授乳期の母体の乳腺および離乳期の仔の腸管におけ
る免疫・微生物環境の形成機序解明
著者
新實 香奈枝
学位授与機関
Tohoku University
学位授与番号
11301甲第19322号
URL
http://hdl.handle.net/10097/00127867
授乳期の母体の乳腺および離乳期の仔の腸管
における免疫・微生物環境の形成機序解明
1
目次………
第一章 序論……… 1.1 幼若期の感染症と予防・治療……… 1.2 腸管の免疫および微生物環境……… 1.3 本研究の目的……… 第二章 授乳期の母体の乳腺組織における免疫および微生物環境の形成機序解明 ……… 2.1 はじめに……… 2.2 材料および方法……… 2.2.1 供試動物……… 2.2.2 採材……… 2.2.3 HE 染色……… 2.2.4 免疫組織化学染色……… 2.2.5 mRNA 抽出……… 2.2.6 cDNA 合成……… 2.2.7 リアルタイム PCR 反応……… 2.2.8 乳汁からのゲノム DNA 抽出……… 2.2.9 乳腺組織からのゲノム DNA 抽出……… 2.2.10 組織内細菌数測定……… 2.2.11 16S rRNA メタゲノム解析……… 2.2.12 統計解析……… 2.3 結果……… 2.3.1 生殖サイクルに伴うマウス乳腺組織の形態的変化と IgA 産生細胞の分布 ……… 1 5 10 13 11 13 6 6 8 13 13 14 15 16 17 18 18 19 19 20 21 212 2.3.2 生殖サイクルに伴う乳タンパク質および IgA 産生関連分子の遺伝子発現 ……… 2.3.3 カゼイン遺伝子と CCL28 および PigR 遺伝子発現の相関……… 2.3.4 ヒト由来乳汁とウシ由来乳汁中の微生物環境……… 2.3.5 マウス乳腺組織内の微生物環境……… 2.3.6 CCL28 および PigR 遺伝子発現に対する乳腺組織内微生物と授乳刺激の影響 ……… 2.4 考察……… 2.5 小括………
図表………
第三章 離乳期の仔の腸管における免疫および微生物環境の形成機序解明……… 3.1 はじめに……… 3.2 材料および方法……… 3.2.1 供試動物……… 3.2.2 採材……… 3.2.3 パイエル板からの細胞抽出……… 3.2.4 小腸・大腸からの細胞抽出……… 3.2.5 小腸内容物および糞便からのゲノム DNA 抽出……… 3.2.6 パイエル板細胞からのゲノム DNA 抽出……… 3.2.7 16S rRNA メタゲノム解析……… 3.2.8 フローサイトメトリー解析……… 3.2.9 蛍光免疫化学染色……… 3.2.10 ELISA(サンドイッチ法)……… 3.2.11 統計解析……… 21 22 22 23 23 25 29 30 39 40 42 42 42 43 43 44 44 45 45 47 48 493 3.3 結果……… 3.3.1 仔パイエル板における微生物の取込み……… 3.3.2 仔パイエル板における機能成熟……… 3.3.3 離乳後の IgA 産生に対する哺乳期の微生物環境の作用評価……… 3.4 考察……… 3.5 小括………
図表………
第四章 離乳期の仔の腸内免疫および微生物環境形成に関わる微生物の探索 ……… 4.1 はじめに……… 4.2 材料および方法……… 4.2.1 供試動物……… 4.2.2 採材……… 4.2.3 ELISA(サンドイッチ法)……… 4.2.4 糞便からのゲノム DNA 抽出……… 4.2.5 IgA 陽性細菌の単離……… 4.2.6 IgA 陽性細菌のフローサイトメトリー解析……… 4.2.7 IgA 陽性細菌からのゲノム DNA 抽出……… 4.2.8 16S rRNA メタゲノム解析……… 4.2.9 腸管組織のフローサイトメトリー解析……… 4.2.10 蛍光免疫化学染色……… 4.2.11 統計解析……… 4.3 結果……… 4.3.1. パイエル板欠損時における幼若期の IgA 産生……… 50 50 51 53 54 58 59 67 68 70 70 70 71 71 71 72 72 73 73 73 74 75 754 4.3.2 幼若期に誘導されるパイエル板依存的 IgA の微生物反応性評価……… 4.3.3 幼若期から成体にかけてパイエル板依存的に微生物環境に作用する菌種選抜 ……… 4.3.4 選抜菌種の特徴解析……… 4.4 考察……… 4.5 小括………
図表………
第五章 総括……… 謝辞……… 参考文献……… 76 76 78 79 82 83 92 98 1005
第一章
6 1.1 幼若期の感染症と予防・治療 哺乳期と離乳期を含む幼若期は、離乳や摂取時の食物構成、そして環境などの変化 による影響を受けて、体内の微生物および免疫環境が劇的に変化する時期である。この 時期の免疫機能は未熟であり、さらに微生物環境もまた不安定であることから、幼若期 は、ウイルスや微生物を含む病原体によって引き起こされる感染症に対する罹患リスク が非常に高い時期である。畜産の生産現場では、幼若期に感染症を罹患することによる 家畜の発育不全が経営成績に大きなダメージを与えることから、幼若期における慎重な 飼養管理が重要視されている(亀上知世子 2014)。幼若期の家畜の感染症予防と治療に は、主にワクチン接種と抗生剤投与が使用される(永井英貴 2011; 大塚浩通 2014)。ワ クチン接種は、感染症の罹患率の低減や罹患後の症状の緩和に効果が認められる一方で、 感染症の罹患を完全に抑制するほどの効力を有していないこと、そして、ワクチン自体 が未だ開発されていない感染症も数多く存在しているというのが事実である。また、抗 生剤は、畜産の生産現場において、病原性微生物により生じる感染症に対する治療に利 用されているほか、成長促進を目的とした飼料添加剤としても常用されている。細菌性 感染症の治療および成長促進剤としての抗生剤の効果は広く認められてきた一方で、そ の多用は、薬剤耐性菌の出現を助長する危険性を有していることが世界レベルで問題視 されており、現在の畜産現場では、ワクチンや抗生剤のみに頼らない幼若期の家畜自身 の免疫強化を誘導する飼養管理方法が必要とされている。 1.2 腸管の免疫および微生物環境 腸管免疫系は、IgA 産生を主とする液性免疫系が発達する。IgA は、腸管粘膜面に 侵入した病原性微生物に結合することで、病原性微生物が上皮へ接着するのを抑制する ほか、同じく粘膜面に常在する微生物に結合することで微生物環境を中心とする腸内の ホメオスタシスの維持に寄与することが知られている(Belkaid and Harrison 2017)。幼
7 若期の個体、特に哺乳期の仔は自身で IgA を産生することが出来ず、IgA による腸内の 免疫防御の殆どを母体から乳汁を介して供給される移行 IgA 抗体に依存する。仔自身 の IgA 産生は、離乳後に母体からの移行 IgA 抗体が仔の腸内で減少することに伴い、 発達し始める。そのため、母体の免疫機能が低下した場合、哺乳期の仔の腸管に直接的 な負の影響が加わり、哺乳期の感染症罹患率の上昇をもたらす(Blanchard-Rohner and Eberhardt 2017; Blanchard-Rohner and Siegrist 2011; Gopalakrishna et al. 2019)。また、 離乳後は仔自身の IgA 産生が開始されるが、成熟までには時間がかかることから、離乳 期もまた感染症に対する罹患リスクが非常に高い。故に、幼若期の感染症を予防するた めには、哺乳期の場合、移行 IgA 抗体を産生する母体の免疫強化を、離乳期には、仔自 身の IgA 産生を迅速に誘導することが必要とされる。特に家畜は、胎盤を介した母体か らの抗体移行が行われないことから、出生直後から離乳にかけての免疫機能の殆どを哺 乳により供給される乳汁移行型の抗体に依存している。そのため、幼若期の家畜の感染 症予防には、母体へ免疫し、その初乳を哺乳させることにより免疫を付与する乳汁免疫 型ワクチンが広く使用されており、哺乳期の母体の免疫機能状態がワクチン接種の効果 に大きな影響を与えている。 一方で、粘膜面の感染防御には、IgA を主体とする液性免疫の強化だけでなく、粘 膜面に常在する微生物が形成する細菌叢のバランス維持も重要であることが知られて いる。腸管の粘膜面には、哺乳期から多種多様な微生物が定着し、菌叢が形成されてい る。腸管粘膜面の菌叢構成は、離乳と成育に伴って哺乳期と離乳期の幼若型から成体の 成熟型へと変化していくことが報告されており、時期ごとに適した微生物環境形成を誘 導することが、病原体の定着の抑制に寄与する(Janzon et al. 2019; Clarke et al. 2010; Zeng et al. 2016; Naik et al. 2012; Diehl et al. 2013)。また、腸内に存在する IgA と微生 物叢は互いに作用し合うことが知られている。具体的には、粘膜面の微生物は、IgA の 誘 導 組織 であ る腸 管関連 リ ンパ 組織 に取 り込ま れ るこ とで 抗原 として 機 能す る
