低酸素白血病細胞由来エクソソームによる
血管内皮細胞における血管新生増強に関する研究
田所 弘子
略語一覧
VEGF:vascular endothelial growth factor 血管内皮増殖因子 miRNA(miR):micro-ribonucleic acid
Exosomal miRNA:エクソソーム内に含まれる miRNA CML:chronic myeloid leukemia 慢性骨髄性白血病
FBS:fetal bovine serum ウシ胎児血清
PBS:phosphate-buffered saline リン酸緩衝食塩水
K562: 慢性骨髄性白血病細胞
SU-DHL-4: ヒトびまん性組織球性リンパ腫細胞株 RPMI-8226: 多発性骨髄腫細胞株
HUVECs:human umbilical vein endothelial cells ヒト臍帯静脈内皮細胞
EDTA:ethylenediaminetetraacetic acid エチレンジアミン四酢酸
NTAs: Nanoparticle tracking analysis
SDS:sodium dodecyl sulfate ドデシル硫酸ナトリウム
2-ME:2-mercaptoethanol 2-メルカプトエタノール
HIF:hypoxia-inducible factor 低酸素誘導因子
TLDA: TaqMan low-density miRNA array K5621%O2-exosome:低酸素条件下(O
2 1%)にて培養したK562 細胞より放出されたエクソ
ソーム
K56220%O2-exosome: 通常酸素条件下(O
2 20%)にて培養したK562 細胞より放出されたエ
クソソーム
TEM:transmission electron microscope 透過型電子顕微鏡 HUVECs1%O2: 低酸素条件下(O
2 20%)にて培養したHUVECs
HUVECs20%O2: 通常酸素条件下(O
2 20%)にて培養したHUVECs
K5621%O2/anti-miR-210: anti-miR-210 を遺伝子導入し、低酸素条件下(O
2 1%)にて 24
時間培養したK562 細胞
K5621%O2/anti-miR-210-exosome(K5621%O2/anti-miR-210-exo.): K5621%O2/anti-miR-210
より放出されたエクソソーム
K5621%O2/control: anti-miR-210 に対するコントロールを遺伝子導入し、低酸素条件
下(O2 1%)にて 24 時間培養したK562 細胞
K5621%O2/con-exosome(K5621%O2/con-exo.): K5621%O2/controlより放出されたエクソソー
ム
sensor vector: miR-210 と相補的な結合サイトと EFNA3 3’-UTR をふくむレポーター プラスミド
mutated sensor vector: sensor vector のコントロール
HUVECs20%O2/anti-miR-210: anti-miR-210 を遺伝子導入し、通常酸素条件下(O
2 20%)
にて 24 時間培養したHUVECs
HUVECs20%O2/ control: anti-miR-210 に対するコントロールを遺伝子導入し、通常酸素
条件下(O2 20%)にて 24 時間培養したHUVECs
RPMI-82261%O2-exosome: 低酸素条件下(O
2 1%)にて培養したRPMI-8226 細胞より放出さ
れたエクソソーム
RPMI-822620%O2-exosome: 通常酸素条件下(O
2 20%)にて培養したRPMI-8226 細胞より放
第一章 低酸素環境における白血病細胞由来エクソソームの血管新生増強作用 第一節 目的
現在、腫瘍研究では腫瘍微小環境に注目が集まっている。腫瘍微小環境の中でも低 酸素はとくに重要な要素の 1 つである。低酸素環境は、がん幹細胞の未分化性維持に 重要であり、血管新生を促進させる因子として知られている。また、慢性骨髄性白血 病 (CML: chronic myeloid leukemia)患者の骨髄では顕著な血管新生がみられるが骨
髄は低酸素環境であることが知られている。通常酸素状態で 慢性骨髄性白血病細 胞 (K562 細胞)が放出したエクソソームが血管内皮細胞を血管新生促進へと促すこと が報告されているが(20)、低酸素環境での白血病細胞由エクソソームの作用について は不明の点が多い。そこで、第 1 章では低酸素状態でK562 細胞から放出されたエク ソソームが血管内皮細胞の挙動に及ぼす影響を検討した。 第二節 方法 1. 培地および試薬の調製
RPMI1640 培地:RPMI Medium 1640 1x(Invitrogen, Carlsbad, CA)に 56℃で 30 分間非 動化した FBS を 10% (v/v)となるように添加し、さらに Penicillin を 10 万単位/L、 Streptomycin を 100 mg (力価)/L となるように添加し調製した。
EBM-2 培地:EBM-2 medium(EBM-2;Lonza Inc.)に EGM-2 singleQuots(Lonza Inc.)を 添加し調製した。
PBS:10xPBS 50mL に対し超純水 450mL 加え、オートクレーブ滅菌処理(121℃ 20min) した。
2. 細胞培養
慢性骨髄性白血病細胞株(K562 細胞)およびヒトびまん性組織球性リンパ腫細胞株 (SU-DHL-4 細胞) : RPMI1640 培地を用い、5% CO2 存在下にて 37℃で培養した。コン フルエントとなった細胞を回収し、12000rpmで 3 分間、室温で遠心分離した。遠心分 離後、上清を取り除きRPMI1640 培地を加え懸濁した。トリパンブルー染色による色素
排除法により生細胞数が 5×105 細胞/mLとなるように調整し、滅菌済培養フラスコ
(面積 25cm2)に 5mLずつ播種し培養した。
多発性骨髄腫細胞株(RPMI-8226 細胞): RPMI1640 培地を用い、5% CO2 存在下にて 37℃で培養した。コンフルエントとなった細胞をスクレーパーで回収し、12000rpmで 3 分間、室温で遠心分離した。遠心分離後、上清を取り除きRPMI1640 培地を加え懸濁
した。トリパンブルー染色による色素排除法により生細胞数が 5×105 細胞/mLとなる
処理し回収後、トリパンブルー染色による色素排除法により生細胞数が 2.2×104 細
胞/mLとなるように調整し、滅菌済培養 100mmcell culture dishに 10mLずつ播種し培 養した。
3. 低酸素培養
前処理として、窒素ガスによりチャンバー内の酸素濃度を調整することのできるマ
ルチガスインキュベーター(MCO-5M, Sanyo, Osaka, Japan )内にて 37℃で 5%CO2かつ
1%O2で 24 時間それぞれ培地の酸素濃度を調製した。これらの培地を用い、K562 細胞
とSU-DHL-4 細胞は 5×105細胞/mLの細胞懸濁液を 5mL、RPMI-8226 細胞は 5×105細胞
