F I I I I I I I I I I I I I I I I I I I I I I L
林木の支配 に関する基礎的研 究
(Ⅷ
)ヒノキの球果の発達 に対する受粉の役割 および
単為結果の人為的誘起
橋
詰
隼
ガ
FundamentaI Studies on Mating
Effect of Pollination on Cone
Artificial induction of Parthenocarpy
in Forest Trees(Ⅷ
)Development and
in C力attβ
ecyparis obrtrsa
HayatO、
HASHIZUME※
Summary
Tl・le female strObili of Cんα胸9ctypαT,s οbιttsα did not develop into cones
,vithout ponination. They developed into cones by the ponination with ponen
Of CγρT9sscC9α9 and T9″ο 'テ
αc9α9, but not by the pollination of F),η αc9α9 and Taπαccα9, Parthenocarpy was also induced by spraying with lvater solutions of 500 ppm GA3 and l% fruitone. lt seems tlaat Cん 。 οbιttsα is a plant of
stimulative(aitonomic)parthenOcarpy.
It was found that cone deve10pment in Cん 。οbιttsa is closely connected、vith
ovular development, In the case of non― pollination and ponination 4rith dis―
tantly related pollens, the o■ rtlles degenerated at the early stage of embryosac
development, and the female strobili did not develop into cones, It seems that cone development is controlled by such growth resulators as auxins and gibberellins, being produced in ovllles.
Altllollgh pollinatiOn was necessary for the development of ovules in凱.
Obを切sc, he extremely distant pollens did not ger■ linate in the pollen chamber
or showed negative tropism, and the o、 ′ules degenerated at the early stage of he development. Since the Ovllles of Cん . οbtttsc are developed normally by tlle pollination of dead pollen of he same species,it is hougllt tllat he Ovlllar deヽrelopment is induced by the special stirnulative substances existing in pollen grains, It seems that these substances are insoluble in 、vater and alcohols, and resistant to heat, but inactivated by O.l N HCl and O.l N
NaOH.
※ ′島取 大学農 学部 造 林学 研 究 室;Laboratory of Silviculture,Faco of Agr., Tottori Uni砿 ,Tottori 680
(52)
王
まえが き
