Title マウスの肝臓におけるコネキシン蛋白の発現と肝障害における細胞間コミュニケーションに関する研究( 本文(Fulltext) ) Author(s) 五十嵐, 功 Report No.(Doctoral Degree) 博士(獣医学) 乙第137号 Issue Date 2015-03-13 Type 博士論文 Version ETD URL http://hdl.handle.net/20.500.12099/50996 ※この資料の著作権は、各資料の著者・学協会・出版社等に帰属します。
マウスの肝臓におけるコネキシン蛋白の発現と肝障害
における細胞間コミュニケーションに関する研究
2014 年
岐阜大学大学院連合獣医学研究科
1 目次 略語 .………. 5 緒論 .………. 7 Figure 1 Figure 2 第1 章 健常マウスの肝臓におけるコネキシン (Cx26 および Cx32) の免疫 染色法の確立と加齢変化・性差の検討 1. 小序 ……….11 2. 材料および方法 2-1 使用動物 2-2 使用抗体および試薬 2-3 マイクロウェーブあるいは蛋白分解酵素による賦活化処理 2-4 LSAB 法による染色性 2-5 CSA 法による染色性 2-6 蛍光抗体法による検出 2-7 肝 Cx26 および Cx32 の局在に対する加齢および性差の影響 2-8 統計解析 3. 結果 3-1 LASB 法による染色性 3-2 CSA 法による染色性 3-3 蛍光抗体法による検出 3-4 肝 Cx26 および Cx32 の局在に対する加齢および性差の影響 4. 考察
2 5. 小括 Tables 1-4 Figures 3-4 第2 章 マウスの肝増殖性病変におけるコネキシン(Cx26 および Cx32)の局在 1. 小序 .……….26 2. 材料および方法 2-1 使用動物 2-2 肝増殖性病変の病理学的検査 2-3 肝増殖性病変の Cx26 および Cx32 の免疫染色と画像解析 2-4 統計解析 3. 結果 3-1 肝増殖性病変の病理学的検査 (H&E 染色) 3-2 肝増殖性病変の Cx26 および Cx32 の免疫染色と画像解析 4. 考察 5. 小括 Tables 5-6 Figures 5-7 第3 章 Cx32 ノックアウト(Cx32KO)マウスの肝臓の細胞間コミュニケー ション (GJIC) と増殖性病変 1. 小序 .……….37 2. 材料および方法 2-1 使用動物 2-2 使用試薬
3 2-3 肝臓の Cx26 および Cx32 の免疫染色 2-4 肝臓の IL/DT assay 2-5 自然発生の増殖性病変の病理学的検査 2-6 統計解析 3. 結果 3-1 肝臓の Cx26 および Cx32 の局在 3-2 肝臓の IL/DT assay 3-3 自然発生の増殖性病変 4. 考察 5. 小括 Table 7 Figures 8-11 第4 章 Cx32KO マウスにおける APAP 肝障害性 1. 小序 ……….47 2. 材料および方法 2-1 使用動物 2-2 使用試薬 2-3 血漿肝細胞逸脱酵素,肝薬物代謝酵素活性と肝 GSH+GSSG 含量 2-4 APAP による肝障害性 2-5 統計解析 3. 結果 3-1 血漿肝細胞逸脱酵素,肝薬物代謝酵素活性と肝 GSH+GSSG 含量 3-2 APAP による肝障害性 4. 考察
4 5. 小括 Tables 8-13 Figures 12-16 総括 ………65 謝辞 ……….67 引用文献 .………68
5 略語
ALT alanine aminotransferase アラニンアミノトランスフェラーゼ ALP alkaline phosphatase アルカリホスファターゼ
APAP acetaminophen アセトアミノフェン
AST aspartate aminotransferase アスパラギン酸アミノトランスフェ
ラーゼ CLZH chlorzoxazone 6-hydroxylation クロルゾキサゾン6-ヒドロキシレー ション Cx connexin コネキシン Cx26KO Cx26 knockout Cx26 ノックアウト Cx32KO Cx32 knockout Cx32 ノックアウト CYP cytochrome P450 チトクロムP450 DEN diethylnitrosamine ジエチルニトロソアミン
ECD 7-ethoxycoumarin O- 7-エトキシクマリン O-デエチラーゼ deethylase
FITC fluoresceinisothiocyanate フルオレッセインイソチオシア ネート
GJ gap junction ギャップジャンクション(細胞間) GJIC gap junctional intercellular 細胞間コミュニケーション
communication
GSH glutathione グルタチオン(還元型)
GSSG glutathione-S-S-glutathione グルタチオン(酸化型)
GST glutathione S-transferase グルタチオンS-トランスフェラーゼ H&E hematoxylin-eosin ヘマトキシリン-エオジン
6
HRP horseradish peroxidase 西洋ワサビパーオキシダーゼ IL/DT incision loading/dye transfer 色素移動 (切開,色素添加) LY Lucifer Yellow ルシファーイエロー MCD 7-methoxycoumarin O- 7-メトキシクマリン O- demethylase デメチラーゼ NAPQI N-acetyl-p-benzoquinoneimine N-アセチル-p-ベンゾキノンイミン PB phenobarbital フェノバルビタール PCD 7-propoxycoumarin O- 7-プロポキシクマリン O- depropylase デプロピラーゼ
T.BIL total bilirubin 総ビリルビン
UDP-GT UDP-glucuronosyltransferase UDP-グルクロノシルトランスフェラ ーゼ
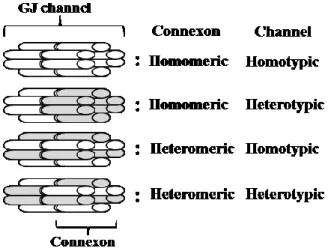
7 緒論 哺乳類の各種組織の細胞膜に存在するギャップ結合 (gap junction; GJ) は,隣接する 細胞同士の接着に関わるとともに細胞間の情報伝達機能を担っている (Fig. 1)。GJ の 分子構造は,コネキシン (connexin; Cx) と呼ばれる膜貫通型蛋白の六量体 (connexon; コネクソン) で構成され,細胞同士の細胞膜でコネクソンが対を作ることで接着し, 2 つの細胞を結びつけている。コネクソンの中心はトンネル構造となっており,無機 イオン (Na+やCa2+) や分子量 1,000 以下の小分子 (cAMP,inositol triphosphate,cGMP, glucose,ATP,ascorbic acid あるいは GSH) が, 隣接する細胞質へと通過できる仕組 みとなっている。また,コネクソンは,チャンネル構造を有し,細胞質内の Ca2+ や pH の状態によって,開閉が調節されている。このような GJ を介した情報伝達は, 細胞間コミュニケーション (gap junctional intercellular communication; GJIC) と呼ばれ, 生体のホメオスタシスを維持する上で極めて重要な役割を果たしている (15, 24, 50, 68)。 GJ を構成する Cx は,組織あるいは細胞によって異なり,分子レベルでの解析か ら多くの分子種が同定されている。例えば,肝臓では,Cx26 および Cx32,腎臓で は,Cx30.3,Cx31,Cx32 および Cx46,心臓では,Cx37,Cx43,Cx46 および Cx50, 肺では,Cx26,Cx37 および Cx46,脳では,Cx30,Cx32 および Cx43,皮膚では,Cx26, Cx30,Cx30.3,Cx31 および Cx43 が,ホモマーまたはヘテロマーとして存在し,コネ クソンを構成している (35, 50)。ホモマーとは,1 つの同じ Cx によってコネクソン が形成され,ヘテロマーとは,2 つ以上の異なった Cx によってコネクソンが形成さ れることである。細胞間では,ホモマーとホモマー,ホモマーとヘテロマーおよびヘ テロマーとヘテロマー (同じ Cx の結合あるいは異なった Cx の結合) の組み合わ せによって,GJ のチャンネルが構築されている (Fig. 2)。 Cx の減少・過剰発現や変異は,GJIC の正常機能からの逸脱につながり,様々な病 気の発生と関係が深いと考えられている。ヒトの疾患では,先天性の Cx32 遺伝子の
8