8
(Macpherson and Uhr 2004)。さらに、誘導された IgA は管腔内で微生物に結合するこ と で 微 生 物 環 境 の バ ラ ン ス 維 持 に 寄 与 す る こ と が 先 行 研 究 で 報 告 さ れ て き た (Kawamoto et al. 2014; Sutherland, Suzuki, and Fagarasan 2016)。故に、正常な IgA 産 生を誘導するには良好な微生物環境を構築することが必須とされる。 したがって、幼若期の感染症を予防し、仔の健全な成育を促すためには、哺乳期の 母体の免疫強化と離乳後の仔の腸管での迅速な免疫誘導に加えて、仔の腸内における良 好な微生物環境の形成が重要とされる。そして、これらを実現するためには、移行抗体 を誘導する母体における抗体産生機構と仔の腸内の微生物および免疫環境の形成機序 に関する知識を基に、授乳期の母体と幼若期の仔に適した飼養環境を構築する必要があ る。しかしながら、母子の免疫・微生物環境における発達機序の多くは明らかとされて おらず、更なる研究が必要とされている。 1.3 本研究の目的 本研究では、ワクチンや抗生剤のみに頼らない感染症の予防可能な幼若期の飼養管 理方法を考案することを目的として、仔自身が腸内の微生物および免疫環境を獲得する 過程を明らかにすべく、幼若期を、免疫誘導の主軸が母体にある哺乳期と仔自身にある 離乳期の 2 つの期間に区別し、母体と仔の双方から免疫学的かつ微生物学的アプローチ を試みた。具体的に第 2 章では、移行抗体を媒介する乳汁の産生場所である母体の乳腺 組織に着目し、仔へ移行する IgA 抗体の産生過程における分子メカニズムを明らかに することを目的として、マウス乳腺組織における IgA 産生関連分子の発現解析を行う と共に発現の誘導因子を検討した。そして第 3 章では、仔の腸内環境に着目し、仔自身 の腸内微生物環境および免疫環境の発達機序を明らかにすべく、哺乳期に腸内微生物環 境が形成される際の母体の役割と、離乳後の IgA 産生の発達における哺乳期の微生物 環境の作用を調査した。最後に第 4 章では、離乳後の仔の腸管免疫機能を賦活化させる
9 微生物の探索を行った。
10
第二章
授乳期の母体の乳腺組織における免疫
および微生物環境の形成機序解明
11 2.1 はじめに 母体の乳腺組織は、外分泌組織の中でも妊娠、分娩、授乳、離乳から成る生殖サイ クルを通して、組織の形態が劇的に変化する特徴的な組織である。乳腺組織の主な役割 は、仔へ供給する乳汁の合成と移行 IgA 抗体を産生する形質細胞の遊走および分泌さ れた IgA の乳腺房腔への輸送であり、哺乳期の仔が正常に発育するためには、乳腺機能 の正しい理解とその機能の向上は欠かせない。 乳汁を媒介にして仔へ供給される移行 IgA 抗体は、乳腺上皮細胞からケモカイン の一種である CCL28 の分泌によって誘導される。具体的には、CCL28 が上皮細胞から 分泌されることで、その受容体である CCR10 を発現する IgA 形質細胞が粘膜関連リン パ組織から乳腺へ遊走し、間質部位で IgA を分泌する。この際、間質で分泌された IgA は、乳腺上皮の基底側に発現する Polymeric immunoglobulin receptor(以下、PigR)に結 合することで上皮細胞内を通り、乳汁中へと放出されるようになる(Wilson and Butcher 2004; Brandtzaeg 2010)。故に、CCL28 および PigR は乳腺の IgA 産生の中心的な役割 を担っている。よって、乳腺におけるこれらの発現機序を理解し、正常に誘導すること は、乳腺組織の IgA 産生機能の向上、延いては、IgA を享受する哺乳期の仔の健全育成 に寄与すると考えられる。
CCL28 と PigR は乳腺組織だけでなく、腸管組織にも発現することが知られてい る(Brandtzaeg 2010)。特に大腸は、CCL28 が盛んに分泌されている組織であり、CCL28 および PigR の発現制御機構に関する研究が多く報告されている(Ogawa et al. 2004; Kvale and Brandtzaeg 1995; Kaetzel 2014)。大腸の CCL28 は、管腔内の微生物の代謝
産物である短鎖脂肪酸や
bacterial flagellin によって発現が誘導される(Ogawa et al.
2004)。また、PigR の場合も、微生物の代謝産物の一種である酪酸によって発現
が恒常的に維持される(Kvale and Brandtzaeg 1995)。そのため、腸管で CCL28
および PigR の発現を誘導し、IgA 産生を正常に機能させるためには腸管粘膜面
12
に常在する微生物叢を良好な状態に維持することが重要である。一方、乳腺では、
大腸と同じく CCL28 と PigR による IgA 産生が機能する一方で、これら分子の
発現誘導機構の殆どは、明らかとされていない。
近年、次世代シーケンサーを用いた 16S rRNA メタゲノム解析の進展により微生物 の検出能力が格段に飛躍してきた。具体的には、培養法による検出が不可能であった菌 が DNA レベルで検出可能となり、特に、無菌環境であると思われていた乳汁の中にも 多種多様な微生物種が存在することがウシやヒトの臨床研究で明らかとされてきた (Taponen et al. 2019; McGuire and McGuire 2017)。しかしながら、ウシやヒトの乳汁 中で検出された微生物が乳汁中だけでなく、乳汁を産生する乳腺組織内において微生物 環境を形成しているのか、そして、乳腺組織内にも腸管のように微生物が存在していた 場合、組織内の微生物は乳腺の IgA 産生を誘導する因子として CCL28 や PigR の発現 を誘導するのか、その詳細は不明である。 そこで、本章では、母体の乳腺組織内の IgA 産生の制御機構を明らかにすべく、乳 腺組織における微生物の存在の有無を調査したほか、乳腺内の免疫および微生物環境の 関係性を評価した。13 2.2 材料および方法 2.2.1 供試動物 SPF 環境下で飼育した BALB/c マウス(熊谷商店)を交配し、分娩 1 週間前(妊娠期)、 分娩直後、分娩 1 週間後、分娩 2 週間後(授乳期)、分娩 6 週間後(離乳後)の各タイミン グで乳腺組織を採材した。また、無菌環境下で飼育し、体内に微生物を保有しない無菌 マウス(三共ラボ)を交配し、分娩 2 週間後の乳腺を採材した。さらに、SPF 環境下で飼 育した BALB/c マウスの一部は、分娩 1 週間後に仔マウスを母体から離すことで強制 的に離乳し、その後、分娩 2 週間後に通常飼育した個体と同じく乳腺組織を採材した。 ウシの乳汁は、静岡県畜産試験場で飼育された 2 頭のウシから 6 月と 9 月に分房 別に乳汁を回収した。また、ヒトの乳汁は、東北大学病院へ通院する被験者から分娩後 1 か月目の健診日に乳汁を回収した。 2.2.2 採材 マウスから採材した乳腺組織は 4%パラフォルムアルデヒドに一晩、固定後にパラ フィン包埋を行った。その後、ミクロトームを用いてパラフィンブロックを 5µm の厚 さに薄切することで組織切片を作製し、組織学的解析に供した。また、乳腺組織の一部 は mRNA を抽出後、cDNA 合成することでリアルタイム PCR に供した。さらに、マウ スから採材した乳腺組織の一部、そしてウシとヒトから回収した乳汁からゲノム DNA を抽出し、細菌特有の遺伝子配列である 16S 配列(V3-4 領域)を PCR で増幅後、次世代 シーケンサーを用いて、マウスの乳腺組織内およびウシとヒトの乳汁中の細菌叢のメタ ゲノム解析を行った。 2.2.3 HE 染色 パラフィン組織切片から下記の工程※1でパラフィンを取り除いた後、3 分間、流水
14 で洗浄し、ヘマトキシリン溶液に 3 分間反応させた。その後、流水で 15 分以上洗浄し、 エオジンに 15 分間反応させた。エオジン反応後、95%エタノールで一回、100%エタノ ールで二回、分別した。100%エタノールで 3 分間脱水した後、キシレンで 3 分間透徹 を 3 回行い、マリノール(武藤化学)を用いて封入した。染色した組織は、光学顕微鏡 (OLYMPUS)を用いて観察および撮影した。 ※1 脱パラフィン ①キシレン1 3 分液浸 ②キシレン2 3 分液浸 ③キシレン3 3 分液浸 ④100%アルコール1 3 分液浸 ⑤100%アルコール2 3 分液浸 ⑥95%アルコール 3 分液浸 ⑦80%アルコール 3 分液浸 2.2.4 免疫組織化学染色 パラフィン組織切片から上述した方法でパラフィンを取り除いた後、5 分間、流水
