2 略 語 CA:Carbonic anhydrase LC50: 半 数 致 死 濃 度 EC50: 半 数 影 響 濃 度 TPO: 甲 状 腺 ペ ル オ キ シ ダ ー ゼ EZA: エ ト キ シ ゾ ル ア ミ ド AZA: ア セ タ ゾ ル ア ミ ド DZA: ド ル ゾ ル ア ミ ド AR: ア ン ド ロ ゲ ン 受 容 体 PTU: プ ロ ピ ル チ オ ウ レ ア MMI: メ ル カ プ ト メ チ ル イ ミ ダ ゾ ー ル dpf:days-post fertilization hpf: hours-post fertilization MO: Morpholino oligonucleotide PBS:Phosphate buffered saline
4 諸 言 近 年 、 産 業 の 発 展 に 伴 い 環 境 に 多 く の 化 学 物 質 が 使 用 さ れ て お り 、 そ の う ち 人 工 的 に 作 ら れ た 化 学 物 質 は 世 界 全 体 で 現 在 十 万 種 以 上 と も 言 わ れ て い る 。 多 種 多 様 な 化 学 物 質 が 人 々 の 生 活 を 支 え る 反 面 、 大 半 は 環 境 や ヒ ト へ の 安 全 性 が 全 て 明 ら か に な っ て い な い た め 世 界 的 な リ ス ク 管 理 が 進 め ら れ て い る 。 日 本 国 内 に お け る 化 学 物 質 の リ ス ク 管 理 は 、 生 体 内 へ の 残 留 性 や 長 期 毒 性 を 有 す る 物 質 の 製 造 お よ び 使 用 に 制 限 が 設 け ら れ て い る 。 新 規 化 学 物 質 お よ び 一 定 の 生 産 量 を 超 え る 化 学 物 質 に 対 し て は 化 学 物 質 の 生 産 量 、 環 境 中 へ の 排 出 量 を 把 握 、 届 け 出 が 必 要 と な る と と も に 、 ヒ ト や 環 境 生 物 へ の リ ス ク 評 価 が 義 務 付 け ら れ て い る 。 農 薬 の よ う に 環 境 中 で 使 用 さ れ 、 河 川 や 湖 沼 へ 流 入 し 環 境 へ の リ ス ク が 想 定 さ れ る 化 学 物 質 は 、 環 境 生 物 へ の 影 響 評 価 が 必 須 と さ れ て い る 。 そ の 結 果 、 生 態 系 に 影 響 を 及 ぼ す 可 能 性 が あ る 農 薬 は 、 使 用 量 や 使 用 回 数 の 制 限 が 講 じ ら れ 生 物 へ の 影 響 を 与 え な い よ う な 規 制 が な さ れ る (Hayasaka et al., 2013; Komoike et al., 2016)。
5
6 第 一 章 ゼ ブ ラ フ ィ ッ シ ュ 胚 発 生 過 程 に 影 響 を 及 ぼ す 化 学 物 質 の 影 響 比 較 背 景 細 胞 や 組 織 、 生 体 の 恒 常 性 維 持 は 細 胞 分 裂 や 細 胞 新 生 、 組 織 形 成 が 活 発 化 す る 胚 発 達 期 に お い て 重 要 で あ る 。 細 胞 や 組 織 は 発 生 初 期 か ら イ オ ン 輸 送 体 や 酵 素 等 が 発 現 し 細 胞 内 の イ オ ン 濃 度 や pH を 制 御 す る こ と で 恒 常 性 が 維 持 さ れ る 。 化 学 物 質 に よ り 制 御 機 能 が 破 た ん す る と 正 常 な 細 胞 分 裂 や 分 化 を 阻 害 さ れ 、 重 篤 な 場 合 に は 細 胞 死 や 奇 形 を 引 き 起 こ さ れ る(Scott et al., 1990)。 一 方 で 、 生 体 と し て の 恒 常 性 維 持 は 神 経 系 と 内 分 泌 系 に よ り 制 御 さ れ て い る 。 神 経 系 に よ る 生 体 制 御 は 、 脳 お よ び 脊 髄 か ら な る 中 枢 神 経 系 と 中 枢 神 経 か ら 延 び る 末 梢 神 経 に 大 別 さ れ 、 ホ ル モ ン や 神 経 伝 達 物 質 と し て 働 く 神 経 ペ プ チ ド を 合 成 ・ 分 泌 し 、 内 分 泌 系 の 調 節 な ど 様 々 な 生 理 作 用 の 調 節 も 行 っ て い る 。 内 分 泌 系 は 甲 状 腺 、 副 腎 、 す い 臓 、 卵 巣 、 精 巣 等 の 分 泌 細 胞 が 集 合 し た 腺 組 織 で あ り 、 内 外 か ら の 刺 激 に よ り ホ ル モ ン を 合 成 し 、 標 的 臓 器 へ 分 泌 す る 。 成 長 過 程 で は 神 経 系 お よ び 内 分 泌 系 の 機 能 が 活 発 化 し て い る た め 、 こ れ ら 機 能 へ の 化 学 物 質 の 影 響 は 、 恒 常 性 機 能 異 常 や か く 乱 作 用 を 引 き 起 こ し 分 化 や 形 態 形 成 に 影 響 を 与 え る こ と が 懸 念 さ れ る(van Gelder et al., 2010)。
そ こ で 、 上 記 に 示 し た 組 織 や 生 体 の 恒 常 性 を 司 る 酵 素 や ホ ル モ ン 等 を 阻 害 す る 化 合 物 を ゼ ブ ラ フ ィ ッ シ ュ 胚 に 暴 露 し 、 胚 発 達 過 程 で の 化 学 物 質 の リ ス ク 把 握 を 行 っ た 。 先 ず 、恒 常 性 維 持 に 関 わ る 酵 素 で あ る 炭 酸 脱 水 酵 素(CA)を 阻 害 す る 化 合 物 を 選 択 し 評 価 対 象 と し た 。CA 阻 害 剤 は 、哺 乳 類 の 胎 児 に 対 し 細 胞 内 pH 制 御 機 能 を 破 た ん さ せ 骨 形 成 異 常 や 心 臓 奇 形 を 誘 発 す る こ と が 報 告 さ れ て い る(Scott et al., 1990)。CA は 魚 類 の 初 期 胚 に お い て も 多 数 の 組 織 に 局 在 し 、 器 官 や 臓 器 形 成 や 発 達 に 大 き く 貢 献 す る 酵 素 で あ る(Aspatwar, 2013; Lin et al., 2008; Liao et al., 2009; Ito et al., 2013)。
8 方 法 使 用 生 物 (独 )理 化 学 研 究 所 か ら 分 譲 さ れ た 5~ 10 か 月 齢 の 雌 雄 の ゼ ブ ラ フ ィ ッ シ ュ (RIKEN WT, Danio rerio) を 用 い た 。 飼 育 条 件 飼 育 に は 流 水 式 水 槽(名 東 水 園 )を 用 い 、 水 温 28.5 ℃ 、 光 条 件 (明 14 時 間 、 暗 10 時 間)に 雌 雄 別 に 飼 育 し 、 1 日 4 回 粉 末 飼 料 、 う ち 2 回 は 雌 の み ブ ラ イ ン シ ュ リ ン プ も 給 餌 し た 。 受 精 卵 採 取 受 精 卵 を 採 取 す る 前 日 夕 方 に 雌 雄 の ゼ ブ ラ フ ィ ッ シ ュ を 交 配 さ せ 、 翌 朝 受 精 卵 を 得 た 。 得 ら れ た 受 精 卵 は シ ャ ー レ に 移 し 10%ク ロ ラ ミ ン T 含 有 0.3×Danieau’s