変異による X-linked Charcot-Marie-Tooth disease があり,主に脱髄をともなう神経障 害をきたすと報告されている (9)。また,他疾患として,Cx26 の変異による皮膚疾患 や難聴,Cx43 の変異による心筋障害,Cx46 および Cx50 の変異による白内障あるい はGJIC の機能異常が促進する腫瘍発現のような種々の障害が知られている (53, 60)。 したがって,これらの病態解析に関する研究成果の蓄積により,Cx あるいは GJIC の 制御によって起こる疾病のメカニズムが明らかにされつつある (53, 60)。また,GJIC は,細胞障害に関わる毒性情報を隣接する細胞に伝えることから,放射線やアポトー シス刺激による細胞ダメージ情報が GJIC を介して隣接する細胞に伝播することも 報告されている (3, 20, 37)。 肝臓における Cx26 および Cx32 の異常は,肝腫瘍の発現と進展と密接に関わり, GJ 構築不全に起因する GJIC の低下がプロモーションあるいはプログレッション作 用を引き起こすと考えられている。例えば,mRNA や蛋白レベルの解析から, プロモ ーター作用を有するフェノバルビタール (PB) は,マウスあるいはラットの肝臓の Cx32 を減少させ GJ 構築異常を誘発すると考えられている (34, 69)。一方,ジエチ ルニトロソアミン (DEN) に代表される種々の発がん物質によるゲッ歯類の肝臓前が んあるいは腫瘍病変では,Cx26 および Cx32 が減少し,腫瘍細胞同士あるいは正常 細胞との間で GJIC が阻害されるが (49),これら腫瘍においては Cx の遺伝子変異 が原因となることは稀であり,GJ 構築異常によって引き起こされるとの考えも示さ れている (73)。 近年,Cx32 の遺伝子をノックアウトした Cx32KO マウス (48) や Cx32 のドミナ ントネガティブ遺伝子を導入した Cx32Tg ラット (2) が作出され,GJIC の欠損ま たは抑制された遺伝子改変動物の利用が可能となってきた。Cx32KO マウスは,ネオ マイシン耐性遺伝子を Cx32 遺伝子のエクソン 2 内に導入することによって Cx32 遺伝子を不活性化し,Cx32 の発現とともに GJIC 機能が欠損している。一方, Cx32Tg ラットでは,Cx32 の発現が認められるものの,Cx32 変異蛋白の発現によ
9 る GJ 構築異常が起こり,GJIC が抑制される。しかし,DEN による肝腫瘍発現に対 しては,両モデル動物とも GJIC の抑制によって高感受性を示すことが確認されてい る (16, 26, 62,)。このように,遺伝子改変動物を用いた実験から,Cx32 は,腫瘍の発 現と進展に抑制的に働くことが明らかであり,がん抑制遺伝子の一つとして認識され ている (11, 65)。興味深いことに,Cx32 とともに肝臓で高発現している Cx26 の遺伝 子をノックアウトしたCx26KO マウスでは,GJIC 機能に異常がみられないことから, GJIC には,Cx32 が必須蛋白であることが示されている (38)。また,Cx26KO マウ スでは,DEN 投与後の腫瘍増加は認められていないことから (38),Cx26 は,肝腫 瘍の進展においては,Cx32 とは異なる作用を持つと考えられている (38, 40, 58)。 GJIC と腫瘍との関連性についての従来までの一連の研究では,化学物質による肝 腫瘍を対象にした実験が多い (16, 26, 34, 38, 40, 45, 49, 62, 69)。一般的に,長期飼育し たゲッ歯類での自然発生病変は,主として腫瘍であり,肝腫瘍の発現は,加齢ととも に上昇し,雌よりも雄の方が高いことがよく知られている (61, 46)。上述したごとく, Cx32 は,化学物質による発がん過程において抑制作用を示すことから,自然発生腫 瘍病変の発現にも関与する可能性が考えられた。しかし,これまで,マウス肝臓にお ける Cx26 および Cx32 の局在の加齢変化あるいは性差についての検討はなく,Cx の自然発生病変への関連性を調べる上で,精査が必要と考えられた。 そこで,本研究では,マウスの肝臓における Cx 蛋白の発現と肝障害における GJIC の関連性について以下4 章にわたり論述する。 第1 章では,健常マウスの肝組織内 Cx26 および Cx32 の局在を定型的な病理組織 学的検査で汎用されるホルマリン固定したパラフィン切片を利用するために,熱処理 あるいは蛋白分解酵素処理を施し, 免疫組織化学的染色が可能か検討した。その後, 肝組織内Cx26 および Cx32 の局在の加齢変化と性差について調べた。 第2 章では,マウスに自然発生する肝増殖性病変における肝組織内 Cx26 および Cx32 の局在を明らかにするために,免疫学的および病理組織学的手法を用いて検討
10 した。 第3 章では,長期飼育した Cx32 ノックアウト (Cx32KO) マウスに発症した肝増殖 性病変とGJIC の阻害の関連を明らかにするために,免疫学的および病理組織学的手 法を用いて解析した。 第4 章では,肝障害時における Cx32 の役割を明らかにするために,Cx32KO マウ スにアセトアミノフェン (APAP) を投与し,臨床病理学的手法を用いて検討した。 以上の検討から,マウスの肝臓でのCx の局在と肝障害への関与について明らかに し,論述を加えることを試みた。
Fig. 1 Gap junction (GJ) structure and its intercellular communication (GJIC)
11 第1 章 健常マウスの肝臓におけるコネキシン (Cx26 および Cx32) の免疫染色法の 確立と加齢変化・性差の検討 1. 小序 細胞間コミュニケーション (GJIC) に関する研究は,これまで化学物質により誘発 された肝臓腫瘍病変おいてコネキシン (Cx) の局在を確認することで解析が行われ てきた (34, 49)。このため,組織内 Cx の局在を確認するためには,新鮮材料を用い, 冷アセトンで固定後,凍結切片を作成し,高感度検出ができる蛍光抗体法を用いて免 疫組織化学的 (免疫染色) 検索を行う必要があった (49, 69)。しかし,定型的な組織 形態の観察に用いるホルマリン固定したパラフィン切片では,ホルムアルデヒドが組 織蛋白と結合し架橋を形成するため,免疫染色において,抗原抗体反応が阻害され, 染色性が低下していた (59)。したがって,Cx のように微量な抗原の検出には,ホル マリン固定したパラフィン切片が用いられてこなかった。 近年,この問題を解決するために,マイクロウェーブによる熱処理あるいは蛋白分 解酵素処理によって架橋構造を破壊し,組織内抗原性を賦活化する処理法が注目され ている (59)。例えば,マイクロウェーブについては増殖性細胞のマーカーである proliferating cell nuclear antigen (PCNA),蛋白分解酵素については上皮細胞に発現する cytokeratin や間葉系細胞に発現する vimentin の抗原検出に利用されている (7, 14)。
そこで,これらの手法を応用し,ホルマリン固定したパラフィン切片をマイクロウ ェーブあるいは蛋白分解酵素処理し,使用できないか検討した。Cx の検出には感度 の異なる 2 種類の免疫染色方法として labeled streptavidin (LSAB) あるいは catalyzed signal amplification (CSA) 法を用い,染色性・利便性を評価した。
LSAB 法は,抗原の Cx に対する一次抗体を反応させた後,ビオチン標識二次抗体, 引き続きHRP 標識ストレプトアビジンを反応させ HRP 染色を行なう手法である (Fig. 3)。一方,CSA 法は,LSAB 法と同様にビオチン標識二次抗体,アビジン・ビオチン・
12 HRP 複合物とビオチン標識タイラマイドを反応させ,さらに HRP 標識ストレプトア ビジンにより,多くのHRP を結合させ染色強度を増強する方法である (Fig. 3)。両法 を比較した場合,CSA 法が LSAB 法より検出感度が高いことが知られている。対照と しては,従来から行われている抗原性賦活化処理を必要とせず,検出感度が高いと考 えられている蛍光抗体法を用いた。 上記検討から,最適な染色法を用いて,13,52 および 104 週齢の雌雄 B6C3F1 マ ウス肝臓のCx 発現の加齢と性差について検討した。 2. 材料および方法 2-1 使用動物 実験に供したB6C3F1 マウスは,室温 23 ± 2 C, 湿度 55 ± 10%,照明時間 200 ル クス 13 時間/日,換気回数 10-15 回/時間で制御したバリアーシステムの動物室で飼 育し, basal diet(NMF: オリエンタル酵母工業, 東京)と水道水を自由摂取させた。動 物の取り扱いは,全て「動物の愛護及び管理に関する法律」 (1973 年 10 月 1 日法律 105 号,1999 年 12 月 22 日改正) に従って行い,第一三共株式会社(東京)の動物実 験倫理規定に従って実施した。なお,B6C3F1 マウスは,肝腫瘍病変の研究 (33, 54, 63, 66,) に多く用いられているため,本研究で選択した。 染色性の検討では,健康な4 週齢の 雄性 B6C3F1 マウス (n = 3) を日本エスエル シー (浜松) から購入し,13 週齢まで飼育して実験に供した。 加齢および性差の検討では,4 週齢の雌雄 B6C3F1 マウス (n = 15 ) を同様に日本 エスエルシーから購入し,長期間飼育した。実験には,13, 52 および 104 週齢 (n = 5) を用い,これらマウスをエーテル麻酔下で腹部大動脈を切断し,放血屠殺後,肝臓を 摘出した。10% 中性緩衝ホルマリン液で 3 日間固定後,外側左葉,内側左右葉,外 側右葉および尾状葉の4 部位から組織片を切り出し,定法に従い固定後 (エタノール, キシレン,パラフィン浸透による組織処理),パラフィンブロックを作製し,約 3 m