で洗浄した。その後、切片の周りを SUPPER PAP PEN(Daido Sangyo)で囲い、3% H2O2
/メタノール溶液を 100µL 添加し、室温で 15 分間反応させ、内在性ペルオキシダーゼ を阻害した。その後、切片を Wash buffer(500mM Tris-HCl(p.H.7.4), 1.4M NaCl, 0.5% Tween20)を用いて、3 分間洗浄した。洗浄を 2 回行った後、Blocking buffer (100mM Tris-HCl(pH7.4), 150mM NaCl, 0.5% Blocking reagent: perkinelmer)を各組織上に 100µL 添加し、30 分間反応させた。反応後、液を除き、Blocking buffer で 1µg/mL に 希釈したラット抗マウス IgA 抗体(BD Pharmingen, clone No; C10-3)を乗せ、一晩反応
15
させた。反応後、Wash buffer によって 5 分間洗浄した。洗浄を 4 回繰り返した後、シ ンプルステインマウス MAX-PO【RAT】(ニチレイバイオサイエンス)を組織に添加し、 室温で 1 時間反応させた。その後、Wash buffer を用いて、洗浄を 4 回行い、DAB 発色 用溶液を組織切片に添加し、室温で 4 分間発色した。4 分経過後すぐに、イオン交換水 へ切片をつけることで発色を止め、流水洗浄を 5 分間行った。その後、ヘマトキシリン 溶液に 30 秒間反応させ、直ぐに流水洗浄を 15 分間行った。洗浄後に 80%、90%、95%、 100%(3 回)の順で 3 分間ずつ組織切片を移すことで脱水した。さらに、キシレンに 3 回移すことによって透徹し、最後にマリノールで封入した。染色した組織は光学顕微鏡 (OLYMPUS)を用いて観察および撮影を行った。 2.2.5 mRNA 抽出
mRNA は、ReliaPrepTMRNA Tissue Miniprep System(Promega)を使用して抽出し
た。採材した組織片を、3.2mm3ステンレスビーズ(TOMY)を入れたサンプルチューブ
(TOMY)に移し、System 付属の LBA Buffer;500µL と 1-Thioglycerol;10µL を添加後、 4000rpm で 60 秒間、破砕機(WAKENYA)にかけることで組織片を破砕した。組織片を 破砕後、15000rpm で 3 分間遠心操作し、上清を 1.5mL チューブに全量移した。新しい チューブへ移した上清にイソプロパノールを 170µL 添加し、5 秒間ボルテックスを用 いて攪拌後、Miniprep System に付属のカラムチューブへライセートを全量移した。ラ イセートを添加したカラムチューブを 15000rpm で 1 分間遠心操作し、濾液を捨てた。 その後、カラムへ Miniprep System 付属の RNA wash solution を 500µL 添加し、 15000rpm で 1 分間遠心操作した。濾液を捨て、カラムへ Miniprep System 付属の
DNase incubation mix (Yellew core buffer;24µL/sample 、 MnCl2;3µL/sample 、
DNaseI;3µL/sample)を 30µL、カラムのメンブレン中央部に添加し、室温で 15 分間、 静置した。15 分後、Miniprep System 付属の Colum wash solution を 200µL 添加し、
16
15000rpm で 1 分間遠心操作した。500µL の RNA wash solution をカラムへ添加し、 15000rpm で 1 分間再度、遠心操作した。濾液を捨て、300µL の RNA wash solution を カラムへ再び添加した後、15000rpm で 2 分間遠心操作することでメンブレンを洗浄し た。濾液を捨て、空のカラムを 15000rpm で 2 分間遠心操作し、メンブレンを乾燥させ た。Miniprep System 付属の Nuclease free water を 30µL、メンブレンへ添加し、 15000rpm で 1 分間遠心操作することで、mRNA を溶出させた。抽出したmRNA は -80℃で保存した。
2.2.6 cDNA 合成
Prime ScriptTM RT Reagent kit(Takara)を使用して、cDNA 合成を行った。分光光
度計(SHIMADZU)を用いて抽出した Total RNA の濃度を測定後、濃度が 500ng/µL 以
下なるように mRNA を希釈した。希釈した mRNA と Prime ScriptTM RT Reagent kit に
付属の5×PrimeScript Buffer、PrimeScript RT Enzyme、Oligo dT Primer、RNase Free
dH2O を下記の使用量で混和し、サーマルサイクラー(ASTEC)を用いて逆転写反応※1
を行った。作成した cDNA は-20℃で保存した。
<1 反応あたりの使用量>
試薬 使用量 最終濃度
5×PrimeScript Buffer(for Real Time) 2μL
PrimeScript RT Enzyme Mix I 0.5μL
Oligo dT Primer(50μM) 0.5μL 25 pmol
Random 6 mers(100μM) 0.5μL 50 pmol
total RNA(<500ng) X μL
Rnase Free dH2O 10-(3.5μL+XμL)
17 ※1 逆転写反応条件
逆転写反応 37℃ 15 分 逆転写酵素の熱失活 85℃ 5 秒
2.2.7 リアルタイム PCR 反応
作出した cDNA を TB GreenTM Premix Ex TaqTM Ⅱ(Takara)を用いて増幅し、リ
アルタイムPCR解析システム Thermal Cycler Dice Real Time System Single (Takara )
を用いて解析を行い、csn1s1、csn2、csn3、ccl28、pigr、iga の発現量を定量した※1。
PCR プログラムは、2 ステップ PCR を採用し、下記の反応条件※2で 40 サイクル行っ
た。1 サイクル毎に蛍光を測定し、melting curve から発現量を検出した。また、standard curve よりプライマーの増幅効率を計算することで結果に反映した。それぞれの遺伝子 の発現量をgapdhで補正し評価した。 ※1 プライマー情報 ※2 PCR 反応条件(2 ステップ PCR を採用) 熱変性 95℃ 5 秒 アニーリングおよび伸長反応 60℃ 30 秒
Name Gene name Sequence F Sequence R 増幅サイズ
casein alpha Csn1s1 TCTTGGCAACCATGAAACTCCT TCACTGCTGCTATGCTGTTGCT 130bp casein beta Csn2 TTCCACAAGATATGTCTGTCCAAG TGTAGCATGATCCAAAGGTGAA 171bp casein kappa Csn3 AATCCTGAAGCTTCCACAGTGTCTA TCATGGATCTTTCCAGGAATGGTAT 161bp
CCL28 Scya28 AACTGGCTGCTATGGTTTGGGTAA CCAGGAGCACCACAGGATGA 84bp
Polymeric immunoglobulin receptor Pigr TGCCTGTGACAATCAACTTGAGAA GAACCAGCACACCTCTGAGGAC 101bp
IgA iga ATGACTCTAACGCCGTCCAAGAA ACCCAGGAGCAGGTCCTCAA 131bp
18 2.2.8 乳汁からのゲノム DNA 抽出