/mLの細胞懸濁液 10mL、また HUVECsは 2.2×104細胞/mLの細胞懸濁液を 10mLおのおの
調製して播種し、これらをC02 5%、02 1%の低酸素条件下 37℃で培養した。
4. 細胞増殖試験
K562 細胞を 5×104細胞/well、24 well dishで播種し、24、48、72 時間培養した。
培養後に細胞を回収し、トリパンブルー染色による色素排除法により生細胞数を測定 した。 5. エクソソームの回収法 細胞懸濁液を 3000×g で 15 分間、室温で遠心分離し、細胞と上清を採取した。細 胞上清 5mL に ExoQuick を 1mL 加え、十分に転倒置換し 4℃で 12 時間以上静置した。 その後、3000×g で 30 分間、室温で遠心分離し、軽く上清を取り除き、さらに 1500 ×g で 5 分間、室温で遠心分離し上清をすべて取り除いた。
6. Nanoparticle tracking analysis (NTAs)
NTA測定は、blue laser (405 nm)を搭載したNanosight LM10 system (Nanosight, Amesbury, UK)を用いて行った。エクソソームはレーザーを照射されブラウン運動に 基づく移動が 90-s sample videosに記録され、Nanoparticle Tracking Analysis (NTAs) 2.0 analytical software (Nanosight)で解析された。PBSでサンプルを希釈
し、NTAs測定に適した濃度(1 × 108-2.5 × 109 particles/mL)にした。capture
超純水を用いて 10 倍に希釈し Bradford 法によりタンパク質の濃度を測定した。 HUVECs: 培養した 100mmcell culture dish の上清を取り除き PBS を洗浄し、再度 PBS を取り除いた。100mmcell culture dish に Lysis Buffer を加え、スクレーパー を用いて細胞を剥がし、マイクロチューブに採取した。室温で 10 分間穏やかに攪拌 し、14000×g で 15 分間、4℃で遠心分離を行い、上澄みを採取した。採取した上澄み を、超純水を用いて 10 倍に希釈し、Bradford 法によりタンパク質の濃度を測定した。 8. タンパク質濃度測定-Bradford 法
Bradford 法を用いてタンパク質の濃度測定を行った。Dye reagent 試薬(Bio-Rad
Laboratories, Inc.,Hercules, CA)を超純水で 5 倍に希釈し、ひだ折り濾紙にて濾過
した。濾液を 96well plate に 200μL 入れ、それぞれの濃度に調製したスタンダード とサンプルを 10μL ずつ加えた。5 分間室温で静置した後、マイクロプレートリーダ ーにて波長 550nm の吸光度を測定し、タンパク質の濃度を算出した。
9. Immunoblotting
プレミックスバッファー10×トリス/グリシン/SDS(Bio-Rad Laboratories, Inc.) を超純水を用いて 10 倍に希釈し泳動バッファーを調製した。プレミックスバッファ ー10×トリス/グリシン 100mL(Bio-Rad Laboratories, Inc.)にメタノール(Wako Pure Chemical Industries, Ltd., Osaka, Japan)200mL と超純水 700mL 加え、トランスバ ッファーを調製した。バッファーは調製後、4℃にて冷やした。Laemmli サンプルバッ フ ァ ー (Bio-Rad Laboratories, Inc.) に 2- メ ル カ プ ト エ タ ノ ー ル (2-ME: Sigma Aldrich, St. Louis, MO)を 5%となるように加えてサンプルバッファーを調製した。1 ウェルに対して 20μg のタンパク質が泳動されるようにサンプルを調製し、100℃で 5min 加熱した。サンプルを加熱した後遠心し、氷上に静置した。Mini-PROTEAN TGX Precast Gels(Bio-Rad Laboratories, Inc.) を 泳 動 層 (Mini-PROTEAN Tetra Cell(Bio-Rad Laboratories, Inc.))にセットした。泳動層に泳動バッファーを加え た。サンプルをウェルに注入した。
泳動層を電源装置(パワーパック HC (Bio-Rad Laboratories, Inc.))へつなぎ、
電気泳動(200V)を行った。泳動が終わったゲルをトランスバッファーに 15 分間浸し、
泳動層を電源装置へつないで電気泳動(50V, 2 時間)を行い、ゲルからメンブレンへ転 写 し た 。 転 写 終 了 後 、 回 収 し た メ ン ブ レ ン を Wash Buffer(ATTO CORPORATION,Tokyo,Japan)で、洗浄しブロッキングエース(DS Pharma Biomedical Co., Ltd., Osaka, Japan)を加えて一時間室温で振盪した。
Inc.):1/250)、一晩 4℃で振盪した。溶液を捨て、Wash Buffer にて 3 回洗浄(5 分間/ 回)した。ブロッキングエースを加え、2 次抗体をそれぞれの濃度加えて一時間、室温 で振盪した。溶液を捨て、Wash Buffer にて 3 回洗浄(5 分間/回)した。Detection Reagent 1, 2(GE Healthcare Bio-Science Corp., Piscataway, NJ) を等量混合した ものを、メンブレンの上に加え、1 分間静置した。メンブレンをシーリングし、フィ ルムに感光した。ImageJ software(http://rsb.info.nih.gov/nih-image/)にて解析 した。
10. Tube formation assay
凍結している BD Matrigel(BD Biosciences)を 4℃にて解凍し、また 24 well culture plate とピペットを 4℃にて冷却した。氷上に 24 well culture plate を置き、十分 冷却したピペットで、1 ウェル当たり 0.289mL の BD Matrigel を、各ウェルに泡だて ないように加えた。24 well culture plate を一時間、37℃でインキュベートした。 BD Matrigel に HUVECs を播種し、採取したエクソソームを添加し培養した。顕微鏡に て形態観察を行い、ImageJ にて解析した。
11. miRNA の抽出
K562 細胞: 回収した細胞懸濁液を 2500×g で 10 分間、4℃で遠心分離し、上清を 取り除いた。PBS を添加し、再懸濁(洗浄)した後、再度遠心分離を行った。上清を 除去し、細胞ペレットに QIAzol Lysis Reagent(Qiagen, Hilden, Germany)を加え、 再懸濁した。キット(miRNeasy kit (Qiagen))のプロトコールに従い、miRNA を採取し た。