solution [17.4 mM NaCl, 0.21 mM KCl, 0.12 mM MgSO4, 0.18 mM Ca(NO3)2, 1.5
mM HEPES pH 7.2; Gustafson et al., 2012]に て 消 毒 し た 後 、 0.3×Danieau’s solution で 3 回 洗 浄 し た 。 洗 浄 後 28℃ で 受 精 後 6 時 間 ま で イ ン キ ュ ベ ー ト し た 。
化 合 物 暴 露
12 考 察 魚 類 の 胚 発 達 過 程 に 対 し て 特 異 的 な 影 響 を 与 え る 化 学 物 質 を 検 索 す る た め 、 胚 発 達 ・ 成 長 に 必 要 な タ ン パ ク や ホ ル モ ン に 影 響 を 及 ぼ す と 想 定 し た 化 合 物 を ゼ ブ ラ フ ィ ッ シ ュ 胚 に 暴 露 し た 。 評 価 し た CA 阻 害 剤 、 性 ホ ル モ ン 産 生 阻 害 剤 、 甲 状 腺 ホ ル モ ン 産 生 阻 害 剤 の う ち 、 最 も 低 濃 度 ま で ゼ ブ ラ フ ィ ッ シ ュ 胚 発 生 に 影 響 を 及 ぼ し た 化 合 物 は CA 阻 害 剤 の EZA で あ り 、 EC50で あ る 0.08ppm に お い て 耳 石 低 形 成 や 遊 泳 不 良 が 生 じ た (Table
13
17
29
Fig. 4 ロキサピンによるゼブラフィッシュ胚 への影 響
ロキサピン 1.3ppm ヒレ低 形 成
32 方 法 使 用 生 物 (独 )理 化 学 研 究 所 か ら 分 譲 さ れ た 5~ 10 か 月 齢 の 雌 雄 の ゼ ブ ラ フ ィ ッ シ ュ (RIKEN WT, Danio rerio) を 用 い た 。 飼 育 条 件 飼 育 に は 流 水 式 水 槽(名 東 水 園 )を 用 い 、 水 温 28.5 ℃ 、 光 条 件 (明 14 時 間 、 暗 10 時 間)に 雌 雄 別 に 飼 育 し 、 1 日 4 回 粉 末 飼 料 、 う ち 2 回 は 雌 の み ブ ラ イ ン シ ュ リ ン プ も 給 餌 し た 。 受 精 卵 採 取 受 精 卵 を 採 取 す る 前 日 夕 方 に 雌 雄 の ゼ ブ ラ フ ィ ッ シ ュ を 交 配 さ せ 、 翌 朝 受 精 卵 を 得 た 。 得 ら れ た 受 精 卵 は シ ャ ー レ に 移 し 10%ク ロ ラ ミ ン T 含 有 0.3×Danieau’s
solution [17.4 mM NaCl, 0.21 mM KCl, 0.12 mM MgSO4, 0.18 mM Ca(NO3)2, 1.5
33 耳 石 の Calsein 染 色 各 発 生 ス テ ー ジ の 胚(1, 2, 3, 5 dpf)を 0.2% Calcein 水 溶 液 に て 30 分 間 染 色 し 、 蒸 留 水 で 洗 浄 し 、 ト リ カ イ ン 過 麻 酔 で 安 楽 死 さ せ 、 耳 石 を 摘 出 し た 。 摘 出 し た 耳 石 は 蛍 光 顕 微 鏡(Leika DM2500)に て 観 察 し た 。 耳 石 面 積 の 測 定 各 発 生 ス テ ー ジ の 胚(1, 2, 3, 5 dpf)を ト リ カ イ ン で 麻 酔 し 、 実 体 顕 微 鏡 (M125, Leica) 下 で 耳 石 面 積 を 測 定 し た 。 面 積 測 定 に は ImageJ software (version 1.49, National Institutes of Health)を 用 い た 。 組 織 標 本 作 成 各 発 生 ス テ ー ジ の 胚(1, 2, 3, 5 dpf)を ト リ カ イ ン 過 麻 酔 で 安 楽 死 さ せ 、 10%中 性 ホ ル マ リ ン 緩 衝 液 に て 1 週 間 程 度 固 定 さ せ た 。 常 法 に 従 い 、 パ ラ フ ィ ン 切 片 (厚 さ 3μm)を 作 製 し 、HE 染 色 、 von KOSSA 染 色 も し く は 免 疫 組 織 化 学 染 色 を 施 し 、 光 学 顕 微 鏡 に て 観 察 し た 。 Morpholino oligonucleotide の マ イ ク ロ イ ン ジ ェ ク シ ョ ン
Ito ら の 方 法 に 従 い 、 CA2a お よ び CA15a に 対 す る Antisense Morpholino
35 結 果
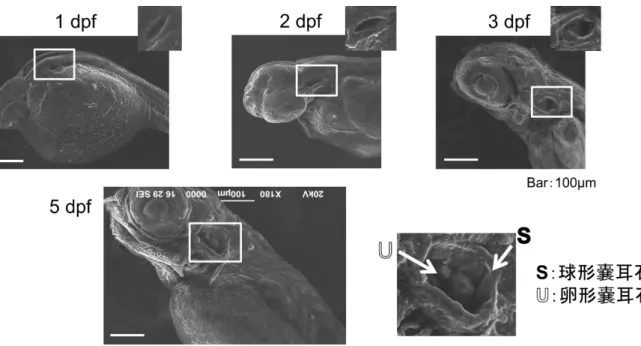
ゼ ブ ラ フ ィ ッ シ ュ 胚 に お け る 内 耳 お よ び 耳 石 の 発 達
耳 石 は 聴 覚 お よ び 平 衡 感 覚 を 司 る 器 官 で あ り 、 魚 類 で 発 生 学 的 に 最 も 初 期 に 形 成 さ れ る 硬 組 織 で あ る こ と が 知 ら れ て い る(Radtke and Dean, 1982; Murayama et al., 2002; Inoue et al., 2013)。 ゼ ブ ラ フ ィ ッ シ ュ の 内 耳 形 成 は 受 精 後 1 日 よ り み ら れ 、 耳 胞 と な る く ぼ み が 形 成 さ れ 、 そ の 中 に 2 つ の 耳 石 (卵 形 嚢 耳 石 、 球 形 嚢 耳 石 )が 観 察 さ れ た 。 孵 化 が み ら れ る 受 精 後 3 日 に は 、 耳 胞 部 に 嚢 状 構 造 が 観 察 さ れ 、 球 形 嚢 耳 石 は 卵 形 嚢 耳 石 に 比 べ 大 き く 成 長 し た(Fig. 1-3)。 ま た 耳 胞 部 の 組 織 切 片 を 作 製 し 詳 細 に 内 耳 を 観 察 し た と こ ろ 、 受 精 後 1 日 で は 内 耳 の 原 基 と な る 耳 胞 部 に く ぼ み(内 腔 )が 観 察 さ れ 、 内 耳 上 皮 細 胞 層 上 部 に は 耳 石 を 形 成 す る 多 数 の 微 細 な 粒 子 が 確 認 さ れ た 。 受 精 後 2 日 に は 、 粒 子 が 集 合 、 凝 集 し 部 分 的 に カ ル シ ウ ム 沈 着 が 確 認 さ れ た 。 受 精 後 3 日 は 、 内 腔 が 拡 張 し 嚢 状 構 造 が 形 成 さ れ た 。 耳 石 は KOSSA 染 色 に よ り 外 層 か ら 石 灰 化 し て い る よ う に 観 察 さ れ た 。 受 精 後 5 日 で は 耳 胞 の 内 腔 が さ ら に 拡 張 し 、 内 柱 の 伸 長 も み ら れ た (Fig. 4)。 耳 石 も 成 長 し カ ル シ ウ ム 濃 度 は 受 精 後 3 日 に 比 べ 1.7 倍 増 加 し 、 耳 石 石 灰 化 が 急 速 に 進 ん で い る こ と が 確 認 さ れ た(Fig. 5, 6)。 ゼ ブ ラ フ ィ ッ シ ュ 胚 の 内 耳 発 達 過 程 で の 炭 酸 脱 水 酵 素(CA)の 発 現 と 局 在 魚 類 の 耳 石 の 主 成 分 は 炭 酸 カ ル シ ウ ム で あ り 、 卵 形 嚢 、 球 形 嚢 と よ ば れ る 袋 状 の 器 官 内 の 中 に 浮 遊 し て い る 。 袋 は 内 リ ン パ 液 で 満 た さ れ て お り 、 外 界 か ら の イ オ ン 供 給 に よ り 重 炭 酸 イ オ ン お よ び カ ル シ ウ ム イ オ ン か ら 結 晶 化 さ れ る こ と が 知 ら れ て い る 。 内 リ ン パ 液 へ の イ オ ン 輸 送 は 、 内 耳 上 皮 細 胞 イ オ ノ サ イ ト に 局 在 す る 炭 酸 脱 水 酵 素(CA)や Cl-/HCO3-transporter を 介 し 供 給 さ れ る (Tohse et al., 2001)。 ゼ ブ ラ フ ィ
