CS 添加修正 PBS を凍結溶液として融解後に段階的に耐凍剤を希 釈除去し胚の生存性を判定した後に受胚牛に移植した 10G-SW、融解後にストロ ー内で 12.5 % (W/V)シュークロースおよび 20% CS 添加修正 PBS と混合して一 段 階 で 耐凍 剤 を希 釈除 去 し 胚 の 生存 性 を 判定 し た後 に受 胚 牛に 移 植し た 10G-ISD、10%エチレングリコールおよび 20% CS 添加修正 PBS を凍結溶液とし て融解後に耐凍剤の除去操作を行わず直ちに受胚牛に移植した 10E-DR、10%エ チレングリコール 0.1 mol/L シュークロースおよび 20% CS 添加修正 PBS を凍結 溶 液 と して 融 解後 に耐 凍 剤の 除 去操 作 を 行わ ず 直ち に受 胚 牛に 移 植し た 10E1S-DR、10%グリセリン 0.25 mol/L シュークロースおよび 20% CS 添加修正 PBS を凍結溶液として融解後に耐凍剤の除去操作を行わず直ちに受胚牛に移植 した 10G25S-DR、5%グリセリン 0.1 mol/L シュークロースおよび 20% CS 添加修 正 PBS を凍結溶液として融解後に耐凍剤の除去操作を行わず直ちに受胚牛に移 植した 5G1S-DR の 6 種類を用いてインタクト胚とバイオプシー胚の生存性と受 胎性を調査した。 その結果、インタクト胚の受胎率は凍結方法の間には有意差は認められなか ったが(P > 0.05)、バイオプシー胚では 10E1S-DR が新鮮胚、10G-SW および 10G25S-DR に比べて有意に受胎率が低かった(P < 0.05)。また、いずれの方法 においてもバイオプシー胚の受胎率はインタクト胚に比べて低い成績となって おり、バイオプシー胚に適した緩慢凍結方法について更なる検討が必要と考察 された。また、緩慢凍結したインタクト胚の移植により双子分娩が確認された 事例から、ウシにおける自然発生的な一卵性双子の発生様式を考察した。
22 2.6 図表 PBS PBS 熱シール 綿栓 凍結液 希釈液 胚を含む凍結液 希釈液 図2.1 10G-SW および 10G-ISD のためのストロー内の構成 希釈液:12.5%シュークロースおよび 20% CS 添加修正 PBS 凍結液:10%グリセリンおよび 20% CS 添加修正 PBS PBS:20% CS 添加修正 PBS
23 6% 3% 0% 20%CS-PBS a b c d 図2.2 10G-SW の融解と耐凍剤除去 a ストローを空気中で 10 秒間保持 b ストロー内容が溶解するまで 35℃温水にストローを保持 c 胚を含む凍結液のみをシャーレ内に押し出す d 6%、3%、0%グリセリンおよび 0.3 mol/L シュークロース添加した修正 PBS に5 分間隔で胚を浸せきして耐凍剤を除去
24 a b c d 図2.3 10G-ISD の融解と耐凍剤除去 a ストローを空気中で 10 秒間保持 b 希釈液が溶解するまで 35℃温水にストローを垂直に保持 c ストロー熱シール部を弾いて希釈液と凍結液を混合し 10 分間保持 d ストロー内容をシャーレ内に押し出し生存性評価
25 熱シール 綿栓 凍結液 胚を含む凍結液 凍結液 図2.4 10E-DR および 10E1S-DR のためのストロー内の構成 凍結液 10E-DR:10%エチレングリコールおよび 20% CS 添加修正 PBS 10E1S-DR:10%エチレングリコール 0.1 mol/L シュークロースおよび 20% CS 添加修正PBS