Milk Bacteria DNA Isolation Kit(NOLGEN)を使用して、乳汁からゲノム DNA を 抽出した。採材したウシおよびヒトの乳汁 200µL を 1.5mL チューブに移し、2 分間遠 心操作を行った。上清を取り除き、チューブ内に残存したペレットを破砕後、そこへ Lysozyme を添加した Resuspention Solution A(Kit 付属)を 100µL 添加し、ボルテック スを用いて混和した。混和後、37℃で 45 分間静置した。反応後、Kit に付属の Buffer SK を 300µL と Proteinase K を 10µL 添加し、ボルテックスを用いて、混和した。その 後、55℃で 45 分間静置した。反応後に 100%エタノールを 200µL 加え、ボルテックス を用いて混和後、ライセートを付属のカラムへ添加した。カラムを 15000rpm で 2 分間 遠心操作し、濾液を捨てた後、Buffer SK を 500µL 添加した。再度、15000rpm で 2 分 間遠心操作を行い、濾液を廃棄した後、付属の Wash solution を 500µL 添加し、2 回洗 浄した。その後、濾液を廃棄し、カラムを空のまま遠心操作することでメンブレンを乾 燥させた。最後に付属の Elusion buffer B を 100µL 添加し、2000rpm で 2 分間遠心操 作後、追加で 15000rpm 1 分間遠心操作することでゲノム DNA を溶出した。抽出した ゲノム DNA は-20℃で保存した。 2.2.9 乳腺組織からのゲノム DNA 抽出
Promega Wizard SV Genomic DNA Purification System(Promega)を使用して乳腺 組織からのゲノム DNA 抽出を行った。DNA Purification System 付属の Digestion Solution Master Mix(Nuclei Lysis Solusion;200µL/sample、0.5M EDTA;50µL/sample、 proteinase K;0.4mg/sample、RNase A Solution;0.02mg/sample)を 2mL 滅菌スクリュー
キャップマイクロチューブへ 275µL 分注し、そこへ、1mm3角の組織片となるまで細か
くした乳腺組織を加えた。その後、55℃で一晩静置することで、組織を溶解させた。反 応後、DNA Purification System 付属の Wizard SV Lysis Buffer を 250µL 添加し、ボル
19 テックスを用いて混和した。15000rpm で 3 分間遠心操作を行い、上清を付属のカラム に添加し、再度 15000rpm で 3 分間遠心操作した。濾液を捨て、付属の Colum wash solution を 650µL 添加し、15000rpm で 1 分間遠心操作することで、カラムを洗浄し た。さらに洗浄過程を 3 回行うことで不純物を洗い流した。洗浄後、濾液を捨て、カラ ムが空のまま 15000rpm で 2 分遠心操作することで、カラムを乾燥させた。Nuclease free water を 250µL 添加し、室温で 2 分間静置後、15000rpm で 1 分間遠心操作を行う ことで、ゲノム DNA を溶出した。抽出したゲノム DNA は-20℃で保存した。 2.2.10 組織内細菌数測定
Bacteria(Tuf gene) Quantitative PCR Kit(Takara)を用いて、乳腺組織から抽出した ゲノム DNA を鋳型に Tuf 遺伝子を増幅し、リアルタイム PCR 解析システム Thermal Cycler Dice Real Time System Single (Takara 社)により Tuf 遺伝子の発現量を定量し た。PCR プログラムは 2 ステップ PCR を採用し、熱変性を 95℃で 5 秒、アニーリン グと伸長反応を 60℃で 3 秒とし、40 サイクル行った。1 サイクル毎に蛍光を測定し、 melting curve から発現量を検出した。また、standard curve よりプライマーの増幅効 率を計算することで結果に反映した。
2.2.11 16S rRNA メタゲノム解析
Prime STAR® HS DNA polymerase(Takara)を用いて、乳腺および乳汁から抽出し たゲノム DNA を鋳型に 16S rRNA 遺伝子配列内 V3-4 領域を増幅した。PCR プログラ ムは 3 ステップ PCR を採用し、熱変性 98℃で 10 秒、アニーリングと伸長反応を 57℃
で 15 秒、72℃で 30 秒とし、40 サイクル行った。作成した 1stPCR 産物は、2%アガロ
ースゲルを用いた電気泳動にかけることで目的遺伝子の増幅を確認した。さらに、プラ イマーダイマーを取り除く為に、Agencourt AMPure XP(BECKMAN COULTER)を用
20
いて 1stPCR 産物の精製を行った。精製後、精製した 1stPCR 産物内の 16S rRNA 遺伝子
配 列 に サ ン プ ル を 識 別 す る タ グ 配 列 を 添 付 す る 為 に 、 Prime STAR® GXL DNA
polymerase(Takara)を用いて 2nd PCR を行った。この際、PCR プログラムは 98℃で 10
秒、54℃で 15 秒、68℃で 30 秒の 12 サイクルを実施した。その後、MultiNA(SHIMADZU)
を用いて、2nd PCR 産物(以下、ライブラリ)の増幅を確認した。さらに、PCR 反応物は
塩濃度が高いため、Agencourt AMPure XP を用いたバッファーの置き換えとライブラ リの精製を行った。ライブラリ精製後、MultiNA を用いて、精製後の増幅産物の状態を 確認した。精製後のライブラリは、Library Quantification kit(Takara)を用いて増幅し、 PCR efficiency calculator(KAPA)を使用して、反応物の濃度を測定した。PCR プログラ ムは、98℃で 10 秒、60℃で 15 秒、68℃で 45 秒の 35 サイクル行い、1 サイクルごと に蛍光を測定し、melting curve から発現量を検出した。濃度を算出したライブラリは、 Elusion Buffer(QIAGEN)を用いて 2nM に希釈後、0.2N NaOH で 1nM に希釈した。さ らに、Hyb Buffer(Ilumina)により 20pM にまで希釈した。最後に、20pM PhiX と Hyb Buffer を用いて 12pM へ調整し、これを最終濃度にして Miseq の次世代シーケンス解 析を実行した。 2.2.12 統計解析 実験結果は、Prism を用いて多重比較検定を行うことで統計学的に比較検討した。 有意水準は5%未満とした。また、次世代シーケンサーを用いて検出した微生物種は、 サンプル内の存在割合について R を用いた FDR 検定に供した。
21 2.3 結果 2.3.1 生殖サイクルに伴うマウス乳腺組織の形態的変化と IgA 産生細胞の分布 生殖サイクルを通した乳腺組織の形態および IgA 産生の特徴を明らかにすべく、 分娩 1 週間前(妊娠期)、分娩直後(授乳期)、分娩 1 週間後、分娩 2 週間後、分娩 6 週間 後(離乳後)、未経産のマウスから乳腺を採材し、HE 染色と抗 IgA 抗体を用いた免疫化 学染色による組織学的解析に供した。HE 染色を用いた組織学的解析の結果、妊娠期の 乳腺組織には、乳汁を貯蔵する腺房の前駆構造が確認された。この腺房の前駆構造は、 分娩時には腺房腔の発達が認められ、授乳日数の経過と共にさらに発達した。また、離 乳後になると発達した腺房構造は、未経産と同程度まで退行することが確認された(図 1A)。 乳腺内の IgA 産生細胞を免疫化学染色により染色した結果、妊娠期と分娩直後の 乳腺内には IgA 産生細胞が殆ど認められなかった。IgA 産生細胞は分娩 1 週間後から発 達した腺房周辺の間質部位に認められるようになり、授乳日数の経過と共に徐々に IgA 産生細胞の集積が増加した。また、離乳後には腺房構造の退行と同じく IgA 産生細胞も 認められなくなった(図 1B)。さらに、免疫化学染色画像から 0.05cm2当たりの細胞数 を測定した結果、分娩直後から分娩後 2 週間後にかけて IgA 産生細胞数が有意に増加 し、離乳後には細胞数が有意に減少することが確認された(図 1C)。このことから、乳 腺の IgA 産生は分娩後の授乳期に発達することが示唆された。 2.3.2 生殖サイクルに伴う乳タンパク質および IgA 産生関連分子の遺伝子発現 乳腺組織の形態的な変化に伴う乳合成を主とする生理機能と IgA 産生を主とする 免疫機能を評価すべく、乳タンパクの代表であるカゼインα、β、κの遺伝子発現と IgA 形質細胞の遊走因子である CCL28 および IgA の輸送因子である PigR、そして IgA の 遺伝子発現をリアルタイム PCR により測定した。その結果、カゼイン 3 種の遺伝子発