36
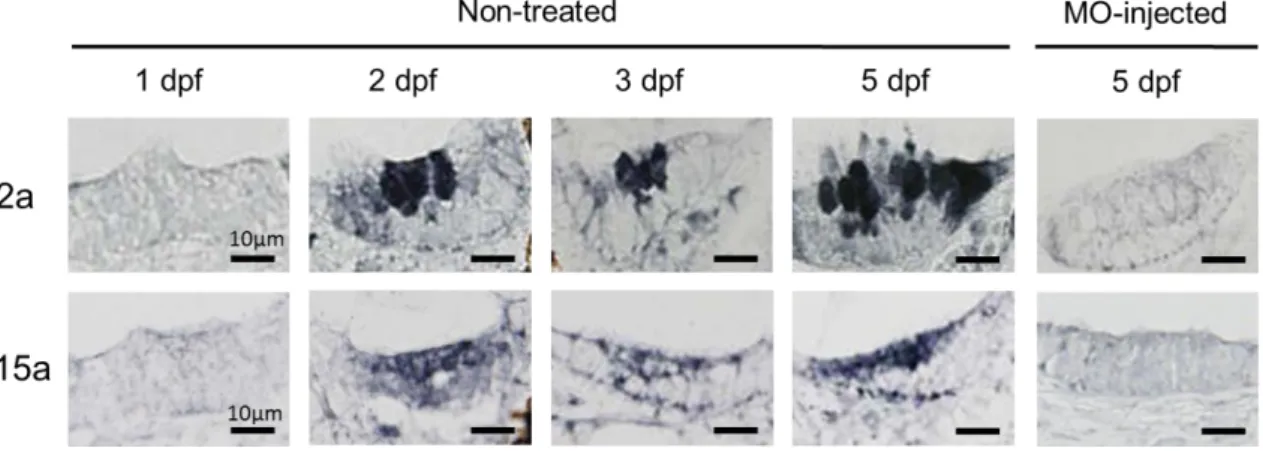
37 考 察 ゼ ブ ラ フ ィ ッ シ ュ 胚 の 耳 石 形 成 は 受 精 後 1 日 よ り 始 ま り 、 耳 胞 と な る く ぼ み が 形 成 さ れ 2 つ の 耳 石 (卵 形 嚢 耳 石 、 球 形 嚢 耳 石 )も 観 察 さ れ た 。 2 日 目 に は 耳 石 に カ ル シ ウ ム 沈 着 が 確 認 さ れ 、 そ の 後 孵 化 以 降 急 速 に 石 灰 化 が 進 行 し た(Fig. 1-4)。 ま た 耳 石 石 灰 化 開 始 と 同 時 期 に 内 耳 上 皮 細 胞 に は CA2a お よ び 15a の 発 現 が み ら れ て お り 、 こ れ ら CA の 発 現 が 内 リ ン パ 液 へ の 重 炭 酸 イ オ ン 供 給 に 寄 与 し 石 灰 化 を 進 行 さ せ て い る と 推 察 さ れ た(Fig.5-7 )。 細 胞 質 タ ン パ ク で あ る CA2a は 内 耳 上 皮 細 胞 の 有 毛 細 胞 全 体 、 お よ び イ オ ノ サ イ ト に 発 現 し 、 膜 タ ン パ ク で あ る CA15a は 有 毛 細 胞 の 頂 端 部 お よ び 底 部 、 お よ び イ オ ノ サ イ ト に 局 在 し て い た(Fig. 7)。 魚 類 の 内 耳 に お け る CA の 局 在 に つ い て の 報 告 は 限 ら れ て い る 。 先 行 研 究 で は サ ー モ ン や ニ ジ マ ス の 成 魚 、 Cichild fish 胚 で は イ オ ノ サ イ ト に CA の 発 現 が 確 認 さ れ て い る が 、 有 毛 細 胞 に CA は 局 在 し て い な い(Tohse et al., 2006, Mayer-Gostan et al., 1996)。 し か し な が ら 、 ゼ ブ ラ フ ィ ッ シ ュ 成 魚 に は CA が 有 毛 細 胞 に 発 現 し て お り (Shiao et al., 2005)、 CA の 局 在 は 種 間 差 が あ る 可 能 性 が 示 唆 さ れ て い る 。 本 研 究 で は ゼ ブ ラ フ ィ ッ シ ュ の 内 耳 有 毛 細 胞 に CA が 発 現 す る こ と を 示 し た 先 行 研 究 を 支 持 し 、 さ ら に CA2a お よ び 15a と い う ア イ ソ フ ォ ー ム を 確 認 し た 初 め て の 報 告 で あ る 。 マ ウ ス の 胎 児 お よ び 新 生 児 の 内 耳 上 皮 有 毛 細 胞 に は CA2 お よ び CA11 が 局 在 す る こ と が 確 認 さ れ て お り(Wu et al., 2012)、 音 を 感 知 す る 際 に 繊 毛 先 端 部 で の イ オ ン 輸 送 や 細 胞 内 酸 塩 基 平 衡 に 関 与 す る と 報 告 さ れ て い る(Wu et al., 2012; Okamura et al., 1996)。 ゼ ブ ラ フ ィ ッ シ ュ で 有 毛 細 胞 に 確 認 さ れ た CA2a は 哺 乳 類 有 毛 細 胞 に 存 在 す る CA と 相 同 性 が 高 く 、 ゼ ブ ラ フ ィ ッ シ ュ の 聴 覚 や 平 衡 覚 の 機 能 制 御 に 関 与 し て い る 可 能 性 が あ る と 考 え ら れ た 。 今 後 は CA の 機 能 お よ び CA を 介 し 産 生 さ れ る HCO3-を 輸 送 す る Cl-/HCO3-交 換 体 、HCO3-ATPase 等 と の 関 わ り 含 め 詳 細 に 解 析 を
38
Fig. 1 ゼブラフィッシュ胚 および内 耳 の発 達
39
Fig. 2 走 査 型 電 子 顕 微 鏡 によるゼブラフィッシュ胚 の観 察
42
Fig. 5 ゼブラフィッシュ胚 から単 離 した耳 石 と Calcein 染 色
43
Fig. 6 ゼブラフィッシュ胚 発 生 過 程 における耳 石 中 のカルシウム濃 度
44
Fig. 7 ゼブラフィッシュ胚 発 生 過 程 における CA2a および CA15a の発 現
45
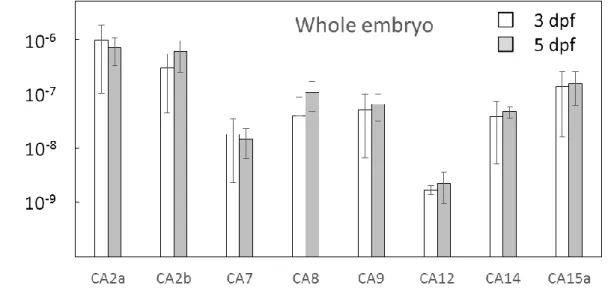
Fig. 8 ゼブラフィッシュ胚 に発 現 する CA アイソフォーム mRNA 発 現 量
46
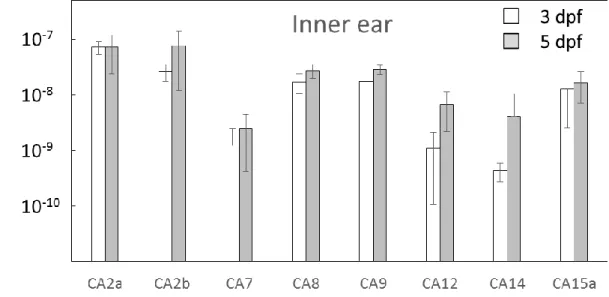
Fig. 9 ゼブラフィッシュ内 耳 に発 現 する CA アイソフォーム mRNA 発 現 量
47
第 三 章 炭 酸 脱 水 酵 素 阻 害 剤 が ゼ ブ ラ フ ィ ッ シ ュ 胚 発 達 に 及 ぼ す 影 響
背 景
48 方 法 使 用 生 物 (独 )理 化 学 研 究 所 か ら 分 譲 さ れ た 5~ 10 か 月 齢 の 雌 雄 の ゼ ブ ラ フ ィ ッ シ ュ (RIKEN WT, Danio rerio) を 用 い た 。 飼 育 条 件 飼 育 に は 流 水 式 水 槽(名 東 水 園 )を 用 い 、 水 温 28.5 ℃ 、 光 条 件 (明 14 時 間 、 暗 10 時 間)に 雌 雄 別 に 飼 育 し 、 1 日 4 回 粉 末 飼 料 、 う ち 2 回 は 雌 の み ブ ラ イ ン シ ュ リ ン プ も 給 餌 し た 。 被 験 物 質 本 試 験 に 用 い た 化 合 物 は エ ト キ シ ゾ ル ア ミ ド(EZA, シ グ マ ア ル ド リ ッ チ )、 ア セ タ ゾ ル ア ミ ド(AZA, シ グ マ ア ル ド リ ッ チ )、 ド ル ゾ ラ ミ ド (DZA, 東 京 化 成 )を 用 い た 。 受 精 卵 採 取 受 精 卵 を 採 取 す る 前 日 夕 方 に 雌 雄 の ゼ ブ ラ フ ィ ッ シ ュ を 交 配 さ せ 、 翌 朝 受 精 卵 を 得 た 。 得 ら れ た 受 精 卵 は シ ャ ー レ に 移 し 10%ク ロ ラ ミ ン T 含 有 0.3×Danieau’s
solution [17.4 mM NaCl, 0.21 mM KCl, 0.12 mM MgSO4, 0.18 mM Ca(NO3)2, 1.5