林木の育種においては種間交雑による雑種の育成,無
受粉,活
力のない花粉の交配あるいは遠縁花 粉の交配などによる半数体の育成などが試験 されているが,落
果,不
結果,交
雑不捻などの現象があ って必ず しも良い結果が えられていない。針葉樹には受粉 しなければ結果 しない ものと,無
受粉で も 結果するものとがある?∼°カラマツ属, モ ミ属,
トウヒ属, ツガ属,
トガサワラ属, スギ属,
ヒノキ 属,
ネズコ属,
ビャクシン属などの多 くの樹種では容易に単為結果することが知 られているが,マ
ツ 属は単為結果のむずか しい ものが多 く,勝
田らつめの報告によると,ク ロマツ,
アカマツでは個体によ っては無受粉のとき落果が著 しく多い ものがある。落果せず,球
果が正常に発達しなければ種子をと ることができないか ら,球
果の発達の仕組み,つ
ま り結果,結
実の生理を研究す ることは林木育種上 大変重要である。本研究においては,
ヒノキについて無受粉および色々な花粉を受粉 したときの球果 の発達の状況,ホ
ルモン剤の散布による単為結果の誘起などについて研究 したので報告する。Π
材料 と方 法
鳥取大学農学部樹木園内の約15年
生 ヒノキを母樹 として,無
受 粉,各
種花粉およ び花粉抽出物の 人工受粉,無
受粉雌花 に対す るホルモ ン剤処理な どの試験 を行 った。人工受粉 に用 いた花粉 は樹木 園 内の植栽木か ら開花期の直前 に枝 をきりと り,室
内で水挿 して採取 した。アカマツは前年採取 の貯蔵 花粉 を,そ
の他の樹種は当年採取の新鮮花粉 を用 いた。生存花粉の発芽率は,
イ トスギ46%,ア
スナ ロ,コ
ノテガシヮ61%,キ
ャラボク69%,そ
の他の樹 種 は90%以
上 で あ った。死滅施 粉は,9 0CO で24時
間乾燥処理 して用いた。花粉成分の抽出方法は,
ヒノキの新 鮮花 粉1量を水,メ
タノール,0.lNHClお
よび01 N NaOHで
抽出 して ,抽 出物 と残冶 に分 けた。それぞれ を乾圃 させた後,抽
出 物 は500野
のタルクと混 合 して受粉 し,残
澄 は粉砕 して直接受粉 した。袋掛は,東
洋理 光器KK製 交配 袋 (ヒ ノキ用 )を 用い,開
花 の直前 に行 った。人工受粉は雌花の満 開初期 に行 った。混合花粉の受粉 は,ヒ ノキの死減花粉 と他種の生存花粉 とを1:1の
比率(重量比)で混合 して用いた。 単為結果誘起のための ホルモ ン剤の処理 は,粉
斉」処理法 と水溶液散布法の二つの方法 によ って行 っ た。粉剤処理法 は,GA3,IBA,NAAを
それぞれ1%と
5%の
割 合で,ま
た フル ー トン(d-2,4,5-TP)を
5%と
20%の
割 合 で タルクに混 合 して受粉 した。水溶液散布法は,表
2の如 く,GA3とIBA
は100p即と500p側の濃度で,NAAは
100p脚で,カ イネチ ンは2 0 0ppmで,フ ルー トンは0.1%と
1%
で用い, 4月 1日 か ら4月28日
の間に4回,交
配袋 をか けた無受粉のllC花着生枝 に葉面散布 した。Ⅲ
結 果 と考 察
1.結
果 ・結実 におよぼす受粉の影響
ヒノキの生存花粉 と死滅花粉の受粉および無受粉 の ときの球果の生長経過は図1の如 くであ る。 ヒ 隼 詰 橋林木の交配に関す る基礎 的研究(Ⅷ ) ノキの雌花 は受粉 によ って正常な球果 に発 達す るが
,無
受粉では球果 に発達 しない。 球果の発育 は生存花粉 の受粉で最 もよ く, 死滅花粉の受粉ではやや劣 る。lLt花は受粉 後 4月 と 5月 に急速 に生長 して球果 に発達 し, 6月
下旬 には生長が ほば停止す る。受 精期 は 6月 上旬で あるか ら,受
精期 まで に 球果 の生長 はほぼ終 わ る。以上の如 く,
ヒ ノキの雌花 は花粉の生死 と無関係 に受粉 に よ って球果 に発達す る。す なわ ち,
ヒノキ は刺激的(他動 的)単
為結果 をす る植物 で あ るとい える。 ヒノキの雌花 に同種 および異種の色 々なApr. May Jun, 」ul, Augo Sep. 図
1
球果 の発達におよぼす受粉の影響出
g・設乳想
tp♀学研
lbιttD C°nc A I PollinatiOn Of ive pollen