26 熱シール 綿栓 凍結液 希釈液 胚を含む凍結液 希釈液 希釈液 図2.5 10G25S-DR および 5G1S-DR のためのストロー内の構成 10G25S 希釈液:0.25 mol/L シュークロースおよび 20% CS 添加 m199 凍結液:10%グリセリン 0.25 mol/L シュークロースおよび 20% CS 添加 m199 5G1S 希釈液:0.1 mol/L シュークロースおよび 20% CS 添加 m199 凍結液:5%グリセリン 0.1 mol/L シュークロースおよび 20% CS 添加 m199
27 図 2.6 10G25S-DR で凍結保存したウシバイオプシー胚の形態 左上:凍結前 右上:融解 4 時間後 左下:融解 24 時間後 右下:融解 48 時間後 スケールバー:100 µm
28
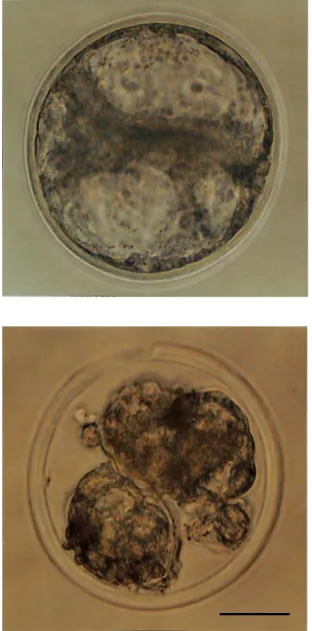
図 2.7 一胚移植後に双子分娩例が得られたウシ胚の形態 上:採取時
下:凍結融解時 スケールバー:50 µm
29
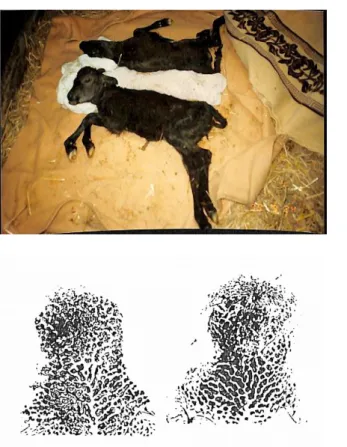
図 2.8 一胚移植後に誕生したウシ双子とその鼻紋 性別:♀、♀
30 表 2.1 ウシ胚の凍結融解後の生存性 融解胚数 生存胚数 生存胚率 融解胚数 生存胚数 生存胚率 a b a/b(%) c d c/d(%) 10G-SW 20 18 90.0 15 12 80.0 10G-ISD 75 51 68.0 12 7 58.3 凍結方法 インタクト胚 バイオプシー胚 10G-SW:10%グリセリンおよび 20% CS 添加修正 PBS で凍結保存し融解後は段 階的に耐凍剤を希釈 10G-ISD:10%グリセリンおよび 20% CS 添加修正 PBS で凍結保存し融解後はス トロー内で耐凍剤を一段階希釈 融解後の胚は、5% CS 添加 TCM199 で卵丘細胞と共培養し、融解後1時間に生 存性を判定
31 表 2.2 ウシ体外受精由来胚の凍結融解後の生存性 融解胚数 生存胚数 生存胚率 融解胚数 生存胚数 生存胚率 a b a/b(%) c d c/d(%) 10G25S-DR 31 26 83.9 12 12 100.0 5G1S-DR 15 13 86.7 14 10 71.4 凍結方法 インタクト胚 バイオプシー胚 10G25S-DR:10%グリセリン 0.25 mol/L シュークロースおよび 20% CS 添加修正 PBS で凍結保存 5G1S-DR:5%グリセリン 0.1 mol/L シュークロースおよび 20% CS 添加修正 PBS で凍結保存 融解後の胚は 0.1 mmol/L メルカプトエタノール添加 TCM199 で培養し、融解後 48 時間に生存性を判定した
32 表 2.3 ウシ胚の凍結融解後の受胎性 移植頭数 受胎頭数 受胎率 移植頭数 受胎頭数 受胎率 a b b/a(%) c d d/c(%) 新鮮胚 有 6 4 66.7 21 11 52.4 a 10G-SW 有 161 77 47.8 23 7 30.4 a 10G-ISD 有 51 22 43.1 - - -10E-DR 無 161 71 44.1 10 3 30.0 10E1S-DR 無 12 6 50.0 c 30 2 6.7 bd 10G25S-DR 無 22 14 63.6 13 6 46.2 a 5G1S-DR 無 18 7 38.9 12 3 25.0 凍結方法 生存 判定 インタクト胚 バイオプシー胚 10G-SW:10%グリセリンおよび 20% CS 添加修正 PBS で凍結保存し融解後は耐 凍剤をシャーレ内で段階的に希釈した 10G-ISD:10%グリセリンおよび 