mM HEPES pH 7.2; Gustafson et al., 2012]に て 消 毒 し た 後 、 0.3×Danieau’s solution で 3 回 洗 浄 し た 。 洗 浄 後 28℃ で 受 精 後 6 時 間 ま で イ ン キ ュ ベ ー ト し た 。
化 合 物 暴 露
49 遊 泳 行 動 の 評 価
対 照 お よ び 化 合 物 を 受 精 後 6 時 間 か ら 5 日 目 ま で 暴 露 し た 稚 魚 を 光 照 射 下 で 遊 泳 行 動 量 の 評 価 を 行 っ た 。 行 動 量 は 1 分 間 ×3 回 、 実 体 顕 微 鏡 (M125, Leica)に 接 続 し た カ メ ラ(DP71, オ リ ン パ ス 光 学 工 業 )に て 撮 影 し 、 解 析 に は ImageJ software (version 1.49, National Institutes of Health)を 用 い 、 総 移 動 距 離 を 算 出 し た 。
成 魚 へ の 化 合 物 暴 露
50 耳 石 中 カ ル シ ウ ム 濃 度 の 測 定 対 照 お よ び EZA を 受 精 後 6 時 間 か ら 5 日 目 ま で 暴 露 し た 稚 魚 を ト リ カ イ ン 過 麻 酔 で 安 楽 死 さ せ 、 耳 石 を 単 離 し た 。 単 離 し た 耳 石 に 1N 塩 酸 を 加 え 溶 解 さ せ 、 8N KOH に て pH12~ 13 に 調 整 し た 。 NN 指 示 薬 (同 仁 化 学 )を 数 滴 滴 下 後 EDTA に て 滴 定 し 、 カ ル シ ウ ム 濃 度 を 求 め た 。 ア ク リ ジ ン オ レ ン ジ / エ チ ジ ウ ム ブ ロ マ イ ド 染 色 対 照 お よ び EZA を 受 精 後 6 時 間 か ら 5 日 目 ま で 暴 露 し た 稚 魚 を ア ク リ ジ ン オ レ ン ジ と エ チ ジ ウ ム ブ ロ マ イ ド を 各 5μ g/mL 含 む 0.3×Danieau’s solution で 20 分 間 イ ン キ ュ ベ ー ト さ せ た 。0.3×Danieau’s solution で 洗 浄 し た 後 、 ト リ カ イ ン 過 麻 酔 で 安 楽 死 さ せ 、 蛍 光 顕 微 鏡 に て 観 察 し た 。 有 毛 細 胞 の 染 色 対 照 お よ び EZA を 受 精 後 6 時 間 か ら 5 日 目 ま で 暴 露 し た 稚 魚 に 3 種 の 染 色 試 薬 に て 有 毛 細 胞 の 染 色 を 行 っ た 。DASPEI も し く は YO-PRO-1 (Thermo Fosher Scientific) は 3μ M を 含 む 0.3×Danieau’s solution で 15 分 間 、 FM1-43 (Thermo Fosher Scientific)は 3μ M を 含 む 0.3×Danieau’s solution で 30 秒 間 イ ン キ ュ ベ ー ト し 染 色 し た 。 各 染 色 を 施 し た 後 、 ト リ カ イ ン 過 麻 酔 で 安 楽 死 さ せ 、 蛍 光 顕 微 鏡 に て 観 察 し た 。Parvalbumin 抗 体 を 用 い た 染 色 は 、 稚 魚 を 4%パ ラ ホ ル ム ア ル デ ヒ ド 含 有 PBS に て 4℃ で 一 晩 固 定 さ せ 、 蒸 留 水 で 洗 浄 後 0.1%Triton お よ び 0.5%ヤ ギ 血 清 を 含 む PBS で ブ ロ ッ キ ン グ さ せ た 。 続 い て 、 抗 Parvalbumin 抗 体 (Merck Millipore, 1:500)を 1%ヤ ギ 血 清 含 有 PBS に て 4℃ で 一 晩 反 応 さ せ た 。 PBS に て 洗 浄 後 、 二 次 抗 体 (Alexa Fluor 488, Life Technologies, 1:500)に て 反 応 さ せ 、 蛍 光 顕 微 鏡 下 で 観 察 し た 。
有 毛 細 胞 数 の 計 測
DASPEI に て 有 毛 細 胞 を 染 色 し た サ ン プ ル に つ い て Neuromast 中 の 有 毛 細 胞 数 を 蛍 光 顕 微 鏡 下 で 計 測 し た 。 計 測 対 象 と し た Neuromast は PO, SO1-3, O, OC, D1-2, M, IO1-3, OP, P1-2 と し た 。
Real-time PCR 法 に よ る 細 胞 死 に 関 連 す る 遺 伝 子 の 定 量
51
52 結 果
53 ろ 、EZA 暴 露 に よ り 受 精 後 3 日 で は 約 50%、 受 精 後 5 日 で は 約 40%ま で 低 下 し て い る こ と が 明 ら か と な っ た(Fig.4)。 EZA に よ る ゼ ブ ラ フ ィ ッ シ ュ 胚 の pH 変 動 お よ び 細 胞 死 の 検 出 CA 阻 害 剤 は 細 胞 内 の pH を 上 昇 さ せ 、 そ の 結 果 と し て 細 胞 や 組 織 、 個 体 の ホ メ オ ス タ シ ス が 破 た ん さ せ 、 石 灰 化 や 骨 化 、 器 官 ・ 組 織 形 成 不 全 を 引 き 起 こ す と 言 わ れ て い る(Postel et al.,2012)。EZA を 暴 露 し た ゼ ブ ラ フ ィ ッ シ ュ 胚 の 体 表 pH は EZA 暴 露 に よ り 濃 度 相 関 的 に 体 表 pH が 上 昇 し て い る こ と が 確 認 さ れ (Fig. 5)、 イ オ ン 輸 送 や pH 維 持 に 影 響 を 及 ぼ し て い る 可 能 性 が 示 唆 さ れ た 。 内 耳 は イ オ ン 輸 送 体 や そ れ に 関 わ る 酵 素 が 多 く 存 在 し て い る た め 、 内 リ ン パ 液 の pH は 変 動 を 受 け や す い 。 さ ら に 、 内 リ ン パ 液 の pH 変 動 は 耳 石 形 成 の み な ら ず 有 毛 細 胞 に 対 し て 影 響 が 生 じ る 可 能 性 が あ る と 報 告 さ れ て い る (Stawiki et al., 2014) 。 そ こ で EZA に よ り 有 毛 細 胞 へ 傷 害 が 起 こ っ て い る か を 確 認 す る た め に HE 染 色 で 組 織 学 的 変 化 を 観 察 し 、 さ ら に ア ク リ ジ ン オ レ ン ジ / エ チ ジ ウ ム ブ ロ マ イ ド 染 色 、TUNEL 染 色 を 施 し ア ポ ト ー シ ス を 検 出 し た(Fig. 6)。 EZA を 暴 露 し た 胚 の 内 耳 上 皮 細 胞 は 核 お よ び 細 胞 質 が 濃 縮 し 空 胞 化 が 生 じ 、 ア ポ ト ー シ ス 様 の 変 化 が み ら れ た 。 ア ク リ ジ ン オ レ ン ジ / エ チ ジ ウ ム ブ ロ マ イ ド 染 色 で は 内 耳 に 初 期 ア ポ ト ー シ ス を 示 す 緑 色 の シ グ ナ ル が 検 出 さ れ 、 胸 ビ レ に は オ レ ン ジ 色 の 後 期 ア ポ ト ー シ ス を 示 す シ グ ナ ル が 確 認 さ れ た 。