13 に薄切して,染色した。
2-2 使用抗体および試薬
染色に用いた抗ラット Cx26 ウサギ抗体および抗ラット Cx32 ウサギ抗体は, Zymed Laboratories Inc.(South San Francisco, CA, 米国) から購入し,マウスと cross reaction があることを確認後使用した。正常血清,ビオチン標識ウサギイムノグロブ リン,ビオチン標識タイラマイド,アビジン•ビオチン•HRP 複合物,HRP 標識スト レプトアビジン,FITC 標識ストレプトアビジン と非蛍光封入剤は市販品 (Dako Cytomation, 京都) を使用した。プロテアーゼおよび Tween 20 は,Sigma-Aldrich Japan (東京) から購入した。過酸化水素水,アセトンは,和光純薬 (大阪) から購入した。 また,3,3'-ジアミノベンチジン・四塩酸塩 (DAB) は,同仁化学研究所 (熊本) より購 入した。リン酸緩衝食塩液 (0.01 M,pH7.4;PBS) は,リン酸水素二ナトリウム,リ ン酸二水素カリウム,塩化カリウムおよび塩化ナトリウムを,トリス塩酸緩衝液 (0.05 M, pH7.6; TBS) は,トリスヒドロキシメチルアミノメタンおよび塩酸を,クエン酸緩 衝液 (0.01M,pH 6.0) は,クエン酸二水和物およびクエン酸ナトリウム二水和物をい ずれも和光純薬 (大阪)から購入して調製した。 2-3 マイクロウェーブあるいは蛋白分解酵素による賦活化処理 マイクロウェーブ処理には,蒸留水あるいはクエン酸緩衝液 (0.01M,pH 6.0) を処 理液とし,電子レンジ (MR-A330,日立,東京) で 500 W,10 分を 1 - 2 回処理する 条件を設定した。蛋白分解酵素処理には,0.2 % プロテアーゼ溶液を用い,室温で 10, 20,30,40,50 および 60 分間の処理条件を設けた。これらの処理は,一般的に応 用されており,各々の処理効果を確認するために,賦活化処理を行わない未処理群も 設けた。
14 2-4 LSAB 法による染色性 LSAB 法による染色手法を Fig. 3 に示した。脱パラフィンした組織切片を 3% 過酸 化水素水で 5 分処理した後,PBS (0.01 M,pH7.4) で洗浄した。その後,抗原性の賦 活化処理を上記の条件で行った。PBS で洗浄後,正常血清を室温で 5 分間処理した。 次に,抗ラットCx26 または Cx32 ウサギ抗体 (1,000 倍希釈) を室温で 60 分間反 応させ,PBS で洗浄後,ビオチン標識抗ウサギイムノグロブリン (400 倍希釈) を室 温で 60 分間インキュベートした。その後,PBS 洗浄を行い,HRP 標識ストレプト アビジンを室温で 30 分間反応した。PBS 洗浄後,DAB と過酸化水素の基質溶液で 3 分発色を行った。最後にヘマトキシリンで核染色を実施し,脱水,透徹後に封入した。 なお,各抗体反応では,染色条件を統一するために,自動免疫染色装置 (Ventana XT, Roche Diagnostics, 東京) を用いた。 Cx26 および Cx32 の染色後,光学顕微鏡を用いて各切片の染色性を確認した。染 色性の良否の判断は,従来実施されている蛍光抗体法で検出した結果と同様に,肝小 葉全体の肝細胞の細胞膜上にスポット状の陽性部位が観察されることに着目した。ま た,非特異反応の有無 (強い:++,弱い:+,認められない:- ),染色強度 (強い: ++,弱い:+,染まらない:- ),鑑別の容易さ (明瞭:○,不明瞭:×),抗原性賦活 化処理にともなう組織傷害の程度 (障害なし:◎,軽度:○,中等度:△) について も判断基準とした。 2-5 CSA 法による染色性 CSA 法による染色手法を Fig. 3 に示した。脱パラフィンした組織切片を 3% 過酸 化水素水で 5 分処理した後,0.1% Tween 20 と 0.3 M 塩化ナトリウムを含む TBS (0.05 M, pH7.6) で洗浄した。抗原性の賦活化処理を各条件で行った後,正常血清で室 温 5 分間処理し,抗ラット Cx26 または Cx32 ウサギ抗体 (1,000 倍希釈) を室温で 60 分間反応した。TBS 洗浄後,ビオチン標識抗ウサギイムノグロブリンを室温で 15
15 分間反応した。TBS 洗浄後,アビジン•ビオチン• HRP 複合物を室温で 15 分間反応 した。TBS 洗浄後,ビオチン標識タイラマイドを室温で 15 分反応した。TBS で洗 浄した後,HRP 標識ストレプトアビジンを室温で 15 分間インキュベートした。そ の後,TBS 洗浄を行い,DAB と過酸化水素の基質溶液で 10 - 20 秒発色を行った。 最後にMayer のヘマトキシリンで核染色を実施し,脱水,透徹後に封入した。なお, 各抗体反応では,染色条件を統一するために,自動免疫染色装置を用いた。染色性の 評価は,LSAB 法と同様に非特異反応の有無 (強い:++,弱い:+,認められない:- ), 染色強度 (強い:++,弱い:+,染まらない:- ),鑑別の容易さ (明瞭:○,不明瞭: ×),抗原性賦活化処理にともなう組織傷害の程度 (障害なし:◎,軽度:○,中等度: △) を判断基準とした。 2-6 蛍光抗体法による検出 摘出した肝臓 (内側右葉) のから 10 × 10 mm の組織片を切り出し,未固定のまま OCT コンパウンド (Sakura Fintek Japan Co., 東京) に包埋して,液体窒素を用いて凍 結ブロックとした。その後,クリオスタッド (Tissue-Tek Cryo3, Sakura Fintek Japan Co., 東京) を用いて,凍結ブロックから約 8 m の切片を作製した。切片を風乾させた後, -20 C に冷却したアセトンで 5 分間固定した。免疫組織化学は,FITC 標識ストレプ トアビジンを用いてLSAB 法で実施 (“2-4”) した。その後,PBS で洗浄を行い, 非蛍光封入剤で標本を封入した。Cx の陽性部位は,蛍光顕微鏡により観察した。 2-7 肝 Cx26 および Cx32 の局在に対する加齢および性差の影響 CSA 法 (0.2%プロテアーゼで 30 分間の抗原性賦活化処理を実施) により Cx26 および Cx32 の染色を行なった後,光学顕微鏡を用いて Cx の陽性部位を確認した。 さらに,肝小葉の中心部と辺縁部に分けてそれぞれ約 800 細胞/例における陽性部位 のスポット数,スポット面積および肝細胞の核数を病理画像解析装置 (IPAP-WIN, 住
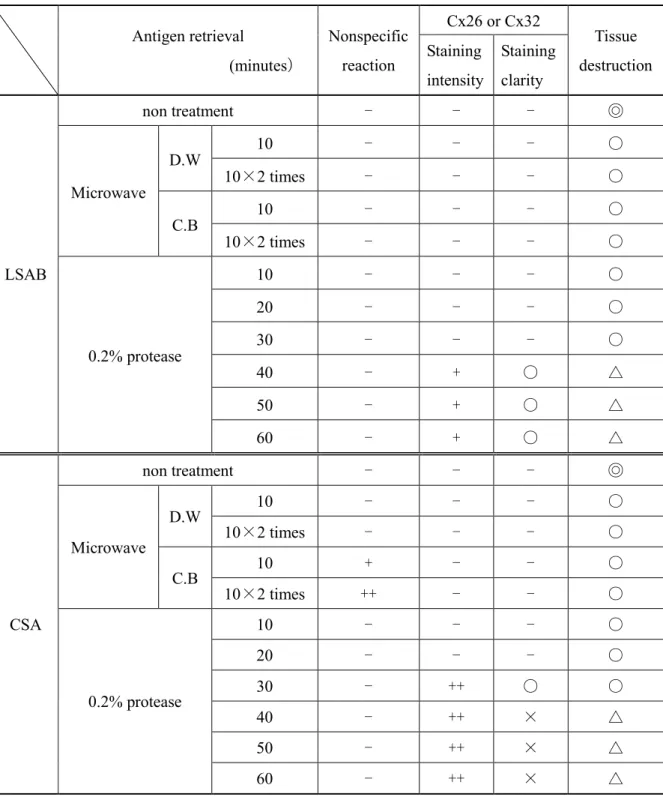
16 化テクノサービス,兵庫) を用いて計測した。それらの計測値から,肝細胞 1 個当 たりのスポット数,スポット総面積を算出し,加齢と性差の有無について計測値を比 較した。 2-8 統計解析 計測データは,群の平均値と標準偏差 (SD) として表し,統計的に Student’s t 検定 により分析した。統計解析における P 値が 5 % 未満となる場合に統計学的に有意で あると判断した。 3. 結果 3-1 LSAB 法による染色性 結果をTable 1 に示した。 Cx26 と Cx32 の染色性は,賦活化未処理,マイクロウェーブによる蒸留水またはク エン酸緩衝液での10 分を 1 回および 2 回,0.2%プロテアーゼによる 10 分~60 分処 理の全ての検討条件で同様であった。 マイクロウェーブ処理では,蒸留水 とクエン酸緩衝液で 10 分を 1 回または 2 回処 理しても,Cx は検出されなかった。また,非特異反応も認められなかった。 0.2% プロテアーゼ処理では,10, 20 および 30 分処理では,賦活化未処理と同様 に陽性部位は全く確認されなかったが,40,50 および 60 分の処理では,肝細胞の細 胞膜上にスポット状に染まる陽性部位が出現した。しかし,それらの陽性部位は,極 めて微小で,Cx の局在を鑑別するのは困難であり,肝細胞に軽度の組織破壊も認め られた。したがって,LSAB 法による染色では,全ての検討条件で良好な結果は得ら れなかった。 3-2 CSA 法による染色性
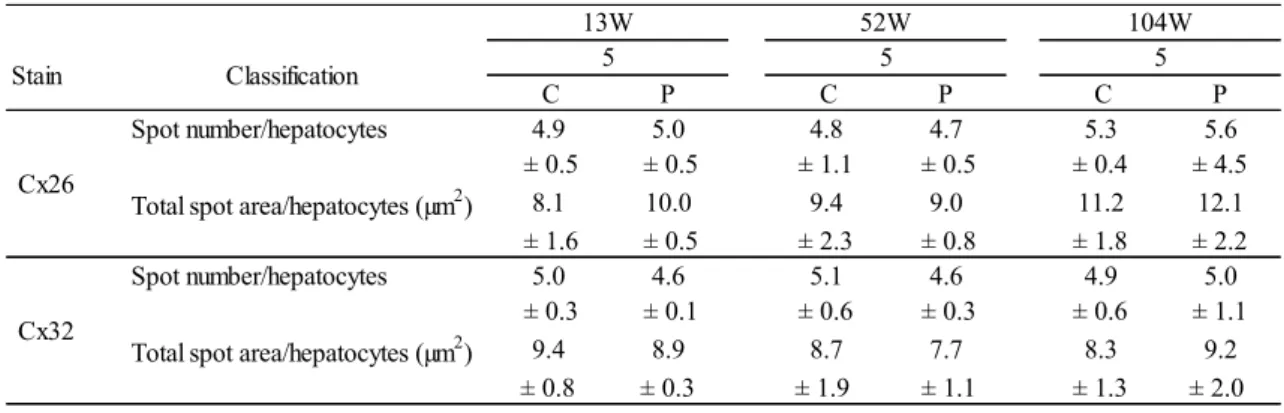
