多細胞性シアノバクテリアにみられる 自己組織的なパターン形成の解析
Analysis of Self-Organized Pattern Formation in Multicellular Cyanobacteria
2015 年 7 月
石原 潤一
Junichi ISHIHARA
多細胞性シアノバクテリアにみられる 自己組織的なパターン形成の解析
Analysis of Self-Organized Pattern Formation in Multicellular Cyanobacteria
2015 年 7 月
早稲田大学大学院 先進理工学研究科 電気・情報生命専攻
細胞分子ネットワーク研究
石原 潤一
Junichi ISHIHARA
1
第1章 序論 4
1-1) 研究の背景(生物でみられるパターン形成とその理論的枠組み)
1-2) 多細胞性シアノバクテリアのヘテロシスト分化パターン形成
1-3) 本論文の概要
第2章 ヘテロシスト分化パターン形成の理論的解析 11 2-1) 要旨
2-2) 序文
2-3) 実験材料と方法
2-4) 結果
2-4-1) 細胞系譜の解析
2-4-1-1) 環境変化に伴う細胞分裂への影響は、35時間目以降に消失した
2-4-1-2) 窒素源飢餓による撹乱が消失して以降、分裂周期がほぼ一定値に収束した
2-4-1-3) ヘテロシスト分化パターン形成のダイナミクスは、窒素源飢餓状態後63-65
時間目と69時間目以降で変化した
2-4-2) ヘテロシスト分化パターン形成の理論モデルの作成
2-4-2-1) セルオートマトンモデルの作成
2-4-2-2) 2段階線形マルコフ近似に基づく解析的な頻度分布の導出
2-4-2-3) 系譜解析から直接決定できなかったパラメータ値の推定 2-4-3) 初期ステージでの分化ダイナミクスの解析
2-5) 考察
2-5-1) ヘテロシスト分化が始まる前に、窒素源飢餓による分裂ダイナミクスへの撹乱
が消失することは、他の研究室の培養条件下でも当てはまるだろうか?
2-5-2) 2段階線形マルコフ近似で用いた仮定は、本研究で明らかにされた分化パター
ン形成ダイナミクスに通じている
2-5-3) 初期ステージでの分化モデル(モデル2)は、どのような分子ネットワークか
2 ら引き起こされるのか?
第3章 ラマン散乱分光計測によるヘテロシスト分化パターンの解析 63 3-1) 要旨
3-2) 序文
3-3) 実験方法と材料 3-4) 結果
3-4-1) Anabaenaのラマンスペクトル計測とそのバンドの帰属
3-4-1-1) 励起波長532、561、785 nmのレーザーを用いたラマンスペクトル計測
3-4-1-2) 励起波長785 nmのレーザーを用いた純物質のラマンスペクトル計測
3-4-2) ヘテロシストの非標識ラべリング 3-4-3) 主成分解析に基づく分化状態の議論
3-4-4) タイムラプス観察に基づく分化運命決定ダイナミクスの解析
3-5) 考察
3-5-1) ラマン散乱分光計測による非標識計測の意義を再考察する
3-5-2) 分化しかけた細胞が分裂しても,その履歴は娘細胞に引き継がれない
第4章 総合討論 93 4-1) 自己組織的なヘテロシスト分化パターン形成のメカニズムは、どのようなダ
イナミクスによって支えられているのだろうか
4-2) ラマン散乱分光計測により、数理モデルの仮定が現象論的に正しいことを実証した
引用文献 97
謝辞 101
3
研究業績一覧 102
4 第1章 序論
1-1)研究の背景
多細胞生物では、分化した細胞が特定の比率で出現するだけでなく、規則的な空間配置 をなすことで機能をもった形態が形成される。このように、空間的に一様なものが非一様 になるためには、外部から何らかの方向付けが行われるか、もしくは自発的に対称性が破 れて空間的非一様性が生じなくてはならない。前者について考えてみると、前後・左右・
背腹軸に対して対称的な形をつくる、もしくは複雑な形をつくるために、例えば母体内で の化学成分の勾配や、精子の侵入に伴う方向決定など、卵の対称性を破る内的・外的要因 が必要となる。Wolpertら(1)は、French Flag Modelにおいて「予め存在する化学成分 がシグナル分子として働き、その濃度に応じて遺伝子発現が誘導されて細胞の状態が変わ るのならば、結果的に空間パターンが形成される」とした位置情報理論を提唱した。実際、
この理論により、ショウジョウバエの胚発生など多くの発生現象の理解が進んだ。
しかし、このような考え方だけでは発生現象を説明するに不十分と言わざるを得ない。
生物の発生にみられるパターン形成現象の多くは、初期条件に必ずしも依存せず、外乱が 加えられても自律的に進行しうる。この自己組織化のプロセスを理解するため、初期条件 に拘束されず、内因性のノイズにもロバストネスを保ちながら形成される空間パターン現 象を実験対象とすべきだろう。具体的に、脊椎動物の体節形成(2)や、魚・動物の体表面 模様の形成(3)が格好の研究対象として挙げられる。
このような形態形成現象を理解するため、Turing(4)は2自由度の力学系を用いつつ、
拡散の存在によって一様な状態が不安定化し、自発的にパターンが形成される一般的な理 論を構築した。この理論では、自己増殖的な変数を活性因子、抑制作用をもつ変数を抑制 因子としたとき、後者の拡散が前者のそれよりも速ければ、対象性の自発的破れが引き起 こされ、結果的に活性因子の高濃度な領域が等間隔に出現することを示した(Turingパタ ーン)。ここで、Turing パターンの形成は反応と拡散の関数型にのみ依存するため、特別 な初期条件を必要とせず、さらに崩されたパターンを修正する調節能の説明にも都合がよ い。実際、熱帯魚をはじめとした動物の体表面の模様は、Turingパターンにより説明され
5
る可能性が高いといわれている(3)。その他にも、Gierer & Meinhardt モデル(5)な ど、反応拡散系に基づく数理モデルが数多く提唱されてきた。
これらの理論提唱から半世紀過ぎた現在、分子遺伝学的手法やライブイメージング技術 の進展に伴い、モデルの変数や条件設定を発生現象のダイナミクスと対応付けようとする 試みが増えてきた。例えば、ゼブラフィッシュ体表面でみられるストライプ模様は、メラ ニン保有細胞と黄色系細胞が接触し、前者が後者から逃避することで形成される(6)。こ の研究を通して、Kondo らは、魚の体表面でみられるパターン形成が、Turing モデルの ようにシグナル分子の拡散(遠距離相互作用)を仮定しなくても、細胞間の接触(近距離 相互作用)によって行われることを示した。このように、ストライプパターン形成に最も 重要なダイナミクスを定性的に検証できた理由は、数理モデルでのダイナミクスを念頭に 置くために、分子レベルの発現、分解、拡散に立ち入らず、モデルに合わせて少ない自由 度で実現象を説明できたことによるだろう。
しかし、生命現象の多くは、極めて多くの生体分子が相互作用するため、一般にモデル の自由度と現象の自由度が必ずしも一致せず、モデルの変数と分子の対応付けが難しい。