22
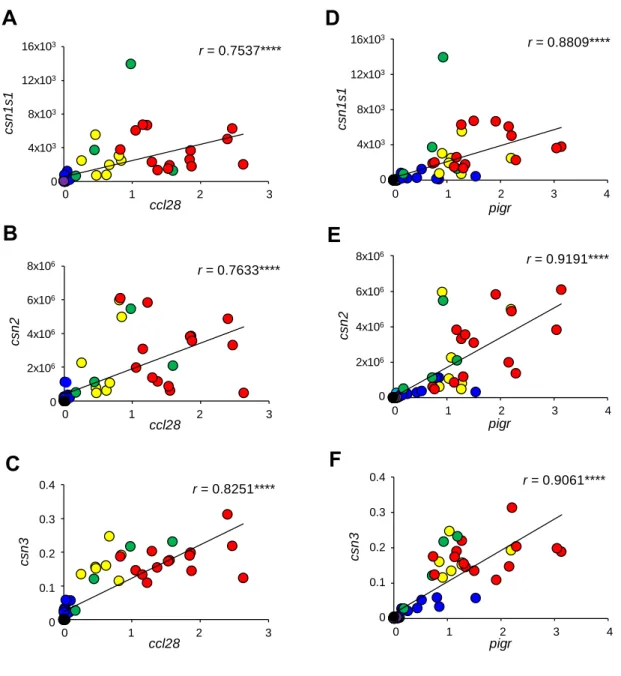
現は、腺房構造の発達と同じく、分娩後 2 週間で有意に増加し、離乳後には発現が減少 した(図 2A)。一方、IgA 産生関連分子である CCL28 と PigR 遺伝子の発現もまた、IgA 産生細胞の免疫化学染色の結果と矛盾なく、IgA 産生細胞が集積する分娩後の 2 週間に 発現が有意に増加し、離乳後には、それらの発現の減少が認められた。さらに、乳腺内 の IgA 遺伝子の発現もまた CCL28 と PigR 遺伝子の発現、そして IgA 産生細胞の集積 と同じく分娩後に増加し、離乳後に減少することが確認された(図 2B)。 2.3.3 カゼイン遺伝子と CCL28 および PigR 遺伝子発現の相関 カゼインα、β、κの遺伝子発現と CCL28 および PigR の遺伝子発現はどちらも 分娩後の授乳期に発現が増加していることから、乳腺の免疫機能には乳汁合成に関わる 授乳期特有の遺伝子発現制御が関与している可能性が考えられた。その知見の一つとし て、カゼイン遺伝子と CCL28 または PigR 遺伝子の発現の相関を解析した。その結果、 カゼインα、β、κ遺伝子と CCL28 遺伝子、そしてカゼインα、β、κ遺伝子と PigR 遺伝子の間にはそれぞれ正の相関が認められた(図 3)。 2.3.4 ヒト由来乳汁とウシ由来乳汁中の微生物環境 細菌叢に関する近年の研究で、乳汁中に微生物の存在が報告されたことから (McGuire and McGuire 2017; Taponen et al. 2019; Fernandez et al. 2013)、本研究では、 16Sr RNA メタゲノム解析を用いてヒトおよびウシ乳汁中の微生物を遺伝子レベルで調 査した。その結果、ヒト乳汁中には 359 菌種、ウシ乳汁中には 1788 菌種が検出された
(図 4,5)。具体的には、ヒト乳汁中ではLatobacillales目、Bacillales目、Bacteroidales目
等が優勢菌種として特定された菌種の中でも高い割合で存在することが確認された(図
4)。さらに、ウシ乳汁中ではClostridiales目、Bacteroidales目、Corynebacteriales目が
23 2.3.5 マウス乳腺組織内の微生物環境 乳腺組織内にも微生物が存在することを明らかにすべく、分娩 1 週間前(妊娠期)、 分娩直後(授乳期)、分娩 1 週間後、分娩 2 週間後、分娩 6 週間後(離乳後)、未経産のマ ウスの乳腺組織からゲノム DNA を抽出し、リアルタイム PCR によって Tuf 遺伝子を 定量することで乳腺組織内の菌数を定量したほか、16S rRNA メタゲノム解析を用いて 組織内に存在する微生物種を特定した。その結果、未経産および分娩 1 週間前には Tuf 遺伝子の発現が殆ど検出されなかったが、分娩直後から Tuf 遺伝子の発現が有意に増 加し、分娩後 2 週間に発現が維持された。さらに、離乳後には乳腺組織内の Tuf 遺伝子 発現量が未経産と同程度まで減少した(図 6A)。このことから、乳腺組織内では分娩後 に授乳期特異的に細菌数が増加することが示唆された。 さらに、菌数の増加が認められた授乳期の乳腺組織内に存在する微生物を種レベル で同定すべく、次世代シーケンサーによる 16S rRNA メタゲノム解析を用いて解析し た。その結果、分娩直後から分娩 2 週間後にかけて 780 種の微生物種が乳腺組織内で
同定された。特に、Bacteroidales目、Clostridiales目、Lactobacillales目を主とする微
生物が同定された微生物の中でも優勢菌種として乳腺組織内に存在することが確認さ れた(図 6B)。一方で、分娩直後と分娩 2 週間後に存在する乳腺組織内の微生物種の構 成を、R を用いた FDR 法によって統計学的に解析した結果、有意な差は認められなか った。以上の結果から、乳腺組織では分娩直後から発達した微生物環境が形成されてい ること、さらに、微生物環境は授乳期間を通して維持されていることが遺伝子レベルで 示唆された。 2.3.6 CCL28 および PigR 遺伝子発現に対する乳腺組織内微生物と授乳刺激の影響 大腸では、管腔内の微生物が腸管上皮細胞を刺激することで CCL28 と PigR 分子 の発現を誘導することが報告されている(Ogawa et al. 2004; Kvale and Brandtzaeg
24 1995; Kaetzel 2014)。一方で、乳腺組織では CCL28 と PigR 遺伝子発現が増加する授 乳期に、微生物が組織内に存在することが本研究を通して示唆された。そこで、乳腺で も大腸と同じく微生物の刺激が CCL28 と PigR の発現に関与するのか否かを調査した。 具体的には、無菌環境下で飼育し、体内に微生物を保有しない無菌マウスまたは SPF 環 境下で飼育した BALB/c マウスを用意し、それぞれ無菌環境または SPF 環境で妊娠、 分娩させた後、乳腺で IgA 産生が発達する分娩後 2 週間目に乳腺を採材し、mRNA を 抽出後にリアルタイム PCR を用いて CCL28 および PigR 遺伝子の発現を比較検討し た。その結果、無菌マウスと SPF 環境下で飼育したマウスの乳腺では、CCL28 と PigR 遺伝子の発現量に大きな差は認められなかった(図 7)。このことから、組織内の微生物 は、乳腺における CCL28 および PigR の発現を主とする免疫機能に深く影響しないこ とが示唆された。 また一方で、本研究を通して乳腺の CCL28 と PigR の遺伝子発現は、カゼイン遺 伝子と同じ発現傾向を有しており、乳汁合成に関わる授乳期特有の刺激によって CCL28 と PigR 遺伝子の発現が誘導される可能性が示唆された。そこで、授乳期特有の 刺激の一つとして仔による哺乳刺激に着目し、哺乳刺激の有無による CCL28 と PigR 遺伝子の発現の変化も調査した。具体的には、SPF 環境下で妊娠、分娩させた BALB/c マウスから、分娩後 1 週間目に仔を排除することで強制的に離乳を行い、無菌マウスお よび通常飼育した SPF マウスと同じく分娩 2 週間後に乳腺組織を採材した後、mRNA を抽出し、リアルタイム PCR により CCL28 と PigR 遺伝子の発現を評価した。その結 果、無菌マウスと通常飼育したマウスでは、CCL28 と PigR 遺伝子の発現に大きな差は 認められなかった一方で、通常飼育したマウスと強制離乳を行ったマウスの遺伝子発現 量を比較すると、強制離乳を行ったマウスの乳腺で CCL28 と PigR 遺伝子の発現が著 しく低値を示していた(図 7)。このことから、乳腺における CCL28 と PigR 遺伝子の発 現には、微生物による刺激よりも仔からの哺乳刺激が重要であることが示唆された。
25 2.4 考察 本章では、母体の乳腺における IgA 産生強化を目的とした、乳腺の IgA 産生の分 子制御メカニズムを解析した。 はじめに、生殖サイクルを通して乳腺組織内の IgA 産生を組織学的および遺伝子 学的な手法を用いて評価した。その結果、乳腺組織では、分娩後の授乳期間で乳汁合成 に伴う腺房構造の発達と共に CCL28 と PigR の遺伝子発現が増加し、腺房周辺に IgA 産生細胞の集積が認められた。その後、離乳後には腺房構造の退行と同じく、CCL28 お よび PigR、IgA 遺伝子発現の減少と IgA 産生細胞の減退が確認された。このことから、 乳腺の IgA 産生は、授乳期に限定した時期特異的な反応であること、そして、乳腺の IgA 産生において中心的な役割を担う CCL28 および PigR の発現は、授乳期間の何ら かの刺激によって発現が誘導される可能性が示唆された。 そこで次に、CCL28 と PigR の発現の誘導に関わる授乳期間の刺激を調査した。 IgA 産生に関する先行研究では、恒常的な IgA 分泌が行われている腸管において CCL28 と PigR の発現は、粘膜面に常在する微生物によって誘導されることが報告されている (Ogawa et al. 2004; Kvale and Brandtzaeg 1995; Kaetzel 2014)。そのため、乳腺でも微 生物刺激が授乳期の CCL28 と PigR の発現を誘導する可能性を考え、乳腺における微 生物環境の存在を評価した。乳腺に関わる微生物に関する先行研究では、乳汁中に多種
の微生物が存在することを報告しており、特に健康なウシでは、Lachnospiraceae spp.
