原 著 ( 東 女 医 大 誌 第55巻 第10・ 問 1 頁 999~1014 昭和60年11月j 岡本糸枝賞受賞論文
肺胞マクロファージの感染防御機構に関する研究
日本大学医学部微生物学教室石 橋
悌 子
( 受 付 昭 和60年8月26日)Studies on Role of Alveolar Macrophages and
Bronchoalveolar Lavage Fluids in Host Defense Mechanisms Teiko ISHIBASHI
Department of Microbiology, Nihon University School of Medicine
1 examined the extracellular release of the bactericidal substances
,
lysosomal enzymes,
lysozyme and long-chain fatty acids from alveolar macrophages (AMφ) of rabbits and a role of bronchoalveolar lavage f1uids (BALF) and AMφin the host defense mechanisms.The results obtained were as fol1ows.
1) There is a c10se correlation between the bactericidal activity of AMφand long-chain fatty acids released from AMφ.
2) Cyc1ic nuc1eotides may play a role in the regulation of the release mechanisms of the bactericidal substances, lysosomal enzymes and long-chain fatty acids from AMφ.
3) In the presence of BALF from complete Freund's Adjuvant (CFA)ーinjectedrabbit, extracel1ular
release of the bactericidal substances, lysozyme and lysosomal enzymes from normal and CFA-AMφ exposed to opsonized zymosan (OZ) were markedly enchanced.
On the other hand, contents of total fatty acids and long-chain fatty acids such as palmitic acid, oleic acid and linoleic acid in the supernatants of OZ-ingested AMφin the presence of BALF were decreased. This decrement may be attributed to incorporation of these fatty acids into the AMφand/or to the selective alteration of fattyacyl composition of AMφphospholipids.
1 conc1ude from the results obtained here that the BALF, co-operated with AMφmay play an im -portant role in the defense mechanisms of lung infections. はじめに 生体内に,細菌や毒性のある異物が侵入すると, 生体の示す感染初期の防御機構として,多形核白 血 球CPMN),マグロファージCMφ〉などのいわ ゆる食細胞による貧食,消化,殺菌作用が重要な 役割を果していることは, 19世紀末にMetchini -ko妊が指摘したところである. 以来,多くの研究,報告がなされてきたが, と くに1975年Klebanoff1ト3)がPMNの殺菌機構に 関して,酸素に依存する殺菌系と,酸素に依存し ない殺菌系とに二大別し報告した. 999 前者はmyeloperoxidase CMPO),またH202 Chydrogen peroxide)
,
0"2Csuperoxide入・
OH Chydroxy radical),102 Csinglet oxygen)などの 活性酸素による殺菌系であり,後者はphagosome 内のpHの酸性化,lysozyme, lactoferrin, lysoso -me酵素およびcationicproteinなどによる殺菌 作用であり,これらの殺菌系はすでに確認されて いる. 食細胞はこれら殺菌機構のいくつかが同時に, あるいは相前後して働き,さらに免疫グロプリン や補体の協力による液性免疫や,細胞性免疫による特異的な防御機構へとリレーされて巧みに生体 の防御が営まれている. 著者らも,ここ数年来,PMNさらに血小板中の lysosomeの殺菌作用に関して一連の研究を報告 してきた4ト10) Klebanoffの 提 唱 し た 以 上 の よ う な 殺 菌 機 構 で,生体内に侵入してきた細菌の多くは殺菌され ることになるが,さらに処理しきれない細菌は
Mrt
が貧食し,細胞内で殺菌して感染の進展を阻 止していくことになる.しかし,病原徴生物の中 にはMycobacteriumtuberculosis,
Mycobacter -ium leprae, Brucella, Listeria, Salmonella, Nocardiaさらにまた原虫のように多量の活性酸 素に曝されている食胞 CPhagosome)内において 生存し,さらに増殖しうる細胞内寄生菌とよばれ る微生物がある. これら細胞内寄生菌に対する殺 菌機構にはPMNのみでは不充分で,活性化され たMφ がこれに大きく関与しており,活性化Mrt
はHzOz
を は じ め 種 々 の 活 性 酸 素 を 大 量 に 産 生 し,当然殺菌能を発揮してくるが,細胞内寄生菌 に対しては,これのみで充分殺菌しうるとは考え られず,著者らは,食細胞が侵入菌その他の異物 を貧食した際に,細胞内のlysosome酵素活性が 瓦 進 し lysosome由来の酵素の一つであるphos -pholipase,Az
によって,細胞膜に存在するphos -pholipidsが分解され遊出してきた殺菌性のある 長鎖脂肪酸もまた,食細胞の殺菌機構において重 要な系の一つで、あることを知り報告した11)-13) 今回は,食細胞の中でも最も気道の末端に存在 して,気道を介する肺感染の防御に重要な役割を 果している肺胞マクロファージ CAMφ)につい て, とくに殺菌機構の面から検討した成績を報告 する. 実験材料および実験方法 1.A Mゅの採取 体 重2.0-2.5kgのNew Zealand Whiteウサ ギにCompleteFreund's Adjuvant (Difco製, CFA)を1ml/rabbit静注し, 3週 間 後 に Myrvikl4 )らの方法に従って heparin(5μ/mD加 Hanks balanced salt solution CHBSS)により経 気道的に50mlずつ3回洗浄して,気管支肺胞洗浄 液 Cbroncoalveolarlavage fl.uids, BALF)を得 て, 1,500rpmで10分間遠心し,沈撞の細胞を集 め,さらにHBSSで洗浄して得られた細胞を1x 107cells/mlと な る よ う にHBSSに 再 浮 遊 し AMφの浮遊液を作製した.このAMφ はその都 度, trypanblue exclusion~こより生存性と,併せ てガラス付着性の性状も確認している. 2. opsonized zymosan (OZ)の作製zymosan CSigma)をCa++
,
Mg++加phosphate bufferd saline CPBS+)に3mg/ml~こ浮遊し, 100'C 30分 煮 沸 後PBS+で3回洗浄し, 3mg/ml PBS+浮遊液とした. この1容に,正常ウサギ血清9容を加え37'Cで 30分間振還しつつ処置し, 4 'C8,500rpm 20分間 遠心し,沈置をPBS+で2回洗浄して,最終濃度3 mg/mlのOZPBS+浮遊液とした. 3. lysosome酵素活性の測定 lysosomeのmarker enzymesとして, acid phosphatase CAc-Pase)とβ-glucuronidase(β -Gase)について,前嶋へ西村4)の方法で測定し,Ac-PaseはK-Auで, β-Gaseはu/dl/hで表わし
た
4
.