HUVECs: 培養した 100mm cell culture dish の上清を取り除き、PBS により洗浄し、 再度 PBS を取り除いた。100mm cell culture dish に QIAzol Lysis Reagent(Qiagen) を加え、スクレーパーを用いて細胞を剥がし、マイクロチューブに採取した。製造者 のプロトコ-ルに従い、miRNA を採取した。
12. miRNA の測定
Nano Drop(NanoDrop 1000, Thermo Fisher Scientific Inc., Waltham, MA)を用 いて、miRNA の濃度測定を行った。
13. TaqMan Low-Density miRNA Array
用いるように設定した。逆転写反応と前増幅の過程は製造者の推奨方法に従って行っ た 。 miRNA の 逆 転 写 は 、 Megaplex Primer Pools (Human Pools A v2.1, Applied Biosystems) を用いた。exosome サンプルの逆転写反応産物は、Megaplex PreAmp Primers (Primers A v2.1, Applied Biosystems)を用いてさらに増幅した。
miRNA の発現 profile は、Human TaqMan miRNA Arrays A (Applied Biosystems)を 用いて定量した。細胞と培地それぞれに対して普遍のコントロールとして RNU6B と spike control (ath-miR159)を用いた。定量的リアルタイム PCR は Applied Biosystems
7900HT thermal cycler を用いて製造者の推奨プログラムに従い行った。最後に 7900HT
か ら 得 ら れ た す べ て の 生 デ ー タ は 、 Data Assist Software ver.3.1 (Applied Biosystems)に解析した。 第三節 結果 1.低酸素環境下における K562 細胞の増殖 まず、K562 細胞の低酸素応答について検討した。細胞増殖は、通常酸素状態(酸素 濃度 20%)と低酸素状態(酸素濃度 1%)で、24、48、72 時間後の細胞数を測定した。24 時間後における K562 細胞の増殖は、通常酸素状態と低酸素状態の間で有意な差がみ られなかったが、続く 48-72 時間では低酸素状態において細胞数の有意な減少がみ られた(Fig.1A)。 低酸素応答性マーカーである HIF-1αの発現を計測することで通常酸素状態と低酸 素状態間で細胞増殖の差がみられない 24 時間後のサンプルに対し、低酸素応答を確 認した(Fig.1B)。 2.K562 細胞より放出されたエクソソーム量の測定 K562 細胞を通常酸素状態/低酸素状態でそれぞれ 24 時間培養し、エクソソームを 抽出した。エクソソームの超微細構造を透過型電子顕微鏡(TEM)にて観察した。低酸 素状態で 24 時間後に放出されたエクソソーム(K5621%O2-exosome)の大きさは、通常酸 素状態で 24 時間後に放出されたエクソソーム(K56220%O2-exosome)とほぼ同じで、それ ぞれの形状は二重膜構造を示した”cup-shaped”形態であった(Fig.1C,D)。 次に、エクソソームのマーカーである CD63 を immunoblotting により測定した。通 常酸素状態/低酸素状態で放出された 24 時間後エクソソームにおいて CD63 発現量の 差はみられなかった(Fig.1E)。 さ ら な る エ ク ソ ソ ー ム の サ イ ズ 分 布 を み る た め に 、 K56220%O2-exosome ま た は
K5621%O2-exosome に 対 し Nanosight LM10 system を 用 い て NTAs を 行 っ た 。 K5621 %
O2-exosomeのナノ粒子サイズ分布はK56220%O2-exosomeとよく似ており、粒子径のピー
クは両者共に約 100nmであった(Fig.1F)。ナノ粒子数を細胞数で正規化し、細胞当た
りのナノ粒子数を比較するとK56220%O2-exosomeとK5621%O2-exosomeの間で有意な差が
放出するエクソソームの大きさや数へ影響を及ぼさないことが分かった。一方、72 時間の低酸素状態は、通常酸素状態で 72 時間後に放出されたエクソソームと比べて エクソソームのナノ粒子サイズ分布を変化させた(data not shown)。通常酸素状態 で 72 時間後に放出されたエクソソームでは、横長の粒子サイズ分布を示すためエク ソソームの放出量を計測することが出来なかった。 24 時間の低酸素状態は、がんの細胞増殖とエクソソーム量に対して影響を与えない ことが分かった。そのため、24 時間に放出されたエクソソームを用いて以下の検討を 行った。 3.低酸素環境における K562 細胞由来エクソソームの HUVECs への血管新生増強作用 K56220%O2-exosomeまたはK5621%O2-exosomeを用いて、24 時間低酸素状態で培養した
K562 細胞が放出したエクソソームがHUVECsの管形成へ与える影響を検討した。 K5621 % O2-exosome を 添 加 し た 場 合 (Fig.2A,2D,**p<0.001,*p<0.01) で は 、 K56220 %
O2-exosomeを添加した場合(Fig.2B,2D)や通常酸素状態のHUVECs(HUVECs20%O2)のみの場
合(Fig.2C,2D)と比べ、有意に管形成を促進した(**p < 0.001, *p < 0.01)。また、
K5621%O2-exosomeによるHUVECsの増殖や細胞生存率に対する検討も行ったが、K56220%
O2-exosomeとK5621%O2-exosomeの間に有意な差がみられなかった。このことから低酸素
環境下でがんが放出するエクソソームは、血管内皮細胞の増殖よりむしろ管形成へ作 用していることが分かった。
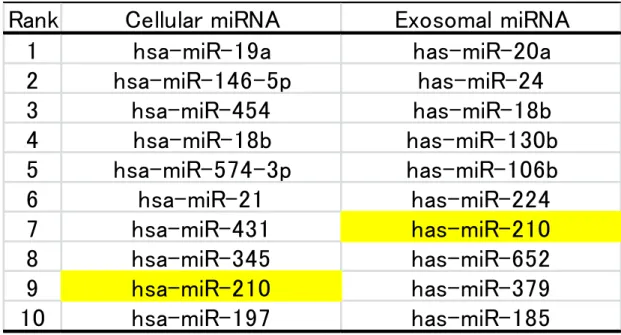
4. 低酸素環境下における K562 細胞内、エクソソーム内の miRNA profiling
低酸素で誘導されたexosomal miRNAを特定するために、TaqMan low-density array 解析を行った。通常酸素状態に比べ低酸素状態で 1.5 倍以上発現が上昇したものを抽
出し、細胞内とエクソソーム内のmiRNAそれぞれに対し 2(-ΔCt)値の順で並び替えた
(Table 1)。並び替えた上位 10 のうち細胞内とエクソソーム内で共通したmiRNAとし て、miR-18bとmiR-210 の 2 つが抽出された。
これら 2 つの miRNA が K562 細胞特異的なものであるかを確認するために、さらに 2 種のがん細胞(SU-DHL-4, RPMI-8226)に対して同様に低酸素で誘導された exosomal miRNA を特定するための TaqMan low-density array 解析を行った。通常酸素状態に比 べ低酸素状態で 1.5 倍以上発現が上昇した細胞内、エクソソーム内それぞれの miRNA を、Ct 値によって分類(20-25, 25-30, 30-35)した(Table S1,S2)。