B I Pollinat10n of dead Ponen
C I Non― pollination P i Time of ponination F:Time of fertilization
Pollen grains of Cん .οb,Ir sα were
used ︵ ミ ミ ︶ H ! o 日 綴 や 0 再 O C 靱 回 Q 昧 皆 表
1
ヒノキの結果 ・結実 にお よぼす各種花粉の影響Table l Effects of variOus pollens on cone bearing and secd bearing in Cん o昴occγPaJrd οbι,sα
交 配 花 粉
P01len used for pollination
交配lH4花数
No Of fe―
naale flo、wers
p01linated 結 果 率 C∩nn sct 球 果 の直 径 COne diametcr e2) 種子 の形成 S eed fOrmatiOn 単為 結果 に 対す る効 果 Effect on partheno― carpy 平 均 Averagc膀 鈎 囲 t 銅 ang R ヒノキ(生存)
0ん。P/ta cc,7婢rιs Obι,dα (hve) アスナ ロ(生存)
TんItyοPs,s Jοどαbrαια(li ve) コノテガシ ワ(生存) Tん巧αο′ιettοιιs(l c) イ トスギ(生存) C,Pressレs sp.(hVe) スギ(生存) CrヵDヶοれc rt, デcPο 'ウ C。(live) アカマツ(生存) Pれ,s」針げJοr,(l e) キ ャラボ ク(生存) Tα
″
ys c,spt」ο
rD vak言与
き
, ヒノキ(死滅) Cん。οb,wsc(dead) スギ (生 存 )十 ヒノキ(死滅) Cr ブDP ο ttιCc(hve)十Cん。 οbιyso(dcad) アカマ ツ(生存 )十 ヒノ キ (死滅)P.」e,sげιο′,(li Ve)十 Cん οbιfrsc(dead) キ ャラボ ク(生存 )十 ヒノキ (死滅)
作灘覧」
π島温が
°
'α(1つ280
250
168
66
459
176
133
413
325
377
352
964
698
385
460
344
12 06894
750
846
618
89∼100
0∼100 3∼100 0∼92 1∼86 0∼ 6 0∼3 58∼100
32∼100 48∼100
12∼100
113
85
88
7175
46
49
107
100
101
96
十 一 ∼ 十 一 ∼ 十 一 ∼ 十 一 ∼ 十 一 ll ―l■l 十 十 十 + 十←) 十←) 十11+()
十 十 十 十花粉 を受粉 した結果 は表
1,写
真1の如 くで あ る。 ヒノキの雌花 は ヒノキ科 とスギ科の花粉の受粉で癬 果 したが,マ
ツ科 とイチ イ科の花粉では殆 ど結果 し なか った。 しか し,マ
ツ科 とイテイ科の花粉 に ヒノ キの死滅花粉 を混合 して受粉す ると結果 した。 ヒノ■ 暇
}.常
宰.彎
!│が
彎 ―彎 「キの球果の発達に対するこれらの花粉の刺激効果は
, 1 2 3 4 56 7 8
ヒノキ(生
存)>ヒ
ノキ(死滅)■混合 受 粉>ア
ス ナ ロ■ コノテ ガ シ ワ>イ
トスギ■ スギ>ア
カマ ツキ写真
1
ヒノキの球果の発達におよぼす 各種花粉の影響 キ ャラボ クの順 で,植
物分 類学 上 の類 縁 関係 と大 体PhOt0 1
号 :≒:is i謂賊:¶
乳 。l: 一 致 し,近
縁 の もの ほ ど効果 が大 き く,速
縁 の ものde℃
bpcme h C/J.οbι,sα l:×Cん οbιysrL(hve) は効果 が み られ なか った。 また ヒノ キの母 樹 に よ っ2:×
Cん。οbιtts。 (deaD 3:× Tん 。 JοιαbrDι。(li ve) て異種花粉 に対す る反応 に著 しい ちがいがみ られた。4:×
Cr,ブ叩οttθ。(li ve) 5:×Ta,c,sP'Jαι。(h ve) 例 えばスギの花粉の交配で結果す る母樹 と結果 しな6:×
P」9,sttιο「α(livc) い母樹 とがあ った。 これ らの交配で結果 した もので・
XT中