殺菌作用の測定 被検液1mlの希釈系列をつくり,これに37'C18 時間培養の被検菌液を添加して,そのO.lml中の 生存菌数を混合平板培養法によって計数し,菌液 添加直後の生菌数 (a) と菌液添加後3TC
2時間 保持後の生菌数 (b) とから次の式によって殺菌 率を求めた.すなわち, 殺菌率(%)=三Q.x100 で,この方法は,従来から当教室で行っている方 法である. 被検菌としては, Klebanoffの提唱した殺菌系 で殺菌される菌として Streptococcus,Escheri -chia coli又リゾチーム活性の対象菌として Mi -crococcus lysodeikticusさらに細胞内寄生菌と してはListeriamonocytogenesを用いた.5. AMφ から heptaneextracts (HE)の作製 法
1 x 107cells/mlの
AMrt
のHBSS浮 遊 液 を プ1000-ラ ス テ ィ ッ ク シ ャ ー レ (Lux,5220)に5mlずつ分 注 し , 一 部 に はOZを 添 加 し て3TCのCO2-in -cubatorで 所 要 時 間 保 持 し た 後 の 遠 心 上 清 液 か ら, Kochan同らの方法に準拠して HEを得た.す なわち,上清液1mlVこ対し, 5mlのisopropanol heptane-2N H2SO. (40 : 10 : 1)を加え室温に10 分間保持後, 2mlのheptaneと3mlの滅菌精製氷 とを加えて,上清のheptane層を採り, 56'CでN2 gas下でheptaneを蒸発さぜてHEを得た. 6. HE のgaschromatographyによる脂肪酸 分 析 HEをmethylester化して,島津製gaschro -matography GC-6 A Mを用いて分析し,さらに島 津製C-RIAchromatopacによって,各種脂肪酸 量を測定した. なおinternalstandard(IS)として pentadeca -noic acid methyl ester (Merk)を使用した. また実験方法でとくに記していない方法は,そ の都度実験成績の項で述べる. 実験成積
1
.
PMNおよびAMφlysateの殺菌作用なら びに lysateから得たHEの指肪酸分析 内藤ら13)は, AMctとPMNの1X 107 cells/ml 浮遊液を凍結融解して得た上清液 (pre-incubate lysate)と,さらにこの上清液を3TC,3日間lll -Preincubated Incubated fo( 3 days at 37 C Incubated with indomethacin (10‘M)for 一InClIbated with
BSA 11%) for 2 J davs atJ7C 5 10 cubateして得たlysateにつき殺菌作用を検討し た このときのPMNはNew Zealand Whiteウ サ ギの腹腔内に0_2%にglycogenを加えた生食水 200mlを注射して 4時間後十こheparin加HBSS で腹腔内を洗浄し細胞を集めたものである. Fig_ 1はその成績で, PMNのlysateは, Stre -ptococcus pneumoniae, Streptococcus pyogen -es, Listera monocytogenes に対し弱し、ながら殺 菌作用が認められるが, AMctのlysateにはこれ ら菌種に対する殺菌作用は全く認められなかっ た.しかし,リゾチームの対象菌である Microco -ccus lysodeikticusに対しては, PMN,
AMct
両 細胞のlysateともに強い殺菌作用を示していた. さらにPMN,AMφ を 凍 結 融 解 し た 浮 遊 液 を 3TC, 3日間保持して得たlysateは, Streptoc -occus pneumoniae, Listeria monocytogenesに 対する殺菌作用は著しく増大しており, Streptoc -occus pyogenesに対しても殺菌作用の増大が認 められた.この凍結融解後,3TC, 3日間保持後 のlysateに み ら れ た 殺 菌 作 用 はphospholipase A2 CPLase A2) の阻害作用をもっindomethacin (IM)を1O-4Mに,あるいは脂肪酸の殺菌作用を中 和する血清アルブミンCfattyacid freeのbovine albumin, BSA)を1%に凍結融解した浮遊液に添 亡=コPMN . 圃 圃A M世 Fig.1 Bactericidal Activities of Lysates from PMN and AM<,b 1001ー加して ,
3rC
,3
日間保持することによって,図 にみられるように, 3日後のlysateの殺菌作用は indomethacinによって影響を受けないリゾチー ム 作 用 の 対 象 菌 で あ るMicrococcus lysodei -kticusに対しては強く殺菌作用を示すが,他の菌 種に対しては著しい殺菌作用の低下を示した. なお,3rc
,3
日間保持後のlysateの示す殺菌 作用はP M Nvこ比べて AMftの方が,その活性は 大であった. さらにTable 1はP M Nの 脂 肪 酸 分 析 の 表 で あるが, incubate前に比べて 3日間incubate後 のHE
中には,総脂肪酸量が約7
.
7
倍と増量し,各 脂肪酸量はノξルミチン酸(16:0),ステアリン酸 (18: 0),オレイン酸(18:1)さらにリノール 酸(18: 2)の各長鎖脂肪酸量の増大が顕著であっ Tこ Table 2はAMφ のHE
中の脂肪酸分析の成績 であるが,3rc
保持前のHE
中の総脂肪酸量に比 べて3日間保持後では,そのlysateのHE
中の総 脂肪酸量は約25.5倍と著明に増大し,各種脂肪酸 量もパルミチン酸(16: 0),ステアリン酸(18: 0),オレイン酸 08:1), りノール酸(18: 2) Table 1 Identification and Quantification of Fatty Acids in Heptane Extracts of Polymor-phonuc1ear LeukocytesIncubated ncubated with Fatty acids Preincubated for 37d℃ ays indomethacin (μg/m]) at 3 or 3day/ms l at (μg/m]) 3TcCμg/m]) 12: 0 1.840 1.075 14:0 0.673 1.602 0.763 16: 0 3.568 31.926 3.655 16: 1 0.308 1.593 0.248 18: 0 2.150 23.737 1.908 18・1 2.414 24.237 1.840 18: 2 5.993 67.137 2.073 18: 3 0.519 2.073 0.365 20: 0 0.420 1.442 0.275 20: 1 0.650 3.420 0.443 20: 4 5.401 5.758 7.488 22: 0 1.327 22: 1 1.491 24: 0 0.45 24: 1 1.877 Tota! 22.096 169.910 23.498 Table 2 Identification and Quantification of Fatty Acids in Heptane Extracts of Alveolar Macrophages Activated with Freund's Adjuvant Incubated Incubated with Fatty acids Preincubated for 37d℃ ays indomethacin (μg/m]) at 3 for 3dagy/ms al) t (μg/m]) 3TcCμ 14: 0 0.147 1.942 16: 0 2.116 56.906 9.540 16: 1 0.489 6.132 1.875 18: 0 1.384 41.479 6.927 18: 1 1.703 48.771 9.630 18: 2 0.665 34.200 5.804 18: 3 0.312 2.618 0.391 20: 0 0.136 1.087 0.457 20: 1 0.149 3.367 0.179 20: 4 9.987 1.877 22: 0 0.392 2.678 2.002 22: 1 0.635 0.219 24: 0 7.740 1.466 24: 1 1.392 4.755 0.995 Tota! 8.894 226.643 41.362 さらにアラキドン酸 (20: 4)のように保持前に は検出されなかった長鎖脂肪酸も検出されてい る.なおこのAMftは
CFA
注射ウサギから,注射 3週間後に採取したものである.またTable 1, Table 2でみられるように1Mを10-4M に添加し incubateすると,総脂肪酸量も各種脂肪酸量も著 明に減少し,とくにP M NのHE