また、従来の遺伝学的手法では、生体分子を蛍光標識し、その蛍光量の空間分布や時間変 化を解析することで、標的分子の発現や分解、拡散を理解してきた。しかし、このような 標識化した計測の場合、本来の分子の性質や機能に与える影響や、蛍光量の上昇・減衰と 生体分子の発現・分解との時間差を無視できないため、例えばシグナル分子の拡散を伴う 細胞間の遠距離コミュニケーションなど、数理モデルとの対応付けが近似的にしか行われ ていない現象も残されている。したがって、数理モデルで仮定されたダイナミクスが、実 際の現象でも同じように適用されるのか、定量的に検証されてきたとは言い難い。
そこで、真に形態形成現象と数理モデルを対応付けるため、現在の計測手法によって解 析できるダイナミクスを反映させたモデルを作らなければならない。言い換えれば、計測 可能なスケール(階層)に沿ったモデルを作るべきだろう。私たちは、数理モデルとの統 合的な理解を可能にする系として、細胞分化を伴う単純なパターン形成(分化パターン形 成)を示し、さらに細胞が直鎖状に連なることで観察・解析が容易となる多細胞性シアノ
6
バクテリアを選んだ。本研究では、この分化パターン形成に必要な遺伝子ネットワークや 代謝経路に踏み込まず、各細胞の運命が決定される時間パターンを解析することで、顕微 鏡下で観察される分化パターン(形態変化)と定量的に結びつけやすい数理モデル作成を 目指した。これにより、一様な細胞集団から分化パターンが形成され始め、それが時間経 過とともに変化する現象のダイナミクスを明らかにする。数理モデルの作成と生命システ ムの解析を相互にフィードバックさせることで、多細胞性シアノバクテリアにとどまらず、
形態形成現象を広く捉えられる手法の提案を目標とする。
1-2)多細胞性シアノバクテリアのヘテロシスト分化パターン形成
シアノバクテリア(ラン藻)は、酸素発生型の光合成を行うグラム陰性細菌の一群であ り、酸素の発生・無機炭素の固定のみならず、空気中窒素の固定も行うことから、海洋・
淡水界や土壌における第一次生産者として重要な生態学的役割を果たしている。その生息 形態として、単細胞で浮遊する種(Synechococcus spp など)、少数細胞の集団を形成す る種(Mycrocystis spp など)、糸状に細胞が並んだ構造をもつ種(Phormidium spp な ど)、分岐する糸状性の多細胞体制をとる種(Nostochopsis spp など)、一部に特異的な細 胞を分化させる種(Anabaena spp など)などが報告されている(7)。
私が研究に使用したAnabaena sp. PCC 7120(以下Anabaena)は、アンモニウムイオ ンや硝酸イオンなどの窒素化合物が存在する培地にて、光合成を行う栄養細胞が直鎖状に 連なる多細胞性形態をとる(8、9)。Anabaenaを培地中の窒素化合物(窒素源)飢餓条件 下で培養すると、栄養細胞がほぼ10細胞間隔でヘテロシストと呼ばれる窒素固定細胞に分 化する。栄養細胞は細胞分裂を繰り返すが、ヘテロシストは細胞分裂を行わず、ヘテロシ スト間の栄養細胞数が増加すると、中間領域に新しいヘテロシストが分化する(8-10)。こ の繰り返しによって観察される形態を「ヘテロシスト分化パターン」とよぶ(図1A、B)。 窒素固定の触媒酵素(ニトロゲナーゼ)は、光合成によって発生する酸素によって失活す るため、窒素固定と光合成は同一細胞内で両立しない(8、9、11)。したがって、栄養細胞 はグルコースをヘテロシストに供給し、一方でアンモニウムイオンなどの窒素化合物をヘ
7
図 1 窒素源飢餓条件下でのAnabaena sp. PCC 7120 のヘテロシスト分化パターン形成。(A)
Anabaenaフィラメントの分化パターン形成の概略図。アンモニウムイオンや硝酸イオンなどの 窒素化合物が存在する条件下では、すべての細胞が栄養細胞である。培地から窒素化合物を除 去すると、窒素固定を行うヘテロシストがほぼ 10 細胞おきに分化する。四角内の概略図は、
栄養細胞とヘテロシスト間の代謝分業機構を示す。窒素固定に特化したヘテロシストは、周囲 の栄養細胞にアンモニウムイオンなどの窒素化合物を供給し、光合成に特化した栄養細胞から グルコースを提供される。(B)ヘテロシスト分化パターンの顕微鏡撮影画像(左:明視野画像、
右:フィコビリソーム蛍光画像)。ヘテロシストは肥大化し、フィコビリソーム蛍光が退色し た細胞として観察される(白矢印)。スケールバーは 10 m を表す。(C)ヘテロシスト分化に 関与する生体分子、関連遺伝子の発現カスケード。分化を促進する hetR 遺伝子、若いヘテロ シストで発現して隣接細胞の分化を抑制する patS 遺伝子、成熟したヘテロシストで発現して 隣接細胞の分化を抑制するhetN遺伝子の発現カスケードを、それぞれ赤、青、緑で示した。(D)
フィラメント内での HetR、PatS タンパク質の分布概略図。hetR-patS遺伝子のフィードバック がヘテロシストで発現するとき、ヘテロシスト近傍の栄養細胞では PatS タンパク質の分子数 が多くなるが、ヘテロシストから離れた細胞では HetR タンパク質の分子数が多くなるだろう。
8
テロシストから受け取る。このような両細胞間の代謝分業体制が、ヘテロシスト分化パタ ーンとして観察される(12-14)。
それでは、栄養細胞のみで構成された一様な個体(フィラメント)から、どのように最 初のヘテロシストが分化し、その後もほぼ等間隔にヘテロシストが分化し続けるのだろう か?まず、培地に含まれる窒素化合物が減少すると、各細胞内でα-ケトグルタル酸(2-OG)
が蓄積し、ntcA遺伝子、nrrA遺伝子の発現を介して、hetR遺伝子の発現が活性化される
(図1C)(15-18)。hetR遺伝子は、窒素固定に必要なnif遺伝子群の発現を促すため、分
化を促進する主要な遺伝子として知られている(8)。HetR タンパク質は、自己触媒的な フィードバックによってhetR遺伝子の発現をさらに活性化させるため、一過的にhetR遺 伝子の発現が高くなった細胞はますます分化しやすくなる(19)。一方、HetRタンパク質 は、hetR遺伝子の発現を抑制するpatS、hetN遺伝子の発現も活性化する。PatSタンパ ク質は 51 アミノ残基でコードされる低分子であり、さらにそのレセプターが栄養細胞で 発現することを根拠として(20)、PatSタンパク質がペリプラズム内を拡散し、栄養細胞 に取り込まれることが示唆されている(12、18)。一方、HetNタンパク質はケトアシル還 元酵素の一種であり、成熟したヘテロシストでのみ発現する(21)。PatSとHetNタンパ
ク質は、RGSGRペプチド鎖を共通して保有するため(22)、HetRタンパク質2量体がhetR
遺伝子のプロモーター領域に結合することを阻害し、(HetRタンパク質の)自己ポジティ ブフィードバック活性を抑制する(19)。Callahan ら(23、24)は、PatSや HetN タン パク質などの分化抑制因子がヘテロシスト近傍の細胞内で蓄積することにより、hetR遺伝 子はヘテロシストから離れた細胞ほど発現しやすくなると考えている(図1D)。このよう なhetR –patS遺伝子間の発現ネットワークは、Turingモデルの活性因子-抑制因子ネット ワークを想起させる(図1D)。ここで、本論文では、ヘテロシストから離れた細胞ほど分 化しやすくなる性質を「分化の距離依存性」と呼ぶことにする。