夢J防°(hveJ■路 を糾 は種子が形成 されたが,
ヒノキ生存花粉の受粉 を除8:×
P.」9,sげιο′C(li ve)十駿ぢを密│ き種子は不捻であ った。 各種花粉の交配 にお け る花粉の行動 と胚珠組織 の発達 の状況 を顕微鏡で調べ た結果 は次の通 りで あ る。 ヒノキ生存花粉の受粉:花粉粒 は 4月 下旬 に花粉室で発芽 して珠心 に侵入 した。胚珠 は正常に発達 して造卵器 を形成 し, 6月
上旬 に受精 して胚が発生 した。 ヒノキ死滅花粉の受粉 :花粉粒 は発芽 しなか ったが,珠
心頂部 に付着 した。胚珠 は正常 に発達 して 造卵器 を形成 したが,受
精せず 7月 以降に退化 した。 アス ナロ,
コノテガシワ,ス
ギの受粉:花粉粒 は 4月 下旬に花粉室で発芽 したが,珠
心への侵入は ヒノキ生存花粉 の場合 よ りも少 しお くれた。花粉管の伸長 は緩慢で,受
精期 まで に造卵器 に達 しなか った。 ヒノキ×アスナ ロの交配では, 6月
25日
に花粉管 は珠心肉部 に約170μ
侵入 し,珠心頂部か ら造卵器 までの距離の約1/3の
所で停滞 していた。胚珠の発達状況 は,胚
の う発生の初期に退化す る 場合,遊
離核形成期か ら胚乳体形成期で発育が停止 し,造
卵器が形成 されない場合,正
常 に発育 して 造卵器が形成 され る場合の二つがみ られた。いずれの場合 も受精 しない。 アカマツ花粉の受粉:花粉粒 は花粉室内で発芽 しなか った。胚珠 は胚の う発生の初期 に退化 し,収
縮褐変 した。 キ ャラボ ク花粉の受粉 :花粉粒 は花粉室内で発芽 したが,珠
心 に侵入せず,珠
心 と逆 の方 向に伸長 し負の屈 向性 移示 した。胚珠 は胚の う発生の初期 に退化 し,収
縮褐変 した。 無受粉:胚珠 は胚の う発生の初期(5月
上旬)に
退化 し,収
縮褐変 した。 遠縁花粉の受粉 におけ る花粉 の行動 と胚珠組織 の発達の状況 を要約す ると次の如 くである。F I I I I I I I I I I I I I I I I I I I I I I I I L 林本の交配 に関す る基礎 的研究(Ⅷ )
(a)花
粉は発芽しないか,発
芽す るが珠心に侵入せず,胚
珠は退化枯死する。 (bl 花粉 は発芽 して珠心に侵入するが,花
粉管の伸長が緩慢である。また胚珠組織の発達が不十分 で造卵器が形成されず,受
精できない。(c)胚
珠は正常に発達 して造卵器が形成 され るが,花
粉管の伸長が緩慢で受精で きない。2.結
果・結実におよぼす花粉抽出物およびホルモン剤の影響
ヒノキの新鮮花粉
1分を冷水
(室
温で24時 間
, 50π″×
3回),温
水(70℃ で5時 間
,5 0 Tlly×3回
),80%冷
メタノール
(0℃
で24時 間
,50腿
×
3回)お
よび
80%温
メタノール
(70℃
で
5時 間, 50紀″X3回
)で
抽 出 し,抽
出物 と残冶 に分 けて受粉 した。 さ らに新 鮮花粉 1量 を01NHClお
よ び0.lN NaOH(100℃
で5時
間,50腿
×2回 )で加 水 分 解 して,
抽 出物 と残 湾 に分 け同様 に 人工受粉 した。 実験の結果,水
お よび メタノール抽 出物の受粉 では雌花 は球果 に発達せず,残
澄の受粉 で結果 した 。O.l N HClお
よび0,l N NaOHの
抽 出物 および残澄の受粉で は結果 しなか った。 ヒノキの花 粉 の中 に含まれてい る球果 の発達 を誘起す る物質は水や アルコールに不溶で,熱
にかな り安定な物質のよ う に思われ る。 しか し,酸
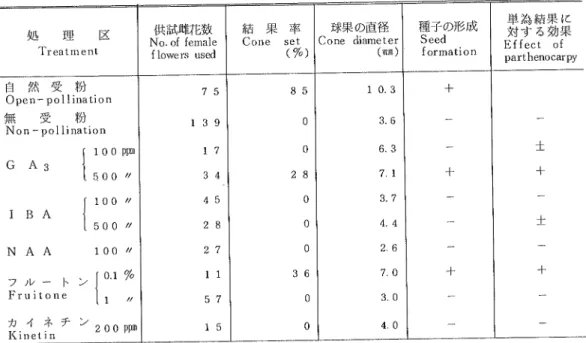
や アルカ リで は分解 され るよ うである。 表2
ホルモン剤によるヒノキの単為結果の誘起Table 2 Artificial ねduction of parthenocarpy in Cん。οb′,sc by spraying with growth regulators
処
理 Treatmcnt
区 供 試 雌花 数
No.of fernale
f iO■veぃ used
結 果 率 Cone set
(%)