中にはincubate 前の脂肪酸量とほとんど相異がみられなかった. これらの成績からしても食細胞の殺菌機構の一 つの系として,長鎖脂肪酸の関与していることが わかる. 2. AMftからのlysosome酵素,殺菌性物質な ら び に 長 鎖 指 肪 酸 の 細 胞 外 放 出 機 構 に 及 ぼ す cyclic nucleotidesの影響 1958年Sutherlandら16)によって cyclic A M P (cAMP)が,ついで1963年Ashman17 )らによって cyclic G M P CcGMP)が発見されて以来,生体内 における cyclic nucleotidesの機能に関して多数 の研究報告が行われ, cyclic nucleotidesがヒトも 含めて動物細胞の中で各種情報のsecond mes-sengerとして働き, cAMPとcGMPとは互に措 抗しながら生命現象をcontrolしていることが明 らかになってきている. -1002一そこでAMφ は異物責食時に種々の生物活性 を有する物質を細胞外に放出してくるが,との放 出機構にcyclicnucleotidesが関与しているもの と思惟して実験を行なった.その成績の一部を述 べる cyclic nucleotidesと し て は phosphodieste r-aseの作用を受けにくく,かつ細胞内への透過性
の高いdibutyrylcAMP CDB-cAMP)と8bromo cGMP C8Br-cGMP)を選び,所要濃度に PBS+に 溶解して, AMφ 浮遊液に添加し, 370 C, 60分lll -cubateし, cyclic nucleotides処理 AMrtとした. 1)AMφ の lysosome酵 素 の 細 胞 外 放 出 に 及 ぼすcyclicnucleotidesの影響 Table 3は DB-cAMPと, 8Br-cGMP処 理 AMrtを5mlずつシャーレに分注し,さらに
o
z
を添加して3TC45分, CO2-incubator中に保持し, 上清液を集め,ついで5.000rpm20分間遠心して 得た上清液について,lysosome酵素活性を検討し た成績で、ある.Ac-Paseとβ-Gaseは dosedepen -dentに DB-cAMP処理によって細胞外放出は抑 制され, 8Br-cGMP処理によって増大が認められ る 2) AMφ からの殺菌性物質の細胞外放出に及 ぼすcyclicnucleotidesの影響 Table 3で lysosome酵素活性を測定した遠心 上清液について,細胞内寄生菌の一つで、あるLis -teria monocytogenesに対する殺菌作用の成績を Fig. 2に示した.図にみられるように controlで ある無処理のA M<tの遠心上清液中には殺リス テリア菌性物質のreleaseはみられないが, OZの 添加により上清液中に殺リステリア菌性物質が releaseしてくる.また A M<tを8Br-cGMPで前 処理したのみでも遠心上清液中には,この20倍希 釈液でも約50%の殺菌率を示した. また8Br-cGMP前 処 理AM<
T
vこOZを 添 加 す ると殺リステリア菌性物質の細胞外放出は更に増 大していた.しかし, DB-cAMP前処理 A M<tか ら は 殺 リ ス テ リ ア 菌 性 物 質 の 上 清 液 中 へ の releaseは全くみられず, OZの添加によっても上 清液中へのreleaseは完全に抑制されていた. Fig.3は,被検菌としてリゾチームの対象菌で ある Micrococcuslysodeikticusを用いて行った 殺菌作用の成績である.被検上清液中への殺菌性 物質,すなわちリゾチームの放出は8Br-cGMP前 処理によって増大し, これにOZを添加すること によって,そのreleaseは顕著な増大を示した.しかしDB-cAMP前 処 理 AMφ からは OZの添加
によっても, リゾチームの細胞外放出は全く抑え られており, Fig. 2の成績と同じ傾向を示した. 3) cyclic nucleotides前 処 置
AM
<tの 遠 心 上 清液から得たheptaneextracts (HE) 中の脂肪 酸分析 Fig. 2, 3で実験した遠心上清液から HEを得 て, HE中の脂肪酸分析を行った成績を Table4, 5に示した.表でみられるように controlの無処理 A Mゅの上清液から得た HE中の総脂肪酸量に比 べて, DB-cAMP前処理 AMφ の上清液から得た HE中の総脂肪酸量は減少しており,さらに DB-cAMP前処理後に OZを添加しでも, A Mゆから releaseされた総脂肪酸量は, OZ添 加 の み の AMrtからの HE中の総脂肪酸量に比べて減少し ていた.これに反して8Br-cGMP前処理 A Mゆか Table3 E妊ectof Cyclic N ucleotides on Release of Lysosomal Enzymes from Alveolar Macrophages Exposed to Opsonized Zymosan二記者手
Acid Phosphatase(K-Au) β.Glucuronidase(u/dl/h) cAM!.' cGMP cAMP cGMP 10-2M c.Nucleotide'OZ 2.5 4.2 320 1025 10-3M c-Nucleotide'OZ 2.6 3.0 350 750 1O-2M c-Nucl 巴otide 2.0 3.1 250 400 1O-3M c-N ucleotide 2.1 2.6 275 375 OZ 2.7 2.6 550 500 untreated 2.1 2.1 300 275 ー 1003向 説
t
i
e
s
tr吋
;
;
;
ト
¥
A
;
:
;
ty 1O-3M DB-cAMP 1O-3M 8 Br-cGMP 50 100 50 100% c-Nucleotide OZ r ヘ υ ハ U ハ U ?ょっム c-Nucleotide OZ untreated 一 -n u n / “ Fig. 2 Effect of Cyclic Nucleotid巴son Release of Bactericidal Substances fromAlveolar Macrophages Exposed to Opsonized Zymosan Test organism: Listeria monocytogenes c Nucleotides
BfEE
百 iv王子 treated、
νith I Dil DB-cAMP 8 Br-cGMP 50 100 50 100% c-Nuc leotide 10 oz I 20 40 c-Nucleotide ハU ハU ハU 1 2 4 oz ハυ ハU ハU 1 i η L A せ ハU ハU ハ り 1L つ 乙 A μ T JU ρ i v t a e r t n u Fig. 3 E妊ectof Cyclic N uc1eotides on Release of Bactericidal Substances from Alveolar Macrophages Exposed to Opsonized ZymosanTest Organism : Micrococcus lysodeikticus ら得た
HE
中 の 総 脂 肪 酸 量 はcontrolのAMφ の上清液のHE
中に比し,約2
.
5
倍の増量がみら れ, 8Br-cGMP前処理AMφ にo
z
を添加するこ とにより,HE
中の総脂肪酸量はさらに増大し,各 脂肪酸も特にパルミチン酸 (16・0),パルミトレ イン酸(16:1),ステアリン酸 (18: 0),さら -1004Table 4 Identification and Quantification of Fatty Acids in Heptane Extracts from Supernatants of CycJic AMP-treated Alveolar Macrophages Exposed to Opsonized Zymosan
(μg/mJ) F¥a¥ttすAtc¥同id日¥twEd ¥ith 10-3乱f 10-3 M untreatedDB-c AMP 02 DB-c AMP, 02 14: 0 0.05
。
.03 0.06 0.06 16: 0 0.56 0.67 1.68 1.34 16: 1 0.06 0.07 0.31 0.23 18: 0 0.42 0.26 0.48 0.62 18: 1 0.22。
.30 1.27 1.22 18: 2 0.18 0.12 1.07 0.99 18: 3 0.06 0.13 0.24 0.23 20: 0 0.20 0.02 0.04 0.02 20: 1 0.07 0.03 0.04 0.05 20: 4 0.47 0.45 1.14 0.73 22: 0 0.02 0.02 0.02 22: 1 0.08 0.03 0.03 0.02 24: 0 0.02 0.08 0.12 24: 1 0.15 0.01 0.15 total 2.56 2.29 6.55 5.65 にアラキドン酸 (20:4),ベヘン酸 (22: 0), リグノセリン酸 (24: 0)などが顕著に検出され たFig.