FIGURE 1. The response to hypoxia in K562 cells. (A) 5 × 104 cells/mL of K562
cells were seeded, and cell growth was measured following 24, 48, and 72 h of normoxic (20% O2) or hypoxic (1% O2) cultivate conditions. (B) The expression
level of HIF-1α protein in K562 cells cultured under 20% O2 or 1% O2 conditions
for 24 h. (C) Photomicrograph of an exosome derived from hypoxic K562 cells. Scale bar, 100nm. (D) Photomicrograph of an exosome derived from normoxic K562 cells. Scale bar, 100 nm. (E) CD63 (exosomal marker) immunoblot of exosome derived from K562 cells cultured under 20% O2 or 1% O2 conditions for 24 h. Equal
amount of exosome (300 ng) were used for the assay. (F, G) The nanoparticle size distribution for the exosome derived from K562 cells cultured under 20% O2 or
1% O2 conditions for 24 h were obtained by NTA (F), and the nanoparticle
FIGURE 2. The exosome derived from K562 cells in hypoxic conditions enhance tube formation of HUVECs. (A, B, C) The formation of tube-like structures was observed under bright field. Endothelial tube formation of HUVECs cultured on Matrigel with K5621%O2-exosome (A), and with K56220%O2-exosome (B), or without K562-exosome
(control) (C). The scale bar indicates 500 μm. (D) The tube-like structures determined by pixel density are significantly enhanced by the addition of K56220%O2-exosome (*p < 0.01) or K5621%O2-exosome (**p < 0.001) as compared with
control (HUVECs20%O2). (E, F, G) Endothelial tube formation of HUVECs with
RPMI82261%O2-exosome (E), and with RPMI822620%O2-exosome (F), or without
RPMI8226-exosome (control) (G). The scale bar indicates 500 μm. (H) The tube-like structures were significantly enhanced by the addition of RPMI822620%O2-exosome (*p < 0.01) or RPMI82261%O2-exosome (**p < 0.001) as compared
Table 1. Top 10 cellular and exosomal miRNAs elevated under hypoxic conditions in K562 cells
Ranking: Cellular and exosomal miRNAs which were up-regulated less
than 1.5-fold under hypoxic conditions are ranked by the expression level.
Rank
Cellular miRNA
Exosomal miRNA
Table S1. Cellular miRNAs elevated under hypoxic condition in K562, RPMI8226, and SUDHL-4 cells K562 RPMI8226 SUDHL-4
Ct values
Table S2. Exosomal miRNAs elevated under hypoxic condition in K562, RPMI8226, and SUDHL-4 cells K562 RPMI8226 SUDHL-4
第四節 考察 腫瘍細胞から放出されたエクソソームの生物学的機能についてはすでに広範囲で 研究が行われている(12, 24, 25)。しかしながら、低酸素環境における腫瘍細胞から 放出されたエクソソームの定量的変化は、完全には明らかにされていない。Kingらに よる最近の研究で、著しい低酸素状態(0.1% O2)での 24 時間培養または、適度な低酸 素状態(1% O2)での 48 時間以上の培養により乳癌細胞からのエクソソーム放出が促進 されることが明らかになった(26)。このことから、低酸素環境下における腫瘍細胞か ら標的細胞へのエクソソームの機能的な伝達を考える際にエクソソームの放出量を 考慮することは重要である。本研究でも、適度な低酸素状態(1% O2)での 48 時間以上 の培養によりエクソソームの放出量が増加しNanosight LM10 systemによるエクソソ ーム粒子径の測定では変化が大きく、Kingらの知見と一致した(26)。一方、低酸素状 態で 24 時間培養したK562 細胞から放出されるエクソソームの粒子径は通常酸素状態 で放出されたエクソソームの粒子径とほとんど差がなかった。長期間の低酸素曝露が どのようにして腫瘍細胞のエクソソームの放出と機能に影響を及ぼすかはまだ分か っていない。それ故、本研究ではエクソソームの量ではなく含有物による影響を特定 するために、低酸素状態(1% O2)で 24 時間培養した低酸素腫瘍細胞から放出されるエ クソソームを用いた。 エクソソームは特異的なタンパク質や RNA、miRNA、脂質を含む。最近の研究で、受 容細胞へのエクソソーム由来の特異的な miRNA の輸送は、直接的な細胞間相互作用や 化学的な受容体を介した事象等の古典的な機序に加えて、遺伝子に基づいた細胞間伝 達を可能とする別の機序であることが提唱されている(14)。低酸素シグナル伝達は、 重要な転写抑制因子である低酸素誘導因子(HIF)によって制御されることが知られて いる(28)。従来、HIF を介する経路の上流や下流シグナルの制御には、いくつかの miRNA