等また、ヒトの場合は、Staphylococcus、Streptococcus、Pseudomonas spp.等の存在が
知られている(McGuire and McGuire 2017; Taponen et al. 2019)。本研究においてもウ シおよびヒトの乳汁中の微生物を 16S rRNA メタゲノム解析によって同定した結果、先 行研究で報告されている菌種を含む多種多様な微生物の存在が認められた。興味深いこ とに、本研究では乳腺に関する微生物の新たな知見として、乳汁だけでなく、乳汁を合 成する乳腺組織内にも微生物が存在することを遺伝子レベルで明らかにした。マウスの
26
乳腺組織内では、分娩直後から細菌数が増加し、授乳期間に菌数が維持されることが認 められた。また、この乳腺内の細菌数は離乳後になると未経産と同程度まで減少した。 さらに、菌数の増加が認められた授乳期の乳腺組織内の微生物種を特定した結果、
Bacteroidales目、Clostridiales目、Lactobacillales目を主とする多種の微生物の存在が 確認された。これらの結果は、乳腺組織内で授乳期特異的に微生物環境が形成されるこ とを示唆している。
本研究で乳汁および乳腺組織内に検出された微生物の由来は未だ明らかではない。 乳汁中の微生物に関する近年の研究では、菌の由来として 2 つの仮説が報告されている (Fernandez et al. 2013; Moossavi and Azad 2019)。1つは、仔の口腔細菌が由来である という仮説である。この仮説では、乳汁中の微生物構成と仔の口腔内の微生物構成には 類似性があるという報告があることから、哺乳を介して仔の口腔内の微生物が母体の乳 頭口から侵入することで、乳汁中に微生物環境が形成される可能性が考えられている (Asnicar et al. 2017; Biagi et al. 2017; Ferretti et al. 2018)。また、もう一つの仮説は、 母体の腸内細菌が由来となり、免疫細胞によって腸内細菌が乳腺組織および乳汁中へ輸 送されるという仮説である。これは、乳汁中の微生物構成と母体の腸内の微生物構成に は類似性があること、そして、腸内の一部の樹状細胞は管腔内の微生物を補足後に細胞 内で補足した微生物を生かしたまま輸送するという報告があることから提唱され、一般 的 に Entero-mammary 経 路 と 呼 ば れ て い る (Jost et al. 2014; Farache et al. 2013; Macpherson and Uhr 2004)。これらの仮説は、未だ直接的な実証は行われていないもの の、ヒトの臨床研究やマウスによる部分的な実証研究などを通して、その可能性が示唆 されてきた。本研究では、乳汁合成の場である乳腺組織内における微生物の存在を明ら かにしたが、このような微生物もまた、乳汁中の微生物に関する研究で議論されている 仔の口腔内細菌または母体内の腸内細菌等を由来としていると考えられる。
27 誘導するのか否かを明らかにすべく、体内に微生物を保有しない無菌マウスと SPF 環 境で通常飼育した BALB/c マウスの乳腺組織における CCL28 と PigR の遺伝子発現を 比較検討した。その結果、両マウス間で CCL28 と PigR の遺伝子発現に大きな差は認 められなかった。これは、乳腺組織内の微生物環境が CCL28 と PigR による乳腺への IgA 形質細胞の遊走および分泌された IgA の腺房腔への輸送に深く影響しないことを 示している。このことから、乳腺の IgA 産生は、微生物非依存的に発達することが明ら かとなった。 乳腺の IgA 産生が発達する授乳期には、乳汁合成を主とする生理機能もまた発達 する。本研究では、乳腺組織における CCL28 と PigR の遺伝子発現は、乳タンパクで あるカゼインα、カゼインβ、カゼインκの遺伝子発現と正の相関を有することが示さ れた。このことは、乳腺の免疫機能が乳汁合成に関わる授乳期特有の制御によって誘導 される可能性を示唆している。乳汁合成は、分娩後に仔からの哺乳刺激を受けて、視床 下部から分泌されるオキシトシンと下垂体前葉から分泌されるプロラクチンなどのホ ルモンに加え、インスリンや糖質コルチコイドが乳腺上皮細胞に作用することで開始さ れる(Horseman 1999; Brisken and Ataca 2015)。また、CCL28 と PigR の発現に関する 先行研究では、子宮または鼻咽頭においてエストロゲンや糖質コルチコイドなどのホル モンが組織の CCL28 と PigR の発現を誘導することが報告されている(Cha et al. 2011; Gutierrez-Meza et al. 2017)。故に、乳腺における CCL28 および PigR の発現が哺乳刺 激下で乳汁合成を誘導するホルモンなどの内因性因子によって誘導される可能性が考 えられた。そこで本研究では、乳汁合成に必須のシグナルとなる哺乳刺激に着目し、哺 乳刺激が CCL28 と PigR の発現を誘導するか否かを調査した。分娩後 1 週間目に仔を 排除し、強制的に離乳させることで哺乳刺激を除いたマウスと通常飼育したマウスの乳 腺における CCL28 と PigR の遺伝子発現を比較検討した結果、哺乳刺激を排除したマ ウスで CCL28 と PigR の遺伝子発現が著しく低値を示すことが確認された。このこと
28 から、乳腺における IgA 産生には仔の哺乳刺激が必須であり、おそらく、哺乳刺激下で 誘導されるホルモンなどの内因性因子が機能することで乳腺の IgA 産生が誘導される と考えられる。 また、IgA が豊富に産生される粘膜組織では、多種の微生物が管腔内の IgA と結合 することで粘膜面に定着し、微生物叢を形成することが腸管を主とする粘膜組織で知ら れている(Donaldson et al. 2018; Pabst, Cerovic, and Hornef 2016)。本研究では、母体 の乳腺組織にも授乳期に多種の微生物が存在することが遺伝子レベルで明らかなった ほか、同時期に哺乳刺激を受けて IgA 産生が発達することが示された。そのため、IgA 産生と微生物環境が発達する授乳期の乳腺の場合もまた、腸管を含む他の粘膜組織と同 じく、IgA が作用することで微生物環境が形成される可能性が考えられた。しかしなが ら、授乳期の乳腺では、IgA の分泌が少ない分娩直後の時点で既に、IgA 分泌が盛んに 行われる分娩後 2 週間目と同程度の細菌数が存在することが確認された。さらに、分娩 直後と分娩後 2 週間目に存在する微生物種の構成を統計学的に比較しても有意な差は 認められなかった。このことから、IgA 産生が未成熟な分娩直後から乳腺組織内には既 に微生物環境が形成されることが示唆された。故に、乳腺組織では腸管と異なり、IgA 非依存的に微生物環境が形成されると考えられる。 以上のことから、乳腺では、授乳期に IgA を主とする免疫環境と微生物環境が其々、 非依存的に発達することが明らかとなった。このことは、母体の IgA 産生強化を図るた めには、免疫および微生物環境に対する個々のアプローチが必要であることを示唆して いる。特に、乳腺の IgA 産生の誘導には哺乳刺激が必須であることが本研究によって明 らかとなった。哺乳刺激下で CCL28 や PigR の発現に寄与する因子は、乳腺の IgA 産 生を向上させる鍵となると考えられる。
29 2.5 小括 本章では、乳腺における免疫機能の向上を目的として、IgA 産生に関わる分子の発 現機序を検討した。 ⚫ 乳腺の IgA 産生は、乳汁合成と同じく授乳期に限定して発達する時期特有の反応で あり、分娩後の授乳日数の経過に伴い、徐々に発達することが示された。 ⚫ 乳汁合成および IgA 産生が発達する授乳期の乳腺では、組織内で細菌数が増加する ことが確認され、多種の微生物が存在することが明らかとなった。この微生物環境 は、乳腺で IgA 産生が発達するよりも前に形成されることから、IgA 非依存的に乳 腺組織内で発達することが示唆された。 ⚫ 授乳期の乳腺組織内に存在する微生物は、同時期に発達する IgA 産生の誘導に深く 関与しないことが明らかとなった。乳腺組織内の IgA 産生の誘導には、微生物刺激 よりも仔からの哺乳刺激が必要であり、哺乳刺激下で機能する内因性因子が関与す る可能性が示唆された。 これらの知見から、母体の乳腺組織内では微生物環境と IgA を主とする免疫環境 が互いに非依存的に発達することが明らかとなった。そのため、母体の乳腺の免疫強化 を図るためには、乳腺の IgA 産生誘導に必須となる哺乳刺激を中心としたアプローチ が必要であることが示唆された。
30
図表
31
HE
IHC (
IgA
/
nucleus
)
分娩
6
週間後
分娩
2
週間後
分娩
1
週間後
分娩直後
分娩
1
週間前
未経産
Bar=50μmA
B
HE
IHC (
IgA
/
nucleus
)
分娩
6
週間後
分娩
2
週間後
分娩
1
週間後
分娩直後
分娩
1
週間前
未経産
Bar=50μmA
B
A
B
32 図 1 生殖サイクルに伴うマウスの乳腺組織の形態的変化 (A)乳腺組織の HE 染色画像 (B)抗マウス IgA 抗体を用いた乳腺組織の免疫化学染色画像 各染色画像は 400 倍で観察・撮影した。(茶;IgA、青;核) (C) 乳腺組織中の IgA 産生細胞数 0.05cm2の組織画像中に検出された IgA 産生細胞を測定し、グラフとした。 ** p<0.01 0.0 100.0 200.0 300.0 400.0
N
u
m
b
e
r
/
0
.0
5
cm
2400
300
200
0
100
-1
0
1
2
6
未経産
分娩後経過週齢
** **
**
** **
C
C
33 図 2 乳腺における生殖サイクルに伴う免疫関連遺伝子および乳タンパク遺伝子の 発現変化 (A) 乳タンパク遺伝子(csn1s1、csn2 、csn3)の発現量 (B) 免疫関連遺伝子(ccl28、pigr、 iga)の発現量 出産 1 週間前(n=7)、出産直後(n=9)、出産 1 週間後(n=7)、出産 2 週間後(n=9)、出産 6 週 間後(n=5)、無交配(n=5) * p<0.05 , ** p<0.01 , *** p<0.001 , **** p<0.0001
A
B
p ig r / g a p d h 4 3 2 0 1 -1 0 1 2 6 無交配 分娩後経過週齢**
**
***
***
****
ccl 2 8 / g a p d h 3 2 1 0******
*
*
*****
-1 0 1 2 6 無交配 分娩後経過週齢 6 4 2 0 ig a / g a p d h** *
****
** *
-1 0 1 2 6 無交配 分娩後経過週齢 csn 1 / g a p d h 15x103 10x103 5x103 0**
****
****
****
-1 0 1 2 6 無交配 分娩後経過週齢 cs n 2 / g a p d h 10x106 5x106 0*
*
*****
***
****
-1 0 1 2 6 無交配 分娩後経過週齢 csn 3 / g a p d h 0.6 0.4 0.2 0* **
*****
***
****
-1 0 1 2 6 無交配 分娩後経過週齢A
B
34
図 3 3 種類のカゼイン遺伝子に対する CCL28 および PigR 遺伝子の相関