4
はcontrolのAMφを3TC
,
6
0
分incuba-te後 の 遠 心 上 清 液 か ら 得 た
HE
中の gaschro-matography による脂肪酸分析の図である.図で
みられるようにノ4ルミチン酸
06:
0),ステアリD
15 <o
Table 5 Identification and Quantification of Fatty Acids in Heptane Extracts from Supernatants of CycJic GMP-treated Alveolar Macrophages Exposed to Opsonized Zymosan
(μg/ml) F¥at¥ttCried、a¥tWed I¥th 10-3M 10-3 M untreated 8Br-c GMP 02 8Br-c GMP, 02 12: 0 0.06 0.06 0.02 trace 14: 0 0.07 0.07 0.18 0.08 16: 0 0.59 1. 78 1.82 1. 95 16: 1 trace 0.06 0.23 0.25 18 : 0 0.24 0.67 0.94 1. 04 18: 1 0.61 1.55 1.00 0.99 18 : 2 0.07 0.31 1.32 0.45 18: 3 trace 0.17 0.09 20: 0 0.21 0.20 0.08 0.22 20: 1 0.07 trace 0.04 0.07 20: 4 0.48 1.33 0.88 1.46 22: 0 0.04 0.14 0.09 0.08 22: 1 0.03 0.02 0.03 0.29 24: 0 0.02 0.03 0.12 24: 1 0.10 0.14 0.02 total 2.59 6.36 6.82 7.09 ン酸
08:
0),オレイン酸08:
1), リノール 酸08:
2)さらにアラキドン酸 (20:4)が検 出され,少量ではあるが3TC
,6
0
分のincubateに よって脂肪酸の細胞外遊出が認められた.Fig.
5
はDB-cAMP前 処 理AMrtの 遠 心 上 清液中の
HE
から検出した脂肪酸分析の図であるFig. 4 Gas Chromatographic Analysis of Heptane Extracts from Supernatants of Alveolar Macrophages 同 @F F@P O H @ -F 守口口白 1005
15 口 co 同 @ F m o ω d w O N Fig. 5 Gas Chromatographic Analysis of Heptane Extracts from Supernatants of cAMP-treated Alveolar Macrophages F @ -F O @ -r C パ w o u 寸 0 . 刷 円 ∞ -F 何 . @ - F -∞ F O 句 ﹄ ﹁ ぜ U O N Fig.6 Gas Chromatographic Analysis of Heptane Extrcts from Supernatants of cGMP-treated Alveolar Macrophages が, controlの Fig.4の区!と殆んど変化が認めら れなかった. 8Br-cGMP前処理AMftの遠心上清液からの
HE
中の脂肪酸分析の図は Fig.6
に示した.前の Fig. 4, 5にはみられなかったラウリル酸(12: 0), ミリスチン酸04:
0),パルミトレイン酸 (16:1), さらにリノレン酸(18:3)の peakを 認め,前二図でみられたパルミチン酸 (16: 0), ステアリン酸08:
0),オレイン酸08:
1)さ らにアラキドン駿(20: 4)の peakが高くなって いるのが認められる.すなわち ,AMftにo
z
を添 加しなくとも cGMP前処理によって,各種長鎖脂 肪酸の細胞外放出が顕著に認められた.3
.
気管支肺胞洗浄液 (broncoalveolarlavage suide, BALF)のHE
中の脂肪酸分析 AMφ は外部からの刺激により細胞外に多量の 生物活性物質や,長鎖脂肪酸を放出することがわ かったので,細胞外に放出された生物活性物質を 含有する BALFからHE
を得て,脂肪酸分析を 行った.すなわち,無処理の正常ウサギから得た normal-BALFと CFAを静注 3週間後のウサギ から得た CFA-BALFに つ い て 脂 肪 酸 分 析 を 行 なった.その成績は Table6に示した.表にみられ るように nomal-BALFのHE
中の総脂肪酸量に ~1006o o H 。 qH ~ ~ ~ "寸 門 H ∞ ﹂ ︻ Nu ∞叶 叶"∞﹂[ 0 ・ ∞ 同 Fig.7 Gas Chromatographic Analysis of heptane Extracts from CFA-BALF ω H 。 " 。 叶 0 4 守 什 。 叶 刊 で ・ . 一 ∞ ∞ ∞ z ~~ Fig.8 Gas chromatographic analysis of Heptane巴xtractsfrom normal-BALF 。 N 叶"日 Y 。 N l ︻ m H 、D ~ ~ ~ 比 べ てCFA-BALFのHE中 の 総 脂 肪 酸 量 は 約 7倍量検出された.このときの gaschromatogra-phyによる脂肪酸分析の図はFig.7,8の通りであ る.Fig. 7はCFA-BALFの脂肪酸分析の図であ り,カプリン酸(10: 0),ミリスチン酸
04:
0), パルミトレイン酸(16:1),ステアリン酸(18 0),オレイン酸(18:1), リノール酸(18: 2) さらにアラキドン酸 (20: 4)の各脂肪酸が検出 され, Fig.8のnormal-BALFにおける脂肪酸分 ﹂門 O N q o N 叶 内 向 析の図ではFig.7での各脂肪酸は検出されるが, 各 脂 肪 酸 量 はCFA-BALF~こ比べて normal BALF中では少量であることが判明する、 4. normal AMφの各種生物活性物質ならび に 長 鎖 指 肪 酸 の 細 胞 外 放 出 に 及 ぼ す normal-BALFおよびCFA-BALFの影響 従来の実験は,正常あるいは活性化したAMφ を組織培養液中に浮遊させて実験を進めてきた. しかし実際生体内においては, AMφ はFig.7,-1007-(a)
。
・
一
一
一
一
0 _ _ _ 0 (b) (C) 〔 E 100 、 cl コ ω 相 p h ω H C O U-一/
。
50 E。
ー
一
一
一
。
一
一
一
一
。
ム 岳 也 / 一 一 / 一 一 / 一 -A U 企 企 , -/ / 一 / f L ム , K 口 〆 〆 〆 〆 〆 J 口 / 〆 〆 〆 〆 〆 口 hNO の h J-.
・ーー一一一ー掴ト--3'0' 90' 180' 3'。目 9'0' 180' 3'0' 9'0' 180' CFA-BAlF + normal-BAlF + ヲ ι O A ψ A ψ ・ M M A 門 A m-a a m m r r
。 。
+ n n 将 司 司 m E 一一 H 5・
r。
- 0 normal-AM,中OZゐ 6 .normal..AM,ゆOZ・
ー
・
normal-AM骨 企___....normal-AMや 事。~O CFA-BAlF &---&事normal-BAlF
Fig, 9 Lysozyme contents of Supernatants from OZ-ingested normal-AM<tin
the presence of CFA-and normal BALF
E二コwithoz 90 min,
圃
・
・
withoutOZ J 川 副 A B ・ 卜 -4a A m F r c。
n normal-BAlF + normal-AMゆ AV M S 哨 同日+副 E m u H r o n Dil 2 5Fig.10 Bactericidal activities of supernatants from OZ-ingested normal-AMφ in the presence of CFA-and normal-BALF 8でみられたような多量な長鎖脂肪酸類あるいは AMφから細胞外にreleaseした生物活性を有す る物質等を多量に含有する(分泌液〉肺胞被覆層 内に存在し,肺胞被覆層と密接な関係を保ちなが ら,感染防御の役を果していると考えられる.そ こで, より生体内に近い環境,条件下とするため に, normal-BALFお よ びCFA-BALFにAMφ
を浮遊させて, BALFのAMφ におよぼす影響を
検討した.その成績のいくつかを示す.
