九州大学学術情報リポジトリ
Kyushu University Institutional Repository
酸素電極法を用いたイネ光合成能力の種内変異と遺 伝に関する研究
長峰, 司
https://doi.org/10.11501/3070096
出版情報:Kyushu University, 1993, 博士(農学), 論文博士 バージョン:
権利関係:
爪り E廿川川 bubL M FHV』TU BAnn口 】qs,J A附川河馴 A口 ?Q 巾冗ふ?
ふ小間凶内
,aぺF AF 」
た い伝 い 噛 忠 岡肘 tC φ伊』
回 並ハ ゆ叫仙 砂阪 主糸 子宮屯 詳細官 町糊戸市川H 亦 久
長隆司
η4U ハu.v ハ吋υ 4EBB-
、ーー-
目次
頁 第1章 緒言
第2章 多点式酸素放出量測定装置の開発
1 . 開発の背景
2. 開発の重点 3 . 装置の概要 4.
実際の測定第5 酸素放出量の遺伝分析
1 7 7 7 8 2 3 3 6 7 2 3 6 6 7
2
8 8 1 1 1 1 1 2 3 3 3 4 5 5 6
第3章 測定試料の前処理方法と最適測定条件の解明第1節 葉身採取時刻に伴う酸素放出量の変化 第2節 葉片の前処理方法
第3節 酸素放出量に及ぼす園場の気象条件の影響
第4節 酸素放出量の最適測定条件の確立 第5節 生育時期別の酸素放出量の変化 第4章 酸素放出量の種内変異
第1節 アジア栽培イネにおける酸素放出量の種内変異 第2節 酸素放出量の種内分化
第3節 目本のイネ品種における酸素放出量の変異 第4節 突然、変異系統における酸素放出量の変異
第1節 特定の生育時期における酸素放出量の遺伝分析 6 8 第2節 個体の生育時期を考慮した酸素放出量の遺伝分析 8 2 第3節 F3系統における酸素放出量の遺伝分析 8 8
第6章 高 ・ 低酸素放出量系統の選抜 9 2
頁
第7章 総合考察 9
7
1 . 光合成多点測定装置の重要性
97
2 . 葉片前処理法の開発と測定条件の決定
97 3. 光合成能力のイネ種内変異について
9 84. イネ光合成能力の遺伝的分化について
9 95 .
多収性イネ育種の方向 9 96 . 光合成能力の遺伝子解析について 100
摘要
102
謝辞
105
引用文献
106
英文要約 1 1 5
"
冒園田園圃ー
第I章 緒言
作物の多収性は農業のもっとも重要な目標であり、 物質生産性 ・ 耐病性 ・ 耐 虫性に関する品種改良、 施肥技術、 病虫害防除、 土壌改良、 機械化などの個別 技術が総合的に組み合わされて達成されるものである。 光合成は植物の物質生 産にかかわる基礎的機能であり、 植物体の地上部全重を生産の対象とする青刈 飼料作物jでは、 光合成の機能を改良して乾物生産量を増加させることがで きる。
イネのように穀実を生産の対象とする作物では、 光合成能力と穀実収量との関 係は複雑であるが、 その機能
・
能力の改良は間接的に収量増加につながり、 光 合成能力の改良は穀実作物でも重要である。これまで、 作物の光合成能力の向上に関する研究は群落葉量の効率的な増大、
すなわち、 単位土地面積あたりの葉面積の拡大および光合成器官の維持に重点、
があった。 イネの群落構造の変化による群落光合成能力の向上にあたり、 半倭 性遺伝子( sd-1)をもっ品種十石、 低脚鳥尖などが果たした役割は大きい(菊 池ら1985)。 半媛性遺伝子はシンクサイズを減少させずに稗長を短縮し、 多 肥栽培による増収効果を大きくしたのみならず、 同時に葉身を直立させ、 受光 態勢をよくするという効果をもたらした。 草型の改良をとおした群落光合成能 力の向上による多収性品種の育種は草型育種(角田1964) といわれる。 フィ
リピンの国際イネ研究所(1 RR 1)や韓国で育成された最近の品種は多けっ型で 葉身は直立し、 単位土地面積あたりの葉面積が大きく、 太陽光を効率よく受け るという受光態勢の点では理想的な草型をもっ (吉田1976、 金1979、 Chung and Heu 1980)。 現在、 わが国ではインディカ品種から大きなシンク容量(総 籾数x 1籾重)を日本品種にとり込む多収性品種育種が行われており、 穂重型 で半媛性の品種が育成された場合にはソース量(葉面積×面積あたりの葉身光 合成能力)の不足が問題となり、 質的な改良、 すなわち、 単位葉面積あたり光 合成能力を遺伝的lこ向上させる必要が生じると考える。
冒国国-
高等植物の光合成型には C3型、 C4型、 ベンケイソウ型酸代謝(CAM)の3種 が知られており、 植物種は光合成型に基づいてC3植物、 C4植物、 CAM植物に区 別される。 イネなどのC3型植物は、 C4型植物に比べて光飽和点が低く、 光呼吸 を有するために光合成能力が低い特性をもっている。 単位葉面積あたり光合成 能力を遺伝的に改良するためには、 光呼吸がなく光合成能力の高いC4型植物の 形質をC3型植物に導入することも一つの方法である。 しかし、 Bjorkman旦主主L (1971)によるハマアカザ属での試みは、 C4型植物の光合成能力をC3型植物に 導入することの困難性を明らかにした研究として注目される。 すなわち、 ハマ アカザ属のC3穫とC4種の交雑実験によりさまざまの人工的中間種が作出された が、 雑種後代からC3親の光合成能力を顕著に越える個体は獲得できず、 C4型光 合成の特徴とする形質を導入しでも光呼吸や低光合成能力を克服できないこと
が明らかにされた。
これまでに単位面積あたり光合成能力の種間変異に関して多くの研究が行わ れた。 イネ、 トウモロ コシ、 コムギ、 トマトなどの主要作物で光合成能力の種 内 変異が明らかにされ (Heichel and Musgrave 1969、 Murthy and Singh 19
79、 Augustine and Stevens 1976)、 その結果、 種聞には光合成能力に大きな 変異が存在することがわかった。
イネでは村田(195 7)が水稲29品種の最高分けっ期における炭酸ガス交換速 度を測定して、 最高値/最低値比 1. 61の変異を報告した。 秋田(1980)は40 品種で同比1. 70の変異があることを明らかにした。 また、 1
RR
1では50品種を用 いて1. 80という最高値/最低値比が報告された(Anonymous 1968)。 これらの実験で用いられた品種は数が限られ、 また、 世界各地のイネ遺伝資源を網羅し ておらず、 イネ種内における光合成能力の変異を知 るための実験材料としては
不十分であった。
光合成能力の改良が作物の多収性にどの程度寄与するか、 あるいはこの能力 がどのような遺伝様式に基づいているかに関する研究は少なく、 明確な結論は
乙
得られていない。 その理由の一つはこの形質の測定が煩雑であり、 しかも多数 の試料を同時に遺伝分析することが困難なためである。 これまで報告された光
合成能力の遺伝様式に関する報告をまとめると、 量的遺伝であり、 多くの遺伝 子が関与するという報告が多い(1 zhar and Wallace 1967、 Wilson and Coo
per 1969、 小島 1971、 Crosbie旦呈上 1978、 Ellison弘主上 1983 )。
Izhar and Wallace (1967)は、 インゲンマメで光合成能力の遺伝は量的で
あるが、 関与する遺伝子数は比較的少なく、 低光合成能力はある程度優性であ ると結論した。 小島(1971)は、 ダイズでんの光合成能力は正規分布に類似し た分布をとり、 光合成能力の遺伝は量的であると報告した。
一方、 低 能力が劣性の単遺伝子支配であるという報告もある(Augustine et al. 1976、 Hayashi el al. 1977、 鮫島 1985)。 イネでは光合成能力は単遺伝 子支配であるという報告がある。 Hayashi et al. (1977)は炭酸ガス交換速度 からみた光合成能力は、 低い能力が優性の単遺伝子分離をすると報告し、 光合 成能力の遺伝的改良が可能であると示唆した。 鮫島(1985)は炭素同位体分別 能(δ1 3C : 大気中の炭酸ガスには原子量13の炭素からなる分子量45の重い炭 酸ガスが約1 %の割合で含まれている。 これは光合成反応における炭酸の輸送
・
固定の段階で同位体効果による分別をうける。 その程度は標準試料と測定試 料中の13