第二章 低酸素白血病細胞由来エクソソームに含まれる miR-210 による血管新生増強 作用 第一節 目的 エクソソームは特異的なタンパク質や RNA、miRNA、脂質を含む。miRNA は臓器特異 的な発現様式を示し、組織の分化や機能、形態の維持、臓器疾患の分子病態に重要な 役割を果たしていることが示唆されている。低酸素シグナル伝達は、重要な転写抑制 因子である低酸素誘導因子(HIF)によって制御されることが知られているが、HIF を 介するシグナル伝達経路の上流や下流シグナルの制御にいくつかの miRNA が関係して いる(28)。第 1 章の結果より、低酸素腫瘍細胞から放出されるエクソソームにおける miRNA の特徴は、解析した 3 つのがん細胞種間で差があり、exosomal miRNA によって 調整される低酸素シグナル伝達は、がん種に依存することが示された。しかし、解析 されたがん細胞種すべてに共通して miR-210 の発現が上昇した。そのため、miR-210 が低酸素環境に曝露された腫瘍細胞から放出される主要な eoxosmal miRNA と考え、 第 2 章では miR-210 の血管新生増強作用について検討を行った。
第二節 方法
1. K562 細胞への Cy3 標識 pre-mir miRNA 前駆体遺伝子導入と PKH67 標識エクソソー ムの伝達
Pre-mir miRNA 前駆体 (has-miR-210; Applied Biosystems)は、manufacturer’s instructions (Mirus, Madison, WI)に従って、Label IT siRNA Tracker Cy3 Kitに
て標識化した。K562 細胞(1× 105)にHiPerFect (Qiagen)を用いて 10nMのCy3 標識
pre-mir miRNA 前駆体を遺伝子導入した(K562/Cy3-miR-210)。遺伝子導入した日に、 K562 細胞をPBSで 3 回洗浄し、培地を新鮮な無血清のAIM V medium (Invitrogen)に 交換した。
1 日間インキュベートした後、ExoQuick (System Biosciences)によるエクソソーム抽 出のために上清を回収した。Cy3-miR-210 を含むエクソソームを 2 μM PKH67(Sigma Aldrich, St. Louis, MO)と共に、2 分間 25℃でインキュベートされ、Amicon Ultra-0.5 (100 kDa, Millipore, Billerica, MA)を用いて 4 回 PBS で洗浄した。
2. Exosomal miR-210 knock down 試験
提供 細胞 (K562 細胞; 1 × 105) または受容細胞 (HUVECs; 3 × 104)に、HiPerFect
(Qiagen) を 用 い て scrambled control (Negative Control #1, Ambion) ま た は anti-miR-210 miRNA inhibitors (Ambion)を遺伝子導入した。次の日、新しい培地へ
交換した後、細胞を低酸素状態で 24 時間培養した。K5621%O2/anti-miR-210 または
3. Luciferase assay
miR-210 相補的結合部位を含む EFNA3 3′-UTR 関連合成オリゴヌクレオチド(5′ -TTTGTCTTCTGTGAAGACAGGACCTATG CAACGCACAGACACTTTTGGAGACCGT-3′)を製造者のプロ ト コ - ル に 従 っ て firefly Luciferase reporter plasmid pMIR-Report (Applied Biosystems)にサブクローニングした。miR-210 (ACGCACA)のシード配列はボールド・ イタリック書体によって示した。また、miR-210 のシード配列を置換した mutated sensor vector を作成した。ルシフェラーゼ活性を測定するために、HUVECs を 24 well plate に播種し 50%–60% コンフルエンスまで育てた。導入効率のコントロールである 0.01 μg of pMIR-Report β-gal control plasmid (Applied Biosystems)と Luciferase plasmid (0.1 μg)を同時導入した。
1 日間インキュベートした後、低酸素状態(K5621%O2-exosome)または通常酸素状態
(K56220%O2-exosome)で培養したK562 細胞が放出したエクソソームをHUVECsに添加し、
48 時 間 後 の Luciferase と β -gal の 活 性 を 製 造 者 の プ ロ ト コ - ル (Applied Biosystems)に従いDual-Light Systemにて評価した。
4. 免疫染色
HUVECsをカバーガラス(Matsunami Glass, Japan)に播種し、K5621%O2-exosomeまたは
K56220%O2-exosome と 共 に 通 常 酸 素 状 態 (20% O
2) で 培 養 し た 。 polyclonal
anti-Ephrin-A3 antibody (sc-1012; Santa Cruz, Inc.)を用いてEphrin-A3 を検出し た 。 2 次 抗 体 と し て Alexa Fluor 488-conjugated anti-mouse IgG goat serum (Molecular Probes, Invitrogen)を用いた。4′,6-diamidino-phenylidole (DAPI I; Abbott, Abbott Park, IL)にて核染色を行い、fluorescent microscope (Biozero BZ-8000; Keyence, Osaka, Japan)で解析した。
5. 統計的解析
データは mean ± SD で表記した。2 群間比較検定は、Mann-Whitney U test または Student’s t-test にて行った。多重比較検定は ANOVA を用いた。GraphPad Prism version 5c for Macintosh (GraphPad Inc., La Jolla, CA) を用いて統計解析した。
P 値が 0.05 未満のとき(p < 0.05)、両者は有意に異なると判断した。
第三節 結果
1. 低酸素環境下における K562 細胞由来 exosomal miR-210 による HUVECs のターゲッ ト遺伝子の制御
低酸素状態で K562 細胞より放出された exosomal miR-210 と血管新生増強作用との 関連を明らかにするために、exosomal miR-210 がエクソソームを介して HUVECs のタ ーゲット遺伝子を制御するかを検討した。
K562/Cy3-miR-210 細胞由来の PKH67 でラベル化されたエクソソームと共に培養した
後、Cy3-miR210 シグナルと PKH67 シグナルが HUVECs の細胞質内で共局在を示した(Fig.