1)リゾチーム含有量に及ぼすBALFの影響
Table 6 Identification and Quantification of Fatty Acids in Heptane Extracts from Normal -and CF A -BALF (μg/2mO Fatty acids normal-BALF CFA-BALF 10: 0 3.61 ( 0.8) 23.00 ( 0.8) l2:0 1.61 ( 0.4) 0.48 ( 0.01) 14: 0 9.75 ( 2.4) 8.24 ( 0.3) 16: 0 123. 14 (30.1) 643.84 (22.0) 16: 1 31.25 ( 7.6) 178.36 ( 6.1) 18: 0 28.28 ( 6.9) 211.17 ( 7.2) 18: 1 85.76 (21.0) 934.73 (31.9) 18: 2 86.18 (21.1) 716.06 (24.4) 18: 3 12.79 ( 3.1) 110.73 ( 3.8) 20: 0 trace 2.00 ( 0.06) 20: 1 1.05 ( 0.3) 4.24 ( 0.1) 20: 4 21.56 ( 5.3) 79.42 ( 2.7) 22: 0 0.88 ( 0.2) 3.93 ( 0.1) 22: 1 2.11 ( 0.5) 8.74 ( 0.3) 24: 0 0.83 ( 0.2) 9.00 ( 0.3) 24: 1 trac巴 5.00 ( 0.2) Tota! 408.80 2930.70 ( ): % 健常ウサギからBALFをを得て,遠心した沈溢 の 細 胞 はHBSSで2回 洗 浄 し てnormalAMtt とし,遠心上清液はnormal-BALFとして実験に 供した.またCFA静注ウサギから注射3週間後 に,同様の操作でCFA-AMφ とCFA-BALFと を得た.normal AMttを1
x
1W cells/mlになるよ うにnormal-BALF,CFA-BALFさ ら にHBSS に浮遊し,それぞれを5mlずつシャーレに分注し, 一部にOZを添加して ,3TC, 30分, 90分, 180分 とCO2-incubatorに保持後, 5, OOOrpm 20分間遠 心した.その遠心上清液についてリゾチームの含 有 量 をOssermanら18)の方法に準拠して測定し た.その成療を示したのがFig.9である.図に示 すようにnormalAMφ をCFA-BALFに浮遊さ せた場合(a)の遠心上清液中にはCFA-BALF単 独で'-44μg/ml含有していたリゾチームの量は,処 置30分後で、はすで、に75μg/mlと増量し, OZを添 加 す る と , そ の 上 清 液 中 に は110μg/mlとリゾ チーム含有量は更に増大していた.(b)はnormal AMφ とnormal-BALFの組み合せであるが, normal-BALF単独でも42μg/mlと, リゾチーム -1009 含有量はCFA-BALFと同程度に検出された.し かし OZの添加によっても180分後で,上清液中の リゾチーム量は67μg/mlであり,リゾチームの細 胞外放出にはCFA-BALFの方が強く影響を及ぼ していることがわかる.図の(c)はHBSSにnor -mal AMφ を浮遊させた場合で、あるが,処置30分 後に10-15μg/mlとリゾチームの細胞外放出が 僅かにみられ, OZの添加により180分後で、50μg/ mlのりゾチーム含有量を検出した.即ち,normal AMttでも, OZの貧食により,僅かではあるがり ゾチームを細胞外に放出することがわかる. 2)殺菌作用に及ぼすBALFの影響 前 実 験 系 と 同 じ 被 検 液 に つ い て ,L
i
steria monocytogenesを被検菌として行った殺菌作用 の成績はFig.10に示す通りである.CFA-BALF に浮遊したAMφはincubate30分後に,上清液 中に2倍希釈液で65%,10倍希釈液でも40%の殺 菌率を示し,殺菌性物質の放出がみられており, さらに経時的に上清液中への殺菌性物質の放出は 増大していた.この細胞外放出はOZの添加によ り更に増大がみられている. normal-BALFにAMttを浮遊した場合は, OZ の添加により,殺菌性物質の上清液中への放出は みられるが, OZを添加しない場合の放出は僅か であった. 3) normaI AMφ からの脂肪酸の放出に及ぼ すBALFの影響 normal AMttを1X 107 cells/mlになるように CFA-BALF, normal-BALFさ ら にHBSSに 浮 遊させて,夫々にOZを添加しincubate120分, 180分後の遠心上清液から HEを得て, HE中の脂 肪酸分析を行った.その成績はTable7に示す通 りである.表にみられるようにnormalAMφを CFA-BALF に浮遊させ,保持2時間後に上清液 からのHE中に検出された総脂肪酸量は500μg/2 ml, 3時間後で、400μg/2mlであり,これはTable 6に 示 し たCFA-BALF単 独 の 総 脂 肪 酸 量 の 17.4%および13.5%しか検出されていない.また 各種脂肪酸の分析により incubate後にパルミチ ン酸 06: 0), オレイン酸 08:1), リノール 酸 (18: 2), リノレン酸08: 3)の減少が目立Table 7 Identification and Quantification of Fatty Acids in Heptane Extracts from N ormal.AMφin the Presence of Normal.and CFA.BALF
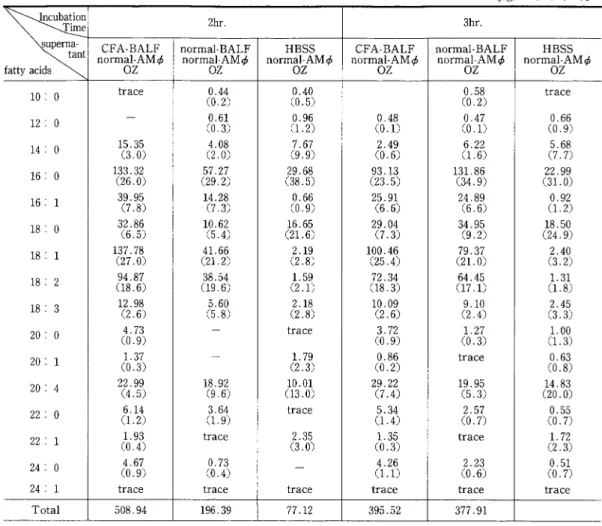
(μg/2m]) ( ):% 2hr. 3hr.