C含量の干分偏差として係数化される〉を測定し、 弱い分別能が優性 の単遺伝子分離にみえる結果を得た。 トマトでもAugustine et al. (1976)は 炭酸ガス固定能力が単一の遺伝子に支配され、 しかも低い能力が優性であるこ とを明らかにした。坂(1985)はイネのクロリナ×正常種のF1個体で光合成暗反応の炭酸固定初 発反応を触媒する酵素Ru BP C (リブロース二リン酸カルボキシラーゼ)活性は 生育期間をとおして 両親より高い傾向を示し、RuBPO (リブロース二リン酸オ キシゲナーゼ)活性は逆に両親より顕著に低く、 酵素から見た光合成、 光呼吸 のヘテロシス的現象があることを報告した。 村山ら(1982)はイネにおいて品
3
種の 組合せによっては光合成能力にヘテロシスがみられる場合があることを報 告した。 玄ら(1989)は雄性不稔系統を利用したF 1雑種において若い止葉で測 定した光合成速度にはヘテロシスはみられないが、 登熟期の止葉ではヘテロシ スがみられるとした。 しかし、 Yamauchi and Yoshida (1985)は35組合せのイ
ネF
1雑種の光合成能力、
葉面積、 分けっ力などを調べて、 F 1には光合成能力のヘテロシスは現れず、 旺盛な分けっと早い葉の展開が雑種強勢をもたらすと報 告した。 トウモロ コシなど他殖性作物ではF 1の光合成能力にヘテロシスが認め られるという報告が多い (Heichel and Musgrave 1969、 Monma and Tsunoda 1979、 Crosbie et al. 1978)が、 イネのF 1雑種において光合成能力にヘテロ シスが現れるかどうか不明である。 以上のように、 イネにおいては光合成能力 の遺伝に関する基礎的知見は少ない。
多収性品種の育種において光合成能力が選抜の対象とされたことはない。 小 島(1971)はダイズで初期世代における光合成能力の選抜が多収個体の選抜に 比較的効果があると報告したが、 Wiebold et al. (1981)は効果はないとしたo
Mahon and Hobbs (1981)はエンドウで初期世代における選抜が比較的効果が あるとした。 栄養繁殖期聞における光合成能力の選抜はダイズやトウモロ コシ で有効であるという報告がある (Secor et al. 1982、 Moss and Musgrave
1971)。 イネでは光合成能力に関する選抜実験は行われていない。 イネの育種 では多収性品種育成の選抜手段として全重や玄米重が用いられており、 乾物生 産性に関する生長解析の各種ノマラメーターや炭酸ガス交換速度などで表した光 合成速度が選抜の対象とされたことはなかった。 多収性品種の育成はイネ育種 の基本目標であるので、 高光合成能力を選抜形質に加えて多収性を追及するこ とは今後の研究において重要と考える。 そこで光合成能力に関する選抜がイネ で有効であるか否かを明らかにする必要がある。
光合成能力の種間 ・ 種内変異の探索 ・ 評価、 その遺伝分析および高能率系統 の選抜を行うためには、 簡便、 迅速、 安定して測定できる装置の開発が必要で
4
ある。 光合成能力の測定には炭酸ガス交換法、 ワールブルグ検圧法 (Sestak et al. 1971)やガスクロマトグラフィ(山末ら 1979)などの方法がある。 炭 酸ガス交換法は葉身にとりこまれる炭酸ガスの濃度を測定して光合成速度を求 めるものであり、 赤外線分析計の改良やチャンパーおよび通気法の改良によっ て測定精度が著しく向上し、 光合成研究に多く用いられている。 しかし、 この 方法は装置が大規模で、 かっ、 測定系の構造が複雑であり、 測定方法が迅速
・
簡便ではないので、 多点数の分析を必要とする変異の探索、 評価、 遺伝解析の 実験に適しているとはいえない。
一方、 ポーラログラフィーの原理を応用して白金電極に一定の電圧を与え、
液相中の溶存酸素が電極で酸化還元を受けることにより生じる電解電流を増幅 して酸素濃度を測定して光合成速度を求める酸素電極法がある。 これは水中の 藻類や単離した葉緑体の光合成活性の測定lこ使われる(Delieu and Walker 19 72)とともに、 高等植物の葉組織の光合成測定にも使われている (Jones and Osmond 1973、 Ishii et al. 1977、 Yamashita et al. 1978、 石井1981、 Saka and Chisaka 1985)。 この方法は葉身全体あるいは一部の葉片を HEPES緩衝液 やリン酸緩衝液などの溶液中に沈めて光を照射して光合成を行わせ、 放出され る酸素の濃度を液相中で測定するものである。 光合成に利用される溶液中の炭 酸分子はHCO 3ーである(Jones and Osmond 1973) 0 HC03ーは葉片の切断面から 葉肉細胞へ入り、 葉面の気孔からとり込まれる量は少なく、 気孔抵抗の影響を 無視できると考えられている(Pitman et al. 1975、 石井 1981)。 すなわち、
炭酸ガスがほぼ飽和した、 しかも気孔抵抗の影響が少ない条件下での光合成を 計測できる。 また、 促進剤や阻害剤などが光合成や呼吸に及ぼす影響を調べる ことができる長所もある (Ishii et al . 1977、 Hyeon et al. 1982)
イネの光合成はこれまで炭酸ガス交換法で測定されてお り、 酸素電極法によ る測定例はきわめて少ない。 Ishii et al. (1977)はイネに本法を最初に適用 し、 葉片の大き さと酸素放出量との関係や最適測定条件を検討した。 その結果、
s
本法を用いて測定した光合成速度は赤外線ガス分析計によって測定した値より 低かったが、 切断面近辺の組織の破壊に起因する光合成速度の低下を補正する とほぼ等しくなることを明らかにした。 坂 ・ 千坂(1981a)は酸素電極法を除 草剤による光合成阻害力の検定に応用した。 本法をイネの光合成能力の遺伝育 種的研究iこ初めて取り入れたのは伊藤(1982)であった。 伊藤は酸素放出量の 品種間変異を調査し、 両親と雑種F 1を材料にして酸素放出量に関する遺伝分析 をするなど酸素電極法を用いた光合成能力に関する先駆的な研究を行なった。
これまでイネの光合成能力の種内変異および遺伝に関する研究は少ないが、
これらは多収性品種の育成にとって重要な課題の一つであると考える。 本研究 は酸素電極法を用いて遺伝分析が可能な迅速多点測定法の開発を行い、 イネの 光合成能力に関する遺伝的変異を明らかにし、 遺伝分析を行い、 高能率光合成 系統の選抜の可能性を検討したものである。 以下、 本論文で用いる「光合成能 力」とは「個葉の単位面積あたりのみかけの光合成速度」をいい、 酸素電極法 で測定した単位葉面積
・
時間あたりの酸素放出量 (μrnol 02/drn2/hr)で示す ものとする。本研究は、 1983年 から1990年にわたって農林水産省旧農業技術研究所
・
生理 遺伝部・
遺伝科・
遺伝第七研究室および農業生物資源研究所・
遺伝資源第一部・ 植物探索導入研究チームに在籍中行なった研究の成果をとりまとめたもので ある。 その一部はすでに Rice Genetics Newsletter (Nagarnine 1985)、 農業 生物資源研究所報告 (Nagarnine et al. 1987、 Nagarnine 1991a)、 JARQ(Na
garnine 1990)、 日本育種学会(Nagarnine 1991b)、 同講演会 (長峰ら 198 4、 長峰
・
中川原19 86、 長峰・
中川原1987)で報告した。6
冒田園4・h
第2章 多点式酸素放出量測定装置の開発
1 . 開発の背景
これまで作物の光合成はおもに赤外線炭酸ガス分析計を応用したガ、ス交換法 により測定されてきた。 しかし、 この分析計による測定装置は構造が複雑で測 定方法が迅速
・
簡便ではないので、 光合成能力に関して多くの試料を同時に分 析する必要がある遺伝変異の探索・
評価や遺伝解析の実験に適していない。 炭 酸ガス交換法で初めてイネF2雑種集団の光合成能力を測定した実験では雑種集 団をポット栽培で養成し個体ごとに測定したので、 1日lこれから40個体しか測 定できず、 1集団の測定に7日から8日を要し、 分析の量、 質ともに制約が多 かった(林 健一氏私信)。 また 、 ダイズF2雑種集団の光合成能力を測定した 小島(1971)も同じ方法を用いたが、 分析した個体数は少なく、 全体でポット 栽培した70から90のF2個体しか測定できなかった。 炭酸ガス交換法では着生葉 を供試するため扱える材料の数が限られ、 材料の扱い方にも繊細な注意が必要 とされる。 すなわち 、 葉を同化箱にセ ットする際に急激な温度変化に合わせな いこと、 実験室にもちこんだ材料はなるべく明るい場所において気孔を閉じさ せないことなど十分な注意が必要とされるので、 多点数を扱いにくく、 遺伝変 異の探索 ・ 評価や遺伝解析の実験には不向きである。 そこで、 炭酸ガス交換法 に代わる光合成測定方法を開発しようと考えた。2 .