TUNEL 染 色 で は 内 耳 上 皮 細 胞 、 特 に 支 持 細 胞 に TUNEL 陽 性 が 検 出 さ れ た 。 EZA に よ る 内 耳 有 毛 細 胞 傷 害 の 解 析 お よ び ア ポ ト ー シ ス 関 連 遺 伝 子 の 関 与 EZA に よ り 異 常 の み ら れ る 内 耳 に ア ポ ト ー シ ス が 検 出 さ れ た こ と か ら 、 有 毛 細 胞 へ の 影 響 の 有 無 を 調 べ る た め 蛍 光 色 素 に て 有 毛 細 胞 を 染 色 し 評 価 し た(Fig. 7)。 Mitochondrial 色 素 で あ る DASPEI 試 薬 に て Neuromast 中 の 有 毛 細 胞 を 染 色 し 、 有 毛 細 胞 数 を 計 測 し た 。 計 測 し た Neuromast は PO, SO1-3, O, OC, D1-2, M, IO1-3, OP, P1-3 の う ち 、 内 耳 に あ る Neuromast で あ る O(Otic hair cells)の み EZA に よ り 有 毛 細 胞 数 が 減 少 し て い る こ と が 示 さ れ た 。 さ ら に mechanotransduction channels の マ ー カ ー で あ る FM1-43FX や 抗 parvalbmin 抗 体 に お け る 染 色 に お い て も 内 耳 の 有 毛 細 胞 数 が 減 少 し て い る こ と も 明 ら か と な っ た 。
54
55 考 察 ス ル ホ ン ア ミ ド 系 CA 阻 害 剤 は ゼ ブ ラ フ ィ ッ シ ュ 胚 の 発 達 過 程 に お い て 耳 石 低 形 成 、 遊 泳 異 常 を 引 き 起 こ し 、 特 に 最 も 阻 害 作 用 の 強 い EZA は 0.08 ppm で も そ の 影 響 が み ら れ た 。 ス ル ホ ン ア ミ ド 系 CA 阻 害 剤 の ゼ ブ ラ フ ィ ッ シ ュ 胚 発 達 過 程 に お け る 影 響 差 を 比 較 し た と こ ろ 、 耳 石 形 成 や 遊 泳 行 動 に 対 す る 影 響 は EZA が AZA に 比 べ 350 倍 程 度 強 い と い う 結 果 と な っ た 。 ス ル ホ ン ア ミ ド 系 CA 阻 害 剤 は ス ル ホ ン ア ミ ド 側 鎖 や 脂 溶 性 が 薬 効 に 大 き く 寄 与 す る と い う 報 告 が あ る(Mohammed, 2013)。 ス ル ホ ン ア ミ ド 基 の pKa(pH7.4)は AZA で 9.1、EZA で 8.0 と 大 き な 差 は な い が 、 脂 質 分 配 係 数 は ア セ タ ゾ ル ア ミ ド の 0.001 に 比 べ エ ト キ シ ゾ ル ア ミ ド は 30 で あ り 脂 溶 性 が 非 常 に 高 く 、 こ の 特 徴 が ゼ ブ ラ フ ィ ッ シ ュ 胚 に 対 す る 影 響 差 に も 大 き く 関 与 し て い る の で は な い か と 考 え ら れ た 。 実 際 に 、 胚 中 へ の 取 り 込 み は 、 エ ト キ シ ゾ ル ア ミ ド が 暴 露 量 の 約 3.5 倍 胚 に 蓄 積 し て い た の に 対 し 、 ア セ タ ゾ ル ア ミ ド は 暴 露 量 の 0.0092 倍 し か 蓄 積 し て い な か っ た 。 こ の よ う な こ と か ら も AZA と EZA の 物 性 差 が ゼ ブ ラ フ ィ ッ シ ュ 胚 発 達 に 対 す る 感 受 性 差 が 生 じ た 要 因 と し て 考 え ら れ た 。 脂 溶 性 の 高 い 化 合 物 は 魚 類 の 体 内 に 蓄 積 し や す く 、 生 物 濃 縮 や 生 物 蓄 積 性 が 高 い と い う 観 点 か ら も 問 題 と な る 。 特 に 稚 魚 や 胚 は 組 織 や 器 官 の 発 達 が 不 十 分 で 免 疫 や 解 毒 機 能 が 十 分 に 発 達 し て い な い た め 、 化 学 物 質 へ の 感 受 性 が 高 い と の こ れ ま で の 見 解 を(Nagel, 2002; Russell et al., 1999; von Westernhagen, 1988, Braunbeck et al., 2005; Lammer et al., 2009; Seiler et al., 2014)、 本 研 究 結 果 は 実 証 す る デ ー タ と な っ た 。
CA は 内 耳 上 皮 細 胞 に 局 在 し 、 内 リ ン パ 液 へ の HCO3-供 給 に 寄 与 す る (Tohse et al.,
58
Fig. 2 スルホンアミド系 CA 阻 害 剤 によるゼブラフィッシュの遊 泳 行 動 量
CA 阻 害 剤 を暴 露 したゼブラフィッシュ稚 魚 (5 dpf)を光 照 射 下 で 1 分 間 の
総 移 動 距 離 を 測 定 し た 。* : p<0.05, ** :p<0.01, *** : p<0.001 (Dunnet
test)
**
EZA
AZA
DZA
59
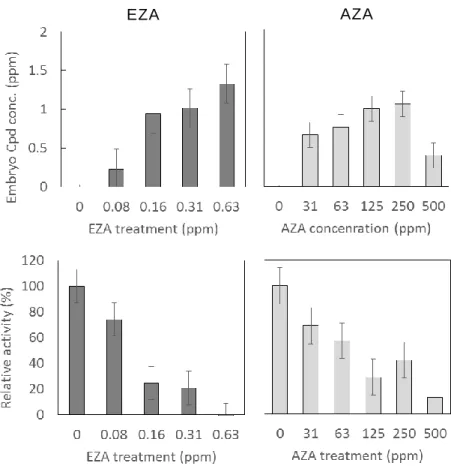
Fig. 3 スルホンアミド系 CA 阻 害 剤 による化 合 物 取 り込 み量 および酵 素 活 性
CA 阻 害 剤 を暴 露 したゼブラフィッシュ稚 魚 (5 dpf)の化 合 物 取 り込 み量 を測 定
した。また稚 魚 から抽 出 したサイトゾル画 分 の CA 酵 素 活 性 を測 定 した。
62
Fig. 6 EZA によるゼブラフィッシュ胚 のアポトーシス誘 導
(A) EZA 0.31ppm を暴 露 したゼブラフィッシュ(5dpf)を
ア クリ ジ ンオレンジ と エチジ ウム ブロマイド で染 色 し 、ア
ポト ーシ スを 検 出 し た。 胸 びれには 後 期 ア ポト ーシ スを
示 すオレンジ色 、内 耳 には初 期 アポトーシスを示 す緑 色
が核 にされた。bar=200μm。
(B) EZA 0.31ppm を暴 露 したゼブラフィッシュ(5dpf)の
内 耳 組 織 切 片 を作 成 し、HE 染 色 、TUNEL 染 色 、CA2a
抗 体 染 色 を 施 し た 。 bar=10μm 。 O : 耳 石 、 HC : 有 毛 細
胞 、SC:支 持 細 胞
64
Fig. 7 EZA によるゼブラフィッシュ胚 の内 耳 有 毛 細 胞 に対 する影 響
65
Fig. 8 EZA がゼブラフィッシュ胚 のアポトーシス関 連 遺 伝 子
および細 胞 周 期 に関 わる遺 伝 子 群 の発 現 に及 ぼす影 響
66 引 用 文 献
Amsterdam A, Nissen RM, Sun Z, Swindell EC, Farrington S, Hopkins N (2004) Identification of 315 genes essential for early zebrafish development. Proc Natl Acad Sci USA 101:12792–12797