17 結果をTable 1 に示した。 Cx26 と Cx32 の染色性は,LSAB 法と同様に賦活化未処理,マイクロウェーブによ る蒸留水またはクエン酸緩衝液での10 分を 1 回および 2 回,0.2% プロテアーゼによ る10 分~60 分処理の全ての検討条件において差はなかった。 マイクロウェーブあるいは0.2% プロテアーゼ処理のどちらも行わない賦活化未処 理では,Cx の陽性部位は全く認められなかった。一方,マイクロウェーブ処理では, 蒸留水とクエン酸緩衝液を用いたいずれの処理条件でも陽性部位は検出されず,クエ ン酸緩衝液においては肝細胞に非特異的な反応が認められた。プロテアーゼ処理につ いては,10 および 20 分処理では,陽性部位を確認することはできなかった。しか し,30~60 分処理した切片では,肝細胞の細胞膜上に陽性スポットが確認され,い ずれも LSAB 法で検出したものより強い染色強度を示した。40 分間以上処理した条 件では,組織破壊が認められるとともに,陽性スポットが不明瞭になる傾向があり, 30 分間処理したものが最も良好であった。30 分処理の染色結果は,蛍光抗体法で実 施したものと同様に Cx の局在が観察し易かった (Fig. 4)。 3-3 蛍光抗体法による検出 肝細胞の細胞膜上にCx26 または Cx32 の局在を示す陽性スポットが確認された (Fig. 4)。また,それらの陽性部位は,蛍光発色により明瞭であった。 3-4 肝 Cx26 および Cx32 の局在に対する加齢および性差の影響 各週齢におけるCx26 および Cx32 の肝細胞 1 個当たりの陽性スポット数および 総面積の計測の雄の結果をTable 2 に,雌の結果を Table 3 に示した。 13,52 および 104 週齢の雌雄マウスにおける Cx26 と Cx32 の陽性スポットは, 肝細胞の細胞膜上で全て認められ,数および総面積に加齢変化は観察されなかった。 また,性差も確認されなかった。
18 4. 考察 凍結切片を用いる蛍光抗体法は,検出操作が比較的容易で検出感度に優ているため, これまで繁用されてきたが,暗視野で観察するために陽性部位と周囲の組織形態との 位置関係の検索が難しい。これに対してホルマリン固定・パラフィン切片による酵素 抗体法は,明視野での観察により組織形態の観察が容易であり,さらに,病理組織学 的検査で利用するH&E 染色標本や他染色と同一のパラフィンブロックまたは隣接切 片を用いて評価できる利点がある。本検討では,健常マウスの肝組織内Cx26 および Cx32 の局在を定型的ホルマリン固定・パラフィン切片が利用できないか,熱処理あ るいは蛋白分解酵素処理を施し免疫組織化学的染色で可能性を検討した。 ホルマリン固定・パラフィン切片では 既報のとおり,Cx の抗原性が消失あるいは 減弱するため,発現量の少ないCx を検出できなかった。しかし,0.2%プロテアーゼ 処理 を 10~60 分の条件で実施した場合には,LSAB 法では 40~60 分,CSA 法では 30~60 分で抗原性の賦活化効果が確認され,肝細胞の細胞膜上に Cx の陽性スポッ トが出現した。これに対して,マイクロウェーブ処理では 蒸留水およびそれ以上の 賦活化効果が得られるクエン酸緩衝液を用い,10 分を 1 回または 2 回処理しても Cx の陽性部位は検出されず,むしろ非特異反応が確認されるのみであった。 したがって,プロテアーゼによる抗原性の賦活化処理とLSAB 法あるいは CSA 法 を組み合わせれば,ホルマリン固定・パラフィン切片でもCx26 および Cx32 蛋白を 検出可能であると考えられた。 そこで,LSAB 法と CSA 法の検出力を,プロテアーゼ処理切片を用いて陽性部位 で比較すると,LSAB 法では染色強度が弱く Cx の局在を鑑別するのが困難であった。 一方,CSA 法で検出した Cx の局在は極めて明瞭であった。なお,プロテアーゼ処 理は,処理時間の延長に伴ってCx の陽性スポットが現れたが,同時に組織破壊が強 く認められることから, 30 分処理が至適であり,蛍光抗体法と同様に陽性部位を検 索し易かった。
19 以上, ホルマリン固定・パラフィン切片において, 0.2 %のプロテアーゼで 30 分 の抗原性賦活化処理を実施しCSA 法で免疫染色を施せば,蛍光抗体法とは遜色のな い染色性が得られ,Cx26 および Cx32 の組織内局在を容易に検索できた (Table 4)。 次に,これら至適手法を用いて,肝Cx の局在に対する加齢および性差の影響を調 べたところ,13,52 および 104 週齢の雌雄マウスとも,Cx26 および Cx32 の肝細胞 1 個当たりの陽性スポット数と総面積および染色性に差異は見られなかった。既報に よると,SD ラットでは,肝 Cx26 の局在に加齢による影響は認められないが,Cx32 は加齢とともに減少し,100 週齢では GJIC の機能が約 60%に低下することが報告 されている (45)。これに関連して,85 週齢のラットに肝発がん性物質であるジエチ ルニトロソアミン (DEN) を腹腔内投与すると,細胞変異巣が 10 および 30 週齢に投 与したラットより増加することから,Cx32 の減少により肝腫瘍プロモーションある いはプログレッション作用が増強されるとの指摘がある (45)。性差に関しては,ラッ トにヘキサクロロベンゼンを経口投与すると,雄よりも雌で肝 Cx32 の減少が強く認 められると報告されている (55, 56)。これらの実験では 各肝毒性物質の有する作用機 作の違いがあるため,今後精査が必要であるが,Cx32 プロモーター遺伝子の応答や 発現に, マウスとラット間では加齢あるいは性差に関して差異があるかもしれない。 以上,B6C3F1 マウスでは,Cx26 および Cx32 の発現に加齢や性差は認められず, Cx32 および GJIC の減少に起因する肝腫瘍プロモーションあるいはプログレッショ ン作用がないと推察された。 5. 小括 肝Cx26 および Cx32 の局在を免疫組織化学的に検索するために,病理組織学的検 査で汎用されるホルマリン固定したパラフィン切片が利用できないか検討した。その 結果,0.2 %のプロテアーゼで 30 分の抗原性賦活化処理を実施した後,CSA 法を採
20
用することで,良好な染色結果が得られることが明らかとなった。確立した手法と104 週齢までのB6C3F1 マウスを用いて,Cx26 と Cx32 の局在を加齢および性差の点から 調べたが,影響はみられなかった。
21
Fig. 3 Immunochemical procedure for Cxs stains by labeled streptavidin (LASB) and catalyzed signal amplification (CSA) methods
22
Table 1. Results of Immunohistochemical staining of Cx26 and Cx32 in the liver of male B6C3F1 mice using paraffin sections
Antigen retrieval (minutes) Nonspecific reaction Cx26 or Cx32 Tissue destruction Staining intensity Staining clarity LSAB non treatment - - - ◎ Microwave D.W 10 - - - ○ 10×2 times - - - ○ C.B 10 - - - ○ 10×2 times - - - ○ 0.2% protease 10 - - - ○ 20 - - - ○ 30 - - - ○ 40 - + ○ △ 50 - + ○ △ 60 - + ○ △ CSA non treatment - - - ◎ Microwave D.W 10 - - - ○ 10×2 times - - - ○ C.B 10 + - - ○ 10×2 times ++ - - ○ 0.2% protease 10 - - - ○ 20 - - - ○ 30 - ++ ○ ○ 40 - ++ × △ 50 - ++ × △ 60 - ++ × △
Note: LSAB: Labeled streptavidin biotin method, CSA: Catalyzed signal amplification method. Microwave with: D.W, distilled water: C.B, citrate buffer (0.01M, pH6.0). Cx32 or Cx26: The Cx26 or 32 positive gap junctions appear between neighboring hepatocytes. Staining intensity: ++, strong; +, weak; -, negative. Staining clarity: ○, clarity; ×, not clarity. Nonspecific reaction appear in cytoplasmic of hepatocytes: ++,strong; +,weak; -, negative. Tissue destruction: ◎, no; ○, mild; △, moderate.
23
Fig. 4 Fluorescent and immunohistochemical staining of Cx26 and Cx32 in the liver of male B6C3F1 mice. Note numerous positive spots for Cx26 (A and C) and Cx32 (B and D) on cell membranes. A and B, fluorescent antibody method; B and D, immunoenzymatic method.Original magnification: 120×.