CFA.BALF normal.BALF HBSS CFA.BALF normal.BALF HBSS
決
normal.AM1> normal.AM1> normal.AM1> normal.AM1> normal.AM1> normal.AM1>oz oz 10: 0 trace (00..424) 12: 0 (00..631) 14: 0 1(53..305) (42..008) 16: 0 1(3236..302) (5279..227) 16: 1 3(97..985) 1(47..238) 18: 0 3(26..856) 1(05..642) 18: 1 l(3277..708) (4121.. 266) 18: 2 (9148..867) (3189..564) 18: 3 1(22..968) (55..680) 20: 0 (40..793) 20: 1 (1.0.337) 20: 4 2(24..959) 1(89 6.92) 22: 0 (61..124) (31..694) 22: 1 (1.0 9.43) trace 24: 0 (40..697) (00..743) 24: 1 trace trace Total 508.94 196.39 ち,逆にアラキドン酸
(
2
0
:
4),ベヘン酸(
2
2:
0), リグノセリン酸 (24: 0)の含有率は増大し ていた.とくにアラキドン酸(
2
0
:
4)はincubate 2時間後より 3時間後の方が更に含有率は大で あった.一 方normalAMrtをnormal-BALFに 浮 遊 し,normalAMゆからの脂肪酸の放出を検討する と, incubate
2
時間後に総脂肪酸量は約2
0
0
μ
g
/
2
mlとnormal-BALF単独の総脂肪酸量の48%し か検出されていないが 3時 間 後 に は normal-BALF単独の総脂肪酸量に近づいていた. 考 察 生体内に侵入した細菌や有毒物質に対する防御 oz 0.40 (0.5) 0.96 (1.2) 7.67 (9.9) 29.68 (38.5) 0.66 (0.9) 16.65 (21.6) 2.19 (2.8) 1. 59 (2.1) 2.18 (2.8) trace 1. 79 (2.3) 10.01 Cl3.0) trace 2.35 (3.0) trace 77.12 1010 oz oz oz 0.58 trace (0.2) 0.48 0.47 0.66 (0.1) (0.1) (0.9) 2.49 6.22 5.68 (0.6) 0.6) (7.7) 93.13 131. 86 22.99 (23.5) (34.9) (31. 0) 25.91 24.89 0.92 (6.6) (6.6) 0.2) 29.04 34.95 18.50 (7.3) (9.2) (24.9) 100.46 79.37 2.40 (25.4) (21. 0) (3.2) 72.34 64.45 1.31 08.3) (17.1) (1.8) 10.09 9.10 2.45 (2.6) (2.4) (3.3) 3.72 1.27 1.00 (0.9) (0.3) 0.3) 0.86 trace 0.63 (0.2) (0.8) 29.22 19.95 14.83 (7.4) (5.3) (20.0) 5.34 2.57 0.55 (1.4) (0.7) (0.7) 1. 35 trace 1.72 (0.3) (2.3) 4.26 2.23 0.51 (1.1) (0.6) (0.7) trace trace trace 395.52 377.91 機構のーっとして食細胞による殺菌機構があげら れ,食細胞による異物のとり込み,消化,殺菌, 排除と一連の流れにより生体の恒常性が保たれて いる.この感染初期に生体防御として働く食細胞 による殺菌機構に関してはKlebanoffl)-3)の提唱 した酸素依存性の殺菌系と酸素非依存性の殺菌系 の二つの系がよく知られ,その後多くの研究者に よる報告があることは前に述べた.しかし,生体 内に侵入した徴生物が,すべてこの系によって殺 菌 さ れ る と は 考 え ら れ ず , 著 者 ら12)-13)は Klebano妊の唱えた殺菌系の他に,責食時に食細 胞内の lysosome酵素活性が允進し,その酵素の 一つである phospholipaseA2によって,細胞膜に存在する phospholipidsから分解遊出してきた殺 菌性のある長鎖脂肪酸もまた食細胞の殺菌機構に 関 与 す る 重 要 な 系 の 一 つ で あ る こ と を 報 告 し た11)12) すなわちCorynebacteriumanaerobium CPropionibacterin acunes)投与ウサギから得た AMctあるいはComplete Freund's Adjuvant CCFA)投与ウサギから採取したAMctのlysate からのheptaneextracts CHE)について,各種細 菌に対する殺菌作用と長鎖脂肪酸の定性と定量分 析を行った.その結果, authenticな長鎖脂肪酸の 純試薬が各種細菌とくにグラム陽性菌に強L、殺菌 活性を示すこと,またphospholipaseA2 CPLase A2) 活性の阻害作用をもっindomethacin(IM), あるいは脂肪酸の殺菌作用を中和する血清アルブ ミ ンCBSA)を添加して,殺菌作用を検討し, Mct 由来の長鎖脂肪酸と殺菌作用との聞に密接な関係 のあることが判明したのである. これらの成績からMφなどの食細胞の侵入菌 に対する殺菌機構として,従来から知られていた 酸素に依存する系と,依存しない系の他に,貧食 時にlysosome酵素の一つで、あるphospholipases によって,細胞膜のphospholipidsが分解して生 じる長鎖脂肪酸も食細胞の殺菌機構に関与し,生 体の感染防御機構に重要な役割を果していると思 惟した. さらに,炎症のmediatorとしての役割をもっ prostaglandinsのprecursorで あ る arachidonic acid (C20: 4)が異物を貧食し,活性化したMφ から多量に放出されることは注目されるところで あり, この面からも長鎖脂肪酸が感染防御に重要 な役を演じていることが思惟される. 今回はmononuclearphagocytesの中でも,生 体内で解剖学的に特殊な場に存在するAMctに ついて,感染防御に関しとくに殺菌機構の面から 検討した.AMφは好気的環境下にあって酸化的 代謝機構が発達し, とくに酸素依存性の殺菌機構 が優位に立つことは当然考えられる.AMctはま た異物の貧食により細胞内の代謝は克進して,活 性酸素も多量に産生され多くの細菌は消化,殺菌 されるが, この殺菌系では殺菌されない細胞寄生 菌(本実験ではその中の1菌種Listeri amonocy-togenesを被検菌とした〉はAMct由来の長鎖脂 肪酸が主体となって殺菌されることが判明した. 1958年Sutharlandに よ る cyclicAMPの 発 見, 1963年にAshmanらによるcyclicGMPの発 見により,生体内における cyclicnucleotidesの 機能に注目が集まり,多くの研究が行われ報告さ れている.それによってcAMPとcGMPとは互 に桔抗しながら作用して生命現象をcontrolして いることが明らかになってきた.