供試した全ての群における各遺伝子発現量を用いて、2 つの遺伝子間の相関を求めた。 (A) Casein alpha(csn1s1)遺伝子と CCL28 遺伝子の相関
(B) Casein beta(csn2)遺伝子と CCL28 遺伝子の相関 (C) Casein kappa(csn3)遺伝子と CCL28 遺伝子の相関 (D) Casein alpha(csn1s1)遺伝子と PigR 遺伝子の相関 (E) Casein beta(csn2)遺伝子と PigR 遺伝子の相関 (F) Casein kappa(csn3)遺伝子と PigR 遺伝子の相関
3 2 1 0 ccl28 r = 0.7537**** 8x103 4x103 0 12x103 csn 1 s1 16x103
A
3 2 1 0 ccl28 r = 0.7633**** 6x106 4x106 2x106 0 8x106 cs n 2B
3 2 1 0 ccl28 r = 0.8251**** 0.4 0.3 0.2 0 0.1 cs n 3C
3 2 1 0 pigr 4 8x103 4x103 0 12x103 csn 1 s1 16x103 r = 0.8809****D
6x106 4x106 2x106 0 8x106 csn 2 3 2 1 0 pigr 4 r = 0.9191****E
0.4 0.3 0.2 0 0.1 cs n 3 3 2 1 0 pigr 4 r = 0.9061****F
-1 0 1 2 6nulliparaweeks after parturition
A
B
D
E
35 図 4 ヒト母乳中の微生物環境 分娩後 1 か月時のヒトの乳汁中微生物叢をメタゲノム解析により明らかにした。 16s rRNA メタゲノム解析によって特定された 359 種の内、2%以上の存在が確認された 菌種でグラフを作成し、2%以下の細菌種は Others として表記した。 被験者番号 1 2 3 4 5 0 10 20 30 40 50 60 70 80 90 100 P e rce n ta g e ( % ) Family Species Order Streptococcus vestibularis Streptococcus pseudopneumoniae Streptococcus infantis Streptococcus tigurinus Streptococcus oralis Streptococcus infantarius Streptococcus bovis Streptococcus anginosus Streptococcus parasanguinis Lactobacillus antri Staphylococcus hominis Staphylococcus aureus Staphylococcus epidermidis Staphylococcus caprae Staphylococcus lugdunensis Gemella haemolysans Gemella sanguinis Gemella cunicula Prevotella multisaccharivorax Prevotella disiens Prevotella stercorea Dysgonomonas wimpennyi Parabacteroides goldsteinii Bacteroides acidifaciens Corynebacterium simulans Corynebacterium kroppenstedtii Corynebacterium tuberculostearicum Agrobacterium tumefaciens Haemophilus parainfluenzae Treponema succinifaciens Brevundimonas diminuta Megasphaera elsdenii Clostridium histolyticum Streptococcaceae Lactobacillaceae Staphylococcaceae
Bacillales Family XI. Incertae Sedis
Prevotellaceae Dysgonamonadaceae Tannerellaceae Bacteroidaceae Corynebacteriaceae Rhizobiaceae Pasteurellaceae Spirochaetaceae Caulobacteraceae Veillonellaceae Clostridiaceae Lactobacillales Bacillales Bacteroidales Corynebacteriales Rhizobiales Pasteurellales Spirochaetales Caulobacterales Veillonellales Clostridiales Others
36 図 5 ウシ乳汁中の微生物環境 ウシ乳汁中微生物叢をメタゲノム解析により明らかにした。 16s rRNA メタゲノム解析によって特定された 1788 種の内、4%以上の存在が確認された 菌種でグラフを作成し、4%以下の細菌種は Others として表記した。 4 分房別に乳汁を回収し、解析に供した。分房の表記は以下のとおり。 RF:右前分房由来の乳汁サンプル RB:右後分房由来の乳汁サンプル LF: 左前分房由来の乳汁サンプル LB: 左後分房由来の乳汁サンプル Sample No. 0 10 20 30 40 50 60 70 80 90 100 P e rce n ta g e ( % ) Family Species Order 1 2 3 4 RF RB LF LB RF RB LF LB RF RB LF LB RF RB LF LB Lachnospiraceae Clostridiaceae Ruminococcaceae Hungateiclostridiaceae Lachnospiraceae Bacteroidaceae Prevotellaceae Porphyromonadaceae Corynebacteriaceae Pseudomonadaceae Moraxellaceae Intrasporangiaceae Micrococcaceae Dermabacteraceae Staphylococcaceae Planococcaceae Bacillaceae Leuconostocaceae Streptococcaceae Bifidobacteriales Erysipelotrichaceae Sphingobacteriaceae Flavobacteriaceae Comamonadaceae unclassified Calotrichaceae Coriobacteriaceae Acetobacteraceae Caldilineaceae Chromobacteriaceae unclassified Clostridiales Bacteroidales Corynebacteriales Pseudomonadales Micrococcales Bacillales Lactobacillales Bifidobacteriales Erysipelotrichales Sphingobacteriales Flavobacteriales Burkholderiales Nostocales Coriobacteriales Rhodospirillales Caldilineales Neisseriales unclassified Blautia coccoides Blautia hansenii Pseudobutyrivibrio xylanivorans Blautia wexlerae Blautia producta Caloramator mitchellensis Alkaliphilus peptidifermentans Alkaliphilus crotonatoxidans Clostridium histolyticum Ruminococcus callidus Ruminococcus bromii Clostridium alkalicellulosi Oribacterium sinus Bacteroides denticanum Bacteroides sartorii Paraprevotella clara Prevotella buccalis Prevotella aurantiaca Porphyromonas canis Corynebacterium stationis Corynebacterium xerosis Corynebacterium amycolatum Corynebacterium ammoniagenes Corynebacterium lubricantis Corynebacterium accolens Corynebacterium nuruki Pseudomonas viridiflava Pseudomonas lutea Pseudomonas luteola Pseudomonas azotoformans Pseudomonas mendocina Acinetobacter indicus Acinetobacter lwoffii Acinetobacter johnsonii Kocuria rhizophila Micrococcus yunnanensis Citricoccus alkalitolerans Brachybacterium conglomeratum Brachybacterium faecium Serinicoccus marinus Staphylococcus haemolyticus Staphylococcus chromogenes Sporosarcina pasteurii Bacillus infernus Weissella thailandensis Streptococcus parauberis Bifidobacterium catenulatum Bifidobacterium pseudolongum Erysipelothrix muris Coprobacillus cateniformis Pedobacter kwangyangensis Sphingobacterium shayense Wautersiella falsenii Chryseobacterium joostei Comamonas testosteroni Tepidimonas taiwanensis Calothrix parietina Collinsella aerofaciens Roseomonas terpenica Caldilinea tarbellica Aquitalea magnusonii
Microvirus Enterobacteria phage PhiX174 Others