Aspatwar A, Tolvanen MEE, Jokitalo E, Parikka M, Ortutay C, Harjula SKE, Rämet M, Vihinen M, Parkkila S (2013) Abnormal cerebellar development and ataxia in carp VIII morphant zebrafish. Hum Mol Genet 22:417–432
Beier M, Anken R (2006) On the role of carbonic anhydrase in the early phase of fish otolith mineralization. Adv Space Res 38:1119–1122
Beier M, Hilbig R, Anken R (2008) Histochemical localisation of carbonic anhydrase in the inner ear of developing cichlid fish, Oreochromis mossambicus. Adv Space Res 42: 1986–1994
Bian C, Zhao Y, Guo Q, Xiong Y, Cai W, Zhang J (2014) Aromatase inhibitor letrozole downregulates steroid receptor coactivator-1 in specific brain regions that primarily related to memory, neuroendocrine and integration. J Steroid Biochem Mol Biol 141:37-43
Brubaker KD, Mao F, Gay CV (1999) Localization of carbonic anhydrase in living osteoclasts with bodipy 558/568–modified acetazolamide, a thiadiazole carbonic anhydrase inhibitor. J Histochem Cytochem 47:545–550.
Braunbeck T, Böttcher M, Hollert H, Kosmehl T, Lammer E, Leist E, Rudolf M, Seitz N (2005) Towards an alternative for the acute fish LC50 test in chemical
assessment: the fish embryo toxicity test goes multi –species – an update. ALTEX 22:87–102
Coffin AB, Williamson KL, Mamiya A, Raible DW, Rubel EW (2013 ) Profiling drug– induced cell death pathways in the zebrafish lateral line. Apoptosis 18:393–408
67 Environ Pollut 4:65-152
Doğan S (2006) The in vitro effects of some pesticides on car bonic anhydrase activity of Oncorhynchus mykiss and Cyprinus carpio carpio fish. J Hazard Mater 132:171–176
Dukes M, Edwards PN, Large M, Smith IK, Boyle T (1996) The preclinical pharmacology of "Arimidex" (anastrozole; ZD1033) a potent, selective aromatase inhibitor. J Steroid Biochem Mol Biol 58: 439-45
Eastin WC Jr, Spaziani E (1978) On the mechanism of calcium secretions in the avian shell gland (uterus). Biol Reprod 19:505518.
Fushimi S, Wada N, Nohno T, Tomita M, Saijoh K, Sunami S, Katsuyama H (20 09) 17beta-Estradiol inhibits chondrogenesis in the skull development of zebrafish embryos. Aquat Toxicol 95:292-298
Gorelick DA, Watson W, Halpern ME (2008) Androgen receptor gene expression in the developing and adult zebrafish brain. Dev Dyn. 237 :298 7-95
Gustafson AL, Stedman DB, Ball J, Hillegass JM, Flood A, Zhang CX, Panzica -Kelly J, Cao J, Coburn A, Enright BP, Tornesi MB, Hetheridge M, Augustine – Rauch KA (2012) Inter–laboratory assessment of a harmonized zebrafish developmental toxicology assay –progress report on phase I. Reprod Toxicol 33:155 -164
He Y, Yu H, Sun S, Wang Y, Liu L, Chen Z, Li H (2013) Trans - 2 - phenylcyclopropylamine regulates zebrafish lateral line neuromast development mediated by depression of LSD1 activity. Int J Biol 57:363 - 373
68
Ito Y, Kobayashi S, Nakamura N, Miyagi H, Esaki M, Hoshijima K, Hirose S (2013) Close association of carbonic anhydrase (CA2a and CA15a), Na+/H+ exchanger
(Nhe3b), and ammonia transporter Rhcg1 in zebrafish ionocytes responsible for Na+ uptake. Front Phys 4:1–17
Inoue M, Tanimoto M, Yoichi O (2013) The role of ear stone size in hair cell acoustic sensory transduction. Sci Rep 3:2114