3A-F)。
次に、exosomal miR-210 がHUVECs内のmiR-210 のターゲット因子であり抗血管新生 因子であるEphrin-A3(EFNA3)を直接制御するかを調べるためにluciferase reporter assayを行った。miR-210 と相補的な結合サイトとEFNA3 3’-UTRを含むreporter
plasmids (sensor vector)を形質導入したHUVECsをK5621%O2-exosomeとともに培養し
た場合、ルシフェラーゼ 活性はK56220%O2-exosomeとともに培養した場合と比べ有意に
減少した(*p < 0.05; Fig. 3G)。対照的に、miR-210 のmutated sensor vectorを用い た場合、HUVECs20%O2のみ、K56220%O2-exosomeまたはK5621%O2-exosomeのそれぞれの場合で
のルシフェラーゼ 活性の差が生じなかった(Fig.3H)。 2. 管形成への exosomal miR-210 関与
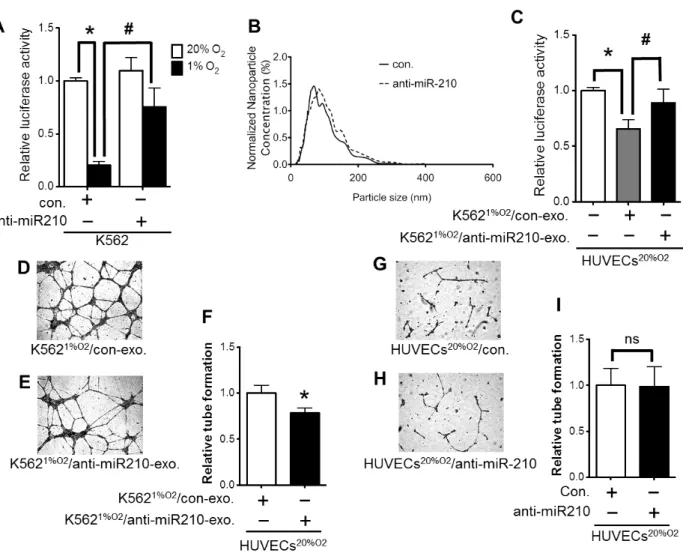
exosomal miR-210 が血管内皮細胞の管形成を促進する主要な役割をはたすことを 示すために knock down 試験を行った。
まず、ルシフェラーゼ活性を指標として、miR-210 阻害剤の有効性を検討した。 sensor vector を形質導入した K562 に miR-210 阻害剤を形質導入した(Fig. 4A)。 miR-210 阻害剤に対するコントロールを形質導入した場合では、K562 に形質導入され た sensor vector が、低酸素状態で発現した miR-210 と反応し、ルシフェラーゼ活性 が低下した(*p < 0.05)。一方、miR-210 阻害剤を形質導入すると低酸素状態での miR-210 発現が阻害されコントロールを形質導入した場合に比べルシフェラーゼ活性 の低下が抑えられた(#p < 0.05)。対照的に、miR-210 の mutated vector を形質導入 した場合は、miR-210 阻害剤はルシフェラーゼ活性に影響を与えなかった(date not shown)。
次に、miR-210 阻害剤またはコントロールを形質導入したK562 細胞を低酸素状態 で培養し、それぞれが放出したエクソソームをNTAsにて測定した(Fig.4B)。miR-210 阻害剤は、細胞内のmiR-210 に加えて、exosomal miR-210 の活性も阻害した。低酸素 状態で培養されたmiR-210 阻害剤に対するコントロールを形質導入したK562 細胞から
放出されたエクソソーム(K5621%O2/con-exo.)をHUVECsへ添加すると、通常酸素状態の
HUVECs(HUVECs20%O2)と比べルシフェラーゼ活性を低下した(*p<0.05)。しかし、miR-210
阻害剤を形質導入して低酸素状態で培養したK562 細胞から放出されたエクソソーム (K5621%O2/anti-miR-210-exo.) は 、 sensor vector を 形 質 導 入 し た HUVECs 内 の
luciderase活性を減少することが出来なかった(#p < 0.05, Fig.4C)。さらに低酸素 状態でmiR-210 をノックダウンしたK562 細胞から放出されたエクソソームを用いて tube formation assayを行ったところ、管形成が低下した(Fig. 4D-F)。
よるものだという可能性は完全には除外出来なかった。
さらに、受容細胞であるHUVECsの内在性miR-210 の影響を調べるために、HUVECsに 直接的にmiR-210 阻害剤を形質導入し、通常酸素状態にて管形成を行った。HUVECsに
anti-miR-210 を形質導入した場合(HUVECs20%O2/anti-miR-210)とコントロールを形質
導入した場合(HUVECs20%O2/control)で有意な差がみられなかった(Fig. 4G-I)。
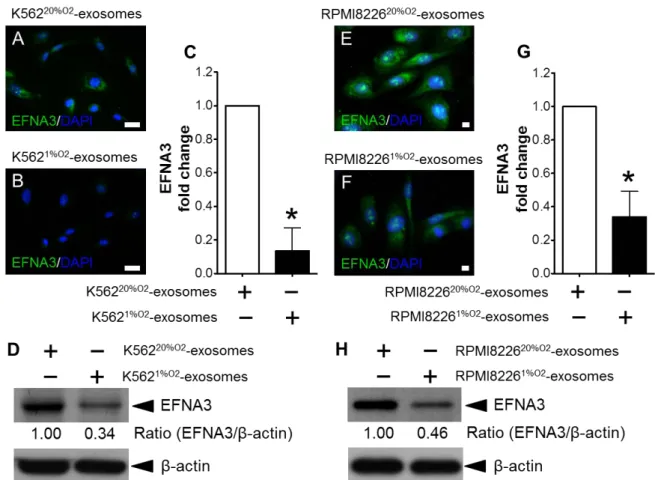
3. exosomal miR-210 による HUVECs における Ephrin-A3 発現阻害
免疫染色を用いてK56220%O2-exosomeまたはK5621%O2-exosomeをHUVECsに添加し培養し
た際のEFNA3 発現を 、免疫染 色法を 用い て検討し た。通 常酸 素状態のHUVECsに
K5621%O2-exosomeを添加することによってEFNA3 の発現は抑制された(Fig. 5A–C, *p <
0.001)。
また、K56220%O2-exosomeまたはK5621%O2-exosomeをHUVECsに添加し培養した際のEFNA3
発現をImmunoblotting法により確認した場合も同様の結果が得られた(Fig. 5D)。低 酸 素 状 態 で 培 養 し た RPMI-8226 細 胞 か ら 放 出 さ れ た エ ク ソ ソ ー ム
(RPMI-82261%O2-exosome)を添加した場合においても同様にHUVECsにおけるEFNA3 の発
現が抑制された(Fig. 5E-H)。
The scale bar indicates 10 μm. (G) Luciferase assay. The luciferase reporter vector for assessing miR-210 specific activity expressed firefly luciferase containing complementary miR-210 sequences in its 3′-untranslated region. The luciferase reporter vector and β-gal control vector allowed simultaneous monitoring of miR-210 activity and transfection efficiency, respectively. Sensor vector: Luciferase activity of HUVECs cultured with exosome from K562 cells cultured under hypoxic conditions (K5621%O2-exosome) was significantly
reduced compared with HUVECs only or cultured with exosome from K562 cells cultured under normoxic conditions (K56220%O2-exosome) (*p < 0.05; n = 3). Mutated
FIGURE 4. Knockdown experiment of exosomal miR-210. (A) The validation of the efficacy of miR-210 inhibitor in K562 cells transfected with the luciferase reporter vector containing miR-210 sequences in EFNA3 3′-UTR. The transfection of anti-miR-210 to K5621%O2 cells inhibited the induction of luciferase activity
(normoxia control vs. hypoxia control: *p < 0.05; anti-miR-210 vs. negative control: #p < 0.05; n = 3). (B) The nanoparticle size distribution for the exosome derived from K5621%O2/con (K5621%O2 cells transfected with negative control miR)
and K5621%O2/anti-miR-210 (K5621%O2 cells transfected with miR-210 inhibitor) were
obtained by NTA. (C) HUVECs20%O2 were transfected with the
luciferase reporter vector containing miR-210 sequences in EFNA3 3 ′ -UTR. Luciferase activity was compared with HUVECs20%O2 cultured with exosome derived
from K5621%O2/anti-miR-210 or K5621%O2/con (with K562-exosome vs. control: *p <
0.05; anti-miR-210 vs. negative control: #p < 0.05; n= 3). (D, E, F) Endothelial tube formation of HUVECs20%02 cultured on Matrigel with K5621%O2/con-exosome (D),