20% CS 添加修正 PBS で凍結保存し融解後は耐 凍剤をストロー内で一段階希釈した 10E-DR:10%エチレングリコールおよび 20% CS 添加修正 PBS で凍結保存 10E1S-DR:10%エチレングリコール 0.1 mol/L シュークロースおよび 20% CS 添 加修正 PBS で凍結保存 10G25S-DR:10%グリセリン 0.25 mol/L シュークロースおよび 20% CS 添加 m199 で凍結保存 5G1S-DR:5%グリセリン 0.1 mol/L シュークロースおよび 20% CS 添加修正 m199 で凍結保存 10G-SW と 10G-ISD は耐凍剤を希釈し胚の生存を確認した後に移植した 10E-DR、10E1S-DR、10G25S-DR、5G-1S は胚の生存性を確認せずに移植した 移植後 35 日に直腸検査により妊娠診断した ab 間、cd 間に有意差あり(P < 0.05)
33
第 3 章 ガラス化保存したウシ体外受精由来胚の加温条件が加温後の胚の生存
性に及ぼす影響 3.1 緒言
胚のガラス化保存は、細胞外液に氷晶形成を伴わないことから胚の生存性を 最も損なわない超低温保存方法であり(Rall と Fahy 1985; Kasai 2001)、緩慢 凍結では生存性や受胎性が低下する体外受精胚やバイオプシー胚の超低温保存 に 有 効 な こ と が 報 告 さ れ て い る( 濱 野 と 濱 脇 2001; 小 渕 ら 2001 ; Yotsushima ら 2004; 小田ら 2005; 的場ら 2006)。しかし、ガラス化保存は 緩慢凍結に比べて高濃度の耐凍剤を用いるため、ウシの胚移植においては加温 後の胚を低濃度の耐凍剤や糖類を添加した溶液に移し替えて耐凍剤を希釈し、 胚の生存性を確認した後に移植用ストローに再度充填して移植に用いられてい る。現在、農家段階で実施されるウシ胚移植の多くは超低温保存された胚を融 解 後 に 耐 凍 剤 の 除 去 操 作 を 行 わ ず に 直 接 移 植 す る 方 法(Massip ら 1987; Voelkel と Hu 1992; Dochi ら 1998)が用いられている。ガラス化保存したウ シ胚についても、農家段階での利用を推進するためには、高い受胎率を安定的 に得ることと同時に、農家の庭先で簡易な操作で加温と耐凍剤の除去を行い、 直ちに受胚牛に移植可能な手法を確立することが望まれる。 ウシガラス化保存胚の加温後の耐凍剤の希釈操作には、1.0 mol/L シュークロ
ース液(Massip ら 1987; 堂地ら 1990; Leewn ら 1997; Vajta ら 1995)
や0.5 mol/L シュークロース液(Ishimori ら 1993; Yotsushima ら 2004)が
用いられ、ストロー内またはシャーレ内で速やかにガラス化液と混合し耐凍剤 を希釈することで、高い生存率や発育率が得られることが報告されている。ま た、Kuwayama ら(1994)、Vajta ら(1995)、Saha ら(1996 )、川島ら(2000)、小 田ら(2005)はガラス化液をストロー内希釈し直接受胚牛に移植できることを報 告した。さらに、小渕ら(2001)、Akiyama ら(2010)はガラス化保存したバイオ プシー胚の耐凍剤をストロー内希釈することにより、新鮮胚と同等の受胎率を 得ることが可能であり、庭先融解に対応可能なことを報告した。 ガラス化保存は加温操作の適否により胚の生存性が大きく変動することが指 摘されており(Kasai 2001)、この技術を農家段階で実用化するためには、高い 受胎率を安定的に得ることができる加温操作の確立が必要である。しかし、ガ ラス化保存したウシ胚の直接移植を想定した加温操作が胚の生存性に及ぼす影 響を検討した報告は少ない。そこで、ガラス化保存したウシ胚の加温温度、加 温後の保持時間、希釈液の組成が加温後の胚の生存性に及ぼす影響を検討した。