開発の重点光合成能力の遺伝変異や遺伝分析を可能にするにはまず多数の材料を扱うこ とが必要であり、 そのための測定機器の開発の重点は測定の迅速性と簡便性の 一点である。 着生葉よりも切葉を用いるほうが多点数を容易に取扱えると考え た。 また 、 測定に用いる葉身の面積はなるべく小さいほうが多数の材料の前処 理や保存を同一条件でできると考えた。 それらの重点を満たすと考えたのが酸
ワ
素電極法である。 すなわち、 酸素電極法の利点である小型で簡潔な構造を利用 し、 光合成lこより切断葉から水溶液中に放出される溶存酸素濃度を同時に多数 測定できる測定装置であった。
3 . 装置の概要
以上をもとに設計
・
開発した測定装置の構造を模式図でF i g. 1に示した。 ま た 、 酸素放出量を測定している装置全体の概観をF i g. 2 (写真〉に示した。式の測定ユニ ットはアクリル製の反応容器、 凸レ ンズ付きの集光器、 タングス テン
・
ハロゲンランプ(Silvania社 EFN 12V-75W)、 水中撹祥用マグネティ ックスターラ一、 クラーク型酸素電極からなる。 1台の測定装置に6基の測定 ユニ ットを水平に設置した。 反応容器の内容積は25mlであり、 容器全体はアク リル製水槽に沈めて温度を調節する。 容器には反応溶液として24mlの50mM HE- PES (N-2 - hydr oxyethyl piperazine - N "- ethane sulfonic acid)緩衝液 と炭素源として炭酸水素ナトリウム液を1m 1入れる(Ishii et al. 1977)。 測 定に用いる葉片をナイロンネットっきアクリル製のリーフホルダーにはさみ、照射光に対して垂直に向くように反応容器に入れる。 安定して高い酸素濃度を 得るため 、 反応溶液はマグネティ ックスターラーにより測定中撹持されている (Fig.
3 、
写真)。タングステン ・ ハロゲンランプの光は集光器をとおして反 応容器に照射される。 照射光の照度はランプと反応容器との距離を変えて2万 から14万ルックスまで調節できる。 反応溶液の水温は温度調節装置でo
,.._,60 oc (精度:t O. 50C )に制御できる。 光照射後、 葉片から放出され反応液中に溶存 した酸素の濃度変化を酸素電極により測定し、 6ペン式記録計で出力する。 酸 素放出量の単位はμmol O:;jdmL/hrであり、 1時間あたり葉身100cm2 ( 1 dm2) あたりの酸素濃度で示した。
本装置の完成により1日で処理
・
測定できる試料数は大幅に増加した。 まず、測定開始前に装置のウオームアップ、 記録計の零点補正、 スパン調整、 酸素電
8
冒田園圃ー
Lens
町一 CE--- 1ム一 J
n一一e一L句一 C E-- 1ム一 ,A
a一一'n一一昨一 ム ・・ Q】一 一
σり一一n一 一
日 十♀ E・E・ 一
5・・・・
一 C E-- 「 し / μん外
四図図
Recorder Oxygen
electrod
仁二コ
。 。
Acrobatic magnetic stirrer
-+-
Water bath
Fig. 1. Diagram of mulli-channel apparatus with six oxygen eleclrodes
守
Fig. 2. A multi-channel apparatus for measurement of oxygen evolution.
10
Fig. 3. Leaf disks in a reaction cell and an oxygen electrode.
極の補正などに30分を要する。 以後、 6基の反応容器への HEPES緩衝液や炭酸 水素ナトリウム溶液の注入、 葉片試料の挿入、 酸素放出量の測定、 試料の摘出、
使用した緩衝液の排出からなる一連の操作は40分で終了するので 、 午前8時半 から午後5時まで測定すると仮定して1日に1台で1 2回反復することができて、
合計で72点の試料を扱うことができるようになった。 メンテナンスとしてはタ ングステン
・
ハロゲンランプの約200時間ごと、 電極膜の約100時間ごとの交換 であり、 HEPES緩衝液がやや高価なことを除けば維持費は廉価である。4. 実際の測定
以上のように、 この装置の開発で扱える点数は増加したが、 本研究ではこの 装置を2台作成して1固に1 2点を同時に測定できるように効率を向上させた。
すなわち、 2台を1日に8時間フル稼働させて少なくても1 4 4点の酸素放出量 を測定できる態勢が整った。 一人でこの装置2台を稼働させ、 酸素放出量を測 定しな がら葉片試料の作成、 緩衝液の作成、 記録紙からの酸素放出量の値の読
みとりが実施可能となった。
これにより、 多数品種を用いる遺伝変異および多くの個体からなる雑種集団 を用いる遺伝分析までの進展を念頭に入れた酸素電極法によるイネ光合成能力 の研究の基礎を確立した。
12
第3章 測定試料の前処理方法と最適測定条件の解明
多点式酸素放出量測定装置の設計 ・ 開発により光合成能力の多点数簡易測定 は可能となったが 、 つぎに問題となる点は、 葉身採取時安IJの推移による酸素放 出量の変化の解明、 安定した値を示すための葉片の前処理方法の開発、 圃場に 栽培する材料を用いることを前提とした場合の酸素放出量に及ぼす園場の気象 条件の影響の解明、 酸素放出量の最適測定条件の決定、 葉令別の酸素放出量の 変化の把握などである。 これらは酸素電極法による光合成能力の測定を安定確 実にするために明らかにしておく必要があるばかりでなく、 多点数を用いる遺 伝分析までを念頭に入れた測定のためにも不可欠である。 本章ではこれらの問 題点を解決するための実験を行った。
第1節 葉身採取時刻に伴う酸素放出量の変化
酸素放出量の最適測定条件を決定するための基礎資料として、 測定材料とす る葉身を1日のどの時刻で採取しでも 安定した放出量が得られるかを検討した。
材料および方法
198 3年4月21日に播種し、 5月1 6 日に圃場に1株1本植えで栽培した水稲品 種コシヒカリを用いた。 本田施肥量は、 元日巴として窒素O. 5 K g/ a 、 分けっ期追 肥として窒素0.2Kg/aとした。 以下の実験もすべて同じ施肥量とした。 葉身採 取時亥IJに伴う酸素放出量の変化を調べるため、 主稗の完全展開した第7葉ある いは第8 葉の葉身を7月7日午前8時30分、 同10時30分、 午後12時30分、 同3 時の4回国場で採取した。 採取後30分、 60分にリーフディスクパンチを用いて 葉身中央部から中肋を含む直径12mmの葉片を打ちぬいた。 葉片作成後ただちに 12反復で酸素放出量を測定した。 測定条件は温度を250C、 光照度を7万ルック ス、 HEPES緩衝液のpHを7 . 2とした。
13
結果および考察
午前8時30分に採取した葉身から30分、 60分後に作成した葉片の酸素放出量 は、 それぞれ 604、 494μmol 02/dm2/hrであった (Table 1 )。 午前10時30分 に採取した葉身では、 それぞれ 341、 357 μmo1 02/dm三/hrで8時30分に採取し た葉身から作成した葉片の値より低下した。 それ以後も葉身の採取時亥IJが遅れ るにつれて両者の酸素放出量は徐々に減少し、 午後3時に採取した葉身から作 成した葉片の値は、 それぞれ 181、 173 μmol 02/dmL/hrであった。 すなわち、
葉身の採取時刻が経過するにしたがって酸素放出量は減少することが明らかと なった。 これは葉身内に蓄積される光合成産物の量に比例して光合成能力が低 下するためと考えたo P 1
112k
(1984)はアルフ アルフ ァで非構造糖類含量が日 中増加することを、 津野・
藤瀬(1965)は甘藷で光合成産物が蓄積されると光 合成能力が低下することを報告している。 葉身の採取時期による酸素放出量の 変化はオオムギでも観察されている(Pitman豆l呈上 1975)0 Pitmanらは8月 から9月に かけて4回オオムギを播種し、 発芽後8 -- 10日の葉身を1日の異な る時刻に採取し葉片を作り、 酸素電極法により光合成能力を測定した。 その結 果、 酸素放出量は1日のうちで朝9時にもっとも高く、 時刻が経過するにつれ て値が低下した。 酸素放出量が低下した理由を酸素電極法の性格上炭酸ガスは 葉片の切断面をとおして直接とりこまれるため、 午後に発生する気孔開度の減 少による炭酸ガスのとりこみ減少ではなくて、 おそらく葉身の固有な光合成能 力の変化によって引き起こされたものとしている。以上から、 イネ葉身の酸素放出量は時刻にともなって変化するので、 時刻を 限って採取した葉身から葉片を作成して酸素放出量を測定するのは多点数測定 には向いていなかった。 したがって、 遺伝的特性を解析するためには新たな葉 片作成方法が必要となった。 そのためには葉片に何らかの処理を行い葉身の採 取時刻に関わりなく安定した値を示す方法を見つける必要があった。
i斗
-田園�
Table
1.