A
24
Table 2. Image analysis for Cx26 and Cx32 stainings of male B6C3F1 mice with ages
C P C P C P
Spot number/hepatocytes 4.9 5.0 4.8 4.7 5.3 5.6 ± 0.5 ± 0.5 ± 1.1 ± 0.5 ± 0.4 ± 4.5 Total spot area/hepatocytes (µm2) 8.1 10.0 9.4 9.0 11.2 12.1 ± 1.6 ± 0.5 ± 2.3 ± 0.8 ± 1.8 ± 2.2 Spot number/hepatocytes 5.0 4.6 5.1 4.6 4.9 5.0
± 0.3 ± 0.1 ± 0.6 ± 0.3 ± 0.6 ± 1.1 Total spot area/hepatocytes (µm2) 9.4 8.9 8.7 7.7 8.3 9.2
± 0.8 ± 0.3 ± 1.9 ± 1.1 ± 1.3 ± 2.0
13W 52W 104W
Stain Classification 5 5 5
Cx26
Cx32
Each value represents the mean ± SD of the respective groups. C: central zone of liver lobule, P: peripheral zone of liver lobule.
Table 3. Image analysis for Cx26 and Cx32 stainings of female B6C3F1 mice with ages
C P C P C P
Spot number/hepatocytes 4.4 4.4 4.68 4.71 4.5 4.2 ± 0.3 ± 0.3 ± 0.2 ± 0.6 ± 0.4 ± 0.5 Total spot area/hepatocytes (µm2) 8.1 8.3 10.4 10.6 8.5 8.6
± 1.0 ± 1.1 ± 1.4 ± 1.6 ± 1.5 ± 1.9 Spot number/hepatocytes 4.2 3.8 4.5 4.4 4.8 4.5
± 0.5 ± 0.6 ± 0.5 ± 0.4 ± 0.3 ± 0.2 Total spot area/hepatocytes (µm2) 8.5 7.7 8.2 8.3 8.9 7.7
± 1.9 ± 2.7 ± 2.3 ± 2.4 ± 0.6 ± 0.3 5 Cx26 Cx32 13W 52W 104W Stain Classification 5 5
Each value represents the mean ± SD of the respective groups. C: central zone of liver lobule, P: peripheral zone of liver lobule.
25
Table 4. Immunohistochemical procedure for Cx26 or Cx32 in the liver using fluorescent antibody or immunoenzymatic method of male B6C3F1 mice
Fluorescent antibody method Immunoenzymatic method
1.Cryosection, 8 μm 1.Fixation (phosphate-buffered 10% formarin) 2.Air dry, 1h 2.Paraffin section, 3 μm
3.Fixation (acetone), -20℃,5 min 3.Deparaffinization
4.Air dry, 30 min 4.3% hydrogen peroxide solution, 5 min 5.PBS(pH7.4), 5 min 5.TBS(pH7.6), 5 min×3
6.Rabbit anti-rat Cx26 or Cx32 antibody, 1 h 6.0.2% protease solution, 30 min
7.PBS, 5 min×3 7.TBS, 5 min×3
8.Rabbit Immunoglobulin/Biotinylated, 1 h ⑧.Protein block, 5 min
9.PBS, 5 min×2 9.Rabbit anti-rat Cx26 or Cx32 antibody, 1h
10.PBS, 5 min 10.TBS, 5 min×3
11.Streptavidin/FITC, 30 min 11.Rabbit immunoglobulin/Biotinylated, 15 min
12.PBS, 5 min×2 12.TBS, 5 min×2
13.Mounting (fluorescence mounting medium) ⑬.Streptavidin-biotin-HRP complex, 15 min 14.TBS, 5 min×2
⑮.Tyramide/biotinylated, 15 min 16.TBS, 5 min×2
⑰.Streptavidin/HRP, 15 min 18.TBS, 5 min×3
19.DAB solution, 10-20 sec 20.Water, 5 min
21. Hematoxylin solution, 5 min 22.Water, 15 min
23.Mounting (water-free mounting medium) ○:Catalyzed Signal Amplification System (Dako Cytomation)
26 第2章 マウスの肝増殖性病変におけるコネキシン(Cx26 および Cx32)の局在 1. 小序 第1 章において,健康な B6C3F1 マウスの肝 Cx26 および Cx32 の局在は,加齢およ び性差がないことを明らかにしたが,マウスでの自然発生の肝増殖性病変における Cx に関する報告は少ない。 これまで肝増殖性病変における Cx の局在は,前がん病変である肝細胞変異巣 (altered cell foci) と腫瘍病変 (tumor) の簡易的な分類で評価されることが多かった (42, 49)。しかし,これら病変では,Cx26 および Cx32 の局在異常または消失が報告さ れているものの (49),簡易的な分類であるため病変のタイプによる Cx の局在や病変 の進展については検討されていない。一方,一般的な組織学的診断では,その形態学 的特徴から,肝細胞変異巣はclear cell foci, eosinophilic foci と basophilic foci に,腫 瘍病変は adenoma と carcinoma に細かく分類 (5, 22) され,かつ増殖性病変への進展 が報告されている (23, 25)。
本章では,肝増殖性病変を有する雄性B6C3F1 マウスを取り上げ,病変のタイプと Cx26 と Cx32 の局在の関連性を明らかにするために,まず,ヘマトキシリン・エオジ ン (H&E) 染色により病変を観察し,5 種 (clear cell foci, eosinophilic cell foci, basophilic cell foci, adenoma および carcinoma) に分類後,第 1 章で確立した手法を用いて Cx の 免疫染色を施した。得られたデータを基にすると,腫瘍病変の adenoma および carcinoma において,H&E 染色における細胞質の染色性によって Cx の発現パターン が異なっていたことから,それぞれeosinophilic cytoplasm と basophilic cytoplasm の 2 種を加え,新たに7 種のタイプの形態学的変化で分類を行った。
さらに, Cx の細胞内局在を画像解析して,肝細胞変異巣および腫瘍病変と Cx の 発現との関連性を調べた。
27 2. 材料および方法 2-1 使用動物 動物の取り扱いは,全て「動物の愛護及び管理に関する法律」 (1973 年 10 月 1 日 法律105 号,1999 年 12 月 22 日改正) に従って行い,第一三共株式会社 (東京) の動 物実験倫理規定に従って実施した。 実験には,4 週齢の 雄性 B6C3F1 マウス (n = 250 ) を日本エスエルシー (浜松) か ら購入し,104 週齢まで飼育して屠殺した。麻酔・飼育条件は,第 1 章の“2-1”と 同様である。対照としては,非病変領域を用いた。 2-2 肝増殖性病変の病理学的検査 安楽死させた後,摘出した肝臓は,剖検所見をとり,10% 中性緩衝ホルマリンで 固定した。その後,外側左葉,内側左右葉,外側右葉および尾状葉の病変を含めて組 織片を切り出し,約 3 m のパラフィン切片を作製して H&E 染色を施し鏡検した。 そのうち,病変と診断された標本を検索材料とした。
肝細胞変異巣については,clear cell foci, eosinophilic cell foci および basophilic cell foci の 3 種類に分類し,各々 4 個の病変を観察対象とした。肝腫瘍については,adenoma と carcinoma に分類し,さらに,これら腫瘍を細胞質が好酸性を示す eosinophilic type と細胞質が塩基性を示す basophilic type に分類し,各々 8 個の病変を観察した。ま た,各病変の特徴 (41) を基に,盲検 (blind) 下で認定病理資格者 (日本毒性病理学会 毒性病理学専門家および日本獣医病理学専門家)が診断した。 2-3 肝増殖性病変の Cx26 および Cx32 の免疫染色と画像解析 H&E 染色標本を用いて確認された病理組織所見に基づき,生存例に認められた肝 細胞変異巣および肝腫瘍を検討対象とした。肝Cx26 および Cx32 の免疫染色は,各 病変を含むホルマリン固定・パラフィン切片を用い,第1 章“2-5”で確立した手法
28 に準じて行った。 次に,光顕的に Cx26 と Cx32 の陽性部位を確認し,各病変部および正常部位 (病 変周囲の非病変細胞) のにおける陽性部位のスポット数,スポット面積および肝細胞 の核数を病理画像解析装置 (IPAP-WIN, 住化テクノサービス株式会社,兵庫) を用い て計測した。計測領域は,肝細胞変異巣については病変全体,肝腫瘍については代表 部分とした。その後,各計測値から肝細胞 1 個当たりのスポット数,スポット総面 積を算出し,病変部と正常部を比較した。 2-4 統計解析 計測データは,平均値 ± 標準偏差 (SD) で表し,統計的に Student’s t 検定により 非病変領域および増殖性領域間について分析した。統計解析における P 値が 5 %未 満となる場合に統計学的に有意であると判断した。 3. 結果 3-1 肝増殖性病変の病理学的検査 (H&E 染色) 得られた所見を基に,下記のように基準を設定した。
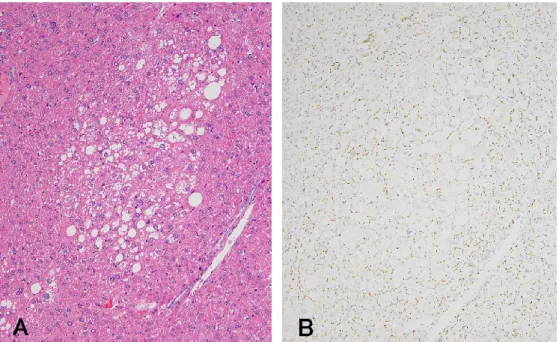
Clear cell foci:空胞を伴う正常または腫大した細胞質を有する細胞からなり,核は 小さく緻密で中央に位置する (Fig. 5)。
Eosinophilic cell foci:好酸性細胞質を有する大型の肝細胞から構成される。また, 核は腫大し,核小体が顕著で中央に位置する (Fig. 6)。