34 3.2 材料および方法 3.2.1 卵子の採取 と畜場で採取したウシ卵巣を実験室に持ち帰り、18G 注射針をつけた 5 mL 注射筒で卵胞液を1% CS 添加ダルベッコ PBS 液で遠心管に吸引した。卵胞液 を実体顕微鏡下で検索し卵子を採取した。 3.2.2 体外受精由来胚の生産 卵子は5%CO2、95%空気、38.5℃の条件で 20~22 時間成熟培養した。成熟
培養液は、0.02 AU/mL FSH、1 μg/mL Estradiol-17、0.2 mmol/L ピルビン酸
およびウシ胎子血清(以下、FBS)添加 TCM199(GIBCO、12340)を用いた。 成熟培養後の卵子は、凍結精液を用いて媒精した。精子の受精能獲得誘起は、 精子洗浄液(機能性ペプチド研究所、IVF100)で 2 回遠心洗浄することで行い、 最終濃度500 万/mL の精子懸濁液に成熟培養後の卵子を導入して 5~6 時間媒精 した。媒精後の卵子は5%FBS 添加修正合成卵管液で卵丘細胞と共培養を行い、 媒精後7~8 日目までに発生した胚盤胞を供試した。 3.2.3 ガラス化保存および加温 胚を25%エチレングリコール(和光純薬、15209-85、以下 EG)、25%ジメチ ルスルホキシド(ナカライテスク、13408-64、以下、DMSO)および 0.4% BSA 添加修正PBS(以下、VSED)(Ishimori ら 1993)をガラス化溶液に用いて次 の方法によりガラス化保存した。 胚を50% VSED に 60 秒間平衡した後、VSED(11μL)に移し、30 秒以内に
VSED とともに 0.25 mL プラスチックストロー(IMV, L’Aigle, France)へ充填し、
液体窒素蒸気中に 2 分間静置し、液体窒素中に浸漬し保存した。ストロー内の 構成は、図3.1 に示すとおり、綿栓側から希釈液、VSED、胚を含む VSED、希 釈液の順に空気層を隔てて充填した。なお、希釈液として5% EG 0.15 mol/L シ ュークロースおよび20% CS 添加修正 PBS(以下、EGS)を用いた。 加温は、図3.2 に示すとおり、ストローを液体窒素より取り出し、空気中で 5 秒間保持後、温水中に入れ氷晶が消えるまで保持して行った。ストローの熱シ ール部を持って2~3 回振って各液層を混合し、綿栓部を上にして温水中に 3 分 間垂直に保持して耐凍剤のストロー内希釈を行った。 試験1 では加温操作に用いる温水の温度を 20℃および 30℃に設定した。試験 2 では 20℃温水中で加温し耐凍剤のストロー内希釈を行ったストローを 38.5℃ の培養器内に0 分、5 分、20 分および 40 分間、水平に保持した。試験 3 ではス トロー内に充填する希釈液として EGS または 0.2 mol/L シュークロース 20%
35 CS 添加修正 PBS(以下、SUC)を用いた。 ストロー内希釈後の胚は5% CS 添加 TCM199 で卵丘細胞との共培養により 48 時間培養した後に形態観察を行い、24 時間後の胞胚腔の再形成による生存胚 率と48 時間後の透明帯からの脱出による透明帯脱出胚率を調査した(図 3.3)。 3.2.4 .統計処理 統計処理はコンピューターソフトSPSS (SPSS 11.5J, SPSS Inc., 東京)を 用い、生存率および透明帯脱出率についてPearson のχ二乗検定または Fisher の直接確率計算法を用いて危険率5%未満を有意差ありとした。