37 出産直後 出産14日目 1 2 3 4 5 6 7 8 9 10 Animal No. 100 90 80 70 60 50 40 30 20 10 0 P e rce n ta g e ( % ) Mucispirillum schaedleri Parabacteroides goldsteinii Porphyromonas canis Blautia coccoides Calothrix parietina Bacteroides acidifaciens Blautia wexlerae Johnsonella ignava Oscillospira eae Lactobacillus taiwanensis Bacteroides xylanisolvens Odoribacter denticanis Dysgonomonas wimpennyi Alkaliphilus crotonatoxidans Escherichia albertii Peptoniphilus coxii Bacteroides sartorii Prevotella dentasini Ruminococcus gnavus Lactobacillus antri Serratia entomophila Sphingomonas desiccabilis Aeromicrobium kwangyangensis Corynebacterium acetoacidophilum Lysobacter deserti Lactobacillus vaginalis Enterococcus gallinarum Aeromicrobium ponti Sphingomonas oligophenolica Campylobacter hyointestinalis Hydrogenophaga defluvii Corynebacterium minutissimum Others Bacteroidaceae Odoribacteraceae Prevotellaceae Bacteroidales Lachnospiraceae Ruminococcaceae Clostridiaceae Clostridiales Lactobacillaceae Enterococcaceae Enterobacteriaceae Enterobacteriales Sphingomonadaceae Sphingomonadales Nocardioidaceae Propionibacteriales Corynebacteriaceae Corynebacteriales Deferribacteraceae Deferribacterales Rivulariaceae Nostocales Peptoniphilaceae Tissierellales Xanthomonadaceae Xanthomonadales Campylobacteraceae Campylobacterales Comamonadaceae Burkholderiales Porphyromonadaceae Family Species Lactobacillales Order A B b c ac bc abc b C o p ie s / mg 0 200 100 -1 0 1 2 6 無交配 出産後経過週齢 授乳期間
A
B
38 図 6 マウス乳腺内における微生物環境 (A)生殖サイクルを通した乳腺組織内における tuf 遺伝子の発現量 分娩 1 週間前(n=7)、分娩直後(n=7)、分娩 1 週間後(n=8)、分娩 2 週間後(n=7)、分娩 6 週 間後(n=8)、無交配(n=7) a,b,c<0.05 (B)分娩直後と分娩 14 日後の乳腺組織内細菌叢の構成。 16s rRNA メタゲノム解析によって特定された 780 種の内、4%以上の存在が確認された。 菌種でグラフを作成し、4%以下の細菌種は Other として表記した。 分娩直後(n=5)、分娩 14 日後 (n=5) 図 7 IgA 産生関連遺伝子(ccl28、pigr)に対する乳腺環境変化の影響 (A) CCL28 遺伝子の発現量 (B) PigR 遺伝子の発現量 分娩 2 週間後の正常授乳群(n=9)、分娩 2 週間後の強制離乳群(n=5)、分娩 2 週間後の無菌 マウス群(n=4) * p<0.05 , ** p<0.01
ccl
2
8
/
g
a
p
d
h
2 0 -2 -4**
*
ns 1 -1 -3 log10p
ig
r
/
g
a
p
d
h
**
*
*
2 0 -2 -4 1 -1 -3 log10A
B
通常授乳 強制離乳 無菌 SPF 通常授乳 強制離乳 無菌 SPFA
B
39
第三章
離乳期の仔の腸管における免疫
および微生物環境の形成機序解明
40 3.1 はじめに
仔の腸管では、離乳後に母体からの移行 IgA 抗体が減少することに伴い、仔自身の IgA 産生が徐々に発達する。しかしながら、仔自身の IgA 分泌が始まる離乳後早期は、 成体と比較して、IgA 産生の発達程度が非常に未成熟であり、感染症に対する罹患率が 高い(Dupont et al. 2016; Green et al. 2018)。故に、感染症を予防し、仔の健全な育成を 促す為には、離乳後早期の IgA 産生機序を正しく理解し、迅速に誘導させることが重要 である。 腸管の IgA 産生は、パイエル板を代表とする腸管関連リンパ組織に微生物を含む 抗原が取り込まれることによって開始されることが成体マウスを用いた研究で明らか とされてきた(Ohno 2015)。組織内に取り込まれた抗原は、樹状細胞などの抗原提示細 胞により捕捉され、細胞内で処理された後、その抗原情報が組織内の T 細胞に伝えら れる。抗原情報を得た T 細胞は、同じ抗原を認識する B 細胞を刺激することで、B 細 胞の IgA クラススイッチと親和性成熟を誘導する。そして、成熟過程を経た B 細胞は、 IgA を分泌する形質細胞へと最終分化し、絨毛下の粘膜固有層へと遊走することで粘膜 面での IgA 分泌を開始することが一般的に知られている。故に、幼若期の仔の腸管にお いて IgA 産生を効率的に開始させる為には、腸内に免疫賦活化能を有する抗原が定着 すること、そして、そのような抗原が腸管関連リンパ組織へ取り込まれることが必要と なる。しかしながら、幼若期の仔の腸管において、腸管関連リンパ組織内に抗原が取り 込まれる時期、そして、取込み後に IgA 産生を誘導する抗原の特徴は、未だ明らかとさ れていない。 本研究では、仔が乳汁を摂取する哺乳期に、母体の乳腺組織内で微生物環境が形成 されることを第二章で明らかとしてきた。乳汁に関するヒトの臨床試験では、乳汁中に 存在する微生物叢の構成と仔の腸内細菌叢の構成に類似性があることが報告されてお り(Heikkila and Saris 2003; Martı́n et al. 2004; Jost et al. 2014)、このことから、乳汁の
41 産生場所である乳腺組織内に存在する微生物は、おそらく、哺乳を介して仔の腸管へ移 行し、仔の腸管粘膜へ定着すると考えられる。さらに、そのような母体の乳腺組織を由 来とする微生物は、母体から供給される哺乳期から腸管関連リンパ組織に取り込まれる ことで仔の IgA 産生の抗原として寄与する可能性がある。 そこで本章では、仔の IgA 産生に対する哺乳期の母体由来微生物の影響を明らか にすべく、腸管関連リンパ組織の一つであるパイエル板に着目し、哺乳期から離乳後の 成育過程を通してパイエル板の微生物環境および免疫環境を調査することで、仔のパイ エル板における母体の乳腺組織由来微生物の取込みと、取り込まれた微生物が IgA 産 生の誘導抗原として機能する可能性を検証した。
42 3.2 材料および方法 3.2.1 供試動物 SPF 環境下で自家繁殖した BALB/c マウスから 2 週齢時(哺乳期)、3 週齢時(離乳 期)、4 週齢時(離乳 1 週間後)、11 週齢時にパイエル板と小腸組織を採材した。また、 無菌環境下で飼育し、体内に微生物を保有しない無菌マウス(SLC)と SPF 環境で飼育 した BALB/c マウス(自家繁殖)から 4 週齢時に大腸組織と糞便を回収した。 3.2.2 採材 2 週齢時、3 週齢時、4 週齢時、11 週齢時の BALB/c マウスから回収したパイエル 板は、細胞を単離後にフローサイトメトリー解析に供した。また、パイエル板から単離 した細胞の一部は、パイエル板内に存在する微生物種を明らかにする為にゲノム DNA の抽出を行い、微生物特有の遺伝子領域である 16S 配列(V3-4 領域)を PCR で増幅する ことで次世代シーケンサーによる細菌叢のメタゲノム解析に供した。パイエル板と同じ タイミングで回収した小腸組織はパイエル板を取り除かずに 4%PFA で固定後、パラフ ィン包埋した。包埋した小腸組織は、ミクロトームを用いて 5µm の厚さに薄切するこ とで組織切片を作製し、組織学的解析に供した。 4 週齢時の無菌マウスおよび SPF 環境下で飼育した BALB/c マウスから回収した 大腸組織は、細胞を単離後にフローサイトメトリー解析に供した。また、同個体から回 収した糞便は、重量を測定後に 3.2mm3ステンレスビーズ(TOMY)を入れたサンプルチ ューブ(TOMY)に移し、糞便 1mg 当たり 10µL の PBS を加えた後、破砕機を用いて懸 濁し、15000rpm で 10 分間、遠心操作することで上清を回収した。回収した糞便上清 は、ELISA を用いた糞便中の IgA 濃度測定に利用した。
43 3.2.3 パイエル板からの細胞抽出 採材したパイエル板は、PBS を用いて 2 回洗浄した。洗浄後、pH8.0、0.5M EDTA を 50µL 添加した 25mL の RPMI(gibco)へパイエル板を移し、37℃の恒温槽で 25 分間、 振盪しながら温めることで上皮層を取り除いた。上皮剥離後、PBS を用いてパイエル板 を 2 回洗浄した。FBS を添加した 15mL の RPMI に DNase(Sigma)0.04mg、コラゲナ ーゼ(Sigma)7.5mg、メルカプトエタノール 0.06mg を加え、そこへパイエル板を移し、 37℃の恒温槽で 60 分間温めながらコラゲナーゼと反応させることにより組織構造を破 壊し、細胞を単離した。コラゲナーゼ反応後、100µm フィルター(FALCON)と 40µm フ ィルター(FALCON)へ細胞液を通し、濾過した。さらに 1500rpm で 10 分間遠心操作 を行い、上清を廃棄後、PBS で再懸濁することで、パイエル板の細胞懸濁液を作製した。 3.2.4 小腸・大腸からの細胞抽出 採材した小腸および大腸組織は、PBS を用いて 2 回洗浄した。洗浄後、組織を 1cm3 角の組織片となるように切断し、pH8.0、0.5M EDTA を 50µL 添加した 25mL の RPMI(gibco)へ加えた。組織片を加えた培地を、37℃の恒温槽で 25 分間、振盪しなが ら温めることで腸管上皮を取り除いた。上皮剥離後、PBS を用いて腸管組織を 2 回洗 浄した。洗浄した組織は、DNase(Sigma)0.04mg、コラゲナーゼ(Sigma)7.5mg、メルカ プトエタノール 0.06mg を加えた 15mL の RPMI に移し、37℃の恒温槽で 75 分間温め ることでコラゲナーゼと反応させ、組織構造を破壊し、細胞を単離した。コラゲナーゼ 反応後の液を 100µm フィルター(FALCON)と 40µm フィルター(FALCON)に通し、濾 過した。さらに 1500rpm で 10 分間遠心操作を行い、上清を廃棄後、PBS で再懸濁す ることで、小腸および大腸組織の細胞懸濁液を作製した。