Basophilic cell foci:好塩基性の細胞質からなる比較的小型の細胞からなる。塊状あ るいは長い帯状に配列する (Fig. 7)。
Adenoma:軽度の異型が認められる肝細胞が不規則に増殖。増殖巣内では正常な小 葉構造が失われ,増殖巣周囲の既存の肝細胞では軽度の細胞の圧迫が認められる (Fig. 6, 7)。
29
Carcinoma:高度の異型が認められる肝細胞が周囲組織へ浸潤性に増殖し,増殖巣 周囲の細胞境界が不明確 (Fig. 6, 7)。
3-2 肝増殖性病変の Cx26 および Cx32 の免疫染色と画像解析
免疫染色所見をFig. 5-7 に,Cx の細胞内局在の画像解析結果を Table 5 に,特に adenoma および carcinoma における Cx の解析結果は Table 6 に示した。
Clear cell foci における Cx26 と Cx32 の陽性スポットは,正常部の肝細胞に比べ減 少傾向がみられたが (Fig. 5),画像解析では非病変領域に対して,有意差はみられな かった (Table 5)。
Eosinophilic cell foci における Cx26 の陽性スポットは,肝細胞間の陽性スポットが 減少傾向を示したが,明らかな変化ではなかった。しかし,類洞の細胞膜上に陽性ス ポットが確認された。画像解析では,有意な変化は認められなかった。Cx32 では, 肝細胞間の陽性スポットが減少し,類洞の肝細胞膜上に強い陽性像が多く認められた (Fig. 6)。画像解析では有意なスポット数およびスポット総面積の減少傾向がみられ, 染色結果と合致した結果を示した (Table 5)。
Basophilic cell foci における Cx26 では,正常な肝細胞に比べて陽性スポットの数が 多く,個々のスポット面積も大きい傾向がみられた。画像解析でも,有意なスポット 総面積に高値が認められ,染色結果と合致していた。Cx32 においても Cx26 と同様 の染色傾向が認められ (Fig. 7),画像解析では有意なスポット総面積の高値を示した (Table 5)。 Adenoma のうち細胞質が好酸性を示す腫瘍細胞 (eosinophilic タイプ) と細胞質が好 塩基性を示す腫瘍細胞 (basophilic タイプ) とでは,Cx26 と Cx32 の発現パターンが 異なっていた (Fig. 6, 7)。Eosinophilic タイプの腫瘍細胞では,肝細胞間のスポットが 減少し,類洞に沿った細胞膜上に陽性像が認められ,eosinophilic foci に類似した発現 パターンを示した (Fig. 6)。肝細胞当たりのスポット数およびスポット総面積の有意
30
な高値が認められ,その変化はeosinophilic foci に比べて明らかであった (Table 6)。 Basophilic タイプの腫瘍細胞では肝細胞間および類洞に沿った細胞膜上に陽性スポッ トが認められ,basophilic foci に類似した染色性を示したが,スポットはさらに大き く,強く染色される傾向が確認された (Fig. 7)。肝細胞当たりのスポット数およびス ポット総面積に有意な高値が認められた (Table 6)。
Carcinoma は,adenoma 同様に,eosinophilic タイプと basophilic タイプでは,Cx26 と Cx32 発現パターンに差異がみられた (Fig. 6, 7)。Eosinophilic タイプでは,肝細胞 間の陽性スポットが減少し,類洞に沿った肝細胞の細胞膜上に陽性像が認められた (Fig. 6)。肝細胞当たりのスポット数およびスポット総面積に有意な高値が認められた (Table 6)。 Basophilic タイプでは,大型のスポットが細胞膜上に均一に増加している 像が認められ (Fig. 7),肝細胞当たりのスポット数およびスポット総面積の有意な高 値が認められた(Table 6)。 4. 考察 これまで,肝増殖性病変における Cx の局在異常については,ラットに発癌物質を 投与して,発生する腫瘍で報告されている (47, 49)。例えば,DEN と PB の併用投 与によって,Cx32 の肝細胞間における減少と類洞の細胞膜での強い発現が認められ ている (49)。 本実験において,104 週齢の雄性 B6C3F1 マウスに自然発生した肝増殖性病変での Cx26 と Cx32 の局在は,病変のタイプによって異なっていた。すなわち,clear cell foci でみられた発現が減少するパターン,eosinophilic foci と細胞質が好酸性を示す
adenoma および carcinoma でみられた肝細胞間の発現が減少し,類洞に沿って細胞膜 に線状に強く発現するパターン (eosinophilic タイプ),basophilic foci と細胞質が好塩 基性を示すadenoma および carcinoma でみられた肝細胞間と類洞の細胞膜に発現して スポット状に大きさや数を増すパターン (basophilic タイプ) である。
31 一般的に細胞変異巣は前がん病変と考えられ,化学物質の発がん性評価の指標とし てよく用いられているが,細胞変異巣からadenoma, さらに carcinoma へと進展すると 考えられている (25)。本検討において確認された Cx の局在パターンは,肝細胞変異 巣からadenoma や carcinoma に病変が進展しても同様な特徴を有していることが明ら かになった。また,肝増殖性病変のうち,eosinophilic タイプと basophilic タイプで Cx32 の局在パターンが異なっていたことは,この局在パターンにより肝増殖性病変が分 類・診断できる可能性を示唆している。なお,Clear cell foci の進展に伴う腫瘍病変つ いては,あまり明確にされておらず,本検索でも不明であった。
Clear cell foci や eosinophilic タイプでみられた肝細胞間での Cx 発現の減少は,コ ネクソンの減少を意味し,basophilic タイプでみられたスポットの増大は Cx が凝集し て正常なコネクソンの構造を保持できていないことが考えられる (47)。また, eosinophilic タイプや bsophilic タイプでみられた類洞側での発現は,肝細胞間の局在 ではないためGJIC には直接関与していない。これらの病変と Cx の局在異常との関 わりは不明でさらなる検討が必要であるが,いずれの病変についても正常なGJIC の 機能が維持されていないと考えられる。一方,細胞質が好酸性を示す肝細胞変異巣や 腫瘍は滑面小胞体やペルオキシゾームの増加を伴い,好塩基性を示す肝細胞変異巣や 腫瘍はリボゾームに富んでいる (5, 22) ことから,細胞内小器官の増加が,Cx26 お よび Cx32 の各病変における局在パターンの差に関与しているかもしれない。
Cx 局在の画像解析では,免疫染色の陽性スポットの数と総面積が clear cell foci, eosinophilic あるいは basophilic タイプでそれぞれ異なることを明らかにした。しかし, 肝細胞質内での分布パターンについては,評価できなかった。Clear cell foci の局在減 少が画像解析で認められなかったことは,大きさの異なる細胞が混在していたことに よると考えられる。また,eosinophilic タイプと basophilic タイプで Cx の局在に細胞 間と類洞側の細胞膜で相違が認められたが,それらの部位を区別して計測できなかっ たため画像解析では違いが示されなかった。
32 Cx26 と Cx32 は,正常細胞では肝細胞膜上で共に局在するが (64, 74),前がん病変 では必ずしも一致しないことが示唆されている。DEN 投与とその後の部分肝切除に より発現するラットの肝細胞変異巣では,Cx32 の減少と同時に Cx26 の局在増加が認 められ腫瘍で減少する (58)。さらに,Cx26 の増加は,DEN とテトラクロロジベンゾ パラダイオキシンの併用投与時にも認められるが,PB の併用投与時には認められな いことから化学物質に依存する変化と考えられている (49)。本検討で対象とした肝増 殖性病変でCx26 と Cx32 の局在に違いがなかったことは,自然発生時の Cx 局在の特 徴であると考えられる。一方,Cx26KO マウスでは,GJIC が阻害されず,自然発生あ るいはDEN と PB の併用投与による肝腫瘍が Wild-type マウス (Cx26KO の系統) と 比べて増加しないため,Cx32 の発現減少あるいは GJIC 阻害がプロモーションあるい はプログレッションに関与すると考えられている (38)。このことから,マウスの肝増 殖性病変にみられたCx32 の局在変化は,腫瘍発現あるいは進展に関わっていること が示唆された。 以上,マウスの肝増殖性病変におけるCx の局在は,病変のタイプによって異なり, 3 つのパターンに分類されることが明らかとなった。 5. 小括 104 週齢の B6C3F1 マウスの肝増殖性病変における Cx26 と Cx32 の局在を明らかに するために,形態学的分類を行い,その後,Cx32 および Cx26 の発現を免疫染色と画 像解析で解析した。その結果,肝病変によってCx の局在が異なり,3 つのパターン に分類された。さらに,病変が進展しても同様な局在パターンが認められることが明 らかとなった。
33
Fig. 5 Microscopic findings of clear cell foci in male B6C3F1 mice. Note reduced Cx32 positive spots in the lesion. A, H&E stain; B, Cx32 immunostaining. Original magnification: 120×.
34
Fig. 6 Microscopic findings of the eosinophilic foci (A and B), and adenoma (C and D) and carcinoma (E and F) having the eosinophilic cytoplasm in male B6C3F1 mice. Note reduced Cx32 positive spots between the hepatocytes, and a strong positive staining in the cell membrane facing the perisinusoidal space. A, C, and E, H&E stain; B, D, and F, Cx32 immunostaining. Original magnification: 120×.
35
Fig. 7 Microscopic findings of basophilic foci (A and B), adenoma (C and D) and carcinoma (E and F) having basophilic cytoplasm in the liver of male B6C3F1 mice. Note an increase in the number of Cx32 positive spots. A, C, and E, H&E stain; B, D, and F, Cx32 immunostaining. Original magnification: 120×.