Komoike Y, Matsuoka M (2016) Application of Zebrafish Model to Environmental Toxicology. Nihon Eiseigaku Zasshi 71:227-235
Komori K, Suzuki Y (2009) Occurrence of pharmaceuticals and their environmental risk assessment of urban streams whose basins have different wastewater treatment conditions. Journal of Japan society on water environment 32:133 –138
Lammer E, Carr GJ, Wendler K, Rawlings JM, Belanger SE, Braunbeck T (2009) Is the fish embryo toxicity test (FET) with the zebrafish (Danio rerio) a potential alternative for the fish acute toxicity test? Comp Biochem Phys C 149:196 –209
Levine AJ, Oren M (2009) The first 30 years of p53: growing ever more complex. Nat Rev Cancer 9:749–758
Liao BK, Chen RD, Hwang PP (2009) Expression regulati on of Na+-K+–ATPase
alpha1–subunit subtypes in zebrafish gill ionocytes. Am J Physiol 296:R1897– R1906
Lin TY, Liao B, Horng J, Yan J, Hsiao C, Hwang P (2008) Carbonic anhydrase 2 -like a and 15a are involved in acid-base regulation and Na+ uptake in zebrafish
H+-ATPase-rich cells. Am J Physiol Cell Physiol 294:C1250–1260
Liu Z, Zuo J (2008) Cell cycle regulation in hair cell development and regeneration in the mouse cochlea. Cell Cycle 7:2129–2133
69
disruption of p27 (Kip1) allows cell proliferation in the postnatal and adult organ of corti. Proc Natl Acad Sci USA 96:4084–4088
Mantela J, Jiang Z, Ylikoski J, Fritzsch B, Zacksenhaus E, Pirvola U (2005 ) The retinoblastoma gene pathway regulates the postmitotic state of hair cells of the mouse inner ear. Development 132:2377–2388
Maren TH (1992) Direct measurements of the rate constants of sulfonamides with carbonic anhydrase. Mol Pharmacol 41:419–426
Maren TH, Conroy CW (1993) A new class of carbonic anhydrase inhibitor. J Biol Chem 268:26233–26239
Mayer–Gostan N, Kossmann H, Watrin A, Payan P, Boeuf G (1997 ) Distribution of ionocytes in the saccular epithelium of the inner ear of two teleosts (Oncorhynchus
mykiss and Scophthalmus maximus). Cell Tissue Res 289:53–61
Miller DS, Kinter WB, Peakall DB (1976) Enzymatic basis for DDE -induced eggshell thinning in a sensitive bird. Nature 259:122 -124
Miyachi S, Tsuzuki M, Avramova ST (1983) Utilization modes of inorganic carbon for photosynthesis in various species of Chlorera. Plant Cell Physiol 24 :441–51
Mohammed A Why are early life stages of aquatic organisms more sensitive to toxicants than adults? New insights into toxicology and drug testing. Chaper3:39 – 62
Mugiya Y (1977) Effect of acetazolamide on the otolith growth of goldfish. Bull Jpn Soc Sci Fish 43:1053–1058
Mugiya Y, Takahashi K (1985) Chemical properties of the saccular endolymph in the rainbow trout, Salmo gairdneri. Bull Fac Fish Hokkaido Univ 36:57–60
70
Nagel R (2002) DarT: the embryo test with the Zebrafish Danio rerio—A general model in ecotoxicology and toxicology. Altex 19:38–48
Nau H, Hauck RS, Ehlers K (1991) Valproic acid-induced neural tube defects in mouse and human: aspects of chirality, alternative drug development, pharmacokinetics and possible mechanisms. Pharmacol Toxicol 69:310 -321