一方、ヘテロシストから供給される窒素化合物も、分化の距離依存性を実現する拡散性 の分化抑制因子の 1つとして考えられている(25-28)。つまり、ヘテロシストから離れた 細胞ほど、供給される窒素化合物の量が少なくなるため、任意の閾値を下回ると分化する
9
という「閾値モデル」が提案されてきた。Allardら(29)は、個々の細胞の生長率がラン ダムであることや、窒素化合物の消費が異なることを仮定し、これらがフィラメント内に 窒素化合物の勾配をつくることを理論的に示した。
このように、ヘテロシスト分化パターン形成には、分化関連遺伝子や代謝ネットワーク 間の相互作用が必要であると分かってきた(10)。しかし、ヘテロシストから拡散する(と 考えられている)低分子ぺプチドや窒素化合物が、どのように協調して細胞運命を決定す るかなど、未解明な問題も多い。これは、前述のように、生体内での分子間相互作用や低 分子の拡散性などを非標識で計測できないことに起因する。また、多くの先行研究では、
任意の時間に培養液から採取したフィラメントを対象としたため、各細胞の運命が決定さ れるまでの詳細な時系列プロファイルが存在せず、同一フィラメントで行うべき解析が滞 っていた。したがって、実際に観察・計測された連続的なダイナミクスの変化を、数理モ デルにフィードバックさせられなかったため、実験と理論の統合的理解が進まなかった。
そこで、Asaiら(30)は、独自に微小培養チャンバーを作製することで、顕微鏡下にて 同一フィラメントの長期間にわたる培養と連続観察を可能とした。この系を用いることで、
窒素源飢餓条件下での分化中枢遺伝子 hetRの発現、光合成活性の変化、細胞分裂および 分化に伴うパターン形成の間の関係について一細胞レベルで解析した。その結果、バクテ リアでは初めてとなる細胞系譜を作製し、ダイナミクスを定性的かつ定量的に描写するこ とができた。興味深いことに、細胞分裂や分化は、直前の分裂からの時間経過に依存し、
一度分化しかけた細胞が分裂すると、分化の進行が止まり、代わって隣の細胞が分化する 様子が捉えられた。このように、ヘテロシスト分化の位置情報は、細胞分裂という内因性 のノイズの影響を受けつつ、細胞間相互作用によって動的に調節されていることが示唆さ れた。
1-3)本論文の概要
本研究では、細胞系譜の解析結果や顕微鏡下での観察結果を取り入れた数理モデルを提 案する。このモデル作成により、一細胞レベルとフィラメントレベルのダイナミクスを定
10
量的に対応付け、母細胞の分裂を経た細胞の運命決定ダイナミクスと細胞間相互作用がど のように位置情報の形成に寄与するか考察した(第2章)。さらに、ラマン散乱分光計測に より、細胞内での生体分子の局在やその濃度を非標識で計測し、分化運命が動的に決定さ れる過程を、光合成系Ⅱへ光エネルギーを伝達するフィコビリソームの分解ダイナミクス を基に議論した。特に、分化しかけた細胞が分裂した後、その履歴がどのように姉妹細胞 に継承されるか解析することで、一細胞レベルで空間情報が形成されるメカニズムを考察 した(第3 章)。第4 章では、第2 章と第3 章で記述した研究成果を総括し、Anabaena が自己組織的にヘテロシスト分化パターンを形成する新しいメカニズムを提案した。特に、
第3章で得られた解析結果を踏まえ、第2章で述べた数学的な仮定が、実際の分化パター ン形成現象のダイナミクスに通じていることを示す。
11
第2章 ヘテロシスト分化パターン形成の理論的解析 2-1) 要旨
私は、細胞分化を伴う単純なパターン形成を示し、さらに細胞が直鎖状に連なる形態を とるため、観察・解析が容易な多細胞性(糸状性)シアノバクテリア Anabaena sp. PCC
7120(以下、Anabaena)を研究対象に選んだ。Anabaena を窒素源飢餓条件下で培養す
ると、栄養細胞のみで構成された状態から、ヘテロシストと呼ばれる肥大化細胞が等間隔 に分化した形態に変化する(ヘテロシスト分化パターン)。さらに、分化パターン形成が進 むにつれて、平均的なヘテロシスト間の細胞数が増加して、ほぼ一定値に収束する。私た ちは、顕微鏡下で同一フィラメントの長期培養と連続観察を行い、マクロな形態変化のみ ならず、一細胞レベルでの遺伝子発現や光合成活性の変化を解析してきた(30)。本研究で は、この先行研究で報告された細胞系譜を再解析し、ヘテロシスト分化の出現頻度やその バラつきを再現した数理モデルを作成することで、どのように分化パターンが形成され始 め、そして変化するか議論した。
はじめに、Anabaenaの生長と分化を模した1次元のセルオートマトンモデルを作成し、
数値シミュレーションを行った。このモデルでは、細胞が成熟する過程を表すため、「細胞 成熟度指数」というパラメータを導入し、この指数が大きくなるにつれて、分裂や分化の 頻度が増加すると仮定した。さらに、ヘテロシストが隣接細胞の分化を抑制する効果(距 離依存性の効果)も取り入れた。
このモデルには、細胞系譜の解析から決定できるパラメータと、実測できないパラメー タが含まれる。後者を推定するため、分裂を伴う細胞数の増加に対応しながら、新しいヘ テロシストの空間情報が形成されるダイナミクスを近似的に解析する数学的手法を考案し た。第1段階の解析として、個々の細胞の運命が独立に決定されるとき、細胞成熟度指数 の遷移ダイナミクスが線形マルコフ過程にしたがうものとして、その密度分布をマスター 方程式から計算した。第2段階の解析として、細胞成熟度指数の定常密度分布が得られた とき、ヘテロシスト間細胞数の遷移ダイナミクスが線形マルコフ過程にしたがうものとし て、その密度分布を同様にマスター方程式から計算した。これらの線形マルコフ近似に基
12
づく解析を統合することで、シミュレーションから得られたヘテロシスト間細胞数の密度 分布を再現できた(「2段階線形マルコフ近似」)。
最後に、この解析的に得られた密度分布と、顕微鏡観察から得られたヘテロシスト間細 胞数の密度分布を比較し、その統計的距離が最小となるパラメータ値を推定した。このパ ラメータ値を、分化が細胞成熟度指数に依存しないモデルに代入することで、時間経過に 伴って変化する分化パターンを再現できた。実際、先行研究(30)のタイムラプスイメー ジングビデオから撮り卸した画像を解析すると、分化促進遺伝子 hetR の発現は、モデル と同じように細胞の成熟レベル(分裂後の時間経過)に依存することが明らかとなった。
このように、一細胞レベルのダイナミクスと分化パターンの変化を時間的に対応付けるモ デルを作成することで、分化の細胞成熟度指数依存性のみが時間変化するという新しいメ カニズムを導出した。
2-2) 序文
私たちは、窒素源飢餓条件下で Anabaena の同一フィラメントを連続観察し、hetR 遺 伝子の発現や分裂・分化ダイナミクスを細胞系譜として書き下した(30)。この系譜には、
運命決定のタイミングや、そのときのフィラメント内での位置情報など、全細胞の時空間 ダイナミクスが記述されている。