36 3. 3 結果 ガラス化保存したウシ体外受精由来胚における加温温度が生存性に及ぼす影 響を表 3.1 に示す。20℃区および 30℃区の加温 48 時間後の生存胚率はそれぞ れ92.1%および 89.7%、透明帯脱出胚率は 50.0%および 44.8%であり、試験区 間に有意差は認められなかった(P > 0.05)。 加温後の保持時間が胚の生存性に及ぼす影響を表3.2 に示す。保持時間 0 分、 5 分、20 分および 40 分の各区の、加温 48 時間後の生存胚率はそれぞれ 92.1%、 73.3%、70.6%および 61.5%であり、0 分に対して 5 分、20 分および 40 分の各 区は有意に生存胚率が低下した(P < 0.05)。また、加温後 48 時間の透明帯脱出 胚率はそれぞれ50.0%、53.3%、38.2%および 7.7%であり、0 分および 5 分に 対して40 分で有意に透明帯脱出胚率が低下した(P < 0.05)。 希釈液の組成が胚の生存性に及ぼす影響を表3.3 に示す。EGS 区および SUC 区の加温後48 時間の生存胚率はそれぞれ 92.1%および 92.1%、透明帯脱出胚率 は50.0%および 50.0%であり、試験区間に有意差は認められなかった(P > 0.05)。
37 3.4 考察 ガラス化保存したウシ胚のストロー内希釈において、加温条件が胚の生存性 に及ぼす影響を検討した。その結果、試験1 では 30℃で加温した場合に 20℃で 加温した場合と同等の胚の生存性と発育性が得られた。試験 2 では直接移植に おける移植所要時間の延長を想定して、ストロー内希釈後のストローを 38.5℃ の条件で保持したところ、保持時間5 分以上で生存胚率が、40 分以上で透明帯 脱出率が有意に低下した。試験3 では希釈液として EGS および SUC を用いた 胚の生存胚率や透明帯脱出胚率は試験区間に有意差は認められなかった。 ガラス化液は過冷却状態でガラス化しているため加温時は脱ガラス化の生じ る温度域をできるだけ急速に加温する必要がある。同時に耐凍剤による化学毒 性の影響を避けるために加温後はできるだけ早く耐凍剤を希釈しなければなら ないとされている(葛西 2001)。
Campos-chillo’n ら(2005)は、7 mol/L EG および 0.5 mol/L ガラクトースを含
む18% (w/v) Ficoll 70 溶液をガラス化液とした際の加温温度は 20℃より 37℃に おいて加温後の胚の胞胚腔再拡張率と脱出胚盤胞率が有意に高く、保持温度を 高めたことにより細胞からの耐凍剤の移動が促されたことがその原因と推察し ている。一方、本研究と同様にVSED をガラス化液に用いてウシ胚をガラス化 保存したこれまでの報告では、加温は 20℃程度で行われている(Ishimori ら 1993; Vajta ら 1997; 小渕ら 2001; 小田ら 2005)。本研究では、これまで の報告と同様の 20℃と、農家段階で普及している緩慢凍結胚の直接移植法