36
Table 5. Image analysis for Cx26 and Cx32 stainings in altered hepatic foci of male B6C3F1 mice
N F N F N F
Spot number/hepatocytes 9.29 9.14 8.58 9.3 11.4 10.3
± 2.9 ± 2.8 ± 2.3 ± 4.1 ± 3.7 ± 4.5 Total spot area/hepatocytes (µm2) 25.6 35.9 22.0 22.7 23.3 35.3
± 13.8 ± 27.3 ± 7.58 ± 14.1 ± 8.3 ± 23.8*
Spot number/hepatocytes 10.3 8.9 8.8 5.8 11.2 12.4
± 1.6 ± 1.6 ± 2.6 ± 3.1* ± 3.9 ± 3.5 Total spot area/hepatocytes (µm2) 25.2 29.7 14.8 7.4 22.2 34.0
± 7.8 ± 7.1 ± 2.7 ± 5.7* ± 8.3 ± 11.0* Clear cell foci Eosinophilc foci Basophilic foci
Cx26
Cx32
4 Classification
Stain 4 4
Significant difference from non-lesion areas. *P<0.05 by the Student’s t-test.
Each value represents the mean ± SD of the respective groups. N: non-lesion areas, F: foci of cell alteration areas.
Table 6. Image analysis for Cx26 and Cx32 stainings in hepatic tumors of male B6C3F1 mice
N T N T N T N T
Spot number/hepatocytes 9.6 19.9 10.2 21.2 8.9 11.5 9.7 18.0
± 1.39 ± 6.9* ± 1.5 ± 9.9* ± 1.1 ± 2.9* ± 2.3 ± 7.8* Total spot area/hepatocytes (µm2) 24.2 48.3 29.9 55.2 25.9 39.0 28.5 55.2
± 7.75 ± 21.4* ± 11.9 ± 51.5* ± 5.5 ± 10.6* ± 10.2 ± 34.9*
Spot number/hepatocytes 9.3 14.0 8.8 21.4 9.5 14.2 8.6 13.2
± 3.7 ± 5.0* ± 2.6 ± 21.1* ± 3.3 ± 6.4* ± 2.1 ± 5.5* Total spot area/hepatocytes (µm2) 20.1 31.6 21.2 70.9 23.3 42.1 17.8 39.9
± 8.4 ± 16.2* ± 9.7 ± 90.6* ± 9.3 ± 21.2* ± 6.9 ± 23.2* Eosinophilic type Cx26 Cx32 Basophilic type Adenoma 8 Carcinoma Adenoma Carcinoma Classification
Stain 8 8 8
Significant difference from non-tumor areas. *P<0.05 by the Student’s t-test.
Each value represents the mean ± SD of the respective groups. N: non-tumor areas, T: tumor areas.
37 第3章 Cx32 ノックアウト(Cx32KO)マウスの肝臓の細胞間コミュニケーション (GJIC) と増殖性病変 1. 小序 前章では,104 週齢の B6C3F1 マウスの肝増殖性病変では,肝細胞間での Cx32 の 発現が変化していることを免疫染色と画像解析で明らかにした。また,Cx32 の発現 異常と肝増殖性病変との関連が示唆されたことから,Cx32 をノックアウトしたマウ ス (Cx32KO マウス) では,肝増殖性病変が増加する可能性が考えられた。 本実験では,Cx32KO マウスを用いた。本系統マウスは,Cx32 の欠損あるいは GJIC 阻害と肝腫瘍発現との関わりについての研究成果が既に報告 (16, 42) されている。し かし,Cx32KO マウスと Wild-type マウス (C57BL) 間には肝増殖性病変の発生に差は なく (16),自然発生の増殖性病変に対して,GJIC 阻害によるプロモーションあるい はプログレッション作用を及ぼさないとの指摘がなされた (16)。一方,C57BL マウ スの肝腫瘍病変は,加齢とともに発生率が高まることが知られているが (46),Cx32KO マウスの自然発生に関する既報の結果は18 ヶ月齢までの検索結果に基づくものであ り,さらに長期飼育することで肝増殖性病変の発現が高まる可能性は否定できない。 また,Cx32KO マウスに発生する肝増殖性病変の組織学的特徴の詳細についても明ら かではなかった。 本章では,まずCx32KO マウスの Cx26 および Cx32 の肝組織内局在を確認する とともに,incision loading/dye transfer (IL/DT) assay で GJIC 機能を Wild-type マウスと 比較し,肝臓における Cx32 の欠損と GJIC の阻害の関連を調べた。IL/DT assay は, GJ を通過できる低分子蛍光色素である Lucifer yellow (LY) を肝臓の切開部位に滴下 し,その拡散の程度からGJIC の状態を評価する手法であり(28),Cx32KO マウスを用 いるに際して,GJIC が阻害されていることを本手法により確認した。その後,長期 飼育したCx32KO マウスおよび対照の Wild-type マウスの自然発生の肝増殖性病変
38 を病理組織学的に検索した。 2. 材料および方法 2-1 使用動物 Cx32KO マウスおよび Wild-type マウスは,エコンケージに 5 例ずつ収容してバリ アーシステムの動物室内で飼育した。動物室の飼育環境は,第1 章の ”2-1” と同 様である。餌には,固形飼料 (Certified Rodent 5002: PMI Nutrition International, 米国) を与え,水道水を自由に摂取させた。なお,本系統は,山崎 洋博士 (関西学院大学, 兵庫) より分与を受け,第一三共株式会社 (東京)で維持・管理している動物である (Fig. 8)。動物の取り扱いは,全て「動物の愛護及び管理に関する法律」 (1973 年 10 月1 日法律 105 号,1999 年 12 月 22 日改正) に従って行い,第一三共株式会社の動物 実験倫理規定に従って実施した。 肝臓のCx26 および Cx32 の免疫染色と IL/DT assay の検討には,健常な 20 およ び104 齢の雌雄 Cx32KO マウスと Wild-type マウス (n = 4) を用いた。 自然発生の増殖性病変の検討には,104 週齢の雌雄 Cx32KO マウスと Wild-type マ ウス (n = 50) を使用した。 2-2 使用試薬
IL/DT assay に用いた Lucifer Yellow (LY) は,Sigma (米国)より購入した。
2-3 肝臓の Cx26 および Cx32 の免疫染色
Cx32KO および Wild-type マウス (n = 4) をエーテル麻酔下で安楽死させ,肝臓を 摘出した。その後,内側右葉から厚さ約4 mm の組織片をトリミングし,OTC コンパ ウンド中に包埋して未固定のまま凍結ブロックを作製した。これらのブロックから約 8 m の凍結切片を作製し,-20 ℃のアセトンで 5 分間固定した後,第 1 章の“2-6”
39
の蛍光抗体法による手法を用いて Cx26 および Cx32 の免疫染色を施した。
2-4 肝臓の IL/DT assay
上記“2-3”で摘出した肝臓を用いて,Hu らの方法 (28) に従って実施した。肝臓 の外側左葉の一部を幅 5 ~ 8 mm,深さ約 2 mm に切開し,0.5% Lucifer Yellow(LY) PBS (0.05 M,pH 7.4) 溶液を滴下した。3 分経過後,切開部を含む組織片をトリミン グし,PBS で 3 回洗浄した後に 10% 中性緩衝ホルマリン液で一晩固定した。固定 した組織片は,99.5% エタノールで脱水し,キシレンを経てパラフィンに包埋した。 5 m の切片を薄切してスライド貼付した後,蛍光顕微鏡を用いて LY の切開部か らの拡散の程度を観察した。 2-5 自然発生の増殖性病変の病理学的検査 Cx32KO および Wild-type マウス (n = 50) を生後 24 ヶ月間飼育し,エーテル麻酔 下で放血して安楽死させ,各種器官・組織の肉眼的変化を調べた。また,一般状態が 悪化または死亡した動物 (Cx32KO マウス:雄 1 例および雌 6 例,Wild-type マウス: 雌8 例) については,その時点で解剖して各種器官・組織の肉眼的変化を観察した。 剖検時に肝臓を採取して,10% 中性緩衝ホルマリン液で 3 日間固定した後,外側 左葉,内側左右葉および外側右葉について約 3 m の病理組織標本 (H&E 染色) を作 製し,光学顕微鏡を用いて観察した。また,以下に示す肝臓以外の器官・組織につい ても病理組織標本を作製し,脳,腎臓,心臓,肺,脾臓,下垂体,甲状腺,副腎,胸 腺,精巣,前立腺,精巣上体,精嚢,卵巣,子宮,唾液腺,ハーダー腺,骨格筋,膀 胱,舌,包皮腺,膣,胃,消化管 (十二指腸,空腸,回腸,盲腸,結腸,直腸),皮膚, 乳腺,大腿骨・胸骨 (骨,骨髄),坐骨神経,脊髄 (頚部,胸部および腰部),腸間膜 リンパ節,腎門リンパ節および眼球の組織学的検査を実施した。
40 2-6 統計解析 病理解剖額的および病理組織学的に観察された所見について,Fisher の直接確率検 定を行った。有意水準は 1% および 5% とした。 3. 結果 3-1 肝臓の Cx26 および Cx32 の局在 Cx32KO マウスおよび Wild-type マウスとも,Cx26 の局在は,肝小葉全体の肝細 胞の細胞膜上でスポット上に確認されたが,Cx32 の局在は,肝細胞で全く検出され なかった (Fig. 9)。本結果には,20 週齢と 104 週齢で違いはなく,性差もなかった。 3-2 肝臓の IL/DT assay
肝臓の IL/DT assay の所見は,Fig. 10 に示した。
Cx32KO マウスの肝臓では,切開部の肝細胞の一部のみに LY が確認されたが, 隣接する肝細胞への拡散は全く認められなかった。一方,Wild-type マウスの肝臓で は,LY の肝細胞内における拡散が認められた。本結果には,20 週齢と 104 週齢で違 いはなく,性差もなかった。 3-3 自然発生の増殖性病変 病理解剖学的および病理組織学的結果をTable 7 に示した。
Cx32KO マウスは Wild-type マウスに比べ,雄のみ肝臓の hepatocellular carcinoma の有意な増加 (Cx32KO: 9 例,Wild-type: 0 例) が認められた。この病変では, eosinophilic タイプと basophilic タイプの両方が確認され (Fig. 11),hepatocellular adenoma も増加傾向 (Cx32KO:4 例,Wild-type:0 例) を示した。雌ではこのような変 化は軽微であった。
41