この先行研究(30)にて、私たちは、平均的なヘテロシスト間細胞数が時間経過ととも に変化し、やがて10細胞程度に収束すること(後期状態)を報告した。つまり、ヘテロシ スト分化が始まったときに観察される分化パターン(初期状態)は、後期状態に観察され る分化パターンと異なることが示唆された。実際、Yoonら(25)やToyoshimaら(31) も、初期にみられる分化パターンではヘテロシスト間細胞数が少ない(=ヘテロシスト数 が多い)ことを報告している。この現象について、Toyoshimaら(31)は、分化に必要と なるエネルギー量が大きいことを挙げ、時間経過に伴ってヘテロシスト数を必要最低限に 調整することで、窒素源飢餓条件での生長を最適化すると考えている。
Asai(30)は、ヘテロシスト分化パターンが変化する理由の手がかりとして、初期状態
13
では後期状態に較べて多くの細胞で hetR 遺伝子の発現が活性化されることを見出した。
特に後期状態では、hetR遺伝子の発現が細胞の成熟レベルに依存し、分化が細胞周期の中 盤以降に起こることを明らかにした(30)。しかし、初期状態でのhetR遺伝子の発現と細 胞の成熟レベルとの関係性は十分に解析されなかったため、分化の運命決定が後期に較べ てどのように変化するか議論できなかった。本研究では、分化パターン形成が始まる以前 の各細胞の振る舞いにも注目しつつ、窒素源飢餓後の時間経過に伴う運命決定と成熟レベ ルの関係性の変化を解析し、ヘテロシスト間の細胞数が時間変化するメカニズムと対応付 ける。
ここで、分化パターン形成が初期と後期で異なるメカニズムについて、他の研究グルー プによって提案された仮説を参照してみたい。例えば、Borthakurら(23)は、分化直後 の若いヘテロシストからPatSが拡散し、一方の成熟したヘテロシストではHetNが代わ って発現するとしたうえで、分化パターン形成の進行に伴って成熟したヘテロシストの数 が増加し、主要な分化抑制因子が置き換わることで、分化パターンの等間隔性が変化する と提案した。しかし、PatS や HetN タンパク質の拡散性を直接的に計測した実験がない ため、これらのタンパク質がどのようにヘテロシストからの距離に依存して分布している か定かではない。また、前述のとおり、これらのタンパク質は同一のRGSGRアミノ鎖に よってhetR 遺伝子の発現を抑制する(22)。したがって、PatSとHetNの拡散性や分化 抑制機能の違いを定量的に比較検討できない段階で、分化パターン形成ダイナミクスの変 化をこれらのタンパク質に求めることは難しいだろう。
また、多くの先行研究では、初期と後期にみられる分化パターンが区別して扱われ、特 に後者が研究対象とされてきた。そのため、栄養細胞のみで構成される均質なフィラメン トから、どのように初期条件がつくられて最初のヘテロシストが分化するか、そもそも議 論されることが少なかった。これは、前述のように、窒素源飢餓条件下で培養し始めた後 の分化パターン形成が連続観察されてこなかったことにもよる。
そこで、本研究では、先行研究の細胞系譜を再解析した後、ヘテロシスト分化の割合や その空間的なばらつきを再現する数理モデルを作成し、分化パターンが窒素源飢餓条件下
14
の初期と後期で変化するメカニズムを考察した。特に、細胞系譜で詳細に記述された各細 胞の分裂・分化のタイミングやそれらの周期に焦点を当てることで、細胞の運命決定と細 胞成熟度指数の関係に注目し、さらにこれらがパターン形成に伴ってどのように変化する か解析した。併せて、フィラメントを窒素源飢餓条件下で培養した後、細胞分裂の細胞成 熟度指数依存性がどのように変化するか解析し、分化パターンが形成され始めるときの各 細胞のダイナミクスを明らかにした。これらの解析を通して、一細胞レベルの運命決定ダ イナミクスと分化パターンの変化を時間的に対応付けることとした。
2-3) 実験材料と方法
・ Anabaenaの培養方法
本研究では、分化促進遺伝子hetRのプロモーター領域にgfp mutant2遺伝子を連結し たPhetR::gfp株(SRhetR-1Gn株)を使用した(30)。液体培養には、窒素化合物が含まれ るBG11液体培地か、窒素化合物が含まれない(窒素飢餓条件)BG110液体培地を使用し た(32)。これらの培養液にAnabaenaを懸濁し、30℃に保ちながら、45 m-2s-1 の連続 光条件下で震盪培養を行った。寒天培地を用いた静置培養には、まず BG11 液体培地で OD730=0.2 まで震盪培養し、その培養液を BG110 液体培地で 4 回洗浄し、最終的に
OD730=0.01まで希釈したうえでBG110寒天培地の下に流し込んだ。そして、この寒天培地
を、30℃かつ50 m-2s-1(FL30SW-B、日立製作所)の連続光条件下に置いた。
・ Anabaenaの観察方法
ヘテロシスト分化パターンの観察に、Olympus IX-71倒立顕微鏡(20× LUCPlanFLN 対物レンズ、分解度0.45、オリンパス)を使用した。ランプやミラーユニットの交換、観 察対象(寒天培地)のステージ上での位置調整は、Slidebook4.1(Intelligent Imaging
Innovations、Denver Co.)による自動制御を利用した。明視野および蛍光撮影に、
Slidebook4.1によって制御されるCCDカメラ(PIXIS1024、Princeton Instruments)を 使用した。GFP 蛍光とフィコビリソームタンパク質の蛍光観察に、それぞれ U-MNIBA3
15
とU-MWIG3フィルターセット(ともにオリンパス)を使用した。
・ 細胞系譜(Asai et al., 2009、Figure S2A)の再解析方法
本研究で再解析した細胞系譜は、Asai et al., 2009 (30)で発表されたFigure S2Aであ る。また、タイムラプスイメージングビデオから撮り卸した画像は、Asai et al., 2009 (30)
で発表されたMovie S1を時間ごとにTIFFファイルに変換したものである。この画像フ ァイルと細胞系譜を1時間ごとに照合しつつ、分裂や分化が起きたと判断されるタイミン グを見直した。
・ 統計解析手法(33-35)
2 標本間の母代表値に差が認められるか(統計的有意差)を判定するため、直前の細胞 分裂からの経過時間の密度分布、分裂周期の密度分布、本研究で得られた顕微鏡観察に基 づくヘテロシスト間細胞数の密度分布に、それぞれ2標本間コルモゴロフ-スミルノフ検定 を行った(33)。ここで、帰無仮説 H0を「2 標本の母代表値に差はない」として、有意水
準0.1%(α=0.1)で両側検定を行った。本研究では、標本分布間の代表値の差だけでなく、
分散や非対称度の差も判定するため、両側検定を行うことにした。
まず、各密度分布の累積分布関数 Sni (X)(i=1、2)の差分を計算し、その最大絶対値を Dとする(式1)。
D= maximum |Sn1(X) - Sn2(X)| (式1)
ここで、それぞれのサンプル数が40以上存在するとき、K値が自由度2のχ二乗分布に したがうとして、p値を計算した。
K= 4D2 n1n2
n1+n2
(式2)
16