Changes of oxygen evolulion with lhe sampling lime of leaf bladesSampl ing l ime of leaf blades
8:30
a. m.10・30
a. m.12:30
p. m.15 :00
p. m.Oxygen evolulion
(
μmol 02/
dm2/
hr)
Measurement
A
Measuremen lB
604 ::t 50 494 ::t 112 341 ::t 94 35 7 ::t 120
239 ::t 132
240士53
181 ::t 5 6 17 3 ::t 60
t-value
2. 87料 0.36n.s 0.02n.s O. 34
n . sMeasurement
A:
Oxygen evolution was measured at30
minutesaf ter samp
1
i ng.Measurement
B :
Oxygen evolution was measured at60
minutes af ter samp1
i ng.The conditions for measurement of oxygen evolution were as follows; Temperalure
250C.
Light intensity70
klux pll of HEPES buffer solution7.2.
林 Significanl at the
1%
level.n. s.
: N
0 t s i gn i f i c a n t.第2節 葉片の前処理方法
葉身の採取時刻に関わりな く品種の遺伝的能力を反映した酸素放出量を示す 試料を得るため、 葉片の前処理方法を検討した。 多数の材料を扱うという条件 を満たすために は 、 前日に葉身を採取 して葉片を作成したほうが測定の効率が あがると考えた。 また、 作成した葉片の乾燥萎凋を防ぐため、 ガラス製シ ャー レ内の蒸留水に葉片を浮かべ、 ふたを して飽和した水分条件下で保存すること を考えた。 この 方法で測定した値が、 最適条件と考えられる条件と比較して同 程度の安定した値であるならば、 本研究で目指す遺伝分析に までたどりつける と考えたからである。
材料および方法
コシヒカリの葉身を採取後、 1枚の葉身の中央部から近接する2枚の葉片を
リーフディ スクパンチで作成し、 それぞれ蒸留水を入れたガラス製シ ャーレに 浮かべ、 以下に示す 2種類の葉片前処理方法を比較した。 すなわち、 第1は光 照度4万ル yクス 、 照明が午前6時から午後8時、 昼温が280Cで夜温が220Cの 人工気象室で1日間葉片をインキュベー卜する方法であった。 第1法は昼の光 照度、 温度条件をなるべく自然、条件に近い条件で葉片をインキュベー卜するも のである。 第2は光照度が 1, 000ルックス、 照明が午前6時から午後8時、 温 度が250Cの実験室で葉片を
1日間インキュベー卜する方法であった。
第2法は 葉片のインキュベートを 実験室で簡便に行 おうとするものである。 酸素放出量 の測定は葉片作成の翌日 に午前9時から午後5時10分まで7回行なった。 材料 および測定条件は第1節と同じであった。結果および考察
第1の前処理方法、 すなわち、 人工気象室でインキュベートした場合の葉片 の酸素放出量を午前9時から午後5時10分まで1日7回測定したCTable 2)。
16
園田園田h
最低値は 506::t 28μmol 02/dm2/hr 、 最高値は 570士85μmol 02/dm2/hrであり、
両者の差は小さかった。 また、 値は7回の測定をとおして安定しており、 平均 値は532μmol 02/dmL/hrであった。 一方、 実験室でインキュベートした葉片の 酸素放出量は、 最低値が344土82μmol 02/dmι/hr 、 最高値が498士38μmol 02/
dm2/hrで、 154μmol 02/dm:こ/hrの差があり、 ば‘らつきが大きかった。 平均値は 430 μmol 02/dm乙/hrであり、 人工気象室でインキュベートした葉片の値より 102μmol 02/dmL/hr低かったCTable 2)。 午前9時における人工気象室でイ ンキュベー卜した葉片の酸素放出量は、 実験室で前処理 を行った葉片の値より 202μmol 02/dm2/hr高く、 t検定の結果5 %水準で有意であった。 10時15分の 測定以降6回の測定では、 いずれも人工気象室でインキュベートした葉片の酸 素放出量は、 実験室でインキュベー卜する方法より値が高く、 得られる値も安 定していると結論した。
第3節 酸素放出量に及ぼす圃場の気象条件の影響
第2節で確立した葉片前処理方法を用いて圃場に栽培した材料の酸素放出量 を測定し、 葉身採取日の気象条件の変化に影響されないで安定した値が得られ るかどうかを検討した。 これは多点数の試料を扱う遺伝変異や遺伝分析の実験 を行う上で前提となるもう一つの条件である。
材料および方法
198 3年5月11日に播種し、 6月13日に圃場に1株1本植えで栽培したコシヒ カリを用いた。 主稗の最上位完全展開葉を用いて7月22日から7月31日までの 10日聞にわたり、 第2節で採用した葉片前処理方法を行って酸素放出量を測定 した。 すなわち、 測定前日の午前9時に葉片を作成し、 人工気象室で葉片前処 理を1日間行った。 毎日午前9時から午後5時まで1時間ごとに(途中12時を
"回�
Table 2. Effects of pretreatment for leaf disks on o xygen evolution
Measurement Oxygen evolution (μm 01 0 ')どdm2どhr) time (0' clock) Pretreatment Pretreatment II
9 :00 a. m 546士 46 344 ::t 82
10:15 a.m. 553 ::t 120 382 ::t 41 11:15 a.m. 526 ::t 98 435 ::t 34 12:15 p.m. 506 ::t 28 488 ::t 60 15:30 p.m. 508 ::t 49 414士 38 16:25 p.m. 570 ::t 85 452士 54 17:10 p.m. 518 ::t 68 498 ::t 38
(Average) 532 430
t-va1ue
3. 73本 2.35n s 1. 53 n
O. 47 n 2. 63 n 2. 04 n O. 45 n
Pretreatment 1: Leaf disks were incubated in a growt h chamber for 24 hours under i 11 umi na tion from 6: 00 a . m. to 8 : 0 0 p. m. a t 2 8 oC d u r i n g d a y t i m e a n d 2 2 oC a t n i g h t.
The light intensity was 40 klux.
Pretreatment II Leaf disk were incubated in an air- c 0 n t r 0 1 1 e d r 00 m a t 2 5 oC f 0 r 2 4 h 0 u r s u n d e r i 1 1 u m i n a t i 0 n
from 6:00 a. m. to 8:00 p. m. The light intensity was 1 klux.
キ : Significant at the 5% level n. s. : Not significant.