そこで食細胞由 来の殺菌性のある長鎖脂肪酸の細胞外放出機構に おいても cyclicnucleotidesが関与するであろう と思惟して実験を行った.cyclic nucleotidesは phosphodiesteraseの 作 用 に よ り 容 易 に5'-AMP または5'-GMPとなるため, cyclic nucleotidesと しての機能が失われるので, phosphodiesterase の作用を受けにくく,かっ細胞内への透過性の高 い8bromo-cGMP C8Br-cGMP)とdibutyryl -cAMP CDB--cAMP)を用いた. cyclic nucleotidesでPMNを処理することに よって lysosome酵素の細胞外放出は影響され, cGMP前処理で促進され,cAMP前処理によって 阻害されることは報告されており11ト18) 川 村19)は PMNからの lysosome由来の殺菌性物質の細胞 外放出がcyclicnucleotidesにより調節されてい ると報告している.さらにPMNのみならず石橋 ら川』土ウサギの血小板から耐熱性殺菌性物質 (β -lysinまたはplakin)の細胞外放出にcylic nu -cleotidesが関与していることを報告した.また内 山28)は正常およびCorynebacteriumanaerobium 死菌体投与ウサギから得た肺胞および腹腔Mφ の殺菌性物質とその放出機構を検討し,ここでも cyclic nucleotidesにより調節されていることを 報告している. 吉岡ら20)は責食時にみられる食細胞の殺菌系の 一つの系である食細胞由来の長鎖脂肪酸の細胞外 放出機構にも cyclicnucleotidesが関与している ことを明らかにした.すなわちFig.2, 3, Table 4, 5さらにFig_4, 5, 6~こ示すように AMφ から遊 出する殺菌性物質ならびに長鎖脂肪酸はAMct を8Br-cGMPで前処理することによって増大し, DB陶cAMPの前処理によって抑制されていた. -1011ー
これらの成績から,貧食時に
A M
ゆから細胞外 に遊出してきた長鎖脂肪酸は,同時に放出されたl
y
s
o
s
o
m
e
由来の酵素や殺菌性物質と相侯って働 き,生体の感染防御における食細胞の殺菌機構に その役割を果し,この殺菌機構にはc
y
c
l
i
c n
u
-c
l
e
o
t
i
d
e
s
が関与して,これを調節していることが 判明した. 生体内における解剖学的位置からしてもAMct
は外界と常に接触があるために,絶えず外からの 刺激があり,ある程度活性化された状態におかれ ている.これは感染防御の面から見て当然のこと と考えられる.さらに,ある程度活性化されたAMct
の 周 回 や 肺 胞 腔 内 にl
y
s
o
s
o
m
e
酵素,l
y
s
o
s
o
m
e
由来の殺菌性物質さらに殺菌的に働く 長鎖脂肪酸を含有する分泌液(BALF)
の存在する ことは,感染防御の面から見ても,その意義は大 きい.また肺胞被覆層にはII型肺胞上皮細胞が合 成し分泌された肺表面活性物質が存在している. この中に含有されているリン脂質も当然AMct
に何らかの影響を及ぼしていると考えられる.R
e
y
n
o
l
d
'
s
2
1lは気管支洗浄法をはじめて臨床の 病態解析に応用した.これによって洗浄液成分に ついて各種免疫グロプリンや補体活性を検討し, 洗浄によって得られた細胞の分類をも,健康者と 患者の病巣反対側の肺胞洗浄法によって比較検討 している. わが国でも,螺良,安藤,安岡23)-問らその他多 くの報告がみられている. 木 文 のF
i
g
_ 9
,1
0
に 示 し た よ う にn
o
r
m
a
l
網AMct
をnormal-BALF
あるいはCFA-BALF
に浮遊させ,
normal-AMφ
に及ぼすBALF
の影響 を観察すると,normal-AMct
をnormal-BALF
に浮遊させた場合よりもCFA-BALF
に浮遊させ た方が,より早く,且多量にl
y
s
o
s
o
m
e
酵素,殺菌 性物質,l
y
s
o
z
y
m
e
を細胞外に放出することが判 明した.一方,T
a
b
l
e
6,F
i
g
.
7にみられたCFA-BALF
中に多量に含有され,検出された長鎖脂肪 酸はT
a
b
l
e7
でみられるようにnormal-AMct
を 浮遊させることにより速やかに細胞内にとり込ま れ,著しい減少を示した.かかる現象はn
o
r
m
a
l
-BALF
にnormal-AMφ
を浮遊させた場合には脂 肪酸量は一過性にやや減少を示すものの,すぐ回 復し殆んど変化がなかった.S
c
h
r
o
i
t
ら2
町工Mφ
の物理的性状を操作するた め に , 外 部 か ら 供 給 し た 脂 肪 酸 , と く にp
a
l
-m
i
t
o
l
e
i
c
a
c
i
d
(16: 1),o
l
e
i
c
a
c
i
d
(
c
i
s
, 18: 1),e
l
a
i
d
i
c
a
c
i
d
(
t
r
a
n
s,
18: 1),
l
i
n
o
l
e
i
c
a
c
i
d
(18 :2
,)l
i
n
o
l
e
n
i
c
a
c
i
d
(18
:
3
)
,さらにa
r
a
c
h
i
d
o
n
i
c
a
c
i
d
(20 : 4)などの不飽和脂肪酸を細胞内に容易 に と り 込 ん で , こ れ に よ っ てMφ
のp
h
o
s
-p
h
o
l
i
p
i
d
s
のf
a
t
t
y
a
c
y
lc
o
m
p
o
s
i
t
i
o
n
を選択的に変 えることを報告している.今回の実験によっても, 明らかにAMφ
の外部に存在する脂肪酸が,細胞 内にとり込まれているので, とり込まれた脂肪酸 が細胞内の脂肪酸の構成にし、かなる変化をもたら しているのか興味のある処であり目下実験中であ る. おわりに 食細胞の殺菌機構について, とくにComplete
F
r
e
u
n
d
'
s
Adjubant (CF
A)
注射ウサギから肺胞Mct
と 気 管 支 肺 胞 洗 浄 液(
b
r
o
n
c
b
o
a
l
v
e
o
l
a
r
l
a
v
a
g
e
f
l
u
i
d
s
:
BALF)
とを得て,AMφ
から細胞 外に放出されるl
y
s
o
s
o
m
e
酵素,殺菌性物質,長鎖 脂肪酸について検討し,さらにc
y
c
l
i
cn
u
c
l
e
o
t
i
d
e
s
がこれら生物活性物質の細胞外放出に及ぼす影響 を,またこれら生物活性物質を含有するBALF
も 併せ検討して,肺内感染防御に果すAMφ
ならび にBALF
の役割について,次の結論を得た. 1)CFA
活 性 化AMφ
は異物責食時にl
y
s
o
s
o
-me
酵素,殺菌性物質さらに殺菌性のある長鎖脂 肪酸を多量に細胞外に放出する. 2) これら生物活性物質の細胞外放出機構にはc
y
c
l
i
c
n
u
c
l
e
o
t
i
d
e
s
が関与し,cAMP
前 処 理 に よって抑制され,cGMP
前処理によって増大して いた.さらにcGMP
前 処 理AMφ
にo
p
s
o
n
i
z
e
d
zymosan (OZ)
を添加すると, これら生物活性物 質の細胞外放出は著しく増大した.とくに,長鎖 脂肪酸においてp
a
l
m
i
t
i
c a
c
i
d
(16
:
0
)
,s
t
e
a
l
i
c
a
c
i
d
(18
:
0
)
,o
l
e
i
c
a
c
i
d
(18
:
1)さらにa
r
a
-c
h
i
d
o
n
i
c
a
c
i
d
(20 :4
)
の細胞外放出は有意に増大 した.これに反しcAMP
前 処 理AMφ
にOZ
を 貧食させても,これら生物活性物質の細胞外放出-1012-は抑制されていた.