ただし、このような総当りの検定の場合、各標本間での検定結果が最終的に「かつ」で 結ばれるため、1 組でも帰無仮説が棄却されてしまうと、全体として帰無仮説が棄却され ることになる。そのため、有意水準 α をあらかじめ小さくしておく必要がある(「多重比 較の問題」)。そこで、ボンフェローニの補正(34)に基づき、(検定全体の)有意水準 α を検定数(サンプル数)で割った値を有意水準α’とした。したがって、直前の細胞分裂か らの経過時間の密度分布を検定したとき、検定数が 9 個となったため、有意水準 α’を 1.11×10-2(=α/9)とした。また、分裂周期の密度分布を検定したとき、有意水準α’を2.50×10-2
(=α/4)とした。このことを踏まえ、帰無仮説H0の採否を決めるとき、p<α’ならば帰無仮 説を棄却し(=「2標本の母代表値に差がある」)、p≥α’ならば帰無仮説を採用した(=「2 標本の母代表値に差がない」)。
一方、サンプル数(標本数)が40未満の密度分布を使用して、代表値の有意差を検定す
るとき、welchのt検定を行った(35)。細胞系譜に記載された各時間でのヘテロシスト間
細胞数の密度分布と、顕微鏡観察から得られたヘテロシスト間細胞数の密度分布との有意 差を検定するとき、帰無仮説H0を「2標本の母平均値に差はない」とし、有意水準0.05%
(α=0.05)で両側検定を行った。帰無仮説H0の採否を決めるとき、p<α’ならば帰無仮説を
棄却し(=「2 標本の母平均値に差がある」)、p≥α’ならば帰無仮説を採用した(=「2 標 本の母平均値に差がない」)。分裂1 時間前の PhetR::gfp シグナル強度に対する分裂前後の シグナル強度比を検定するとき、帰無仮説H0を「2標本の母平均値に差はない」とし、有
意水準0.01%(α=0.01)で両側検定を行った。帰無仮説H0の採否を決めるとき、p<α’なら
ば帰無仮説を棄却し(=「2 標本の母平均値に差がある」)、p≥α’ならば帰無仮説を採用し た(=「2標本の母平均値に差がない」)。31の細胞系譜を解析することで得られた、直前 の細胞分裂からの経過時間の密度分布を、決められた計測時間ごとに総当たりで検定する とき、帰無仮説H0を「2標本の母平均値に差はない」とし、ボンフェローニの補正に基づ いて有意水準を1.11×10-2%(α=1.11×10-2)とした。帰無仮説H0の採否を決めるとき、p<α’
ならば帰無仮説を棄却し(=「2 標本の母平均値に差がある」)、p≥α’ならば帰無仮説を採 用した(=「2標本の母平均値に差がない」)。
17
・ 数値シミュレーションおよび解析に使用したプログラム
セルオートマトンモデルの数値シミュレーションは、C言語で書かれたソースコードを
使用し、VMWare Player(VMWare. Inc)にて実行した。解析解の導出には、C言語プロ
グラミングとMathematica 6(Wolfram Research)を併用した。t検定によるp値の導 出には、R(ver. 3.2.0)とExcel 2010(Microsoft)を使用した。
・ 数値シミュレーションの方法
数値シミュレーションの時間発展に、モンテカルロ法を用いた。フィラメント内のどの 細胞の状態を更新するか決めるため、ランダム・シークエンシャル・アップデートにより 細胞を選んだ。時間の刻み幅Δtは、α ∙ r(i)⁄n(α:モデルでフィラメントの全細胞が細胞 成熟度指数を1つ更新するためにかかる(ことに対応する)観察上の実時間(細胞周期13 時間を細胞成熟度指数10で規格化するので、細胞成熟度指数が1つ増えることに対応する 実時間は1.3 時間)、n:その時のフィラメント内の細胞数)により与えられる。モデル 1 より、細胞分裂・分化・成熟頻度は、それぞれ
p(i)Δt 、 q(i)Δt 、 r(i)Δt
により与えられる。この数値シミュレーションでは、窒素源飢餓条件に移して以降のヘテロシスト分化パタ ーン形成を再現する。初期状態として、フィラメントに含まれる細胞数と各細胞の細胞成 熟度指数をランダムに与え、分化細胞は存在しない。1 フィラメントに含まれる細胞数は 最高5000まで増え、それを10フィラメント発生させた。5000細胞に達する前に全ての細 胞が分化した場合、新しいフィラメントでシミュレーションを行った。
この一連の方法により、同一のキネティクスパラメータのもとで、常に有意差のないヘ テロシスト間細胞数の頻度分布が得られた(p>0.95、2 標本間コルモゴロフ・スミルノフ 検定(両側))。
・ カルバック・ライブラー情報量の計算
パラメータの値を推定するため、顕微鏡観察により得られたヘテロシスト間細胞数の頻
度 g(i) と解析解 f(i│θ) (θ = (a,b,c,d,e,f)、i: ヘテロシスト間細胞数)から、カルバック・
18 ライブラー情報量 I(g;f) を計算した(式3)。
I(g;f) = ∑i=mi=1 g(i) ∙Logf(i |θ)g(i)
= ∑ ni
N
i=mi=1 ∙Log(nNi) − ∑i=mi=1 nNi∙ Log f (i|θ) (∵g(i)= nNi , N=∑i=mi=1ni) (式3)
ここで、右辺第2項に含まれるni は、ヘテロシスト間細胞数iの観察個数を表し、Nは ヘテロシスト間細胞群の観察総数を表す。また、ヘテロシスト間細胞数の最大値 m を 50 とした。
右辺第2項は平均対数尤度 EG[Log f(X│θ)] を表すので、対数尤度L(θ) は
L(θ) = ∑i=mi=1 ni∙ Log f (i |θ) (= N ⋅ EG[Log f(X│θ)] ) (式4)
と表すことができる。したがって、前述のとおり、カルバック・ライブラー情報量を最小 とするパラメータ値は、(平均)対数尤度を最大とするパラメータ値に一致する。
・ MCMCサンプリング法による事後確率の導出
実測できないパラメータの値を推定した後、全てのパラメータ値の事後確率を求めた。
事後確率 Pr(θ│data ) は、ベイズ理論に基づき
Pr(θ│data )= Pr(data│θ)∙Pr(θ)
∫Pr(data│θ )∙Pr(θ) dθ ∝ Pr(data│θ )∙Pr(θ) (式5)
と表される。ここで、Pr(θ) は事前確率を表し、∫Pr(data│θ )∙Pr(θ) dθ により規格化した。
また、Pr(θ) は一定値と仮定した。
事後確率に必要なデータサンプルは、以下のようなメトロポリス法によって求めた。ま ず、aのデータを得るため、他のパラメータを推定値で固定して、a ∈[0.01, 0.1] からラン
19
ダムに1つの値を選んだ。次に、a + 0.01 か a - 0.01の値をランダムに選び、これを anew とした。L(anew) が L(a) より大きいとき、aの値をanew に変更したが、L(anew) が L(a) よ り小さいとき、exp(L(anew)-L(a)) の確率でaの値をanew に変更した。この仮定を1×103回 繰り返した。
b、c、d、e、f のデータを得るときも、同様の手順を用いた。このとき、b ∈[0.1, 1]、
c ∈[0.05, 0.95]、d ∈[0.5, 1.6]、e ∈[5, 30]、f ∈[3, 10] の範囲から、b の値を0.1 間隔ずつ、