determined by pixel density are reduced by the addition of K5621%O2/anti-miR-210-exosome (*p < 0.01) compared with K5621%O2/con-exosome. (G,
H, I) Endothelial tube formation of HUVECs20%O2 transfected with scramble control
miR (G), and with anti-miR-210 inhibitor (H). (I) There was no difference in the tube-like structures with inhibition of endogenous miR-210 in HUVECs20%O2 (ns;
FIGURE 5. EFNA3 expression of HUVECs cultured with exosome derived from K562 cells. (A, B) EFNA3 expression was inhibited by exosomes derived from hypoxic K562 cells. HUVECs were cultured under normoxic conditions with the exosomes derived from K562 cells cultured under normoxic conditions (K56220%O2-exosomes)
for 24 h (A), and with the exosomes derived from K562 cells cultured under hypoxic conditions (K5621%O2-exosomes) (B). The scale bar indicates 10 μm. (C) EFNA3
signal was quantified using ImageJ software. EFNA3 fluorescence signals were drastically inhibited by addition of K5621%O2-exosomes (*p < 0.001) as compared
with K56220%O2-exosomes. (D) Expression of EFNA3 in HUVECs by immune blot. (E,
F) HUVECs were cultured under normoxic conditions with the RPMI822620%O2-exosomes
for 24 h (E), and with the RPMI82261%O2-exosomes (F). The scale bar indicates 10
μm. (G) EFNA3 fluorescence signals were drastically inhibited by addition of RPMI82261%O2-exosomes (*p < 0.001) as compared with RPMI822620%O2-exosomes. (H)
第四節 考察
K562 細胞由来エクソソームの可視化は、Mieno らによって最近報告された(27)。本 研究では、エクソソーム移入の 3 次元再構成によって可視化することは出来なかった が、すでに報告されている手法(20)を用いて、提供 細胞(K562 細胞 )から受容細胞 (HUVECs)への PKH67 標識 エクソソーム と Cy3 標識 exosomal miRNA の輸送を証明し た (Fig. 3A–F)。
引用文献
1. Joyce, J. A., and Pollard, J. W. (2009) Microenvironmental regulation of metastasis. Nat. Rev. Cancer 9, 239–252
2. Hu, M., and Polyak, K. (2008) Microenvironmental regulation of cancer development. Curr. Opin. Genet. Dev. 18, 27–34
3. Harris, A. L. (2002) Hypoxia—a key regulatory factor in tumour growth. Nat. Rev. Cancer 2, 38–47
4. Vaupel, P., and Mayer, A. (2007) Hypoxia in cancer: significance and impact on clinical outcome. Cancer Metastasis Rev. 26, 225–239
5. Heddleston, J. M., Li, Z., Lathia, J. D., Bao, S., Hjelmeland, A. B., and Rich, J. N. (2010) Hypoxia inducible factors in cancer stem cells. Br J Cancer 102, 789–795
6. Albini, A., and Sporn, M. B. (2007) The tumour microenvironment as a target for chemoprevention. Nat. Rev. Cancer 7, 139–147
7. Bennewith, K. L., and Dedhar, S. (2011) Targeting hypoxic tumour cells to overcome metastasis. BMC Cancer 11, 504
8. Serini, G., Valdembri, D., and Bussolino, F. (2006) Integrins and angiogenesis: a sticky business. Exp. Cell Res. 312, 651–658
9. Shibuya, M., and Claesson-Welsh, L. (2006) Signal transduction by VEGF receptors in regulation of angiogenesis and lymphangiogenesis. Exp. Cell Res. 312, 549–560
10. Park, J. E., Tan, H. S., Datta, A., Lai, R. C., Zhang, H., Meng, W., Lim, S. K., and Sze, S. K. (2010) Hypoxic tumor cell modulates its microenvironment to enhance angiogenic and metastatic potential by secretion of proteins and exosome. Mol. Cell Proteomics 9, 1085–1099 11. Svensson, K. J., Kucharzewska, P., Christianson, H. C., Skold, S., Lofstedt,
T., Johansson, M. C., Morgelin, M., Bengzon, J., Ruf, W., and Belting, M. (2011) Hypoxia triggers a proangiogenic pathway involving cancer cell microvesicles and PAR-2-mediated heparin-binding EGF signaling in endothelial cells. Proc. Natl. Acad. Sci. U. S. A. 108, 13147–13152 12. Vlassov, A. V., Magdaleno, S., Setterquist, R., and Conrad, R. (2012)
13. Valadi, H., Ekstrom, K., Bossios, A., Sjostrand, M., Lee, J. J., and Lotvall, J. O. (2007) exosomemediated transfer of mRNAs and microRNAs is a novel mechanism of genetic exchange between cells. Nat. Cell Biol. 9, 654–659 14. Kosaka, N., Iguchi, H., Yoshioka, Y., Takeshita, F., Matsuki, Y., and Ochiya,
T. (2010) Secretory mechanisms and intercellular transfer of microRNAs in living cells. J. Biol. Chem. 285, 17442–17452
15. Camussi, G., Deregibus, M. C., Bruno, S., Grange, C., Fonsato, V., and Tetta, C. (2011) exosome/microvesicle-mediated epigenetic reprogramming of cells. Am. J. Cancer Res. 1, 98–110