3)BALF中にはlysosome酵素, lysozyme,殺 菌性物質さらに長鎖脂肪酸が含有されており,と くにnormal-BALFに比べてCF
A
-
BALF中にこ れら物質の含有量は顕著に増加していた.また, 生 物 活 性 物 質 を 多 量 に 含 有 す るCFA-BALFに normal-AMφを 浮 遊 さ せ る と CFA-BALF中の 長鎖脂肪酸は急激に減少を示した. 以上の成績からAMφからのlysosome酵素, 殺 菌 性 物 質 さ ら に 長 鎖 脂 肪 酸 の 細 胞 外 放 出 に BALFが影響を及ぼしていることが判明した. AMφ が肺感染の防御機構を充分に発揮するた め に は あ る 程 度 活 性 化 さ れ たA M4>の存在と同 時 にA Mゆ か ら 遊 出 さ れ た 種 々 の 生 物 活 性 物 質 を含有する分泌液がAMφ の周りに,肺胞腔内に 存在しAMφ の機能に影響を及ぼしている. 一方,気道を介し大量の細菌が侵入したり異物 の侵入によって AMφの活性化が更に進み,細胞 内の代謝が允進して活性酸素の産生の増大,種々 の生物活性物質の細胞外放出も増大されると,分 泌液中に放出された生物活性物質はさらにA M4> を活性化していくこととなり,生体にとっては必 ずしも有利にのみ働くとは考えられず,むしろ肺 組織に障害性に働いて逆に疾病を惹起したり,増 悪したりしていくことも当然考えられる. 今後はさらにBALFの生化学的性状,組成など を 検 討 し , 肺 感 染 時 に お い て 感 染 防 御 に 果 す A M4>と BALFの役割を追求したい. 稿を終るに当り,長年御指導を頂いた故相津憲教 授,故山口康夫教授,現在御指導を頂いている微生物 学教室小野魁教授に深謝し,併せて共同研究者である 夫,石橋昭に感謝する. 本研究の一部は,岡本糸校研究助成によった.昭和 25年に東京女子医専を卒業し,以来感染と抵抗性を テーマとして,とくに殺菌機構の面から続けてきたさ さやかな研究を,今回,改めてふり返る機会を得たこ とを感謝している. 文 献 1)区lebano宜,S.J.: Role of the superoxide anion in the myeloperoxidase mediated antimicrobial system. J Biol Chem 249 3724 -3728(1974) 1013 2) Klebanoff, S.J.: Antimicrobial systems of the polymorphonuclear leukocyte. In the Phagocytic Cell in Host Resistance (ed. Bal lanti and Dayton), Raven Press, New York (1975) pp. 45-59 3) Klebanoff, S.J. and Rosen, H.: Ethylene for -mation by polymorphonuclear leukocytes. Role of myeloperoxidase. J Exp Med 148 490-506 (1978) 4) Nishimura, M.: Studies of antibacterial activity of lysosomesI.Bactericidal activities of lysosomes. Nihon Univ J Med 14 203-230 (1972)5) Maejima, Y.: Studies of antibacterial activ -ity of lysosomes. II. E妊ectof endotoxin and antilysosomal membrane serum administration on serum bactericidal activities. Nihon Univ J Med 15 329-352 (1973)
6) Ishibashi, A., Ishibashi, T., Oami, S., et al. : Studies of antibacterial activity of lysosomes III. Release of bactericidal and lysosomal en -zyme activities into plasma from whole blood expos巴dto Ehrlich ascites tumor cells. N ihon Univ J Med 16 241-263 (1974) 7)Oami
,
S.: Studies of antibacterial activity of lysosomes. IV. Effect of cortisone on release of lysosomal enzymes and bactericidins. N ihon Univ J Med 17 139 -164 (1975) 8) Ishibashi, A., Ishibashi, T., Oami, S., et al. : Studies of antibacterial Activity of Lysosomes. V. The In Vivo and in Vitro E任ectsof Ehrlich Ascites Tumor Cells on Release of Lysosomal Bactericidins and Enzymes. Nihon Univ J Med 17 165 -184 (1975)9) Otani
,
A.: Studies of Antibacterial Activity of Lysosomes. VI.E妊巴ct of Complement Components on Release of Lysosomal Bacter icidins and Enzymes from Cytochalasin B -treated Polymorphonuciear Leukocyt巴sexpos巴dto Undigestible Particles. Nihon Univ
J Med 18 267 -290 (1976) 10)石橋昭・石橋悌子・蛭問哲雄・他血小板由来 殺菌性物質の放出機構に関する研究.日細菌誌 34731-743 (1976) 11)石橋悌子・石橋昭・鈴木八重子・他・マグロ ファージのFunctionalheterogeneityに関する 研究 Corynebacteriumanaerobium死 菌 体 投 与によるマクロフア ジの非特異的殺菌能増強作 用に関する研究 .日大医誌 39789-797(1980) 12)角田国義・石橋悌子・石橋昭 マクロファージ 由来の長鎖脂肪酸の殺菌作用に関する研究.日大 医誌 41 919-930 (1982)
13) 内藤敏雄・石橋悌子・石橋 昭・他'食細胞由来 の 殺 菌 性 長 鎖 脂 肪 酸 の 放 出 に 及 ぼ すindometh -acinお よ びcytochalasinBの 影 響 . 日 大 医 誌 41 1173 -1183 (1982)
14) Myrvik, Q.N., Leake, E.S. and Faris, B.: Studies of pulmonary alveolar macrophages from normal rabbit. A technique to procure them in a high state of purity.J Immunol 86 128-136 (1961) 15) Kochan, 1., Pellis, E.R. and Pfohl, D.G.: E任ectsof normal and activated cell fraction on the growth of tubercule bacilli. Infect Immun 6 142 -148 (1972) 16) Sutherland, E Fractionation and characterization of a cyclic adeni泊neribonucle抗otid巴邸sformed by tis鎚sueparti. cles.
J
Bio Chem 232 1077 -1091 (1957) 17)Ashman, D.F., Lipton, R., Melicow, M.M., et al.: Isolation of adenosine 3',5' .monophos -phate and guanosine 3',5'-monophosphate from rat urine. Biochem Biophys Commun 11 330 -334 (1963)18) Osserman
,
E.F. and Lawler,
D.P.: Serum and urinary lysozyme in monocytic and monomyelocytic leukemia.J Exp Med 124 921-956 (1966)19) Kawamura
,
S.: Studies of antibacterial activity of Iysosomes. VII. Effect of comple ment components, cyclic nucleotid巴5and relat-ed agents on extracellular release of lysosome -derived bactericidins. Nihon Univ J Med 20
一 1014 87 -106 (1978) 20)吉岡広晶・漆原 彰 , 山 内 典 明 ・ 他 " ク ロ フ ァ ー ジ 由 来 の 殺 菌 性 長 鎖 脂 肪 酸 の 放 出 機 構 に 及 ぼ す cyclic nucleotidesの 影 響 . 日 大 医 誌 441-9 (1985)
21)Reynolds, H.Y., Fulmer, J.D., Kazmierowski, J.A., et al.: Analysis of cellular and protein content of bronchoalveolar lavage fluid from patients with idiopathic pulmonary fibrosis and chronic hypersensitivity pneumonitis.J Clin Invest 59 165 -175 (1977) 22) Schroit, A.J. and Gallily, R.: Macrophage fatty acid composition and phagocytosis. E妊ect of unsaturation on cellular phagocytic activity. Immunol 36 199-205 (1979) 23)螺 良 英 郎 ・ 肺 と 免 疫 の 問 題 点 , 呼 吸 1 63-67 (1982) 24)螺 良 英 郎 ・ 安 岡 勧 , 尾 崎 敏 夫 ・ 他 Bron -choalveolar lavageに 関 す る 研 究 . 日 胸 疾 会 誌 19 792-800 (1981) 25)安 藤 正 幸 肺 胞 マ ク ロ フ ァ ー ジ . 呼 吸 2 645 -649 (1983) 26)安藤正幸:肺における生体防御の特殊性.生体防 御 217-26(1985) 27)安岡 劫,河野知弘・大串文雄・他:肺胞マクロ フ ァ ー ジ の 生 化 学 的 背 景 . 呼 吸 2813-822 (1983) 28)内山秀彦 マクロファージのFunctionalHeter -ogeneityに 関 す る 研 究 とくに肺胞および腹腔 マクロファーシの殺菌性物質とその放出機構につ いて . 日大医誌 39 151-161 (1980)