cの値を0.05間隔ずつ、dの値を0.1間隔ずつ、e、fの値を1間隔ずつ動かした。
・ PhetR::gfpのシグナル強度の解析
PhetR::gfpのシグナル強度を一細胞レベルで解析するため、Image Jソフト(NIH)を使 用してタイムラプス動画を再解析した。この画像データは、タイムラプスイメージングビ デオから撮り下ろしたものである。個々の細胞の輪郭をトレースし、面積当たりの平均 GFP蛍光強度“FL”と、その周囲のバックグラウンド蛍光強度“BG”を計測した。平均 GFP 蛍光強度からバックグラウンド蛍光強度を差し引いた蛍光強度“FS”を、細胞由来
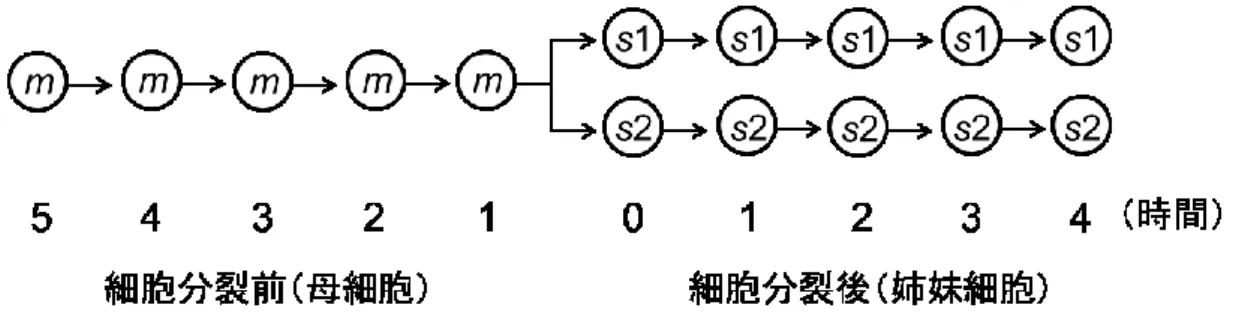
のPhetR::gfp 蛍光強度と考えた。ここで、分裂前後の細胞の系統(母細胞と2 つの姉妹細
胞)をサンプルラインと呼び、分裂前後のhetR遺伝子の発現ダイナミクスを解析した(図 2)。サンプルラインの数Nは、分裂した母細胞の数に等しく、63時間目に21、64時間目
に14、65時間目に12、69時間目に18、70時間目に14となった。分裂i時間前の母細胞
の蛍光強度を “FS m-i, j”( j: サンプルラインの No. 、j ∈[1, N])と表し、分裂 i 時間後の 姉妹細胞の蛍光強度をそれぞれ“FS s1-i, j”、“FS s2-i, j” と表した。
まず、分裂1時間前の母細胞の蛍光強度 FS m-1, j に対し、それ以前の母細胞の蛍光強度
FS m-i, j(i >1)と、同じサンプルラインの姉妹細胞の蛍光強度(FS s1-i, j,FS s2-i, j)の比率を
それぞれ求めた。
20 rm-i, j = FSm-i, j⁄ FSm-1, j
rs-i, j = FSs-i, j⁄ FSm-1, j (sは s1かs2) (式6)
次に、細胞分裂が63、64、65、69、70時間目に起きたそれぞれのサンプルラインにて、
分裂前後の同じタイミングでの比率を異なる細胞間で平均化した。
r m-i = N1(∑j=Nj=1 rm-i, j)
r s-i = 2N1 (∑j=1j=N rs1-i, j+∑j=Nj=1 rs2-i, j) (式7)
21
図 2 細胞分裂前後の母細胞と娘細胞の概略図。母細胞を m と呼び、2 つの姉妹細胞をそれぞ れs1、s2と呼ぶ。分裂前後の時間経過を各細胞の下に示した。
22 2-4) 結果
2-4-1) 細胞系譜の解析
私たち(30)は、先行研究にて、PhetR::gfp 株を顕微鏡下で培養し、4つのAnabaena フィラメントのヘテロシスト分化パターン形成を観察した。今回の研究では、そのうち 1 つのフィラメントを対象とし、細胞系譜(30のFigure S2A)とタイムラプスイメージン グビデオ(30のMovie S1)から撮り卸した画像を使用することで、細胞分裂とヘテロシ スト分化の時空間プロファイルを再解析した(図3)。このとき、改めてタイムラプスイメ ージング画像を見直し、細胞系譜での分裂や分化のタイミングを修正した。このフィラメ ントは、窒素源飢餓条件に移した直後(窒素源飢餓後0 時間目)、24個の細胞から構成さ れ、窒素源飢餓後72時間目に465個まで増加する(39個のヘテロシストを含む)。また、
窒素源飢餓後63時間目に最初のヘテロシストが分化し、窒素源飢餓後69時間目に平均ヘ テロシスト間細胞数がほぼ一定の値に収束した(30)。
2-4-1-1) 環境変化に伴う細胞分裂への影響は、35時間目以降に消失した
まず、窒素源飢餓条件後の時間経過に伴い、個々の細胞分裂のタイミングがどのように 変化するか解析した。この解析では、窒素源飢餓条件下で培養し始めたことによる影響が、
どのくらいの時間で消失したのかを決定する。細胞分裂が起きたタイミングは、私たちの 先行研究と同様に,細胞隔壁が形成されたときとした(36、37)。
Anabaenaを窒素源飢餓条件下で培養し始め、その直後からの決められた時間において、
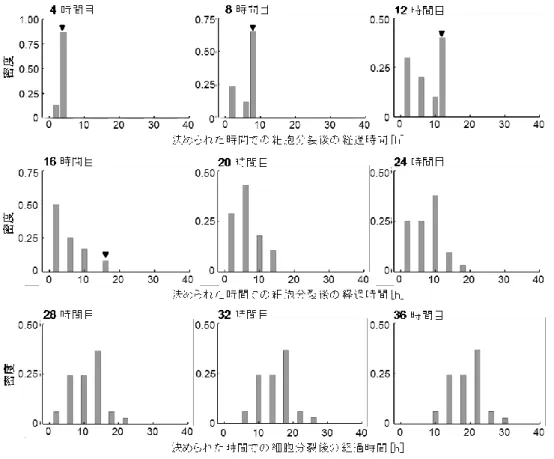
直前の細胞分裂から経過した時間を細胞ごとに計測した(図3C)。この経過時間は、計測 したそれぞれの時間における各細胞の成熟レベルを表している。なお、窒素源飢餓条件下 で細胞分裂が確認されていない細胞では、代わりに同条件下に移してからの経過時間を計 測した。また、分化が確認されたタイミングで、その細胞を計測対象から除外した。私は、
窒素源飢餓後30 時間目以前に分裂した細胞が少なかったことを鑑みて、窒素源飢餓後30 時間目から窒素源飢餓後70時間目まで5時間おきに計測を行い、分裂の密度分布を得た(図 4)。
23
図 3 細胞系譜と,本研究で再解析した「直前の細胞分裂から経過した時間」と「分裂周期」
の算出方法の概略図。(A)窒素源飢餓条件下での同一フィラメントの細胞分裂と細胞分化を記 述した細胞系譜(Asai et al., 2009 を改変)。分岐したところが分裂を表し,橙色で示した細 胞がヘテロシストを表す。(B)PhetR::gfpシグナルを一細胞ごとに計測し,その蛍光強度を(A)
の細胞系譜に重ね合わせた細胞系譜(Asai et al., 2009 を改変)。白色が強くなるにつれ,
PhetR::gfpシグナルが強くなることを示す。(C)図 4 に示す、決められた時間で計測した「直前 の細胞分裂からの経過時間」の計測方法。細胞が tc1時間目で分裂したとき,窒素源飢餓状態 後からt2,t3時間目での「分裂後の経過時間(h)」をd2(=t2-tc1),d3(=t3-tc1)とした。こ こで,窒素源飢餓条件下で分裂していなかった場合,窒素源飢餓状態後からの経過時間t1を分 裂後の経過時間d1とした。(D)図 5 に示す「分裂周期」の計測方法。細胞がtc1,tc2時間目で 分裂したとき,分裂周期の時間(h)をc2(=tc2-tc1)とした。ここで,最初の分裂が観察され るまでの時間をc1(=tc1-0)とし,第 0 世代とした。
24
図 4 決められた時間で計測した「直前の細胞分裂からの経過時間」の密度分布。(A)窒素源 飢餓条件下で培養してから 30、35、40、50、60、70 時間目に計測した密度分布。窒素源飢餓 後 30 時間目に 40 細胞、35 時間目に 48 細胞、40 時間目に 71 細胞、45 時間目に 92 細胞、50 時間目に 132 細胞、55 時間目に 174 細胞、60 時間目に 235 細胞、65 時間目に 329 細胞、70 時 間目に 404 細胞を計測対象とした。計測した時間に細胞分裂が確認された場合、姉妹細胞の時 間経過を 0(時間)とした。窒素源飢餓後 30、35 時間目の分布には、窒素源飢餓条件下での細 胞分裂が確認されなかった細胞が含まれる(黒矢印)。
25
これらの標本分布の母代表値に統計的有意差が認められるかを判定するため、窒素源飢 餓後30時間目以降の分布に総当たりで2標本間コルモゴロフ-スミルノフ検定(両側)を 行った(表1)。その結果、窒素源飢餓後35時間目以降、「密度分布の母代表値に差がない」
と判定された(2-3の統計解析手法を参照のこと)。実際、窒素源飢餓条件下で分裂してい なかった細胞の数が、窒素源飢餓後35時間目から減少していく(=細胞分裂が活発化する)