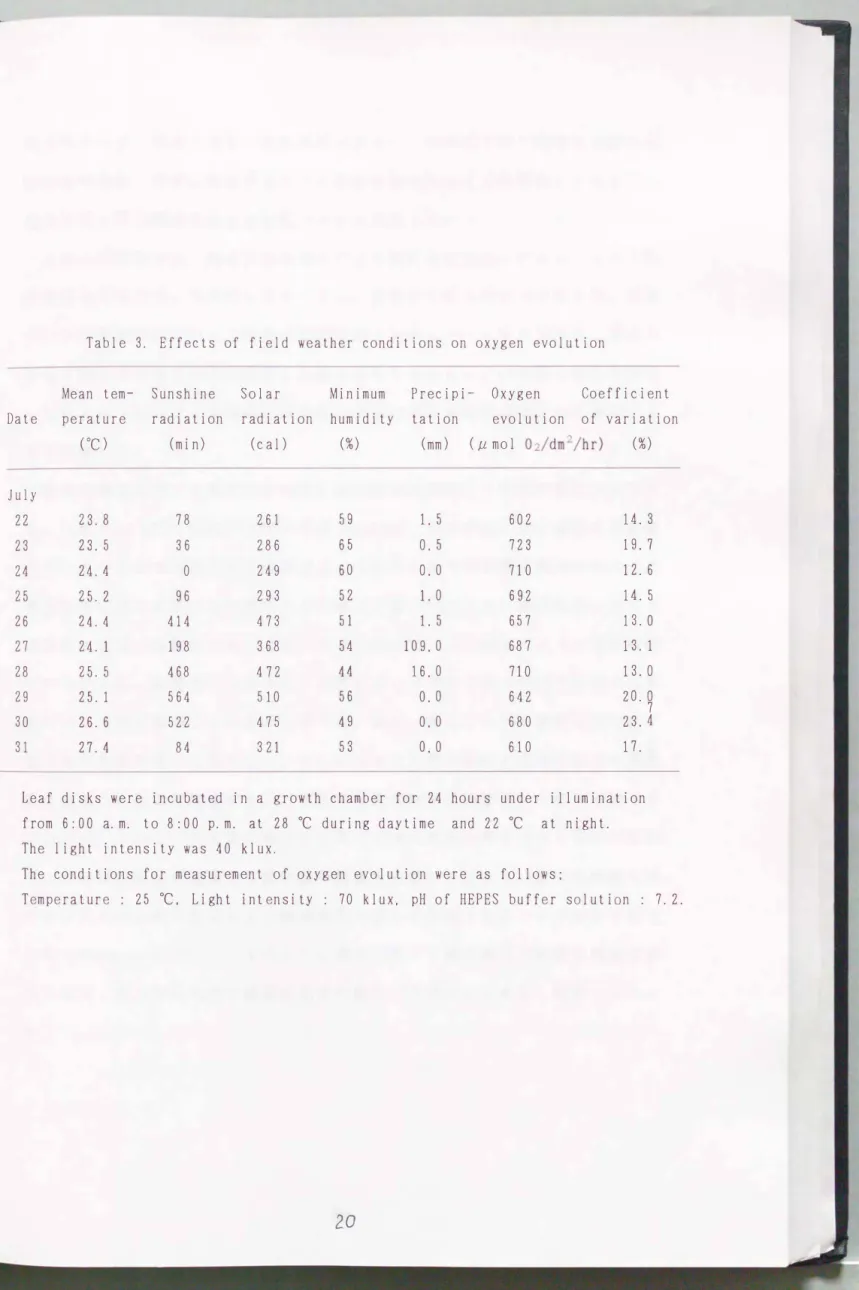
除く)8回、 温度250C、 光照度7万ル yクス、 HEPES緩衝液のpH7. 2の条件で測 定した。 圃場の気温、 日照時間、 日射量、 降雨量などの気象データは供試材料 を栽培する圃場に隣接する旧農業技術研究所物理統計部気象科の露場で測定さ
れたデータを利用した。
結果および考察
日平均気温は7月22日から徐々に高まった。 日照時間は0分(7月24日)か ら564分(7月29日)まで日によって大きく異なり、 日射量も249cal(7月24 日)から 510cal (7月29日〉まで変動した。 7月27日に109rnrnの大雨があった 以外は降雨は微量か無であった(Table 3)。 このように本測定期間は梅雨の 末期にあたり、 日照時間、 日射量の変動が著しかった。
1日8回の測定値を平均した日平均酸素放出量をみると、 最低値は6 02 μrnol
02/drn:ミ/hr( 7月22日)、 最高値は723μrnol 02/drn2/hr ( 7月23日)であった。
両者の差は121μrnol 02/drnι/hrと小さ く、 10日間の測定期聞をとおして放出量 は安定していた(Table 3)。 すなわち、 測定日ごとの酸素放出量の変動係数 の大きさをみると、 最小値は12.6%(7月24日)、 最大値は23.7%(7月30日) であり、 人工気象室で葉片をインキュベートする方法は、 測定材料を栽培する
圃場の気象条件にかかわらず安定した値をもたらすことがわかった。
小島
・
川島(1968)はダイズ品種で測定日の異なる個体聞の炭酸ガス交換速 度の変動係数は、 栽培条件が違っても5-25%の範囲にあることを報告した。 こ の値を他の農業形質の変動係数と比べると比較的小さい値であって、 光合成能力が品種の特性を示す一般的形質としてこれまでの形質と同等に扱うことがで きるとした。 移植栽培水稲における農業形質の変動係数は、 登熟歩合: 11. 8 %、
1穂、籾数: 15. 4児、 穂数: 23.8% (松島 1957)、 最長稗長: 4. 0%、 登熟歩合:
5. 5潟、 最長稗穂長: 6. 2 %、 1穂籾数: 8. 5 %、 穂、数14.4%、 穂、重: 15. 0児、 精玄米 重: 15. 7児
(楠田1990)などである。 すなわち、 長さに関する形質の変動係
19
Table
3.
Effects of field weather conditions on oxygen evolutionMean tem- Sunshine Solar Minimum Precipi- Oxygen Coefficient Date perature radiation radiation humidity tation evolution of variation
( OC
) ( m i n) ( c a1
)(%) (
m m ) (μmol 02/dmL/hr)(完)
Vぜ1i
nJ臼 円‘u an吐 Ed nhu 円t『
nxu n同υ nHU 4lム Hu nJ臼 nJ白 nJb n,b n,L nJb nJ臼 nJ白 円ペυ 円喝U
T『JV
no rhd a叫 つL 4性 1i rhU 41 Fb an
- - - - - - - - -
n‘u n匂U 44A phu a4A aA1 rhu
「「U n'hU 門I' nJb つL つL qL n,L ηL nL 円L n,L 円L
78 36 0 96 414 198 468 564 522 84
261 286 249 293 473 368 472 510 475 3 21
nHd rhu nHu nJU 41よ an吐 anuτ ρhu n同υ 円ぺU
「D PO RU Ed rh.u rhu aH2 phd aH2 rhd
Fhu r、u nHu nHu phu nHu nHu nHu nHu nHU 4li nu nU 1i 41ム nuυ nhu nHU nHU
nHU
nHU 4li
4,S4
602 723 710 692 657 687 710 642 680 610
円‘υ 円tf phv r「u nHU 41ム nHv
nuu円1144ふ
- - - - - - - - -
an吐 nud nJ臼 d乱A n叫U 円‘u 円喝u
nuu
n《U 門It 41ム 40ム 41ム 4lム 4lム 4tム 唱『ム n,L nJ白
4lム
Leaf disks were incubated in a growth chamber for
24
hours under illumination f r 0 m6: 0 0
a. m. t 08: 00
p. m. a t28 oC
d u r i n g d a y t i m e a n d2 2 oC
a t n i g h t.The light intensity was
40
klux.The conditions for measurement of oxygen evolution were as follows;
T e m p e r a t u r e :
2 5 oC,
L i g h t i n t e n s i t y :70
k1
u x , pH
0 fH
E P E S b u f f e r s 01
u t i 0 n :7. 2.
20
数は小さいが、 重量に関する変動係数は大きし1。 本実験で得た酸素放出量の変 動係数は穂、数、 穂、重、 精玄米重などの変動係数の大きさと差がなく、 これらの 農業形質と同じ精度で扱える形質であると判断した。
これらの結果から 、 葉身を採取後ただちに葉片を打ちぬいてシ ャーレに入れ た蒸留水に浮かべ、 光照度4万ル ックス、 照明が午前6時から午後8時、 昼温 が280Cで夜温が220Cの人工気象室で24時間インキュベー卜する方法は、 翌日の 午前9時から午後5時まで安定した値を与えるとともに、 10日聞にわたり安定 した値であったので、 多数の材料を 扱う実験の葉片前処理方法として適用でき ると結論した。
酸素電極法に用いる葉片の前処理lこ減圧脱気処理を行う方法が報告されてい る。 Ishii et al. (1977)と坂
・
千坂(1981b) の方法は小さい葉片を用いる ため、 そのままでは葉片が反応溶液上に浮いてしまうので減圧脱気処理により 葉片を反応液中に沈める必要がある。 坂・
千坂(1982) は、 減圧脱気したイネ の葉片 は室温に放置しておいても、 あるいは270C、 6, 000ル yクスの連続照明 下においても、 調整当日を含めて 2日間くらい は光合成能力の低下が認められ ず、 むしろ高くなることがあると述べた。 また、 種によって は調整後経時的に 低下するものが多いと報告した。 とくにイネでは葉片調整1日後に始めの速度 の2倍以上に高まり漸減するが、 葉片 調整後 72時間 は調整当日の値を下回らな いという。 しかし 、 本実験において は葉片lこ減圧脱気処理を行うと値が不安定 で、 中には切断面が浸潤して低い値の酸素放出量しか示さない葉片を観察した。コムギなどでも減圧脱気により酸素放出が著しく抑制されることが報告されて いる(MacDonald 1975)。 このように葉片に及ぼす 減圧脱気の影響は無視でき ないので、 葉片の前処理で減圧脱気は行わない方がよいと考え、 採用しなかっ た。
21
第4節 酸素放出量の最適測定条件の確立
酸素放出量の最適測定条件のうち、 葉片前処理方法などを確立したので、 つ ぎに照射光の照度、 温度、 HEPES緩衝液のpH、 炭素源としての炭酸水素ナトリ ウムの濃度、 葉身から葉片を打ち抜く部位などの検討に入った。
材料および方法
以下の実験ではいずれも葉片を人工気象室でインキュベー卜して前処理を行 っTこ。
最適測定照度
ランプと反応容器との聞の距離を集光器で変えて2万から10万ルックスまで 2万ルックスずつ増やし、 酸素放出量が光飽和となる光照度を調べた。 光照度 は照度計(東芝5号)で測定した。 暗呼吸は反応容器全体を黒色の暗幕でおお って測定した。 反応液の温度は250C、 HEPES緩衝液のpHは7.2とした。
最適測定水温
温度調節装置で水槽
(Fig.