その他病変としては,非腫瘍性病変として雄の腎臓でvacuolation of tubular epithelium の増加が認められた。 なお,瀕死解剖例または死亡例には,それらの原因と判断される血管肉腫,組織球 性肉腫および悪性リンパ腫が確認されたが,Cx32KO マウスと Wild-type マウスで差 はなかった。 4. 考察 Cx32KO マウスは,Cx32 遺伝子発現の欠損により,Cx32 の肝細胞膜での局在が 消失し,GJIC が阻害されていると考えられた。20 および 104 週齢の Cx32KO マウ スでは,免疫組織化学的染色でCx26 が肝細胞の細胞膜上に確認されたが,Cx32 の 局在は肝細胞で全く検出されなかった。また,IL/DT assay では,隣接する肝細胞への LY の拡散は全く認められなかった。このことから,Cx32KO マウスでは,Cx32 遺伝 子発現の阻害によってCx32 蛋白が発現せず,GJIC が阻害されていることが示された。 Cx26 の局在は確認できたが,コネクソン構築には Cx32 が必須で Cx26 だけではコネ クソンを構築できないことが知られている (38)。 次に,24 ヶ月齢 Cx32KO マウスおよび Wild-type マウスの自然発生する病変につ いて病理組織学的に検索した。Cx32KO マウスの雄の肝臓で,Wild-type マウスに比 べてeosinophilic および basophilic タイプの carcinoma が有意に増加し,Cx32 欠損の関 与が考えられた。本carcinoma は,雄で発現が高く,明確な性差が認められた。一般 的に,肝臓腫瘍の発現は,雌よりも雄の方が高く,性ホルモンの影響に起因すると考 えられている (16)。すなわち,去勢した雄性マウスでは腫瘍が減少,卵巣を摘出した 雌性マウスでは腫瘍が増加 (72),雄性マウスにテストステロンを過剰投与で腫瘍の増 加が確認されている (12, 57, 67)。第 1 章での肝 Cx32 の発現には性差がないことを考 え合わせると,Cx32KO マウスにおける増殖性病変の性差は,性ホルモンの影響が一 部関与しているかもしれない。
42
肝臓以外の器官・組織については,Cx32KO マウスの雌の下垂体で adenoma が Wild-type マウスに比べて有意に減少したが,下垂体では,Cx32 の発現が認められて いないことから,Cx32 の欠損と直接関係のない変化と考えられた。
非増殖性病変として,Cx32KO マウスの雄の腎臓に vacuolation of tubular epithelium の有意な増加が認められた。本病変は C57BL マウスでしばしば認められる変化 (71) であるが,対照としたWild-type マウスで認められなかったこと,腎臓では Cx32 が発 現することから,Cx32 欠損の影響か否かについては精査が必要である。その他,脳 においてもCx32 が発現することが知られているが,光学顕微鏡レベルの組織形態の 観察において,それら器官・組織において変化は確認されなかった。 以上,Cx32KO マウスで,Cx32 の欠損に直接起因する病変は,肝の増殖性病変の 増加と考えられた。また,第 2 章において,肝増殖性病変は Cx32 の局在パターン によりeosinophilic と basophilic タイプ に分類されたが,Cx32KO マウスにおいても 両タイプの腫瘍が発現した。しかし,Cx32KO マウスでは Cx32 が発現していないこ とから,Cx32 の発現と腫瘍の組織形態とは必ずしも一致しないことが示された。
5. 小括
長期飼育のCx32KO マウスに発生する肝増殖性病変と GJIC 阻害との関連を調べる ために,肝Cx の免疫染色と GJIC の機能を把握する incision loading/dye transfer (IL/DT) assay を行った。その結果,20 および 104 週齢の Cx32KO マウスの雌雄で,Cx32 の 局在消失とともにGJIC 阻害が確認された。104 週齢の Cx32KO マウスでは,雄の肝 臓で増殖性病変の発生率がWild-type マウスより増加し,eosinophilic と basophilic タ イプの病変が発現した。したがって,Cx32 の欠損は GJIC を阻害し,肝増殖性病変の 促進に関与していることが示唆された。
43
Fig. 8 Targeted disruption of the mouse Cx32 gene. The Cx32 gene was disrupted by replacing the exon 2 of the Cx32 gene with Neomycin resistant casette (neoPy) in J1 ES cells. 1, Targeting vector construct; 2, The wild allele of the Cx32 gene; and 3, The predicted mutant allele.
44
Fig. 9 Cx26 and Cx32 immunostaining in male Cx32KO mouse liver at age of 20 weeks. Note diffusely localization of Cx26 protein on the hepatocyte membranes (A) and deficient Cx32 protein localization (B). Original magnification: 120×.
Fig.10 Hepatic dye-transfer of Lucifer Yellow (LY). Wild-type mice (A ) and Cx32KO mice (B) at age of 20 weeks. Note spreading of LY (arrows) on the hepatocyte only in wild-type mice and low or deficient spreading of LY (asterisks) in Cx32KO mice. Original magnification: 100×.
B
A
45
Table 7. Spontaneous neoplastic and non-neoplastic lesions in wild-type and Cx32KO mice during a 104-week observation period
Organ
Lesions Pathological findings Wild-type Cx32KO Wild-type Cx32KO
Liver
MACRO Nodule 4 (8) 14* (28) 2 (4) 10* (20)
MICRO Histiocytic sarcoma 6 (12) 7 (14) 3 (6) 7 (14)
Hemangiosarcoma 1 (2) 2 (4) 0 1 (2)
Hamangioma 3 (6) 2 (4) 1 (2) 1 (2)
Malignant lymphoma 4 (8) 5 (10) 3 (6) 5 (10)
Hepatocellular carcinoma 0 9** (18) 0 2 (4)
Hepatocellular adenoma 0 4 (8) 1 (2) 5 (10)
Mast cell tumor 0 0 1 (2) 1 (2)
Pituitary
MACRO Nodule 0 0 8 (16) 0*
MICRO Adenoma in pars distalis 0 0 10 (20) 2** (4)
Adenoma in pars intermedia 0 0 0 1 (2)
Carcinoma 0 0 1 (2) 0
Liver
MACRO Colored area 7 (14) 9 (18) 1 (2) 4 (8)
Rough surface 1 (2) 0 2 (4) 2 (4)
Atrophy 1 (2) 0 0 2 (4)
Discoloration 1 (2) 1 (2) 0 1 (2)
Cyst 0 4 (8) 1 (2) 2 (4)
Enlargement 0 0 2 (4) 1 (2)
MICRO Increased granulopoiesis 2 (4) 1 (2) 0 1 (2)
Inflammatory cell infiltration 1 (2) 0 0 0
T hrombosis 1 (2) 0 0 0
Proliferation of Kupffer cell 2 (4) 0 1 (2) 1 (2)
Angiectasis 1 (2) 0 1 (2) 1 (2)
Congestion 1 (2) 0 0 0
Fatty change of hepatocyte,
centrilobular 1 (2) 1 (2) 0 0
Biliary cyst 0 3 (6) 0 2 (4)
Eosinophilic foci 0 1 (2) 1 (2) 3 (6)
Focal necrosis of hepatocyte 0 1 (2) 0 1 (2)
Clear cell foci 0 0 1 (2) 0
Vacuolated cell foci 0 0 0 1 (2)
Eosinophilic cytoplasmic ateration
of bile duct epithelium 0 0 1 (2) 0
Pituitary
MACRO Colored area 0 0 2 (4) 1 (2)
Enlargement 0 0 1 (2) 0
MICRO Cyst 1 (2) 3 (6) 1 (2) 3 (6)
Focal hyperplasia 0 1 (2) 0 1 (2)
Cyst in pars distalis 0 0 1 (2) 0
Focal hyperplasia in pars distalis 0 0 6 (12) 7 (14)
Hyperplasia in pars intermedia 0 0 2 (4) 1 (2)
Kidney MACRO Discoloration 1 (2) 0 1 (2) 1 (2) Cyst 2 (4) 0 1 (2) 1 (2) Enlargement 0 0 1 (2) 0 Colored area 0 0 1 (2) 1 (2) Rough surface 0 0 0 1 (2)
MICRO Dilatation of renal pelvis 0 1 (2) 0 1 (2)
Mineralization 0 2 (4) 0 0
Cyst 2 (4) 3 (6) 2 (4) 2 (4)
Osseous metaplasia 1 (2) 0 0 0
Vacuolation of tubular epithelium 0 11** (22) 0 0
Chronic nephropathy 5 (10) 5 (10) 9 (18) 10 (20) Glomerulopathy 0 0 2 (4) 0 Hyperplasia of transitional epithelium 0 0 1 (2) 0 Males Females Neoplastic Non-neoplastic
* p < 0.05,** p < 0.01 vs. the Wild-type group by the Fisher's exact probability test. Entries are the number of animals, including those that died or were killed in extremis. Data in parentheses show incidence (% of animals) of each lesion. n = 50. MACRO: Macroscopic examination. MICRO: Microscopic examination.