ことが見て取れた(図4)。ここで、窒素源飢餓後 30、35時間目では、窒素源飢餓条件下 での分裂が確認されなかった細胞が残されていたことに注目したい。これらの細胞が密度 分布に含まれると、密度分布全体の分散や対称性に影響を与えるため、窒素源飢餓後 40 時間目以降の密度分布との統計的有意差が、実際よりも大きく見積もられた可能性が残る。
そこで、窒素源飢餓条件下での分裂が確認されなかった細胞を除外し、改めて各経過時間 の密度を計算したうえで、総当たりで2標本間コルモゴロフ・スミルノフ検定(両側)を 行った(表2)。その結果、前述と同様に、細胞分裂の経過時間の密度分布は、窒素源飢餓 後 35 時間目以降、「密度分布の母代表値に差がない」と判定された(2-3 の統計解析手法 を参照のこと)。以上のことは、細胞分裂が窒素源飢餓により一時的に撹乱されるものの、
その影響は窒素源飢餓後35時間目までに消失することを示唆している。なお、細胞分裂が 撹乱された理由には、タイムラプス観察のために培養環境を変化させたことなども考えら れるが、本研究ではこの影響を考慮しなかった。
2-4-1-2) 窒素源飢餓による撹乱が消失して以降、分裂周期がほぼ一定値に収束した そこで、この撹乱が細胞分裂に及ぼす影響を解析し、それが消失した後での分裂ダイナ ミクスを記述するため、一度分裂してから次に分裂するまでの時間(「分裂周期」)を計測 した(図3D)。この解析では、各細胞が窒素源飢餓条件下で分裂した回数を「世代」とし てカウントした。即ち、窒素源飢餓条件に移した直後の全細胞を第0世代とし、細胞分裂 を経るごとに姉妹細胞の世代が1つずつ増える。窒素源飢餓後72時間目の時点で、フィラ メントには、窒素源飢餓条件下で4回分裂を行った237個の細胞(第4世代)と5回分裂 を行った210個の細胞(第5世代)が含まれる(その後に分化したヘテロシストを含む)。
26
35 h 40 h 45 h 50 h 55 h 60 h 65 h 70 h
30 h 9.26×10-3 0.0622 8.64×10-3 4.16×10-3 5.26×10-3 9.56×10-3 8.12×10-4 7.56×10-3
35 h 0.0101 0.354 0.0112 0.118 0.0354 0.0119 0.155
40 h 0.332 0.494 0.128 0.329 0.568 0.0690
45 h 0.0432 0.2026 0.124 0.0119 0.750
50 h 0.205 0.559 0.524 0.0806
55 h 0.735 0.0429 0.133
60 h 0.109 0.045
65 h 0.0113
表1 決められた時間で計測した「直前の細胞分裂からの経過時間」の密度分布(図 4A)を 2 標本間コルモゴロフ・スミルノフ検定し、求められたp 値の一覧表。p 値は、2-3 の統計解析 手法に記載した方法で求めた。
27
35 h 40 h 45 h 50 h 55 h 60 h 65 h 70 h
30 h 3.26×10-4 0.0562 4.62×10-3 0.0214 0.0139 0.0312 0.0206 6.60×10-4
35 h 0.0181 0.3855 0.0296 0.197 0.0675 0.0166 0.569
表 2 窒素源飢餓後 30、35 時間目で計測した「直前の細胞分裂からの経過時間」の密度分布か ら、分裂していなかった細胞を除外し、改めて密度分布を計算したうえで、ほかの密度分布 と総当たりで 2 標本間コルモゴロフ・スミルノフ検定(両側)を行った(表 2)。その結果、
求められた p値の一覧表。p値は、2-3 の統計解析手法に記載した方法で求めた。
28
全ての細胞は、窒素源飢餓後 35 時間目以降に2 回目の分裂を行ったため、窒素源飢餓 直後の細胞分裂の撹乱(一時停止)は第1世代の終わりまでに消失することが予想された。
そこで、第1~4世代での密度分布に対し、総当たりで2標本間コルモゴロフ・スミルノフ 検定(両側)を行った(表3)。このとき、第0世代の細胞の分裂周期は、窒素飢餓条件下 での連続する細胞分裂間の時間を表していないため、解析の対象から外した。その結果、
第2世代以降、「密度分布の母代表値に差がない」と判定された(2-3の統計解析手法を参 照のこと)。実際、第2世代以降の細胞では、すべての細胞の分裂が20時間以内に行われ、
各世代の90%以上の細胞が13時間以内に分裂したことがわかった。そこで、第2~4世代
で分裂した細胞の各分裂周期を足し合わせ、それを(第2~4世代の)全細胞数で標準化す ることで、窒素源飢餓条件下での細胞周期の密度分布とした(図5B)。
2-4-1-3) ヘテロシスト分化パターン形成のダイナミクスは、窒素源飢餓状態後63-65時
間目と69時間目以降で変化した
次に、ヘテロシスト分化した細胞が、窒素源飢餓後の時間経過に伴い、どのように出現 するかを解析した。ヘテロシスト分化が起きたタイミングは、細胞が肥大化し、フィコビ リソーム蛍光が低下し始めたときとした(8)。最初のヘテロシストが窒素源飢餓後63時間 目に分化したことを鑑みて、窒素飢餓による細胞分裂の撹乱が消失した後、ヘテロシスト 分化パターン形成が始まると考えられた。前述のように、初期の分化パターンでは、hetR 遺伝子を分化細胞と同程度に発現させたものの、最終的に分化しなかった細胞が多く観察 された。特に、Asai(30)は、窒素源飢餓後61~65時間目に、hetR遺伝子の発現が高く なることを報告した。つまり、ヘテロシスト分化が始まるとき、より多くの細胞が分化候 補細胞となるが、分化パターン形成が進むにつれて、限られた細胞のみが分化候補細胞と なることが示された。私は、hetR遺伝子の発現を個々の細胞で活性させるメカニズムが、
分化パターン形成の進行に伴って変化すると考えた。そこで、最初のヘテロシスト分化が 観察されてから、hetR遺伝子の発現活性が維持される窒素源飢餓後63~65 時間目を初期 ステージとし、ヘテロシスト間の細胞数がほぼ一定値に収束すると判断された窒素源飢餓
29
図 5 第 0-4 世代での分裂周期の密度分布。(A)第 0、1、2、3、4 世代で、それぞれ 24、48、
91、171、105 細胞を計測対象とした。これらは、各世代でまだ分化していない生細胞のみを含 む。(B)窒素源飢餓による撹乱が消失した後の分裂周期の密度分布。第 2-4 世代での各分裂周 期の長さの細胞数をすべて足し合わせ、それを(第 2-4 世代の)全細胞数で割ることにより、
改めて密度分布を求めた。
30
第 2 世代 第 3 世代 第 4 世代
第 1 世代 2.12×10
-31.15×10
-45.00×10
-4第 2 世代 0.999 0.999
第 3 世代 0.999
表 3 「第 1-4 世代での分裂周期の密度分布」(図 5A)を 2 標本間コルモゴロフ・スミルノフ 検定し、求められたp値の一覧表。p値は、2-3 の統計解析手法に記載した方法で求めた。
31 後69時間目以降を後期ステージと呼ぶことにした。