Okamura HO, Sugai N, Suzuki K, Ohtani I (1996) Enzyme-histochemical localiza- tion of carbonic anhydrase in the inner ear of the guinea pig and several improvements of the technique. Histo- chem Cell Biol 106:425–430
Pietsch EC, Sykes SM, Mcmahon SB, Murphy ME (2008) The p53 family and programmed cell death. Oncogene 27:6507– 6521
Pocker Y, Beug WM, Ainardi VR (1971) Carbonic anhydrase interaction with DDT, DDE amd dieldrin. Science, 174:1336-1339
Radtke R, Dean J (1982) Increment formation in the otoliths of embryos, larvae and juveniles of the mummichog, Fundulus heteroclitus. Fish Bull US 80:201 –215
Ralston S, Miyamoto GT (1983) Analyzing the width of daily otolith increments to age the Hawaiian snapper, pristipomoides filamentosus. Fishery Bulletin 81:523– 535
Russell RW, Gobas FAPC, Haffner GD (1999) Maternal transfer and in ovo exposure of organochlorines in oviparous organisms: A model and field verification. Environ Sci Technol 33:416–420
Scott WJ, Duggan CA, Schreiner CM, Collins MD (1990) Reduction of embryonic intracellular pH: a potential mechanism of acetazolamide -induced limb
malformations. Toxicol Appl Pharmacol 103: 238 -254
71
Seiler TB, Best N, Fernqvist MM, Hercht H, Smith KEC, Braunbeck T, Mayer P, Hollert H (2014) PAH toxicity at aqueous solubility in the fish embryo test with
Danio rerio using passive dosing. Chemosphere 112:77–84
Shiao JC, Lin LY, Horng JL, Hwang PP, Kaneko T (2005) How can teleostean inner ear hair cells maintain the proper association with the accreting otolith? J Comp Neurol 488: 331–341
Shinohara C, Yamashita K, Matsuo T, Kitamura S, Kawano F (2007) Effects of Carbonic Anhydrase Inhibitor Acetazolamide (AZ) on Osteoclasts and Bone Structure. Journal of hard tissue biology 16:115–123
Stawicki TM, Owens KN, Linbo T, Reinhart KE, Rubel EW, Raible1 DW (2014) The zebrafish merovingian mutant reveals a role for pH regulation in hair cell toxicity and function. Dis Model Mech 7:847–856
Tohse H, Ando H, Mugiya Y (2004) Biochemical properties and immunohistochemical localization of carbonic anhydrase in the sacculus of the inner ear in the salmon Oncorhynchus masou. Comp Biochem Physiol A 137:87–94 Tohse H, Murayama E, Ohira T, Takagi Y, Nagasawa H (2006) Localization and diurnal variations of carbonic anhydrase mRNA expression in the inner ear of the rainbow trout Oncorhynchus mykiss. Comp Biochem Physiol B 145:257–264
Topal A, Atamanalp M, Oruc E, Demir Y, Beydemir S¸ Isık A (2014) In vivo changes in carbonic anhydrase activity and histopathology of gill and liver tissues after acute exposure to chlorpyrifos in rainbow trout. Arh Hig Rada Toksikol 65:377 – 385
Tsuzuki M, Miyachi S (1989) The Function of Carbonic anhydrase in aquatic photosynthesis. Aquat Bot 34: 85–104
72
van Gelder MM, van Rooij IA, Miller RK, Zielhuis GA, de Jong-van den Berg LT, Roeleveld N (2010) Teratogenic mechanisms of medical drugs. Hum Reprod Update 16 :378-94
von Westernhagen H (1988) Sublethal effects of pollutants on fish eggs and larvae. Fish physiology series 11. The physiology of developing fish Part A Eggs and Larvae San Diego: Academic Press 253–346.
Wang T, Wang J, Qiu J (1996) Observation on activity of carbonic anhydrase in the vestibule of guinea pigs. Zhonghua Er Bi Yan Hou Ke Za Zhi 31:24 –25