球果 の直径 Cone diameter (2π) 種 子 の形 成 Seed fOrmatio n 単 為 結 果 に 対 す る効 果 Effect of parthenOcal v 自 然 受 粉 Open―pollinatiOn 無受
粉 Non―pollination
G A3 1:;││)
IBA I:÷
│;
NAA 100〃
盈
雀
津 ン
1干
子
誕
ittt亀チン
2 0 0 ppm75
139
17
34
45
28
27
1 157
15
85
0 028
0 0 036
0 0103
86
63
71
37
4 426
70
30
40
十 十 十 十 ■ 十 十 備考 :4月 1日∼28日の間に 4回 葉面散布す る。 無受粉のlht花に ホルモ ン剤 を散布 した結果 は表2, す るホルモ ン剤の効果 は処理 の方 法 によ って異 な り, 写真2の
通 りである。 ヒノキの球果 の発達 に対 粉剤処 理法 はあま り効果が な く,水
溶液散布法(56) 橋 詰 隼 人 Ч 口 E I I l l l I I I I I I I I I I I I I I I I I I I I I I I I I I I I I I I ゴ
⇔
o●
0
こ
―
│:子
:
魅
忌
筑
竜
塩
手
辻
島
1警-1 2 3 4
写 真2
ホルモ ン却に よ る単 為結果 の 誘起Phot0 2 Altificial inductiOn Of pal―
thenocarpy in Cん. οb ιレdο
by spraying with GA3 and
fl・uitOne
l:NOn―polli natiOn
2:01%fruitone 3:500ppal GA3 4i Open―pollination
3.受
粉 と胚 珠 の発生 との関係
が有効のよ うであ った。単為結果 はジベ レ リンとフ ルー トンの散布区で認め られた。IBA,NAA,カ
イ ネチ ンの散布 区で は耳果 は殆 ど発育 しなか ったが, これは これ らのホルモン剤が単為結果の誘起に無効 であるのか,あ
るいは処理方法が適当で ないために 効果が現 れなか ったのか明 らかで ない。GA3500p脚
区 とフルー トン0.l ppm区の単為結果 した球果 は 自然 受粉球果 の3分
の2程
度 まで大 きくな り,中
に小 さ な種 子が認め られた力L結
果 率 はあま り高 くな く, 中には薬害 によ り枯死 す る もの もあ った。また個体 によ って ホルモ ン剤の効果 に差が あ り,
ジベ レ リン や フルー トンの効果 が顕著に現 れない もの もあるよ うで あ った。 前述の実験の通 り,
ヒノキでは受粉 しなければ胚珠は胚の う発生の初期 に退化 し,配
偶体が形成 さ れない。また雌花 は球果 に発 達 しない。難花 が 球 果 に発 達 す るためには,受
渤 によって胚珠が発育 す ることが必要 で あ る。花粉 とlVHずい(胚珠 )と の間には相互作用が あ り,極
端 な遠 縁花粉の交配で は花粉が発芽 しないか,発
芽 して も珠心に侵入す ることがで きない場合が多 い。 アカマツの花粉 は ヒ ノキの胚珠 内で発芽 しない し,キ
ャラボ クの花粉 は発芽す るが負の屈 向性 を示 し花粉管が珠心に侵入 しない。いずれの場合 も胚珠は早期 に退化す る。 ヒノキにアスナロ,
コノテガ シワ,
スギな どの花粉 を交配す ると,造
卵器 の形成まで胚珠 を発達 させ ることがで きるが,そ
れ以前の段階,す
なわ ち胚の う発生の初期,遊
離核形成期,胚
乎し体形成期 な どで胚珠 の発達が とま り退化が起 こる場合が多い。針 葉樹ではかな り遠縁の花粉 で も受粉 した花粉が花粉室内で発芽 し,花
粉の発芽に関 してはあま り種特 異性がみ られないが,花
粉管の珠心へ の侵入 と珠心組織 肉の伸長 については植物 の類縁関係が強 く作 用す るよ うで,近
縁 の ものほ どスムースにゆ く。 しか し,
ヒノキの胚珠の正常な発達 は,同
じ種 の死 滅花粉 の交配 で起 こるか ら,花
粉の生死お よび胚珠 肉にお ける花粉の発芽の有無 とは直接 関係がない。 おそ らく花粉の中に含まれてい る特殊 な刺激物 質による もの と思われ る。受粉 によ る胚珠の発達 には 植物の類縁関係が強 く作用す るので,