(Massip ら 1987; Voelkel と Hu 1992; Dochi ら 1998)で用いられている 30℃で加温した場合の生存性を比較した。その結果、30℃で加温した場合の生 存性と発育性は、20℃で加温した場合と同等の成績が得られた。このことから、 ガラス化保存したウシ胚を農場段階で利用する際に、他の緩慢凍結胚と同様の 温度で加温することが可能であり、農場段階でガラス化胚を利用する際に操作 手順の混乱が少ないものと考えられた。 Vajta ら(1995)および Campos-chillo’n ら(2005)は、ガラス化保存したウシ胚 をストロー内希釈後にそれぞれ22℃で 30 分間、37℃で 15 分間保持したが、胚 の生存性や発育性に有意差が認められないことを報告している。一方、Matoba ら(2003)は、1.5 mol/L EG と 0.1 mol/L シュークロースを含む液を用いたウシ 体外受精由来胚の緩慢凍結において融解後のストローを 38.5℃に 30 分間保持 することで胚の生存性が有意に低下することを報告し、Inaba ら(2011)はクライ オトップ上でガラス化保存したウシ体外受精由来胚のストロー内希釈において 胚を希釈液(0.3 mol/L シュークロース液)中に 30 分間曝すことで生存性が低 下することを報告しており、凍結液やガラス化液、希釈液中での保持時間の延
38 長が胚の生存性を低下させることを指摘した。本研究の結果はこれらの報告を 支持するものであった。さらに、Dochi ら(1998)は、1.6 mol/L プロパンジオー ル液または1.5 mol/L EG 液で緩慢凍結したウシ胚のダイレクトトランスファー において、移植所要時間が11 分以上で受胎率が低下し、凍結溶液中に胚を長時 間保持することは有害であることを指摘している。本研究と同様のストロー内 希釈法におけるストロー内の耐凍剤濃度は加温後の短時間で平衡され、移植時 には緩慢凍結のダイレクトトランスファー法(Dochi ら 1998; Voelkel ら 1992)と同程度の耐凍剤濃度に希釈されていることが確認されているが (Akiyama ら 2012)、性判別胚の野外移植試験において、移植所要時間が延長し た場合には有意ではないが受胎率の低下が認められることが報告されている (Akiyama ら 2010)。 このことから、ガラス化保存したウシ胚のストロー内希釈においても、加温 後に体温程度の温度でストローを長時間保持することにより胚の生存性が低下 する可能性があり、直接移植において移植所要時間が延長した場合には受胎性 の低下を招く可能性があると考えられた。 本研究で用いたガラス化保存において、EGS 区では EG 5%を含んだ希釈液 を用いており、ストロー内容液を混合した後の胚は、EG 7%程度、DMSO 3% 程度を含む溶液で保持されている(Akiyama ら 2012)。一方、SUC 区は希釈液 にEG や DMSO を含まないことから、胚からの耐凍剤の除去を速やかに行うと ともに移植時の溶液の耐凍剤濃度を低下させることが可能であり、胚の生存性 の一層の向上につながることを期待した。本研究の結果から、異なる組成の希 釈液を用いてガラス化保存した両区において同等の胚の生存率が得られたこと から、希釈液の組成の違いによる生存性への影響は認められないものと考えら れた。しかし、SUC 区では希釈液とガラス化液の濃度差が EGS 区に比べて大 きいことからストロー内に胚を充填する際にはガラス化液が希釈されることが ないよう操作に注意することが必要と考えられる。 ガラス化保存したウシ胚の移植を農家段階で普及するためには、高い受胎率 を安定的に得ることと同時に、加温後に直ちに受胚牛に移植することが可能な 手法を確立することが望まれる。本研究の結果から、ガラス化保存したウシ胚 のストロー内希釈において、加温温度の違いや希釈液の組成により胚の生存性 の低下は認められなかったが、ストロー内希釈後の保持時間が延長した場合に は胚の生存性が低下する可能性があることが示された。