2)内の水温を200Cから350Cまで5 oCずつ上げ て、 最大の酸素放出量となる温度を調べた。 反応は、 光照度を7万ルックス、HEPES緩衝液のpHを7.2として行わせた。 以上の2実験には1984年4月21日lこ播 種し、 5月1 6日に園場に1株1本植えで栽培したコシヒカリの第7、 8葉を用
し\ t.こ。
最適測定pH
HEPES緩衝液のpHをO. 1規定の水酸化ナトリウムで調節し 6. 6から7 . 6までO.2
ずつ上げ、 最適のpH条件を検討した。 反応液の温度は250C、 光照度は7万ルッ クスで測定した。
22
最適測定炭酸濃度
炭酸水素ナトリウムの反応容器内の最終濃度については、 o mMから50mMまで 10mMずつ変え最適の濃度条件を求めた。 光照度を温度を250C、 7万ルックス、
HEPES緩衝液のpHを7. 2として測定した。 以上の二つの実験に使用した材料は最 適温度条件の実験に供試したものと同じであるが、 第9 、 10葉を用いた。
最適葉身測定部位
葉身から葉片を採取する部位については、 1枚の葉身の先端部、 中央部、 基 部の3ヶ所から葉片を作成し、 温度を250C、 光照度を7万ルックス、 HEPES緩 衝液のpHを7 . 2で測定した。 実験には10枚の葉身を用いた。
直達光と葉片の表裏の関係
光源、からの直達光に葉片の表側を向けた場合、 裏側とした場合に酸素放出量 に差が生じるかどうかを調べるために、 1枚の葉身の中央部から隣接した2枚 の葉片を作成した。 葉片の表側あるいは裏側を照射光に向けて反応容器内に入 れ、 温度を250C、 光照度を7万ルックス、 HEPES緩衝液のpHを7.2で測定した。
実験は15反復で行った。 以上の二つの実験に使用した材料は最適温度条件の実 験に供試したものと同じであるが止葉を用いた。
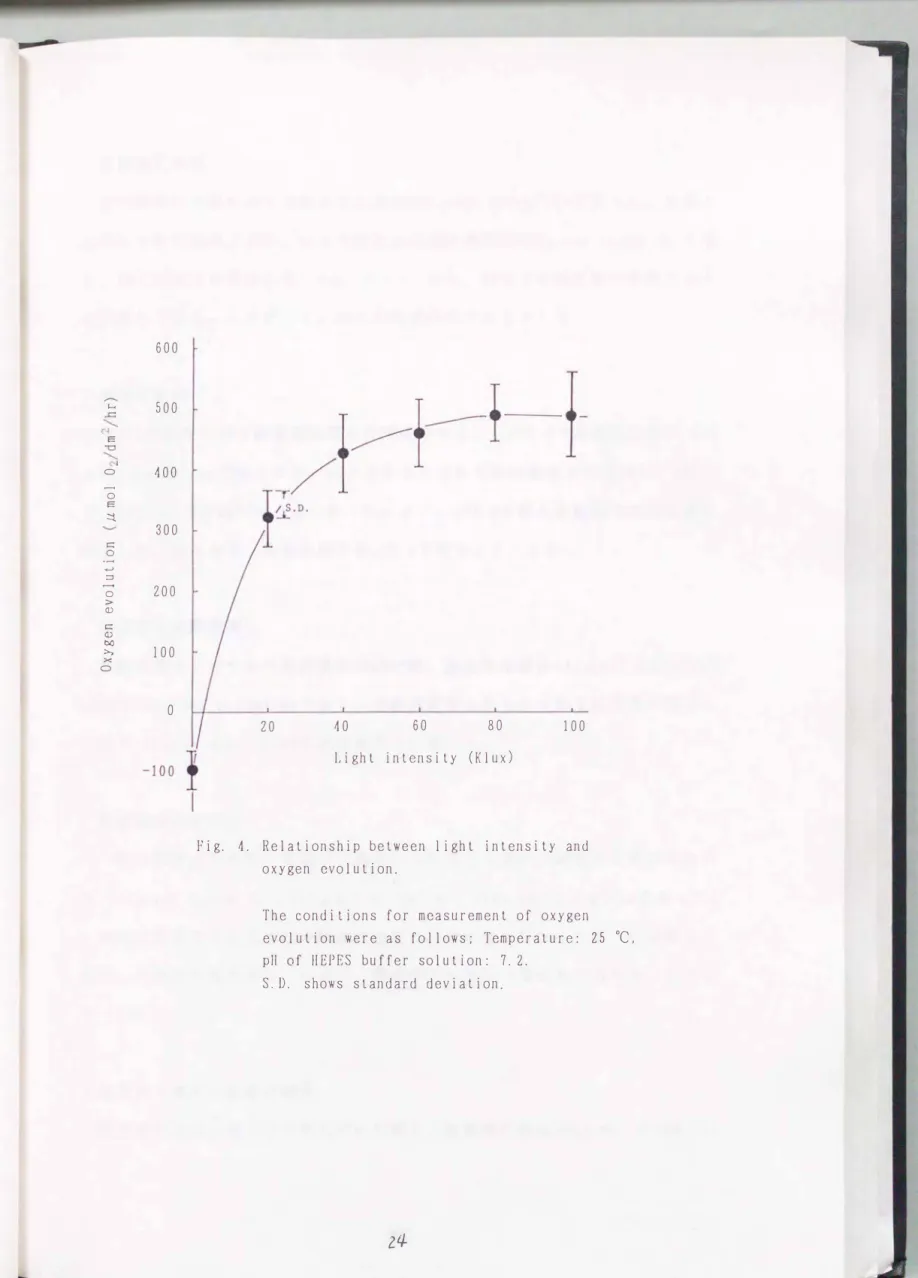
結果および考察 最適測定照度
照度2万ルックスでは酸素放出量は3 19μmol 02/dm 三/hrであったが、 光照度
が高くなるにつれて酸素放出量は増加し、 7万ルックスで最高値480μm01 02
/dmL/hr に達した(Fig. 4)。
また、 測定値は変動も小さく安定していた。 したがって、 今後は光照度は7万ルックスで測定することにした。
2.3
H r
\
」- J
コ\
E
\て r\
コ .,
Cコ
...
。 E三
とえ
亡
.。...
φJ :::J
陶ー...
。
〉 (1) c::
(1) 00
〉ョ x Cコ
600
500
400
300
200
100
。
20 40 60 80 100
Lighl intensity (Klux)
Fig. 4. Relationship belween lighl intensity and oxygen evolulion
The conditions for measurement of oxygen evolulion were as follows; Temperalure: 25 oc,
pl! of HEPES buffer solution: 7.2.
S. D. shows slandard deviation
24-
最適測定水温
反応溶液の水温が 200Cで酸素放出量は554μrn0
1
02/ drn L
/h rであった。 水温の 上昇につれて値は上 がり、 250Cで酸素放出量は最大の660 μrnol 02/drnL/hrに達 し、 300C以上では減少した(Fig. 5)。 また、 250Cでは測定値の変動が小さく安定していた。 したがって、 250Cが最適温度であると した。
最適測定pH
HEPES緩衝液のpHと酸素放出量と の関係をみると、 pH6. 6で酸素放出量は 542 μrnol 02/drn2/hrであったが、 pHが高まるにつれて放出量はやや増加し、 pH7. 2 で 629μrnol Ol/drn2/hrとなった(Fig.
6)
0 pH7.2を越える範囲では測定値は低下した。 以上から、 今後の測定はpH 7. 2で行うこと にした。
最適測定炭酸濃度
炭酸水素ナトリウムの最終濃度10rnMの時、 酸素放出量は641μrnol 02/drn三/hr、
20rnMで、639μrnol Ol/drnL/hrであり、 最終濃度が上がるにつれて放出量は減少し たので(Table 4)、 20rnMを測定濃度と結論した。
最適葉身測定部位
一枚の葉身の中央部、 先端部、 基部から作成した葉片の酸素放出量はそれぞ
れ 40 7μrnol Ol/drn2/hr 、 391μrnol Ol/drn2/hr、 362μrnol 02/drnl/hrであった。
t検定の結果では3部位聞の酸素放出量に有意な差はなかったが、 中央部から 作成した葉片の値が高かったので、 葉身の中央部から葉片を作成すること にし
Tこ(Table
5)
直達光と葉片の表裏の関係
直達光の方向に葉片の表側を向けた場合、 酸素放出量は369μrnol 02/drn2/hr、
2�
800
ぃZ\JE刀\付。
OEミ)
\
巴。 .._,
コ 400
0 〉 Q) c Q) 凶
〉予><
o
300
200
2S 30 3 5
T e m p e r a t u r e 0
f
r e a c t i 0 n s 01
u ti
0 n( oc )
Fig.
5.
Relationship belween temperature of reaction solution and oxygen evolution.The conditions for measurement of oxygen evolution were as follows; Light intensity:
70
klux, pH of HEPES buffer solution:7.2.