この刺激物 質は花粉の中に合 まれてい る一般的な植物 ホルモン で はないよ うに思われ る。4.胚
珠 の 発 達 と球 果 の 発 達 との 関 係 胚珠の発達 と球果の発達 とは密接な関係が ある。発達程度 の異 なる球果 を解剖 して調査 した ところ(表
3),無
受粉や 極 めて遠縁花粉の受 粉で は,胚
珠が胚の う発生の初期 に退化 し,雌
花 は球果 に発 達 しない。球果 に発 達 した ものは少な く とも胚の う細胞が大 きく生長 して遊離核督
零り膨纏鋒行
存
) 球果に発達 しなしヽ 果径4π″前後 球果 に発達す る。 果径7∼ 9 ππ 球果 に発達す る。 果径9∼ 10π″ 球果 に発達す る。 果径11∼1222 林 本 の交配 に関す る基 礎 的研究(Ⅷ ) 表3
胚珠の発達 と球果の発達 との関係Table 3 Relation bet、 veen ovule development and cOnc development
交 配 花 粉 球果 の発達 アスナロ (生 存), テガシヮ (生 存), (生存) ヒノキ (死 滅) ヒノキ (生 存) ノ ギ コ ス 形 成期以降の状態 まで発育が進 んでお り
,ま
た受精期(6月
上旬 )を す ぎて も胚珠は枯死せず7月以 降 に退化す るよ うであ つた。一般 に胚の うが大 きく生長 して遊離核形 成後期 ない し胚乳体形 成期 まで 分化が進 めば,雌
花 は球果 に発達 し,受
精 と無関係 に種子が形成 され る。 もちろん未受精の胚珠 は不 稔種 子 とな る。 ヒノキの研果 の生長期 は 4月 上旬か ら6月 下旬 までで,受
精の前 に球果が急激 に生長す る。雌花が 球果 に発達す るためには,球
果の生長期 に胚珠が生存 して生育 を続 けてい ることが必要のよ うである。 多 くの植物で果 実の発育 は胚珠 あ るい は種 子で生産 され るオー キシンや ジベ レ リンな ど生長調節物質 によ って コン トロール されてい ることが知 られている。針葉樹の場合 も,胚
珠 の発達 と球果 の発達 と は密接 に関連 してお り,ま
た フル ー トンや ジベ レ リンの散布 によ って無受粉 の雌花が あ る程度 まで球 果 に発 達す るので,球
果 の生長 にはB企珠で生産 され るオーキ シンや ジベ レ リンな ど生長調節物質が重 要 な役割 を演 じてい ることが推察 され る。 しか し,速
縁花粉の交配 で結果 しない現象 はオーキシンや ジベ レ リンな ど一般的な植物ホルモ ンのみでは説明で きない複雑な ものが ある。5,考
察 針葉樹 の中 には受粉 しなければ結果 しない もの と,無
受粉で も結果す るもの とが ある。 ヒノキの雌 花 は花粉 の生死 に関係な く受粉 しなければ球果 に発達 しない。 ヒノキは刺 激的(他動的)単
為結果 を す る植 物で ある。 スギで は ')受 粉 しな ければ球果 が発達 しない もの と,無
受粉 で も球果が大 き くな る もの とが あ る。大部分 は後者の栄養 的 (自 動 的)単
為結果 をす るが,無
受粉の球果 は一般 に小 さく,また種子の発育も悪い。アカマツ
,
クロマツでは
P他
家受粉よりも無受粉で落果が多く
,無
受粉の球
果は自然受粉の球果に比べて小 さい。また無受粉でタネがで きるものと,で
きない ものとあるが,タ
ネができて もその量は極めてわずかであるとい う。すなわち,針
葉樹は一般に受粉 しなければ結果・ 結実が不良の場合が多い。 ヒノキでは受粉 しなければ胚珠が早期に退化して結果 しない。またクロマツなどでは無受粉のとき に胚珠が開花後早い時点で崩壊 して落果することが多い という。 これ らのことか ら,針
葉樹では受粉 が胚珠の発達を刺激 して球果の発育 を促す と考えられる。すなわち,花
粉の中には特殊な刺激物質が 胚 珠 の 発 達 胚 珠は胚の う発生の初期に退化枯 死 し,種子が形成 されない。 胚珠の発達は遊離核形成後期∼胚 説で粍馨椒F柔
愁蓬キ壱懇落!罷
胚珠は正常に発達す るが,受精 し ない。不稔種子 となる。 胚珠は正常 に発達 し,受精す る。 充実種子が えられ る。隼 橋 合 まれていて