16. Kosaka, N., Iguchi, H., Yoshioka, Y., Hagiwara, K., Takeshita, F., and Ochiya, T. (2012) Competitive interactions of cancer cells and normal cells via secretory microRNAs. J. Biol. Chem. 287, 1397–1405
17. Webber, J., Steadman, R., Mason, M. D., Tabi, Z., and Clayton, A. (2010) Cancer exosome trigger fibroblast to myofibroblast differentiation. Cancer Res. 70, 9621–9630
18. Skog, J., Wurdinger, T., van Rijn, S., Meijer, D. H., Gainche, L., Sena-Esteves, M., Curry, W. T., Jr., Carter, B. S., Krichevsky, A. M., and Breakefield, X. O. (2008) Glioblastoma microvesicles transport RNA and proteins that promote tumour growth and provide diagnostic biomarkers. Nat. Cell Biol. 10, 1470–1476
19. Hood, J. L., San, R. S., and Wickline, S. A. (2011) exosome released by melanoma cells prepare sentinel lymph nodes for tumor metastasis. Cancer Res. 71, 3792–3801
20. Umezu, T., Ohyashiki, K., Kuroda, M., and Ohyashiki, J. H. (2012) Leukemia cell to endothelial cell communication via exosomal miRNAs. Oncogene doi: 10.1038/onc.2012.295
21. Nazarenko, I., Rana, S., Baumann, A., McAlear, J., Hellwig, A., Trendelenburg, M., Lochnit, G., Preissner, K. T., and Zoller, M. (2010) Cell surface tetraspanin Tspan8 contributes to molecular pathways of exosomeinduced endothelial cell activation. Cancer Res. 70, 1668–1678
22. Sullivan, R., Pare, G. C., Frederiksen, L. J., Semenza, G. L., and Graham, C. H. (2008) Hypoxia-induced resistance to anticancer drugs is associated with decreased senescence and requires hypoxia-inducible factor-1 activity. Mol. Cancer Ther. 7, 1961–1973
24. Corrado, C., Flugy, A. M., Taverna, S., Raimondo, S., Guggino, G., Karmali, R., De Leo, G., and Alessandro, R. (2012) Carboxyamidotriazole-orotate inhibits the growth of imatinib-resistant chronic myeloid leukaemia cells and modulates exosomestimulated angiogenesis. PLoS One 7, e42310
25. Kosaka, N., Iguchi, H., Hagiwara, K., Yoshioka, Y., Takeshita, F., and Ochiya, T. (2013) Neutral sphingomyelinase 2 (nSMase2)-dependent exosomal transfer of angiogenic microRNAs regulate cancer cell metastasis. J. Biol. Chem. 288, 10849–10859.
26. King, H. W., Michael, M. Z., and Gleadle, J. M. (2012) Hypoxic enhancement of exosome release by breast cancer cells. BMC Cancer 12, 421
27. Mineo, M., Garfield, S. H., Taverna, S., Flugy, A., De Leo, G., Alessandro, R., and Kohn, E. C. (2012) exosome released by K562 chronic myeloid leukemia cells promote angiogenesis in a Src-dependent fashion. Angiogenesis 15, 33– 45
28. Pugh, C. W., and Ratcliffe, P. J. (2003) Regulation of angiogenesis by hypoxia: role of the HIF system. Nat. Med. 9, 677–684
29. Kulshreshtha, R., Ferracin, M., Wojcik, S. E., Garzon, R., Alder, H., Agosto-Perez, F. J., Davuluri, R., Liu, C. G., Croce, C. M., Negrini, M., Calin, G. A., and Ivan, M. (2007) A microRNA signature of hypoxia. Mol. Cell Biol. 27, 1859–1867
30. Huang, X., Le, Q. T., and Giaccia, A. J. (2010) MiR-210—micromanager of the hypoxia pathway. Trends Mol. Med. 16, 230–237
31. Hua, Z., Lv, Q., Ye, W., Wong, C. K., Cai, G., Gu, D., Ji, Y., Zhao, C., Wang, J., Yang, B. B., and Zhang, Y. (2006) MiRNA-directed regulation of VEGF and other angiogenic factors under hypoxia. PLoS One 1, e116
32. Fasanaro, P., D'Alessandra, Y., Di Stefano, V., Melchionna, R., Romani, S., Pompilio, G., Capogrossi, M. C., and Martelli, F. (2008) MicroRNA-210 modulates endothelial cell response to hypoxia and inhibits the receptor tyrosine kinase ligand Ephrin-A3. J. Biol. Chem. 283, 15878–15883
33. Davis, S., Gale, N. W., Aldrich, T. H., Maisonpierre, P. C., Lhotak, V., Pawson, T., Goldfarb, M., and Yancopoulos, G. D. (1994) Ligands for EPH-related receptor tyrosine kinases that require membrane attachment or clustering for activity. Science 266, 816–819
B. A., Lickliter, J. D., and Boyd, A. W. (2013) EphA3 Maintains Tumorigenicity and Is a Therapeutic Target in Glioblastoma Multiforme. Cancer Cell 23, 238–248
35. Kvasnicka, H. M., Thiele, J., Staib, P., Schmitt-Graeff, A., Griesshammer, M., Klose, J., Engels, K., and Kriener, S. (2004) Reversal of bone marrow angiogenesis in chronic myeloid leukemia following imatinib mesylate (STI571) therapy. Blood 103, 3549–3551