これ以降、初期ステージと後期ステージにおいて、どのように各ステージでの分化パタ ーン形成が変化するか解析した。そこで、ヘテロシストに挟まれた栄養細胞のクラスター を「ヘテロシスト間細胞群」、その栄養細胞数を「ヘテロシスト間細胞数」と名付け、ヘテ ロシスト分化パターンの指数と見なした。
初期ステージと後期ステージでの分化パターンを定量的に評価するため、先行研究と同 様のPhetR::gfp株を寒天培地で静置培養し、窒素飢餓後65、90時間目に複数フィラメント からヘテロシスト間細胞数を計測した(図6)。窒素源飢餓後 65時間目の密度分布では、
ヘテロシスト間細胞数の平均と標準偏差が 9.14、2.95(個)となった。一方、窒素源飢餓 後90時間目の密度分布では、平均と標準偏差が11.39、3.86となった。2標本間コルモゴ ロフ・スミルノフ検定(両側)を行った結果、「密度分布の母代表値に差がある」と判定さ
れた(p=2.10×10-4、2-3 の統計解析手法を参照のこと)。特に、窒素源飢餓後 65 時間目の
密度分布では8細胞の頻度が最も高く、窒素源飢餓後90時間目の密度分布では10細胞の 頻度が最も高くなった。さらに、後者の分布ではヘテロシスト間細胞数がより広い範囲で 計測された。
ここで、細胞系譜に記録されたフィラメントを解析し、窒素源飢餓後69-72 時間目での ヘテロシスト間細胞数の平均値を、窒素源飢餓後90時間目の(密度分布の)平均値とWelch のt検定した。その結果、「密度分布の母平均値に差がない」と判定された(いずれもp ≥ 0.05、
2-3 の統計解析手法を参照のこと)。一方、窒素源飢餓後 66~68 時間目でのヘテロシスト 間細胞数の平均値を、窒素源飢餓後90時間目の平均値とWelchのt検定した結果、「密度 分布の母平均値に差がある」と判定された(いずれもp < 0.05、2-3の統計解析手法を参照 のこと)。したがって、私たちの培養条件では、窒素源飢餓後69時間目に平均ヘテロシス ト間細胞数がほぼ収束したと考えられた。
以上のように、初期ステージと後期ステージで観察されるヘテロシスト分化パターンは、
それぞれ定量的に異なることが明らかにされた。したがって、運命決定ダイナミクスの変 化が、異なる分化パターン形成に反映されることを時間的に対応付けることができた。
32
図 6 初期ステージ と後期ステージでのヘテロシスト間細胞数の密度分布。(A)窒素源飢餓後 65 時間目の密度分布は、平均と標準偏差が 9.14、2.95 となった(計測対象のヘテロシスト細 胞群は 166 個)。(B)窒素源飢餓後 90 時間目の密度分布は、平均と標準偏差が 11.39、3.86 と なった(計測対象のヘテロシスト細胞群は 164 個)。いずれの時間でも、ヘテロシストが隣接 した細胞群や、細胞数が 30 を超えた細胞群は存在しなかった。
33
そこで、本研究では、初期ステージと後期ステージで観察される分化パターンを、それ ぞれ初期パターン、後期パターンと名付けて区別した。2-4-2-3で述べるパラメータ値の推 定では、窒素源飢餓後65時間目の分布を初期パターン、窒素源飢餓後90時間目の分布を 後期パターンとして参照した。
2-4-2) ヘテロシスト分化パターン形成の理論モデルの作成
2-4-2-1)セルオートマトンモデルの作成
私は、Anabaena のヘテロシスト分化パターン形成を模倣する数理モデルを作成した。
このモデルでは、直鎖状に連なった細胞をフィラメントとして考える。各細胞は、栄養細 胞かヘテロシストのいずれか2状態をとる。主なダイナミクスは、栄養細胞の運命決定が 確率的に行われることと、フィラメント内の細胞間で相互作用が行われることである。
このモデルは連続時間のダイナミクスだが、細胞の状態を離散として考える。栄養細胞 は、1)細胞分裂、2)ヘテロシスト分化、3)成熟(細胞成熟度指数の更新)を確率的に行
う(図7A)。ここで、細胞成熟度指数は分裂後の成熟レベルを表す自然数であり、細胞分
裂と分化はこの量に依存した頻度で起こると仮定する。細胞成熟度指数は細胞分裂直後に 1となり、その細胞が分裂もしくは分化するまで単位時間あたり一定率で 1ずつ増え、最 大値10に達するとそれ以上増えない。細胞分裂によって、母細胞は隣り合った2つの姉妹 栄養細胞に分かれ、それぞれ独立して細胞分裂、ヘテロシスト分化、成熟を行う。一方、
一度ヘテロシスト分化した細胞は、栄養細胞に脱分化することはなく、分裂や成熟も起こ らない。
さらに、フィラメント内での分化細胞の位置は、ヘテロシスト間細胞群を挟む両側のヘ テロシストからの距離(細胞数)にも依存する(図7A)。このような距離依存性の効果を 表すため、両ヘテロシストから離れた細胞ほど、分化抑制因子の影響が弱くなり、結果 的に分化しやすくなるとした。つまり、このモデルでは、PatSや窒素化合物などの分化 抑制因子の拡散性や濃度分布を直接的に表したわけではない。ここで、ヘテロシストの 位置決定に、最も近い左右のヘテロシスト以外からの影響はないとする。
34
図 7(A)細胞分裂、分化、成熟から構成される状態遷移と、ヘテロシストからの(分化の)距 離依存性の効果を示した図。一般に、ヘテロシストで発現する分化抑制因子の濃度は、ヘテロ シストに近い細胞ほど高くなる。そこで、このモデルでは、ヘテロシストから離れている細胞 ほど、分化しやすくなるとした。(B)数理モデルを数値シミュレーションしたときの各細胞の 状態の概略図。各細胞の細胞成熟度指数の値はランダムであり、それは一定率で増加する。ま た、細胞成熟度指数の値が高くなった細胞ほど、分裂もしくは分化しやすくなる。
35
新たにヘテロシスト分化する細胞は、2 つの既存のヘテロシスト間で最終的に一細胞に 決定されるが、それまでにヘテロシスト細胞群中間領域の複数の細胞で hetR 遺伝子の発 現が上昇する。この発現の活性化は、ヘテロシストからの距離に比例して起こるわけでは なく、一定の距離を離れると急激に起こるようになる。さらに、Rivers ら(38)は、距離 依存性の効果がパラクリン的に効いており、分化抑制因子の濃度勾配がヘテロシストから の距離に比例せず、一定の距離を離れると急激に希釈されることを示した。前述のとおり、
HetRタンパク質は、自己ポジティブフィードバックによってhetR遺伝子の発現をさらに 活性化させるため、一過的にhetR遺伝子の発現が高くなった細胞は分化しやすくなる(19)。 したがって、ヘテロシストから離れた領域の栄養細胞では、HetR タンパク質の数が分化 抑制因子の数に較べ、急激に多くなると考えられた(図1D)。したがって、距離依存性の 効果は、ヘテロシストからの距離に依存した指数関数でなく、シグモイド関数で表される べきだろう(図7A)。
以上のことを踏まえ、分化パターン形成を表すモデル1を以下のように定義する。
分裂率:
p(i) = a ∙ i
m1i
m1+ k
1m1分化率:
q(i) = b ∙ i
m2i
m2+ k
2m2(式 8)
分化率に対する距離依存性の効果:
l(n) = d ∙
n f n f +e fヘテロシストからの距離依存的な分化率:
Q(i, n
l, n
r) = q(i) ∙ l(n
l) ∙ l(n
r)
成熟率:r(i) = c
このモデルには、細胞分裂率