S.
D.
shows standard deviation.600
L iご -...
N E
てコ -...
N
ζ〉戸O 400 E丈一co?パザコ{O〉ω
200 Cωmh×O
7.4 7.6 7.2
6.8 7.0
。 6.6
solution pH of HEPES buffer
Rela l ionship bc l wccn pll of lIEPES buffcr solu l ion F i g. 6
evolu l ion
The condi l ions for measurcmen l of oxygen evolu l ion we r c a s f 0 1 1 0 w s . T c m p c r a l u r c: 2 5 oC
,
L i g h t i n t e n s i t y and oxygen70 Klux
Table 4. Relationship between final concentration of NaHC03 in a reaction cell and oxygen evolution
Final concentralion of NaHC03 (mM)
10 20 30 40 50
Oxygen evolution (μmol Ol/dmL/hr)
3 6 641 6 3 9 6 1 5 6 23 5 3 1
The conditions for measurement of oxygen evolution were as f 0 1 1 0 w s; T e m p e r a t u r e 2 5 oc I L i g h t i n t e n s i t y : 7 0 k 1 u X I
pH of HEPES buffer solution 7.2.
2Õ>
Table 5. Relationship between the position of leaf disks in a leaf blade and oxygen evolution
Position of disks in a leaf blade
Top Middle Base
Oxygen evolution (μmol 02/dm乙/hr)
391 ::t 87 407 ::t 89 362 ::t 46
The conditions for measurement of oxygen evolution were as follows;
T e m p e r a t u r e 2 5 oC, L i g h t i n t e n s i t y : 7 0 k 1 u x pH of HEPES buffer solution 7.2.
一方、 裏側を向けた場合は337μmol 02/dm2/hrであり、 表側がやや高い値であ った(Table 6) 0 t検定の結果では両者聞に有意な差はなかったが、 表側を 用いることにした。
以上の結果をまとめると、 イネ葉身酸素放出量の最適測定条件は照射光照度 7万ルックス、 水温250C、 HEPES緩衝液のpHは7. 2であった。 測定試料は葉身の 中央部から作成した葉片が放出量が高く、 また、 葉身の表側を用いるのがよか った。 以下の実験ではすべてこの条件で測定することにした。
イネでは同一葉身で中肋を含む葉片とそれを除いた葉片とで光合成能力に差 が認められない( 坂
・
千坂1981a)ので、 中肋を含む葉片を使用することにし た。 また、 生育時期が揃った葉身を容易に採取するため主稗の最上位の完全展開葉を用いることにした。 以下の実験ではすべてこの条件で測定することにし た。
本実験で得られたイネの酸素放出量の最適測定条件をこれまでの報告と比べ て以下考察する。 Ishii et al. (1977)は、 5万ルックス、 250C、 坂 ・ 千坂 (1981b)は、 6万ルックス、 300Cを最適条件としており、 本実験結果とはや や異なっていた。 Ishii et al. (1977)は面積 1mm 2という極小葉片を試料に 用い、 坂
・
千坂(1981b)は直径3 mmの葉片を減圧脱気した後で用いており、本実験で用いた葉片の大きさ ( 直径12mm)よりかなり小さし1。 最適測定条件の 若干の差異は葉片の大きさの違いによるか もしれない。 また、 Ishii et al.
(1977)と坂
・
千坂(1981b) は極小葉片を反応液中に浮遊させた状態で測定しているが、 本実験では面積の比較的大きな葉片をリーフホルダーで光源に対 して垂直に固定した状態で測定しているという違いもある。 伊藤(198 2)は葉 片の面積と酸素放出量との関係を検討し、 葉片の面積が2.5cm2以下では酸素放
出量は直線的に増加すると報告した。
新しく開発した多点式酸素放出量測定装置2台と葉片の前処理方法とを用い ることによって1日あたり150点の酸素放出量の測定が可能になった。 葉片試
--←
Table 6. Differenc e of oxygen evolution between adaxial and abaxial sides of a leaf disk
Position
Adaxial Abaxial
Oxygen evolution (μmo l 02/dm2/hr)
369士 67 337土64
The conditions for measurement of oxygen evolution were as follows;
T e m p e r a t u r e : 2 5 oC, L i g h t i n t e n s i t y
70 klux, pH of HEPES buffer solution : 7.2.
3/
料作成および葉片前処理の手順を述べると以下のようである。
①測定前日の午前8時に圃場で最上位完全展開葉をサンプリングして、 午前 9時に葉片を作成する。
②葉片をシャーレ内の蒸留水に浮かべ、 ふたをして、 光照度4万ルックス、
照明が午前6時から午後8時、 昼温が280Cで夜温が220Cの人工気象室で24時間 インキュベー卜する。
③葉片のインキュベートが終了後、 すなわち、 測定日の午前9時に人工気象 室からシャーレを取り出し、 実験室に搬入する。
④以後、 シャーレから逐次葉片を取り出し測定に供する。
1日あたり 150点の試料を処理できるということは、 従来の測定方法では困 難であった短時間における多点数測定を達成したことを意味する。 多くの材料 の測定を必要とする光合成能力の遺伝的変異や多数個体からなる雑種集団を用 いる遺伝分析が可能となり、 測定値の精度も高く、 かっ比較的容易に分析でき るシステムを構築したと考える。
伊藤(1982)は日本稲および外国稲14品種を用い、 7月22日から8月1日に 完全展開した上位1ないし2葉を着生のまま炭酸ガス交換速度を3から5反復 で測定した。 一方、 その同一葉から葉片を作成し、 酸素放出量を測定し、 両者 の測定値の関係を検討した。 その結果、 両者聞には1 %水準で有意な正の相関 (r=0.74料)を認め、
酸素電極法による葉身酸素放出量の測定が光合成能力の
推定に有力であることを示した。 このことは、 酸素電極法を用いた酸素放出量
の測定は従来の炭酸ガス交換法と相対的に同じ結果が得られることを示してい
る。第5節 生育時期別の酸素放出量の変化
以上の測定条件の解明により酸素放出量の分析条件はほぼ整ったが、 最後に 植物体の生育時期別に酸素放出量がどのように変化するかが未解決の問題とし て残った。 もし生育時期別に測定値に大きな変動があれば、 最高値となる時期 に測定することなどの措置が必要となるからである。 そ こで、 本節では生育時 期を主稗の葉令から追跡し、 生育時期別の酸素放出量の変化を明らかにしよう とした。
材料および方法
コシヒカリを1985年5月8日に播種し、 6月5日に4葉苗を園場に1株1本 植えで栽培した。 移植後、 旺盛に分けつを発生させた第7葉期から第13葉(止 葉)期まで酸素放出量を測定した。 測定部位は、 完全展開した最上位葉とした。
各測定時で1 2個体を測定し、 平均値をその生育時期の酸素放出量とした。 酸素 放出量の測定日をそれぞれTable 7に示した。 また、 各時期で測定に用いた最 上位葉の葉面積、 葉緑素含量をTable 7に示した。
結果および考察
コシヒカリの葉令別酸素放出量は第7葉期および8葉期がもっとも高く 930
,...., 910μmol 02/dm2/hrであった(Table 7)。 第10葉期までは直線的に低下し、
第10葉期から第13葉期までは値はやや低下するか、 横ばいとなった。 すなわち、
酸素放出量は生育時期の推移につれて変化し、 第9葉期以降は低下することが 明らかになった。 葉身の葉緑素含量も酸素放出量と同じように変化した。
生育時期の推移にともなう光合成速度の変化はイネ(Osada 1964、 坂1977、
伊藤19 82)、 コムギ(Murthy and Singh 1979)、 クワ(村上1978)、 ワタ (Jones and Osmond 1973)でも観察されている。 Osada(1964)はイネ4品種 を用いて移植後経時的lこ炭酸ガス交換速度を測定したところ、 移植後30日に最
Table
7.
Dependency of oxygen evolution on the lea f orderLeaf Measurement Oxygen Sta ndard Lea f a rea Ch lorophy ll
order date evol ut ion devi ation con ten t
(μmol 02!dm2/hr) (μmol 02!dm2 /hr) (cmつ (mg/g
F. W. )
Ju 1.
6 930 88 14.7 2. 80
8 11 910 131 21. 8 2. 97
18 784 72 30. 7 2. 60
10 24 608 125 38. 4 1. 70
11
Aug.1 595 92 4 5. 3 1. 46
12 556 120 1. 37
13
(f lag leaf)22 630 43 27. 5 2. 24
The conditions for measurement of oxygen evolution were as follows;
Temperature :
25 oC.
Light i ntensity :70
klu x. pH of HEPES buf fer solution :7.2.
一 Not measu red.
ミ与
高値となり、 徐々に低下することを観察した。 伊藤(1982)はポット栽培した イネ27品種の第3葉と第5葉の酸素放出量を測定した。 その結果、 第3葉が第 5葉より値が高く、 両者にはr=0.65*本の正の相関があった。 イネでは本実験や Osada (1964)や伊藤(1982)の実験結果から明らかなように生育の早い時期 に光合成能力が最大となるといえる。 伊藤はかなり早い生育時期でも酸素放出 量の測定は可能だが、 測定葉は材料や目的に応じて決定するのがよいとした。
本実験のように遺伝変異や遺伝分析のための多点数測定を目的とする場合には 圃場で栽培した材料が望ましく、 その際には移植後の活着が終了し、 分けつが 開始された7葉期ころが望ましいと考える。
コムギでも光合成能力は最高分けっ期で高く、 開花期になると低下し、 葉緑 素含量や酵素活性は高くなるという (Murthy and Singh 1979)。 クワで村上
(1978)は、 葉身の最大光合成能力は7月中旬から8月中旬までの比較的葉令 の若い葉にみられるとした。 一方、 Jones and Osmond (1973)はワタで第1葉 から第9葉まで測定したところ、 第7葉がもっとも高い値であると報告してい る。
このように葉令の推移につれて光合成能力が変化するので、 多数でしかも生 育時期が異なる個体を含む雑種集団を用いた光合成能力の遺伝分析や品種間変 異の実験にあたっては以下の点に注意をはらって行う必要がある。 すなわち、
第1に、 比較対象とする雑種集団や品種の測定には一定の葉令の材料を用いる ことである。 第2に、 イネは生育の比較的早い時期に高い酸素放出量がみられ るのでなるべく最高分げつ期までに測定を終了することである。
第4章 酸素放出量の種内変異
第3章で多数の材料の酸素放出量を簡便、 迅速、 安定して測定できる条件が 確立されたので、 本章ではイネ種内の酸素放出量の変異を調べた。
第1節 アジア栽培イネにおける酸素放出量の種内変異
イネの光合成能力の遺伝分析を行うためには、 栽培イネ種内における酸素放 出量に関する遺伝的変異を明らかにする必要がある。 栽培イネにはアジア栽培 イネの立主主主主s a t i v a L. と アフリカ栽培イネの立ニーglaberrirn a STEUD. の2種が ある。 本節では栽培面積が広く重要性が高い立ι S a t i v aを材料にとりあげた。
これまでの研究で、 O. s ativ aには品種群レベルでの分化が進んでおり、 古く から議論が多く数種の分類方法が提案されている。 ここでは、 形態形質やエス テラーゼアイソザイ ムの遺伝子型(Nakagahra 1978)などによって、 インデイ 力、 ジャポニ力、 ジャパニ力、 シニカ(中国柏稲)の4 品種群に分類して、 ア ジア栽培イネの酸素放出量の種内変異および品種群間変異を調べた。
材料および方法
農業生物資源研究所植物探索導入研究チームに保存されている品種の中から、
インディカ品種群に属する25品種、 ジャポニカ品種群25品種、 ジャパニカ品種 群25品種、 シニカ品種群16品種の合計91品種の在来品種を選んだ。 供試品種の 原産地と品種数は、 日本10、 北中国13、 南中国24、 フィリッ ピン2、 インドネ シア7、 ラオス5、 ミヤンマー( I日ビルマ)4、 タイ1、 パ、ングラテJシュ5、
インド15、 スリランカ5品種であった。 Append i x 1に供試品種名、 品種群お よび原産地を示した。 これらを198 6年5月8日に播種し、 6月10日に園場に1 株1本植えで栽培した。 すべての品種は茨城県つくば市観音台にある農業生物 資源研究所の圃場条件で出穂した。 測定時期は、 第1期: 198 6年6月30日--7 月6日(移植後20--26日〉 、 第2期: 7月21日--7月26日(同41日--46日)、
第3期: 8月11日--8月 2 3日(同6 2--74日)とした。 酸素放出量は1 品種あた り4個体について測定し、 平均値で表した。 測定には各時期ともに主稗 の最上 位完全展開葉を用いた。 また、 出穂期、 稗長、 穂長、 穂数、 ふ先色、 全性、 籾 長、 籾幅、 籾積、 籾比を測定し、 酸素放出量との関係を調べた。 Appendix 1 にそれぞれの品種の酸素放出量とその他の形質の測定値を示した。
結果および考察
前章第5節で観察された生育時期の推移による酸素放出量の変化は、 本実験 でも観察された。 すなわち、 9 1品種の酸素放出量の変異を測定時期別に最低値
~最高値で示すと、 第1期: 501--895μmol 02/dmL/hr、 第2期: 306--669 μmol 02/dm2/hr、 第3期: 193--547μmol 02/dが/hrであり、 前章第5節で確
認したように測定時期が遅れるにつれて酸素放出量は低下することがわかった
(Fig.7)。 そこで、 本実験において酸素放出量からみた光合成能力としては、
ほとんどの品種がもっとも高い値となった第1期の値を採用するのが妥当と考
えた。供試した品種の中でインディカ品種群に属するSudu Heenatiが最高値 895μ mol 02/dm2/hrであり、 一方、 ジャパニカ品種群のHaohaiは最低値501μmo1 02 /dm2/hrであった (Appendix 1)。
インディカ品種群の酸素放出量の範囲は最低値598μmol 02/dm2/hrから最高
値895μmol 02/dm2/hrで、あり、 最高値/最低値比は1. 50であり、 大きな変異が あった(Table 8、 9 )。 この品種群の平均値は724μmol 02/dm2/hrであり
、
t検定の結果ジャポニカ品種群より有意に低く、 一方、 ジャパニカ品種群より 有意に高く、 シニカ品種群とは有意な差がなかった(Fig. 8)。 ジャポニカ
品種群は平均で772μmol 02/dm2/hrであり、 4品種群の中でもっとも高かった。
この品種群の最低値は6 51μmol 02/dm2/hr 、 最高値は8 84μmol 02/dm2/hrであ り、 最高値/最低値比は1.36と小さかった。 t検定の結果、 ジャポニカ品種群
37
……… υ…
ハハハ一日 ………
問easurement
I
(30/Jun.-6/Ju1. )10
15
5
Measurement II (21/Jul.-26/Ju1. )
Ti、tlγ1i
・ Tinヨ
+しハハnH,I/ HU ρ」勺、Jv
e m2
HU門UJ ra-
s
,,l・、 、EEE・ MH11・ 『dnハρ」Jf' u
。
20
nu
、,aE'
0 20
。
nu
可EEEE
5 1 5
5
15 25
( 守-e
的ω「パ戸ωf'・Lの〉
トO Lω2Eコch←ohucωコσωLL
Changes of oxygen cvolulion wilh lhc growlh indigenous varielies of rice 91
slagc ln
r i g.
Table 8. Variations of oxygen evolution in four varietal groups of rice
Oxygen 551 601 651 701 751 801 851
evolution
(μmol 02/dmι/hr) 550 600 651 700 750 800 850 Total
Indica 。 25
Varietal Sinica1> 。 。 4 16
group Javanica 。 25
Japonica 。 。 。 4 25
Total 10 16 14 20 13 91
1) Chinese Hsien variety
a�
Table 9. Difference of oxygen evolution in four varietal groups of rice
R ice varietal group
Item Indica Sinica1) Javanica Japonica Total
umber of varieties 25 16 25 25 91
Range (μmol 02/dmι/hr) 598-895 644-811 501-799 651-884 501-895
Average (μmol 02/dmι/hr) 724 722 613 772 705
Variance (μmol 02/dm2/hr) 6,791 1, 781 3. 896 4, 931 8, 345 Highest/lowest ratio 1. 50 1. 26 1. 59 1. 36 1. 79
1) Chinese Hsien variety
(724)
n. s. **
*
* ;.ド
、、JJ円ペU41i phu ,,t‘、
、、‘,,,nJL n,L 円tIfl、
* **
Japonica
、、‘,,,円JU円,I円tt,,,‘‘、
ドig. 8. Diffcrcnces of oxygen cvolulion among four rice variclal groups by l-lesl.
Numerals in lhe parenlhcses show lhe averaged value of cvolulion
*, * *
vvv 'ttA 噌PSE&
円d 凸U
t v
ρu
・
'i-tiaELrA FU 円d ρu vv , nν・
円、unu pu
wn rAaTEL
nH po
pu -
ρv pu mw VV 4L ρ) pu -
LU wm
e -
nL
nH .パU DU nH TA 円d ρU 「i V品川
「i
「ヘυ 4L iA
n AU ρV 円A -hU FU ar'b a?tiv
・'EA
nH
pfl 門d +し
・2』
c a n
--l
円ku ri po
--・
1l nν・
cu
nu uu ub nU 4L
-l
r
O
F\U 円KO
n. s